
Ocean-Atmosphere CO2 Fluxes in the North Atlantic Subtropical Gyre: Association with Biochemical and Physical Factors during Spring

Abstract
:1. Introduction
2. Material and Methods
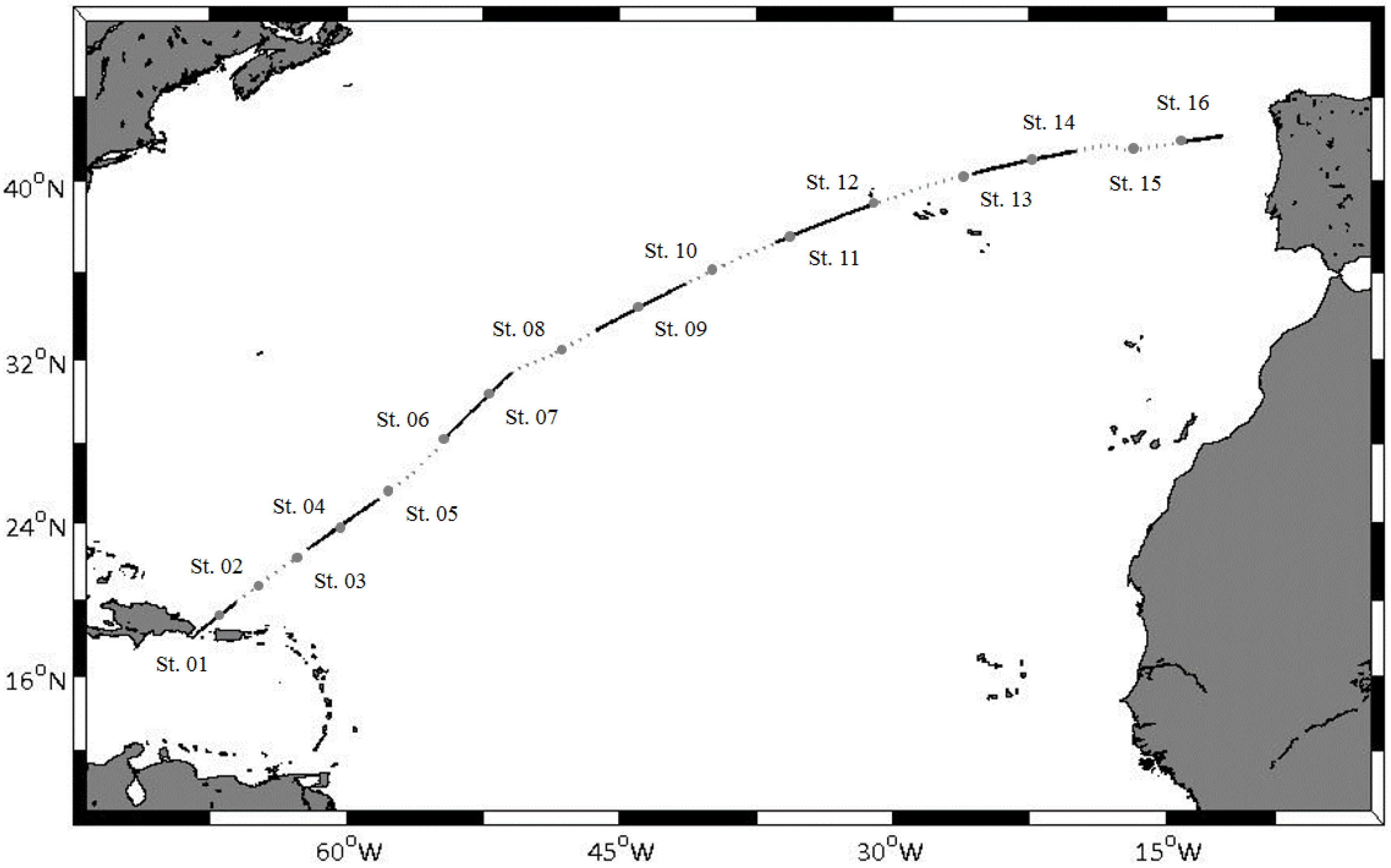
2.1. Study Area and Field Sampling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Location (N, W) | pH | TA (mM) | DIC (mM) | pCO2 (μatm) | ΔpCO2 (μatm) | WS (m s−1) | FCO2 (mmol m−2 day−1) |
|---|---|---|---|---|---|---|---|---|
| 01 | 19.3°, 66.9° | 8.09 | -- | 354.4 | −38.6 | 9.20 | −8.5 | |
| 02 | 20.9°, 64.7° | 8.12 | -- | -- | 348.1 | −44.9 | 5.22 | −3.9 |
| 03 | 22.4°, 65.2° | 8.12 | 2.41 | 1.93 | 346.6 | −46.4 | 4.02 | −2.1 |
| 04 | 24.0° 59.4° | 8.10 | 2.45 | 1.99 | 343.7 | −49.3 | 5.12 | −3.3 |
| 05 | 26.5°, 56.3° | 8.08 | 2.47 | 1.94 | 341.2 | −51.8 | 5.58 | −4.1 |
| 06 | 29.0°, 53.7° | 8.05 | 2.37 | 1.99 | 347.9 | −45.1 | 6.73 | −5.3 |
| 07 | 31.6°, 50.4° | 8.03 | 2.37 | 2.01 | 344.2 | −48.8 | 8.05 | −8.3 |
| 08 | 33.1°, 46.8° | 8.07 | 2.47 | 2.16 | 336.6 | −56.5 | 7.49 | −8.2 |
| 09 | 34.7°, 43.3° | 8.06 | 2.44 | 2.08 | 338.3 | −54.7 | 5.73 | −4.7 |
| 10 | 36.3°, 39.3° | 8.03 | 2.47 | 2.12 | 343.2 | −49.8 | 5.83 | −4.5 |
| 11 | 37.8°, 35.1° | 7.96 | 2.48 | 2.20 | 343.1 | −49.9 | 7.49 | −7.4 |
| 12 | 39.1°, 30.7° | 7.94 | 2.40 | 2.16 | 342.2 | −50.9 | 7.31 | −7.2 |
| 13 | 40.3°, 25,6° | 7.94 | 2.40 | 2.16 | 329.5 | −63.5 | 9.19 | −14.1 |
| 14 | 41.0°, 21.8° | 7.92 | 2.36 | 2.15 | 345.0 | −48.0 | 5.60 | −4.0 |
| 15 | 41.2°, 17.1° | 7.92 | 2.36 | 2.16 | 431.1 | 38.1 | 10.37 | 9.3 |
| 16 | 41.6°, 14.7° | 7.96 | 2.36 | 2.14 | 591.0 | 197.9 | 8.73 | 36.9 |
2.2. Analytical Methods
2.3. Flux Calculation
3. Results
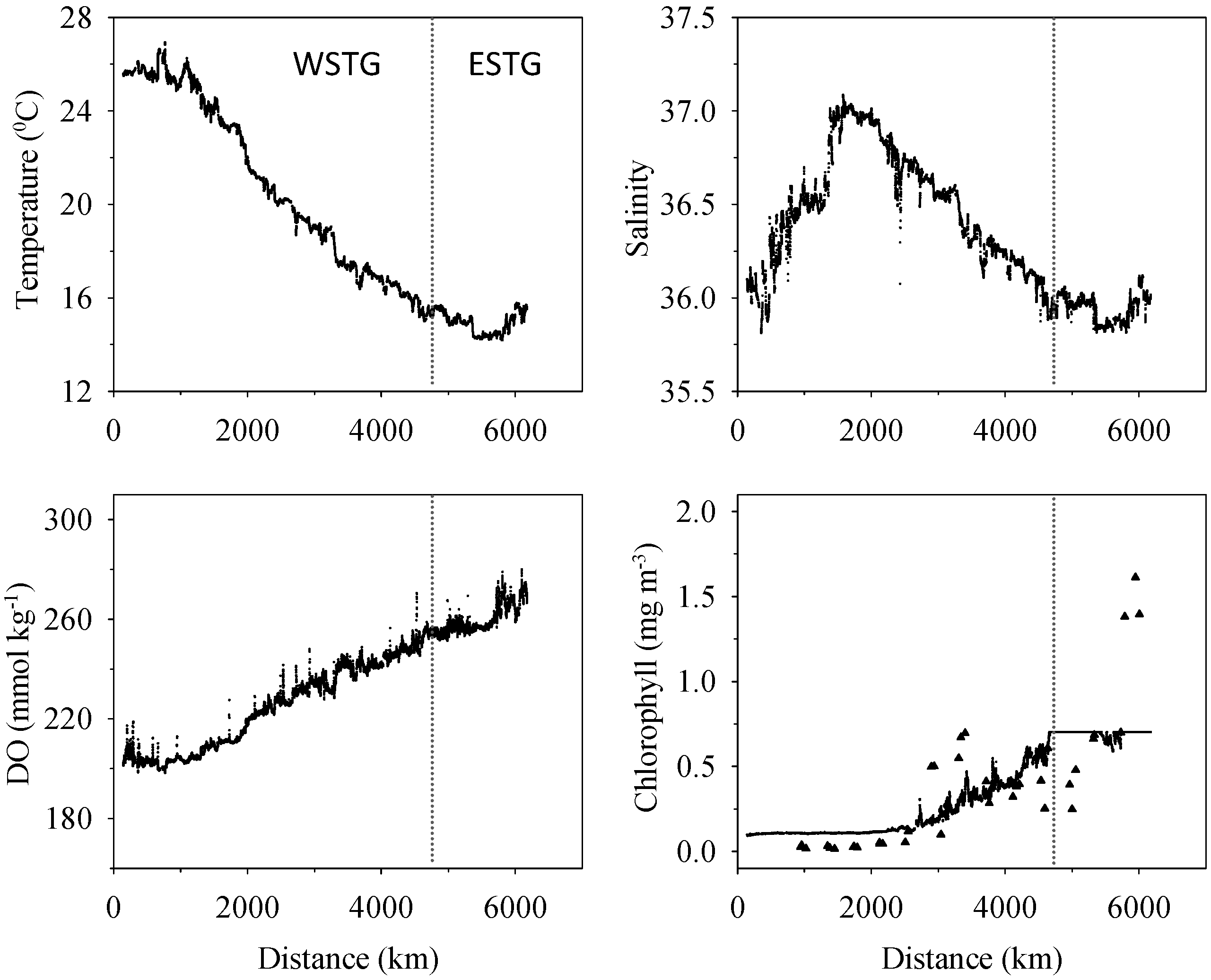
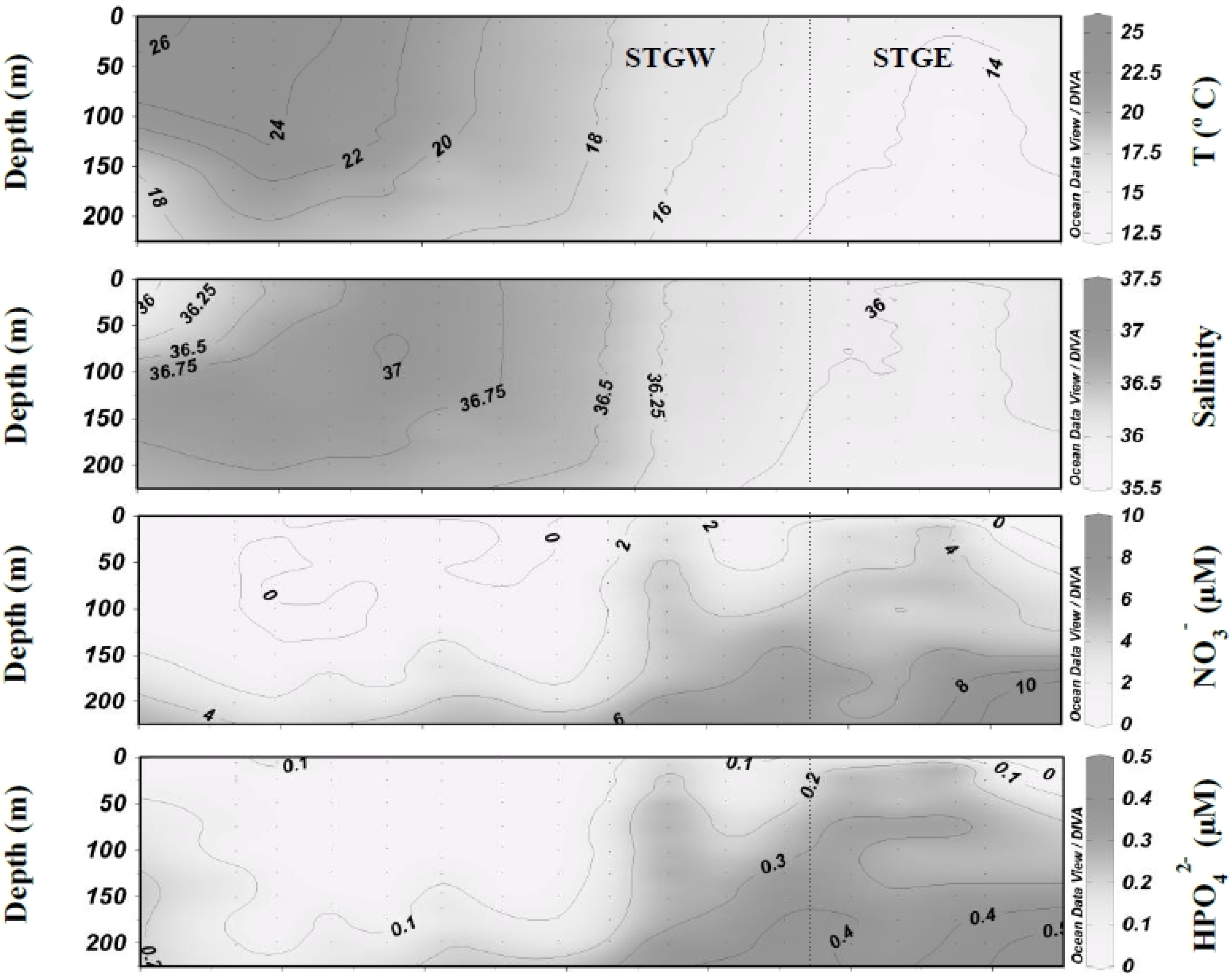
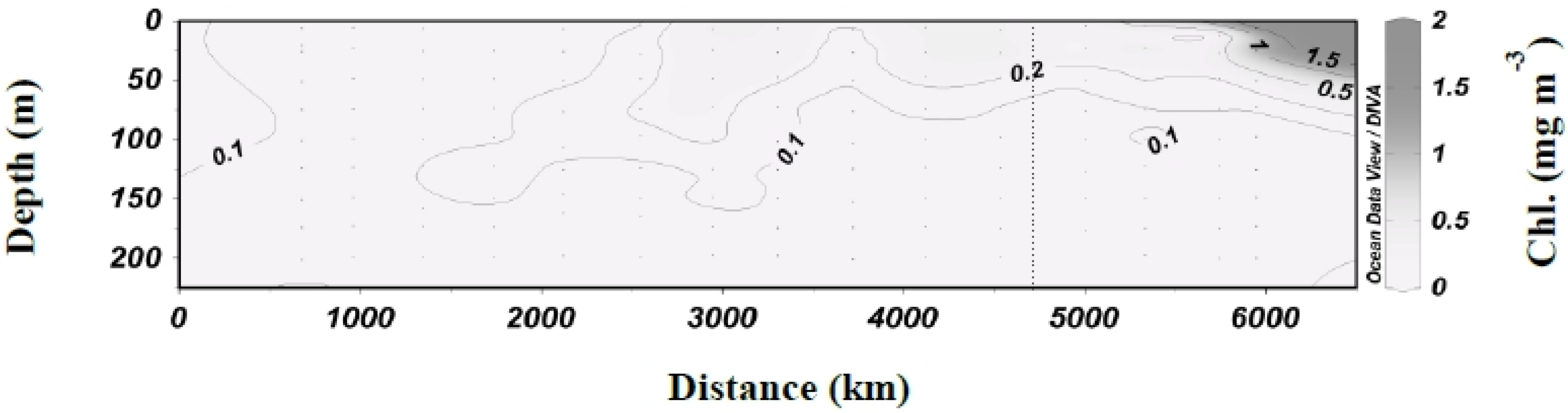
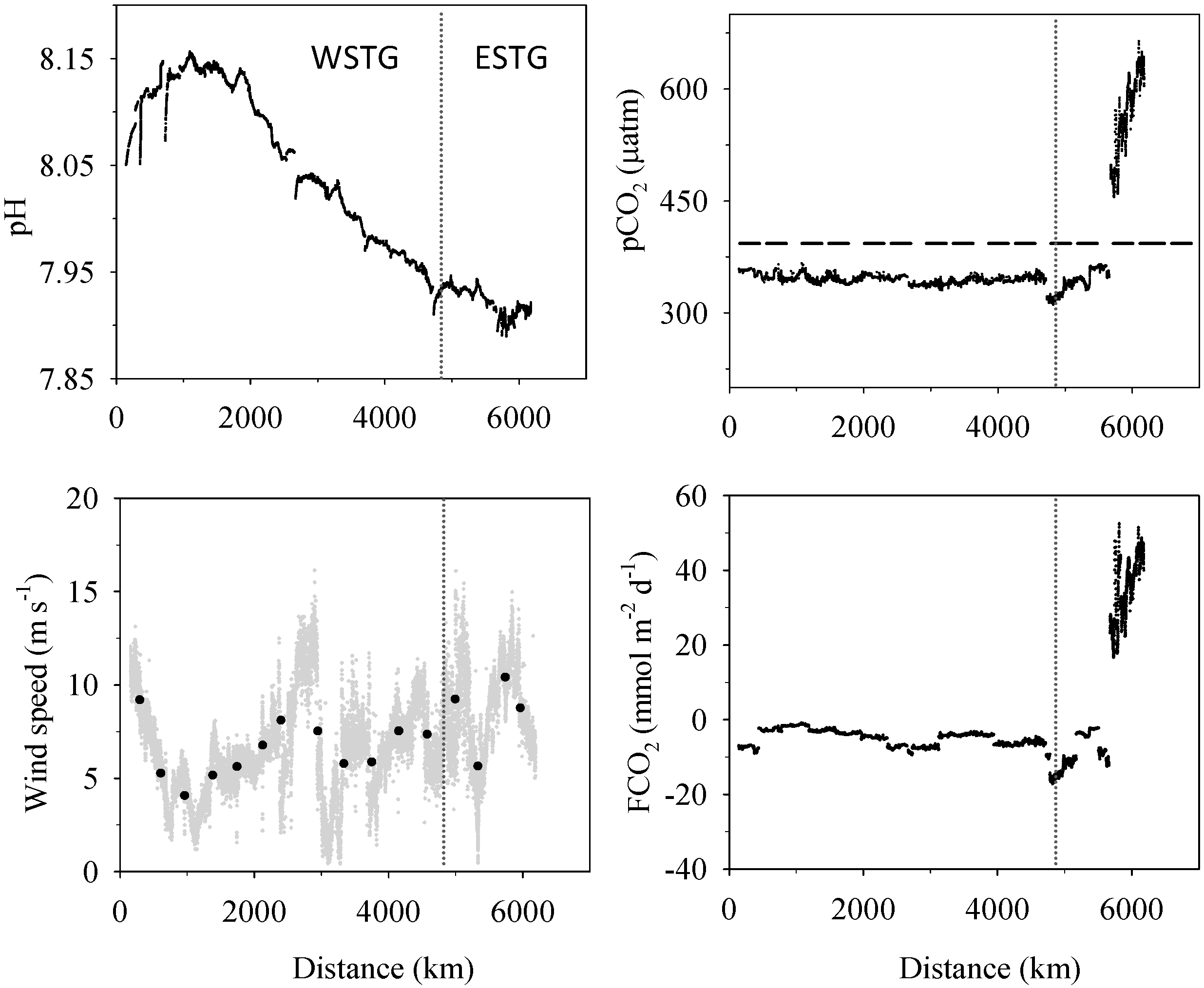
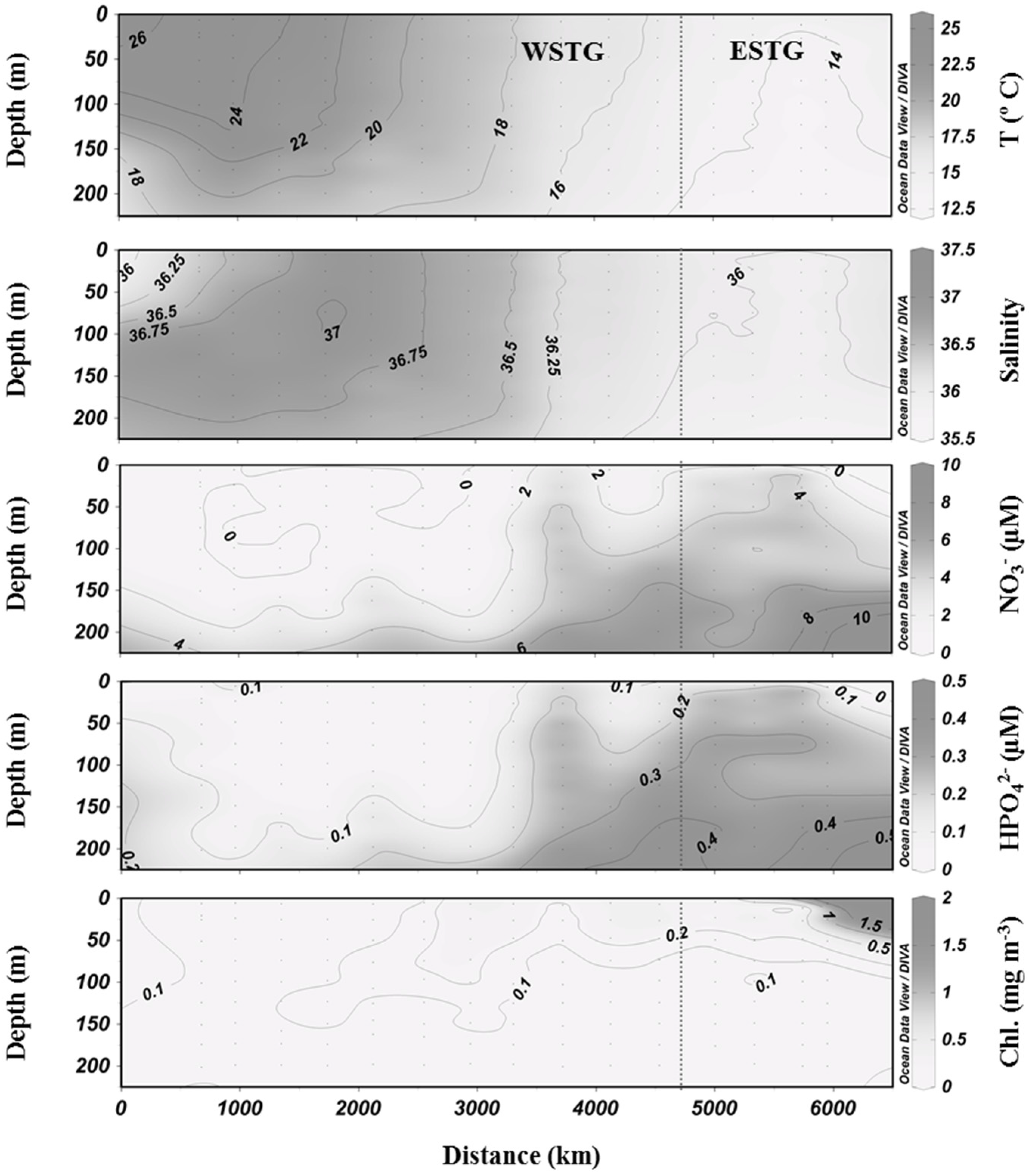
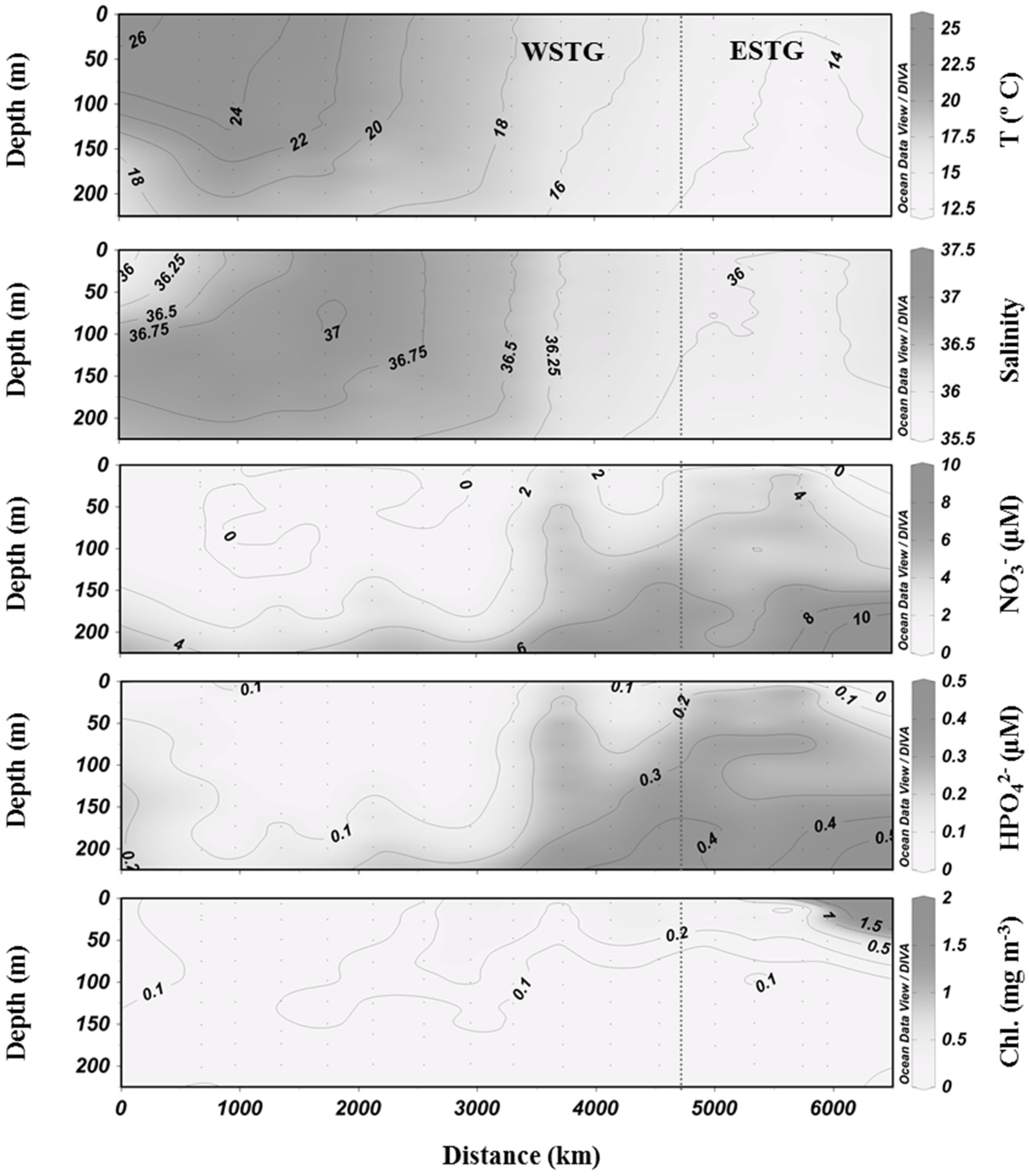
3.1. Biogeochemical Variables
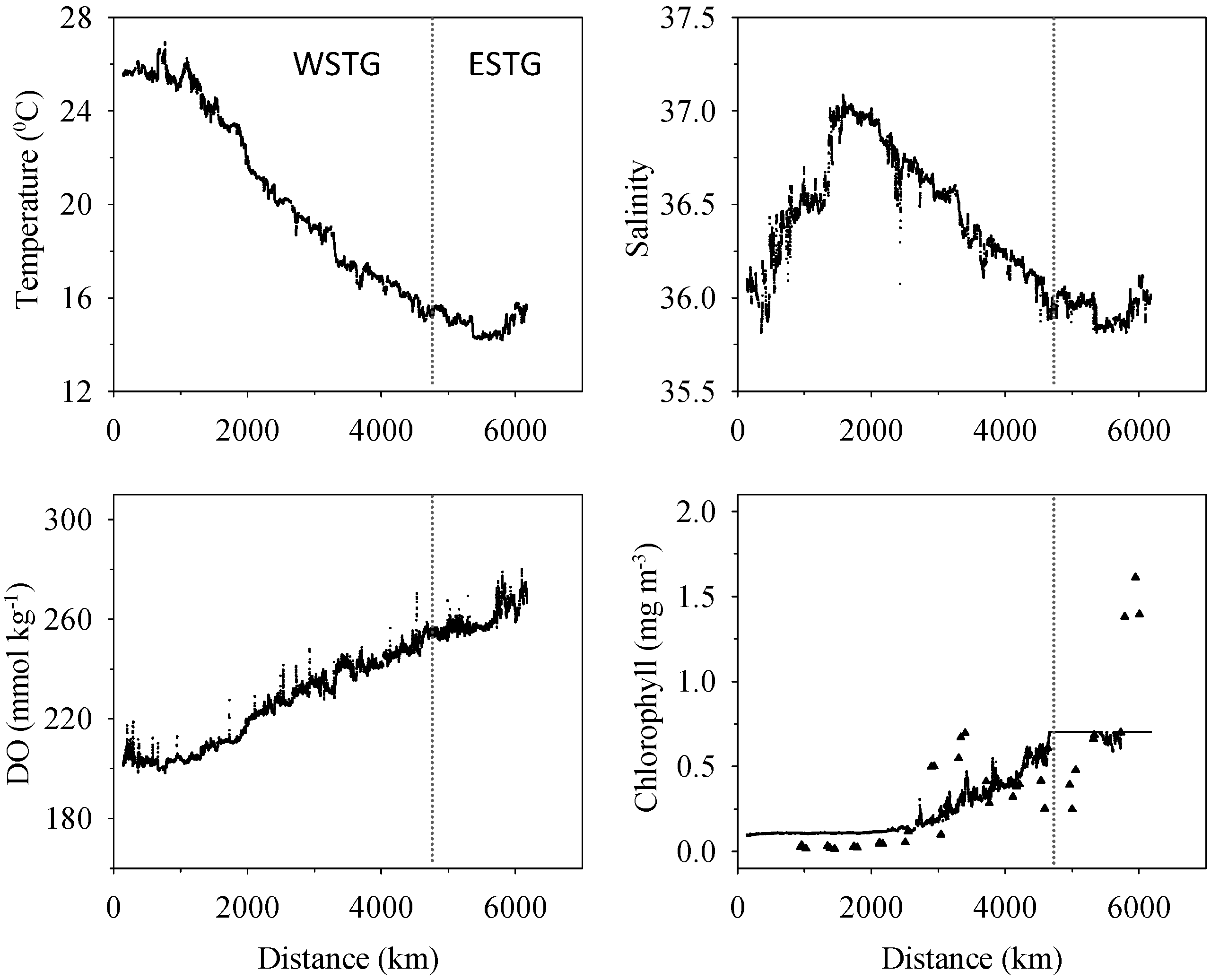

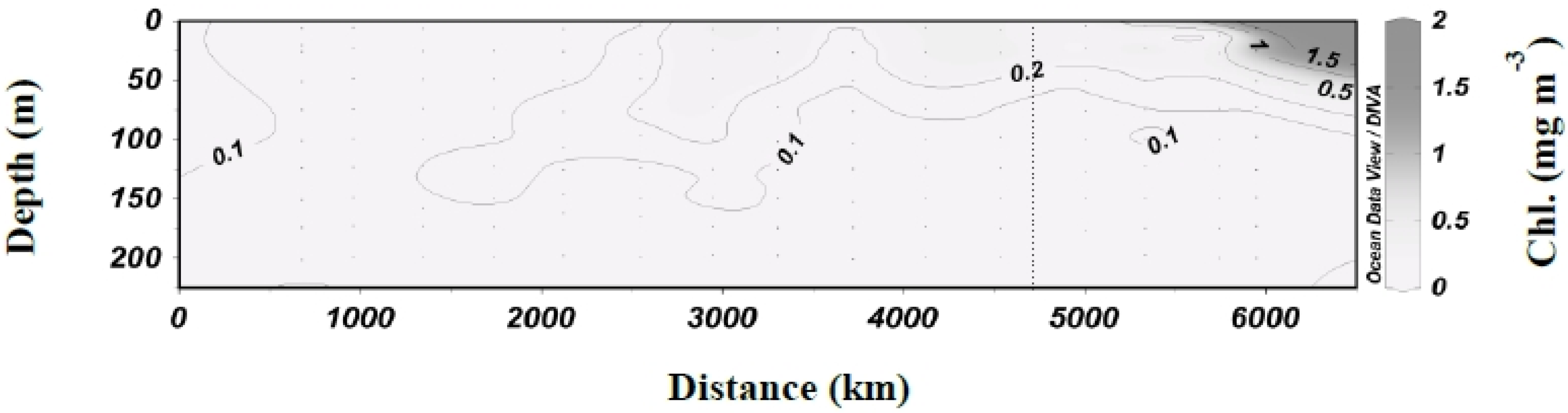
3.2. Inorganic Carbon System and Atmospheric Fluxes
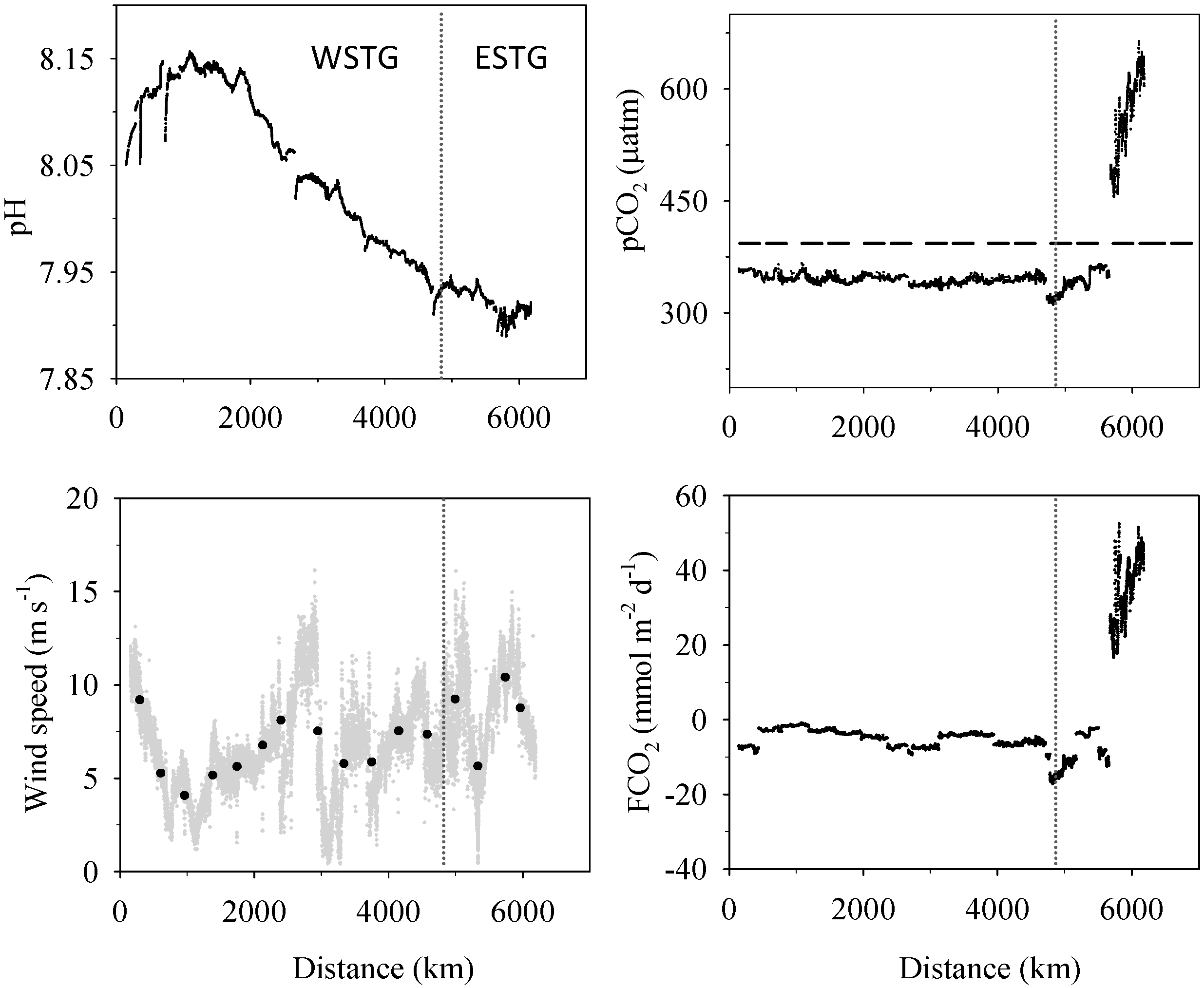
4. Discussion
4.1. Western North Atlantic Subtropical Gyre
| Period | Location | pCO2 (μatm) | SST (°C) | FCO2 (mmol m−2 d−1) | Reference |
|---|---|---|---|---|---|
| 1988–1998 | BATS site | 350 ± 50 | - | - | Bates (2001) [12] |
| 1994–1995 | Atl 20°–50° N | 330 ± 20 | - | −2.7 ± 4.1 | Schuster & Watson (2007) [7] |
| 1984–2000 | BATS site | 350 ± 50 | - | −0.7 ± 2.0 | Gruber et al., (2002) [13] |
| 2002–2005 | Atl 20°–50° N | 370 ± 10 | - | −1.2 ± 1.5 | Schuster & Watson (2007) [7] |
| 2005 | UK-Caribbean | - | - | −1.8 ± 0.1 | Watson et al., (2009) [8] |
| Spring 1970–2006 | Atl 14°–50° N | 310 ± 30 | - | −2.7 ± 2.7 | Takahashi et al., (2009) [6] |
| Spring 2004–2006 | Atl 14°–50° N | 340 ± 40 | 14.5 ± 14.4 | - | Telszwski et al., (2009) [42] |
| Spring 2000–2008 | Atl 27°–39° N | - | 18.6 ± 0.9 | −4.9 ± 3.3 | Padín et al., (2010) [43] |
| Spring 2011 | SGTW | 343 ± 8 | 19.8 ± 4.1 | −5.5 ± 2.2 | This work |
4.2. Eastern North Atlantic Subtropical Gyre
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Behrenfeld, M.J.; O’Malley, R.T.; Siegel, D.A.; McClain, C.R.; Sarmiento, J.L.; Feldman, G.C.; Milligan, A.J.; Falkowski, P.G.; Letelier, R.M.; Boss, E.S. Climate-driven trends in contemporary ocean productivity. Nature 2006, 444, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Schuster, U.; Watson, A.J.; Bates, N.R.; Corbiere, A.; Gonzalez-Davila, M.; Metzl, N.; Pierrot, D.; Santana-Casiano, M. Trends in North Atlantic sea-surface fCO2 from 1990 to 2006. Deep Sea Res. II Top. Stud. Oceanogr. 2009, 56, 620–629. [Google Scholar] [CrossRef]
- Körtzinger, A.; Send, U.; Lampitt, R.S.; Hartman, S.; Wallace, D.W.R.; Karstensen, J.; Llinás, O.; Villagarcia, M.G.; DeGrandpre, M.D. The seasonal pCO2 cycle at 49° N/16.5° W in the northeastern Atlantic Ocean and what it tells us about biological productivity. J. Geophys. Res. Oceans 2008, 113, 1–15. [Google Scholar] [CrossRef]
- Benson, N.U.; Osibanjo, O.O.; Asuquo, F.E.; Anake, W.U. Observed trends of pCO2 and air-sea CO2 fluxes in the North Atlantic Ocean. Int. J. Mar. Sci. 2014, 4, 1–7. [Google Scholar]
- Borges, A.V.; Frankignoulle, M. Distribution of surface carbon dioxide and air-sea exchange in the upwelling system off the Galician coast. Glob. Biogeochem. Cycles 2002, 16, 13-1–13-13. [Google Scholar] [CrossRef]
- Takahashi, T.; Sutherland, S.C.; Wanninkhof, R.; Sweeney, C.; Feely, R.A.; Chipman, D.W.; Hales, B.; Friederich, G.; Chavez, F.; Sabine, C.; et al. Climatological mean and decadal change in surface ocean pCO2, and net sea-air CO2 flux over the global oceans. Deep Sea Res. II Top. Stud. Oceanogr. 2009, 56, 554–577. [Google Scholar] [CrossRef] [Green Version]
- Schuster, U.; Watson, A.J. A variable and decreasing sink for atmospheric CO 2 in the North Atlantic. J. Geophys. Res. 2007, 112, C11006. [Google Scholar] [CrossRef]
- Watson, A.J.; Schuster, U.; Bakker, D.C.E.; Bates, N.R.; Corbière, A.; González-Dávila, M.; Friedrich, T.; Hauck, J.; Heinze, C.; Johannessen, T.; et al. Tracking the variable North Atlantic sink for atmospheric CO2. Science 2009, 326, 1391–1393. [Google Scholar] [CrossRef] [PubMed]
- Le Quéré, C.; Takahashi, T.; Buitenhuis, E.T.; Rödenbeck, C.; Sutherland, S.C. Impact of climate change and variability on the global oceanic sink of CO2. Glob. Biogeochem. Cycles 2010, 24. [Google Scholar] [CrossRef]
- Bates, N.R. Multi-decadal uptake of carbon dioxide into subtropical mode water of the North Atlantic Ocean. Biogeosciences 2012, 9, 2649–2659. [Google Scholar] [CrossRef]
- Santana-Casiano, J.M.; González-Dávila, M.; Rueda, M.J.; Llinás, O.; González-Dávila, E.F. The interannual variability of oceanic CO2 parameters in the northeast Atlantic subtropical gyre at the ESTOC site. Glob. Biogeochem. Cycles 2007, 21, 1–16. [Google Scholar] [CrossRef]
- Bates, N.R. Interannual variability of oceanic CO2 and biogeochemical properties in the Western North Atlantic subtropical gyre. Deep Sea Res. II Top. Stud. Oceanogr. 2001, 48, 1507–1528. [Google Scholar] [CrossRef]
- Gruber, N.; Keeling, C.D.; Bates, N.R. Interannual variability in the North Atlantic Ocean carbon sink. Science 2002, 298, 2374–2378. [Google Scholar] [CrossRef] [PubMed]
- Savidge, G.; Turner, D.R.; Burkill, P.H.; Watson, A.J.; Angel, M.V.; Pingree, R.D.; Leach, H.; Richards, K.J. The BOFS 1990 spring bloom experiment: Temporal evolution and spatial variability of the hydrographic field. Prog. Oceanogr. 1992, 29, 235–281. [Google Scholar] [CrossRef]
- Ducklow, H.W.; Harris, R.P. Introduction to the JGOFS North Atlantic bloom experiment. Deep Sea Res. II Top. Stud. Oceanogr. 1993, 40, 1–8. [Google Scholar] [CrossRef]
- Boyd, P.W.; Pomroy, A.; Bury, S.; Savidge, G.; Joint, I. Micro-algal carbon and nitrogen uptake in post-coccolithophore blooms in the northeast Atlantic, July 1991. Deep Sea Res. I Oceanogr. Res. Pap. 1997, 44, 1497–1517. [Google Scholar] [CrossRef]
- Koeve, W. C:N stoichiometry of the biological pump in the North Atlantic: Constraints from climatological data. Glob. Biogeochem. Cycles 2006, 20. [Google Scholar] [CrossRef]
- González-Dávila, M.; Santana-Casiano, J.M.; Rueda, M.J.; Llinás, O. The water column distribution of carbonate system variables at the ESTOC site from 1995 to 2004. Biogeosciences 2010, 7, 3067–3081. [Google Scholar] [CrossRef]
- McKinley, G.A.; Fay, A.R.; Takahashi, T.; Metzl, N. Convergence of atmospheric and North Atlantic carbon dioxide trends on multidecadal timescales. Nat. Geosci. 2011, 4, 606–610. [Google Scholar] [CrossRef]
- Andersson, A.J.; Krug, L.A.; Bates, N.R.; Doney, S.C. Sea-air CO2 flux in the North Atlantic subtropical gyre: Role and influence of Sub-Tropical Mode Water formation. Deep Sea Res. II Top. Stud. Oceanogr. 2013, 91, 57–70. [Google Scholar] [CrossRef]
- Sathyendranath, S.; Longhurst, A.; Caverhill, C.M.; Platt, T. Regionally and seasonally differntiated primary production in the North Atlantic. Deep Res. 1995, 42, 1773–1802. [Google Scholar] [CrossRef]
- Longhurst, A.; Sathyendranath, S.; Platt, T.; Caverhill, C. An estimate of global primary production in the ocean from satellite radiometer data. J. Plankton Res. 1995, 17, 1245–1271. [Google Scholar] [CrossRef]
- Marañón, E. Phytoplankton growth rates in the Atlantic subtropical gyres. Limnol. Oceanogr. 2005, 50, 299–310. [Google Scholar] [CrossRef]
- Pierrot, D.; Neill, C.; Sullivan, K.; Castle, R.; Wanninkhof, R.; Lüger, H.; Johannessen, T.; Olsen, A.; Feely, R.A.; Cosca, C.E. Recommendations for autonomous underway pCO2 measuring systems and data-reduction routines. Deep. Res. II Top. Stud. Oceanogr. 2009, 56, 512–522. [Google Scholar] [CrossRef]
- Dickson, A.G.; Sabine, C.L.; Christian, J.R. Guide to Best Practices for Ocean CO2 Measurements; North Pacific Marine Science Organization: Sidney, BC, Canada, 2007; Volume 3, p. 191. [Google Scholar]
- Millero, F.J. Seawater as A Multicomponent Electrolyte Solution; DTIC: Fort Belvoir, VA, USA, 1974. [Google Scholar]
- Takahashi, T.; Olafsson, J.; Goddard, J.G.; Chipman, D.W.; Sutherland, S.C. Seasonal variation of CO2 and nutrients in the high-latitude surface oceans: A comparative study. Glob. Biogeochem. Cycles 1993, 7, 843–878. [Google Scholar] [CrossRef]
- Grasshoff, K.; Ehrhardt, M. Automated chemical analysis. In Methods of Seawater Analysis; Springer-Verlag: Berlin, Germany, 1983; pp. 263–289. [Google Scholar]
- Weiss, R.F. The solubility of nitrogen, oxygen and argon in water and seawater. Deep Sea Res. Oceanogr. Abstr. 1970, 17, 721–735. [Google Scholar] [CrossRef]
- Clayton, T.D.; Byrne, R.H. Spectrophotometric seawater pH measurements: Total hydrogen results. Deep Sea Res. I Oceanogr. Res. Pap. 1993, 40, 2115–2129. [Google Scholar] [CrossRef]
- Perez, F.F.; Fraga, F. A precise and rapid analytical procedure for alkalinity determination. Mar. Chem. 1987, 21, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Lueker, T.J.; Dickson, A.G.; Keeling, C.D. Ocean pCO2 calculated from dissolved inorganic carbon. Mar. Chem. 2000, 70, 105–119. [Google Scholar] [CrossRef]
- Weiss, R.F. Carbon dioxide in water and seawater: The solubility of a non-ideal gas. Mar. Chem. 1974, 2, 203–215. [Google Scholar] [CrossRef]
- Wanninkhof, R. Relationship between wind speed and gas exchange over the ocean. J. Geophys. Res. Ocean. 1992, 97, 7373–7382. [Google Scholar] [CrossRef]
- NOAA-ESRL Global Monitoring Division Station. Available online: http://www.macehead.org (accessed on 19 May 2015).
- Keller, K.M.; Joos, F.; Raible, C.C.; Cocco, V.; Frölicher, T.L.; Dunne, J.P.; Gehlen, M.; Bopp, L.; Orr, J.C.; Tjiputra, J.; et al. Variability of the ocean carbon cycle in response to the North Atlantic Oscillation. Tellus B 2012, 1, 1–25. [Google Scholar] [CrossRef]
- Redfield, A.C.; Ketchum, B.K.; Richards, F.R. The Influence of Organism on the Composition of Sea-Water. The Composition of Seawater. Comparative and Descriptive Oceanography. The Sea: Ideas and Observations on Progress in the Study of the Seas; Interscience Publishers: New York, NY, USA, 1963. [Google Scholar]
- Menzel, D.W.; Ryther, J.H. Nutrients limiting the production of phytoplankton in the Sargasso Sea, with special reference to iron. Deep Sea Res. 1961, 7, 276–281. [Google Scholar] [CrossRef]
- Sowell, S.M.; Wilhelm, L.J.; Norbeck, A.D.; Lipton, M.S.; Nicora, C.D.; Barofsky, D.F.; Carlson, C.A.; Smith, R.D.; Giovanonni, S.J. Transport functions dominate the SAR11 metaproteome at low-nutrient extremes in the Sargasso Sea. ISME J. 2008, 3, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Cianca, A.; Helmke, P.; Mouriño, B.; Rueda, M.J.; Llinás, O.; Neuer, S. Decadal analysis of hydrography and in situ nutrient budgets in the western and eastern North Atlantic subtropical gyre. J. Geophys. Res. Ocean. 2007, 112, 1–18. [Google Scholar] [CrossRef]
- Bates, N.R.; Best, M.H.P.; Neely, K.; Garley, R.; Dickson, A.G.; Johnson, R.J. Detecting anthropogenic carbon dioxide uptake and ocean acidification in the North Atlantic Ocean. Biogeosciences 2012, 9, 2509–2522. [Google Scholar] [CrossRef]
- Telszewski, M.; Chazottes, A.; Schuster, U.; Watson, A.J.; Moulin, C.; Bakker, D.C.E.; González-Dávila, M.; Johannessen, T.; Krtzinger, A.; Lüger, H.; et al. Estimating the monthly pCO2 distribution in the North Atlantic using a self-organizing neural network. Biogeosciences 2009, 6, 1405–1421. [Google Scholar] [CrossRef] [Green Version]
- Padin, X.A.; Vázquez-Rodríguez, M.; Castaño, M.; Velo, A.; Alonso-Pérez, F.; Gago, J.; Gilcoto, M.; Álvarez, M.; Pardo, P.C.; de la Paz, M.; et al. Air-Sea CO2 fluxes in the Atlantic as measured during boreal spring and autumn. Biogeosciences 2010, 7, 1587–1606. [Google Scholar] [CrossRef]
- Boye, M.; Aldrich, A.; van den Berg, C.M.G.; de Jong, J.T.M.; Nirmaier, H.; Veldhuis, M.; Timmermans, K.R.; de Baar, H.J.W. The chemical speciation of iron in the north-east Atlantic Ocean. Deep Sea Res. I Oceanogr. Res. Pap. 2006, 53, 667–683. [Google Scholar] [CrossRef]
- González Dávila, M.; Santana-Casiano, J.M.; Merlivat, L.; Barbero-Muñoz, L.; Dafner, E.V. Fluxes of CO2 between the atmosphere and the ocean during the POMME project in the northeast Atlantic Ocean during 2001. J. Geophys. Res. C Ocean. 2005, 110, 1–14. [Google Scholar] [CrossRef]
- Kähler, P.; Oschlies, A.; Dietze, H.; Koeve, W. Oxygen, carbon, and nutrients in the oligotrophic eastern subtropical North Atlantic. Biogeosciences 2010, 7, 1143–1156. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Salgado, X.A.; Rosón, G.; Pérez, F.F.; Pazos, Y. Hydrographic variability off the Rías Baixas (NW Spain) during the upwelling season. J. Geophys. Res. Ocean. 1993, 98, 14447–14455. [Google Scholar] [CrossRef]
- Álvarez-Salgado, X.A.; Nieto-Cid, M.; Gago, J.; Brea, S.; Castro, C.G.; Doval, M.D.; Pérez, F.F. Stoichiometry of the degradation of dissolved and particulate biogenic organic matter in the NW Iberian upwelling. J. Geophys. Res. 2006, 111, C07017. [Google Scholar] [CrossRef]
- Cobo-Viveros, A.M.; Antonio Padin, X.; Otero, P.; de la Paz, M.; Ruiz-Villarreal, M.; Ríos, A.F.; Pérez, F.F. Short-term variability of surface carbon dioxide and sea-air CO2 fluxes in the shelf waters of the Galician coastal upwelling system. Sci. Mar. 2013, 77, 37–48. [Google Scholar] [CrossRef]
- Hales, B.; Takahashi, T.; Bandstra, L. Atmospheric CO2 uptake by a coastal upwelling system. Glob. Biogeochem. Cycles 2005, 19, 1–11. [Google Scholar] [CrossRef]
- Pérez, F.F.; Castro, C.G.; Ríos, A.F.; Fraga, F. Chemical properties of the deep winter mixed layer in the Northeast Atlantic (40°–47° N). J. Mar. Syst. 2005, 54, 115–125. [Google Scholar] [CrossRef]
- Harris, K.E.; Degrandpre, M.D.; Hales, B.R. Continuous Time-Series of Carbonate System Dynamics in the Coastal Oregon Upwelling System. In Proceeding of the 2010 AGU Fall Meeting, San Francisco, CA, USA, 2010; Volume 1, p. 1568.
- Mahadevan, A.; Lévy, M.; Mémery, L. Mesoscale variability of sea surface pCO2: What does it respond to? Glob. Biogeochem. Cycles 2004, 18, 127–142. [Google Scholar] [CrossRef]
- Körtzinger, A.; Koeve, W.; Kähler, P.; Mintrop, L. C :N ratios in the mixed layer during the productive season in the northeast Atlantic Ocean. Deep. Res. I Oceanogr. Res. Pap. 2001, 48, 661–688. [Google Scholar] [CrossRef]
- Backhaus, J.O.; Wehde, H.; Hegseth, E.N.; Kämpf, J. “Phyto-convection”: The role of oceanic convection in primary production. Mar. Ecol. Prog. Ser. 1999, 189, 77–92. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burgos, M.; Sendra, M.; Ortega, T.; Ponce, R.; Gómez-Parra, A.; Forja, J.M. Ocean-Atmosphere CO2 Fluxes in the North Atlantic Subtropical Gyre: Association with Biochemical and Physical Factors during Spring. J. Mar. Sci. Eng. 2015, 3, 891-905. https://doi.org/10.3390/jmse3030891
Burgos M, Sendra M, Ortega T, Ponce R, Gómez-Parra A, Forja JM. Ocean-Atmosphere CO2 Fluxes in the North Atlantic Subtropical Gyre: Association with Biochemical and Physical Factors during Spring. Journal of Marine Science and Engineering. 2015; 3(3):891-905. https://doi.org/10.3390/jmse3030891
Chicago/Turabian StyleBurgos, Macarena, Marta Sendra, Teodora Ortega, Rocio Ponce, Abelardo Gómez-Parra, and Jesús M. Forja. 2015. "Ocean-Atmosphere CO2 Fluxes in the North Atlantic Subtropical Gyre: Association with Biochemical and Physical Factors during Spring" Journal of Marine Science and Engineering 3, no. 3: 891-905. https://doi.org/10.3390/jmse3030891