Water Use and Water-Use Efficiency of Three Perennial Bioenergy Grass Crops in Florida

Abstract
:1. Introduction
2. Experimental Section
2.1. Site Description and Experimental Design
2.2. Weather and Water Inputs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | TAIR (°C) | RH (%) | Q (W·m−2) | Rainfall (mm) | Irrigation (mm) |
|---|---|---|---|---|---|
| 2009 | 23.6 | 79.1 | 195 | 897 | 430 |
| 2010 | 24.4 | 77.0 | 213 | 585 | 405 |
2.3. Cultural Practices and Aboveground Biomass Yield
2.4. Water Use and Water-Use Efficiency
2.5. Soil Volumetric Water Content
2.6. Leaf Gas Exchange and Root Biomass
2.7. Data Analyses
3. Results and Discussion
3.1. Crop Morphology and Aboveground Biomass Yield
| Species | Dry biomass yield (Mg ha−1) | Stem density (per m2) | Stem diameter (mm) | |||
|---|---|---|---|---|---|---|
| 2009 | 2010 | 2009 | 2010 | 2009 | 2010 | |
| GR | 13.4B † | 29.1B | 14.5C | 24.9A | 13.5B | 14.8B |
| EC | 38.4A | 40.9A | 25.3A | 26.7A | 15.0AB | 15.0AB |
| EG | 35.5A | 38.9A | 19.3B | 20.6A | 17.1A | 16.3A |
| s.e ‡ | 2.61 | 2.75 | 1.55 | 2.23 | 0.73 | 0.58 |
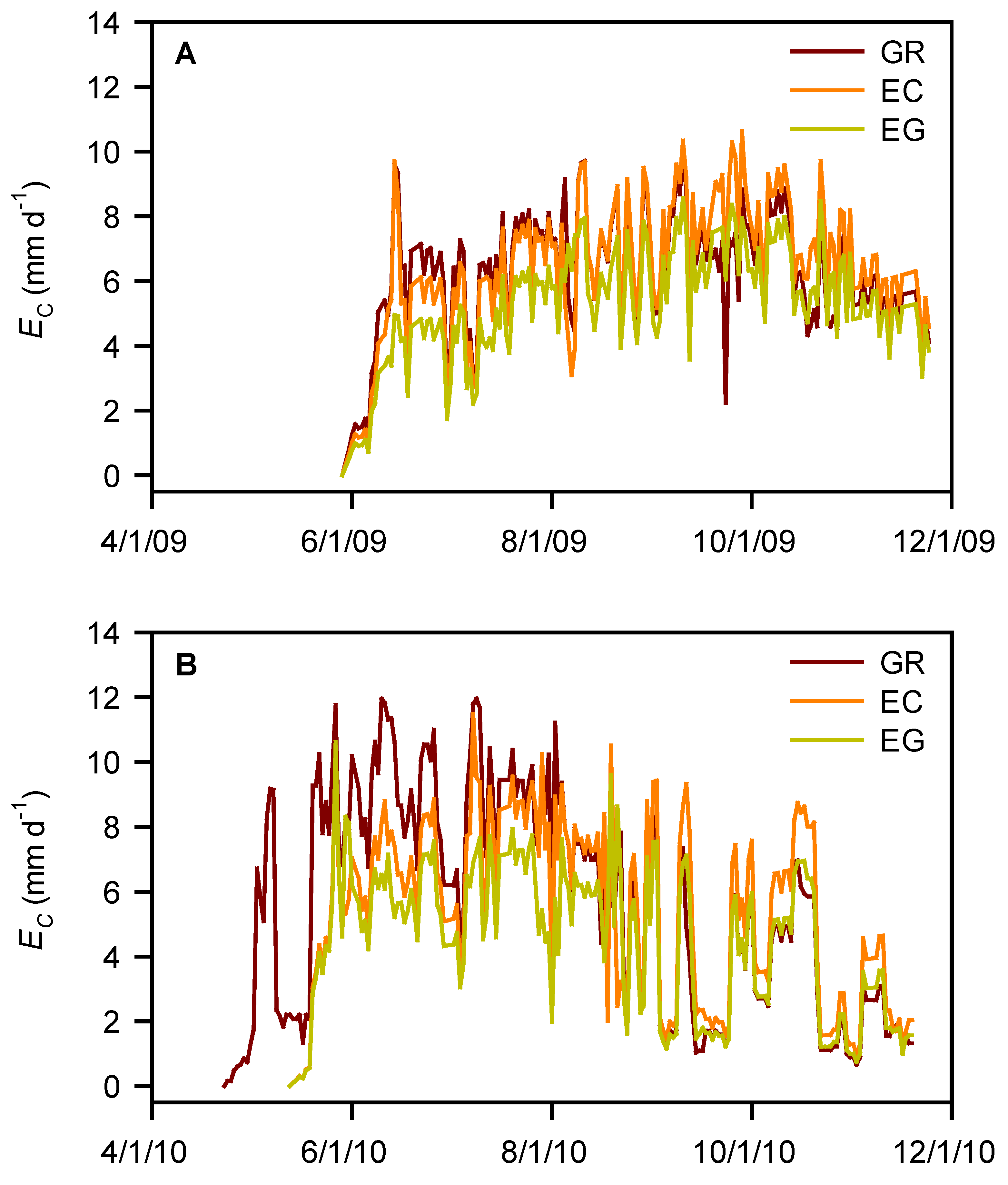
3.2. Water Use and Water-Use Efficiency
| Species | EC (mm) | WUE (g kg−1) | ||
|---|---|---|---|---|
| 2009 | 2010 | 2009 | 2010 | |
| GR | 1113AB† | 1177A | 1.19B | 2.47B |
| EC | 1151A | 1035B | 3.35A | 3.96A |
| EG | 930B | 856C | 3.84A | 4.57A |
| s.e ‡ | 77.1 | 33.3 | 0.24 | 0.32 |


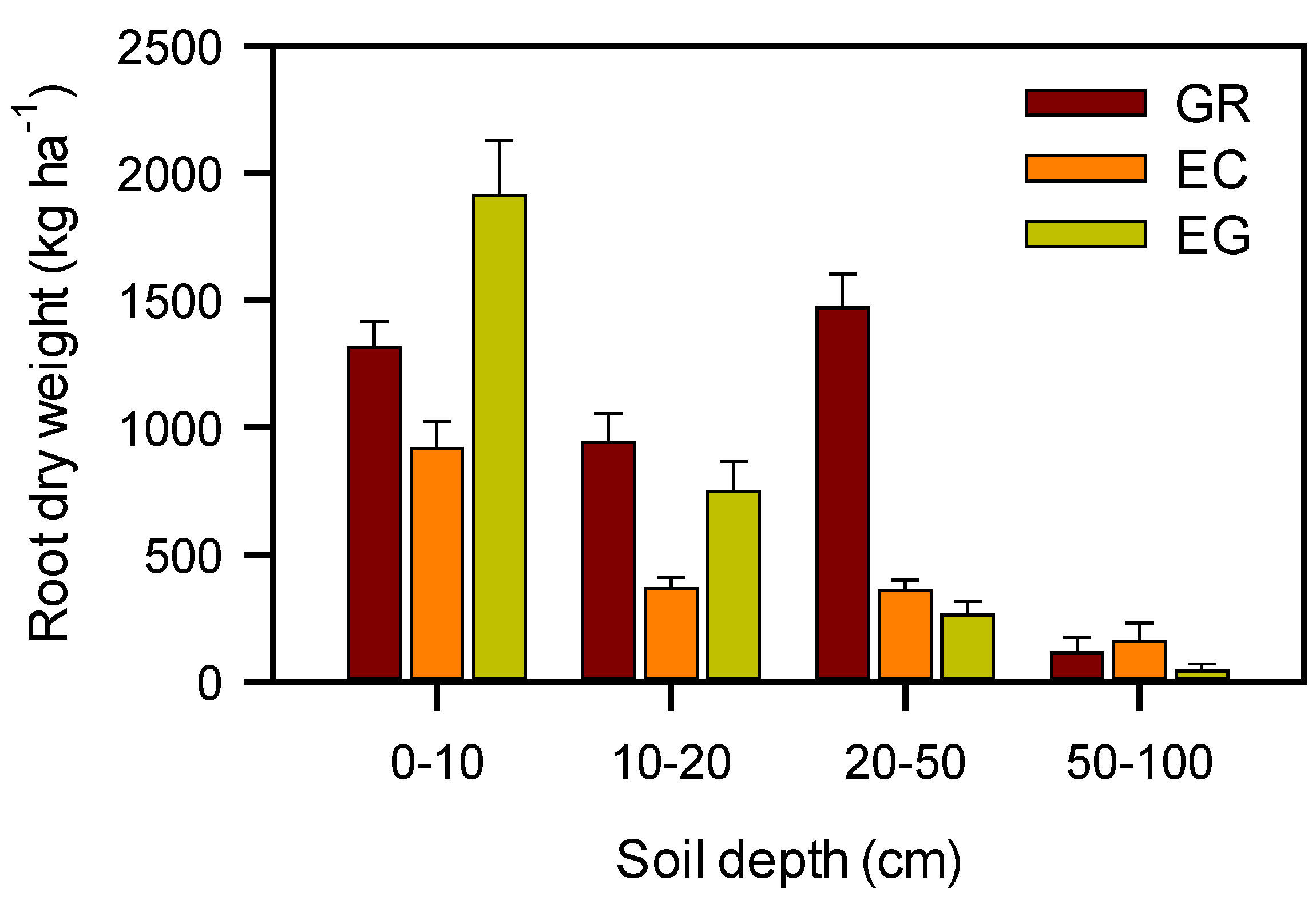
3.3. Leaf Gas Exchange and Root Biomass
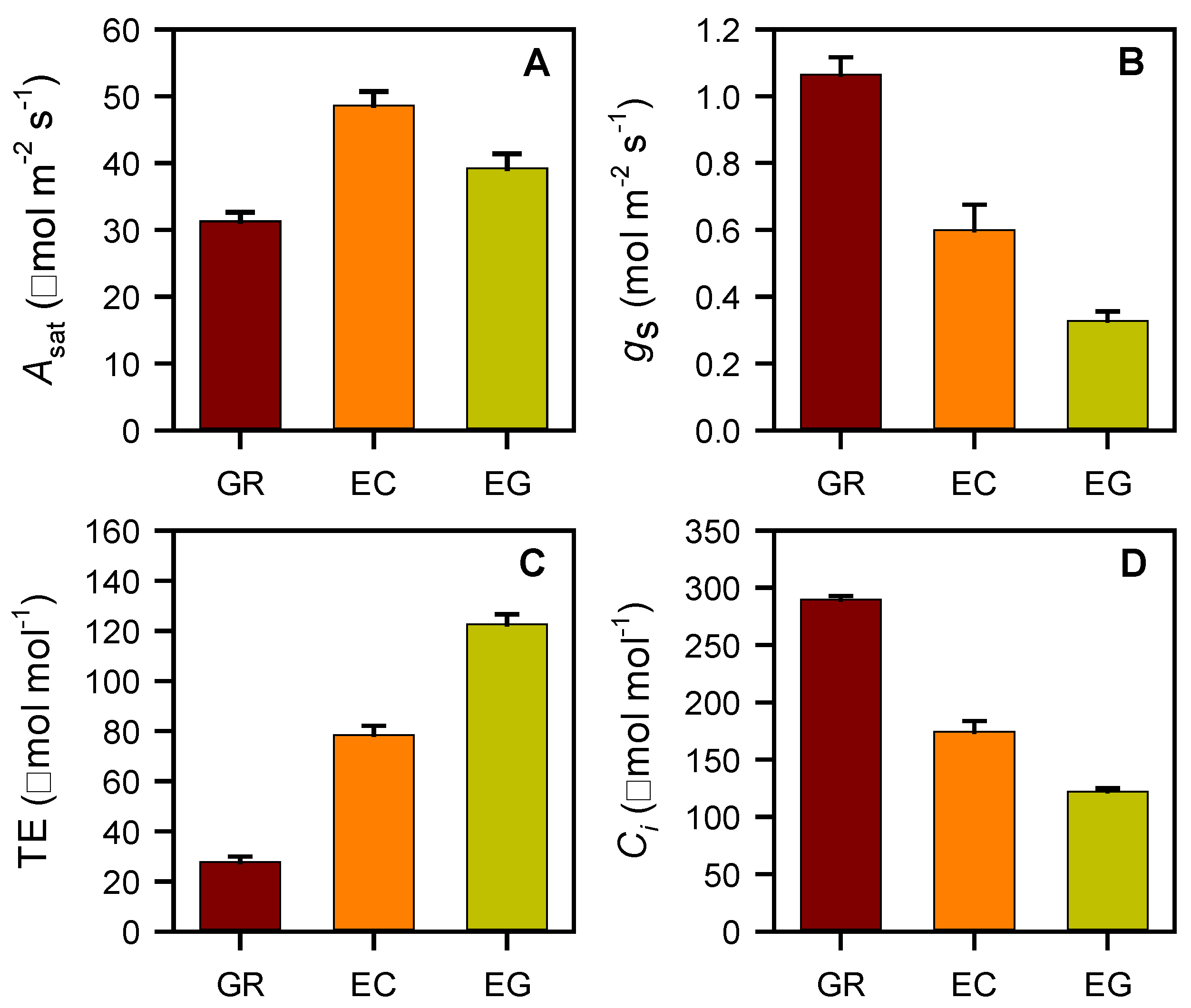
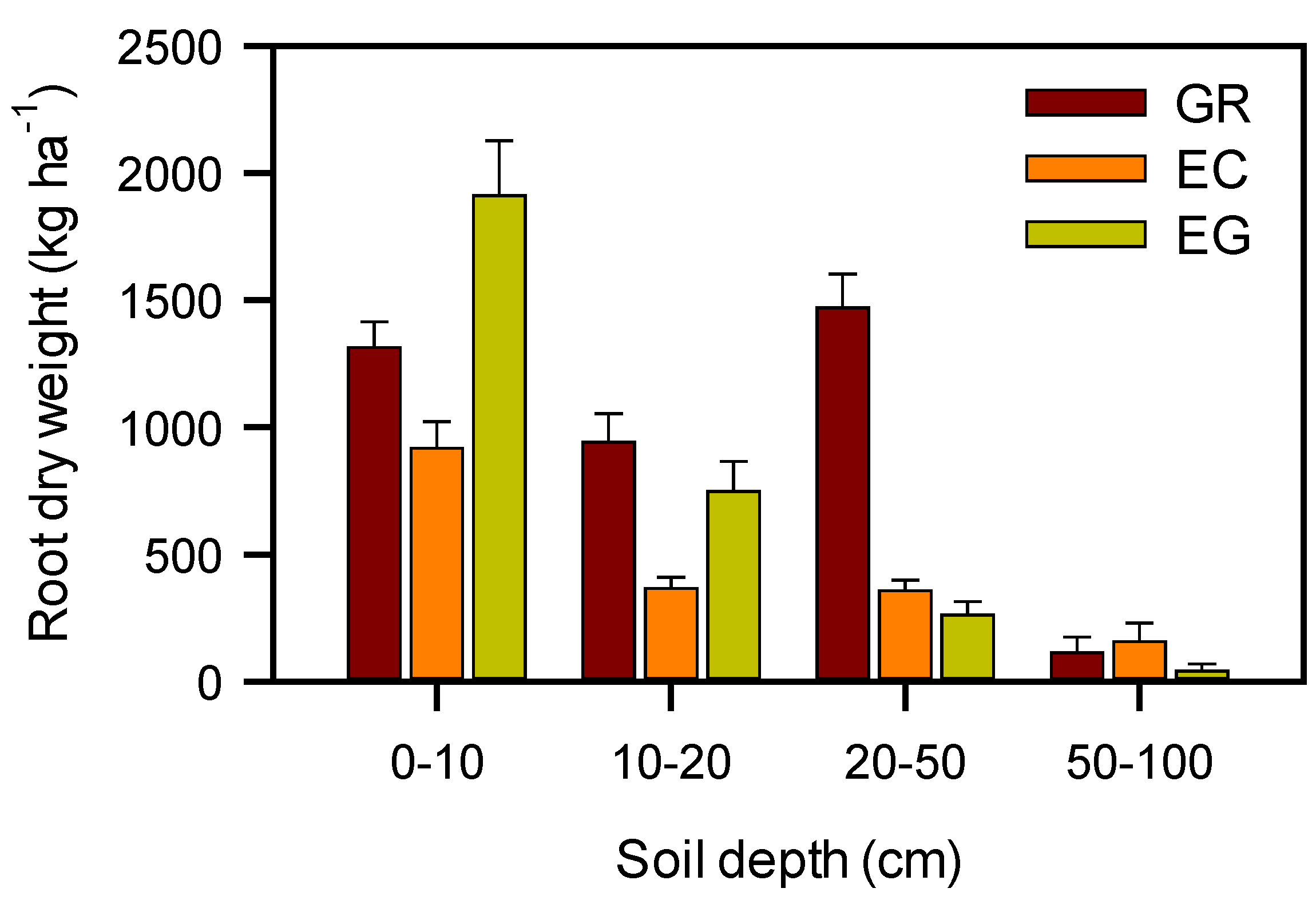
4. Conclusions
Acknowledgments
References
- Balat, M.; Balat, H. Recent trends in global production and utilization of bio-ethanol fuel. Appl. Energy 2009, 86, 2273–2282. [Google Scholar] [CrossRef]
- Margeot, A.; Hahn-Hagerdal, B.; Edlund, M.; Slade, R.; Monot, F. New improvements for lignocellulosic ethanol. Curr. Opin. Biotechnol. 2009, 20, 372–380. [Google Scholar] [CrossRef]
- Woodard, K.R.; Prine, G.M. Dry matter accumulation of elephantgrass, energycane, and elephantmillet in a subtropical climate. Crop Sci. 1993, 33, 818–824. [Google Scholar] [CrossRef]
- Knoll, J.E.; Anderson, W.F.; Strickland, T.C. Low-input production of biomass from perennial grasses in the Coastal Plain of Georgia, USA. Bioenerg. Res. 2012, 5, 206–214. [Google Scholar] [CrossRef]
- Hill, J.; Nelson, E.; Tilman, D.; Polasky, S.; Tiffany, D. Environmental, economic, and energetic costs and benefits of biodiesel and ethanol biofuels. Proc. Nat. Acad. Sci. USA 2006, 103, 11206–11210. [Google Scholar]
- Berndes, G. Bioenergy and water—The implications of large-scale bioenergy production for water use and supply. Glob. Environ. Chang. 2002, 12, 253–271. [Google Scholar] [CrossRef]
- Evans, J.M.; Cohen, M.J. Regional water resource implications of bioethanol production in the southeastern United States. Glob. Chang. Biol. 2009, 15, 2261–2273. [Google Scholar] [CrossRef]
- Angelini, L.G.; Ceccarini, L.; Bonari, E. Biomass yield and energy balance of giant reed (Arundo donax L.) cropped in central Italy as related to different management practices. Eur. J. Agron. 2005, 22, 375–389. [Google Scholar]
- Christian, D.G.; Riche, A.B.; Yates, N.E. Growth, yield and mineral content of Miscanthus × giganteus grown as a biofuel for 14 successive harvests. Ind. Crop. Prod. 2008, 28, 320–327. [Google Scholar] [CrossRef]
- Kering, M.K.; Biermacher, J.T.; Butler, T.J.; Mosali, J.; Guretzky, J.A. Biomass yield and nutrient responses of switchgrass to phosphorus application. Bioenerg. Res. 2012, 5, 71–78. [Google Scholar] [CrossRef]
- Burvall, J. Influence of harvest time and soil type on fuel quality in reed canary grass (Phalaris arundinacea L.). Biomass Bioenerg. 1997, 12, 149–154. [Google Scholar]
- Stanhill, G. Water use efficiency. Adv. Agron. 1986, 39, 53–85. [Google Scholar] [CrossRef]
- Beale, C.V.; Morison, J.I.L.; Long, S.P. Water use efficiency of C4 perennial grasses in a temperate climate. Agric. For. Meteorol. 1999, 96, 103–115. [Google Scholar] [CrossRef]
- Berry, J.; Bjorkman, O. Photosynthetic response and adaptation to temperature in higher plants. Annu. Rev. Plant Phys. 1980, 31, 491–543. [Google Scholar] [CrossRef]
- FAO. FAO Aquastat; FAO: Rome, Italy, 2012. Available online: http://www.fao.org/nr/water/aquastat/water_use/index.stm (accessed on 21 July 2012).
- Marris, E. Water: More crop per drop. Nature 2008, 452, 273–277. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Tanner, C.B.; Bennett, J.M. Water-Use efficiency in crop production. Bioscience 1984, 34, 36–40. [Google Scholar] [CrossRef]
- Baker, J.M.; van Bavel, C.H.M. Measurement of mass flow of water in the stems of herbaceous plants. Plant Cell Environ. 1987, 10, 777–782. [Google Scholar]
- Sakuratani, T. A heat balance method for measuring water flux in the stem of intact plants. Agric. Meteorol. 1981, 37, 9–17. [Google Scholar] [CrossRef]
- Dugas, W.A.; Prior, S.A.; Rogers, H.H. Transpiration from sorghum and soybean growing under ambient and elevated CO2 concentrations. Agric. For. Meteorol. 1998, 83, 37–48. [Google Scholar]
- Bethenod, O.; Katerji, N.; Goujet, R.; Bertolini, J.M.; Rana, G. Determination and validation of corn crop transpiration by sap flow measurement under field conditions. Theor. Appl. Climatol. 2000, 67, 153–160. [Google Scholar] [CrossRef]
- Saliendra, N.Z.; Meinzer, F.C. Genotypic, developmental and drought-induced differences in root hydraulic conductance of contrasting sugarcane cultivars. J. Exp. Bot. 1992, 43, 1209–1217. [Google Scholar] [CrossRef]
- Topp, G.C.; Davis, J.L. Measurement of soil water content using time-domain reflectometry (TDR): A field evaluation. Soil Sci. Soc. Amer. J. 1985, 49, 19–24. [Google Scholar]
- Dirksen, C.; Dasberg, S. Improved calibration of time domain reflectometry soil water content measurements. Soil Sci. Soc. Am. J. 1993, 57, 660–667. [Google Scholar]
- Propheter, J.L.; Staggenborg, S.A.; Wu, X.; Wang, D. Performance of annual and perennial biofuel crops: Yield during the first two years. Agron. J. 2010, 102, 806–814. [Google Scholar] [CrossRef]
- Wiggans, D.R.; Singer, J.W.; Moore, K.J.; Lamkey, K.R. Maize water use in living mulch systems with stover removal. Crop Sci. 2012, 52, 327–338. [Google Scholar] [CrossRef]
- Jara, J.; Stockle, C.O.; Kjelgaard, J. Measurement of evapotranspiration and its components in a corn (Zea mays L.) field. Agric. For. Meteorol. 1998, 92, 131–145. [Google Scholar]
- Azam-Alia, S.N.; Gregorya, P.J.; Monteitha, J.L. Effects of planting density on water use and productivity of pearl millet (Pennisetum typhoides) grown on stored water: II. Water use, light interception and dry matter production. Exp. Agric. 1984, 20, 215–224. [Google Scholar] [CrossRef]
- Sumner, D.M.; Jacobs, J.M. Utility of Penman-Monteith, Priestly-Taylor, reference evapotranspiration, and pan evaporation methods to estimate pasture evapotranspiration. J. Hydrol. 2005, 308, 81–104. [Google Scholar] [CrossRef]
- McGroary, P.C.; Cisar, J.L.; Snyder, G.H.; Erickson, J.E.; Daroub, S.H.; Sartain, J.B. Water use of St. Augustinegrass and bahiagrass under varying nitrogen rates. Agron. J. 2011, 103, 100–106. [Google Scholar] [CrossRef]
- Connor, D.J.; Lommis, R.S.; Cassman, K.G. Crop Ecology: Productivity and Management in Agricultural Systems, 2nd ed; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration. In Guidelines for Computing Crop Water Requirements; FAO Irrigation and Drainage Paper 56 for Food and Agriculture Organization of United Nations: Rome, Italy, 1998. [Google Scholar]
- Chabot, R.; Bouarfa, S.; Zimmer, D.; Chaumont, C.; Moreau, S. Evaluation of the sap flow determined with a heat balance method to measure the transpiration of a sugarcane canopy. Agric. Water Manage 2005, 75, 10–24. [Google Scholar] [CrossRef]
- Fougerouze, J. Quelques problemes de bioclimatologie en Guyanne Francaise. Agron. Trop. 1966, 21, 291–346. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Erickson, J.E.; Soikaew, A.; Sollenberger, L.E.; Bennett, J.M. Water Use and Water-Use Efficiency of Three Perennial Bioenergy Grass Crops in Florida. Agriculture 2012, 2, 325-338. https://doi.org/10.3390/agriculture2040325
Erickson JE, Soikaew A, Sollenberger LE, Bennett JM. Water Use and Water-Use Efficiency of Three Perennial Bioenergy Grass Crops in Florida. Agriculture. 2012; 2(4):325-338. https://doi.org/10.3390/agriculture2040325
Chicago/Turabian StyleErickson, John E., Arkorn Soikaew, Lynn E. Sollenberger, and Jerry M. Bennett. 2012. "Water Use and Water-Use Efficiency of Three Perennial Bioenergy Grass Crops in Florida" Agriculture 2, no. 4: 325-338. https://doi.org/10.3390/agriculture2040325