
Characteristics of Carbon, Nitrogen and Phosphorus Stoichiometry and Nutrient Reabsorption in Alfalfa Leaves with Different Fall-Dormancy Levels in Northern Xinjiang, China

Abstract
:1. Introduction
2. Materials and Methods
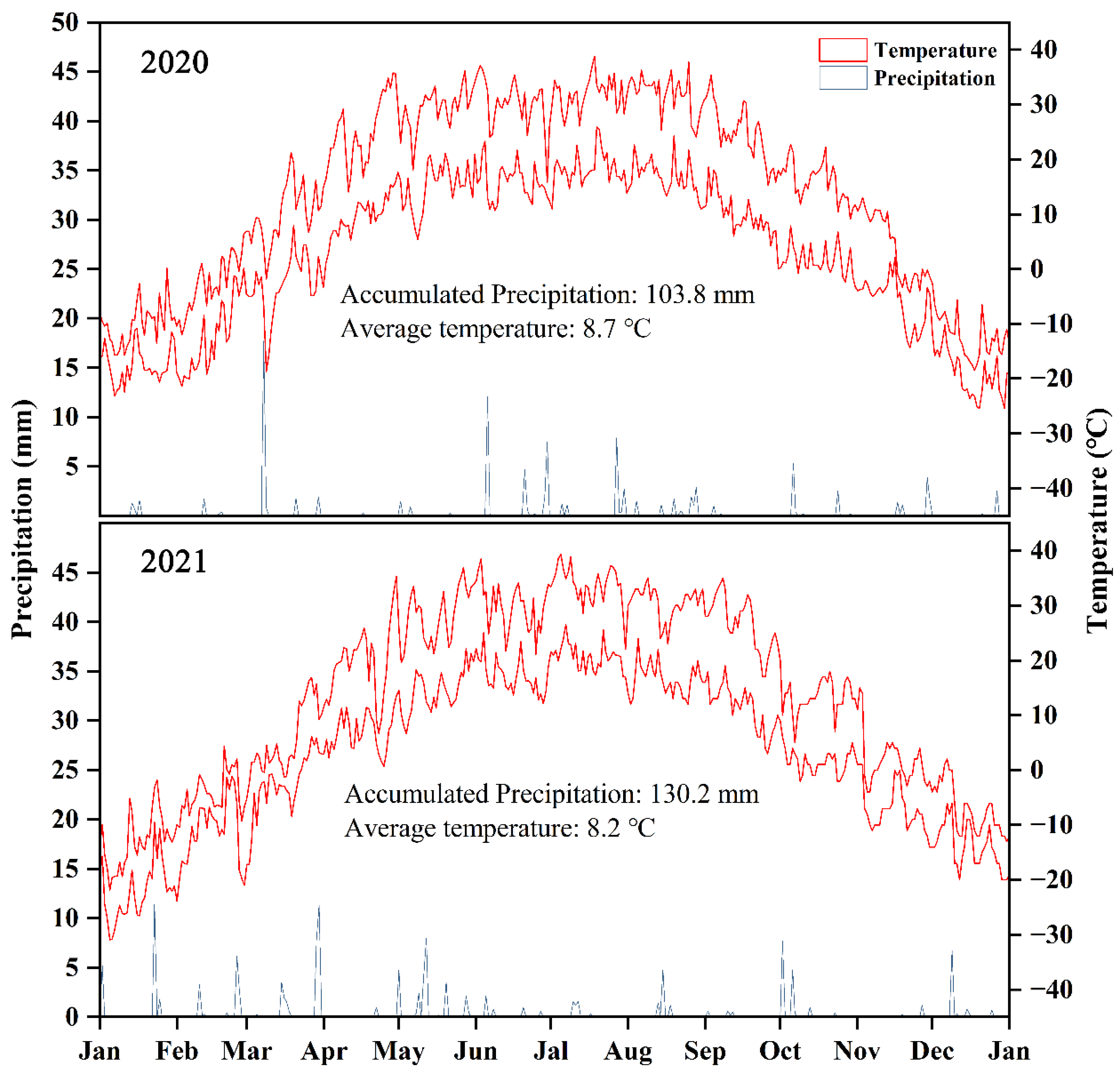
2.1. Experimental Site
2.2. Experimental Materials and Experimental Design
2.3. Measurement Indexes and Methods
2.4. Data Analysis
3. Results
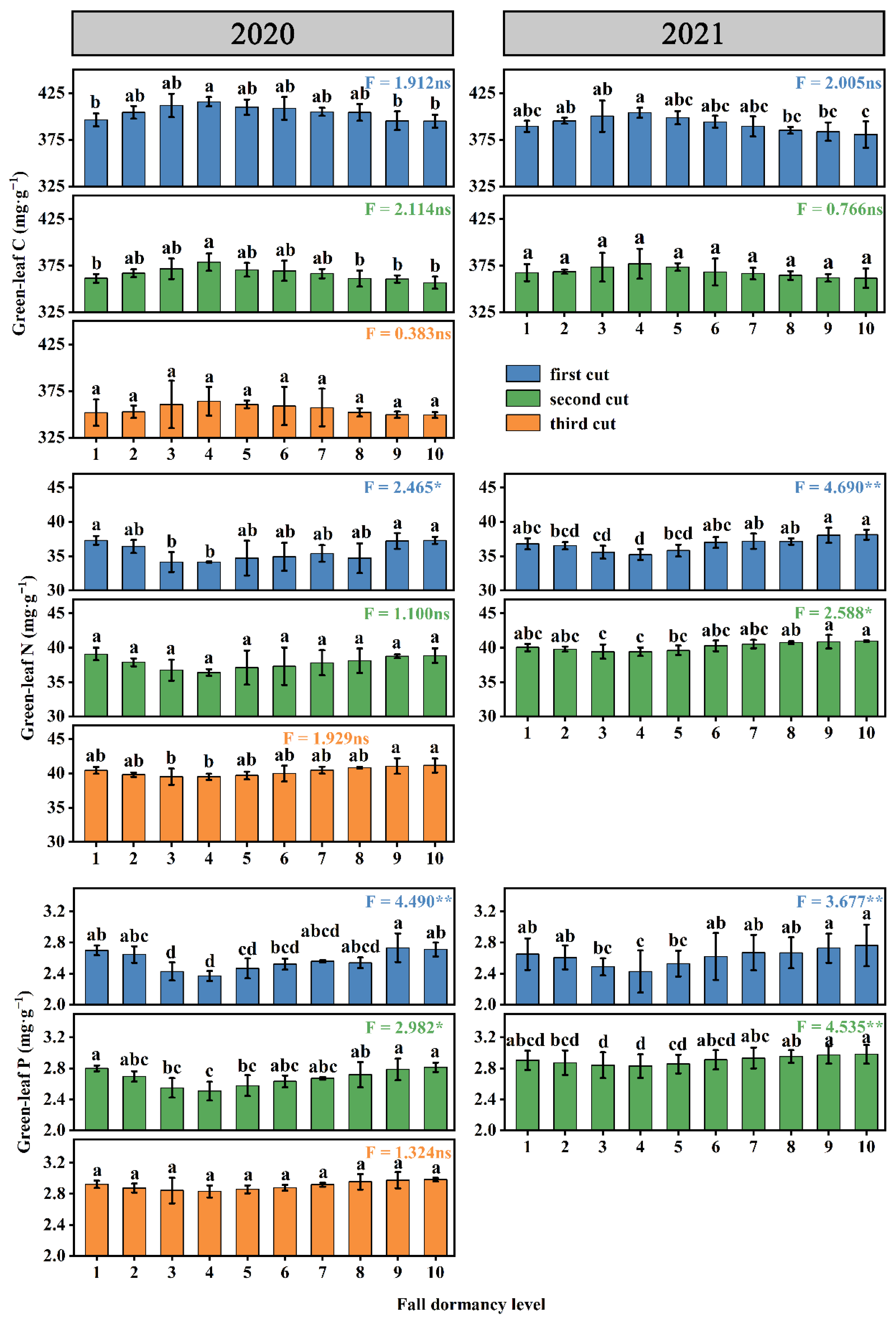
3.1. C, N, and P Concentrations in Mature Leaves of Alfalfa with Different Fall-Dormancy Levels
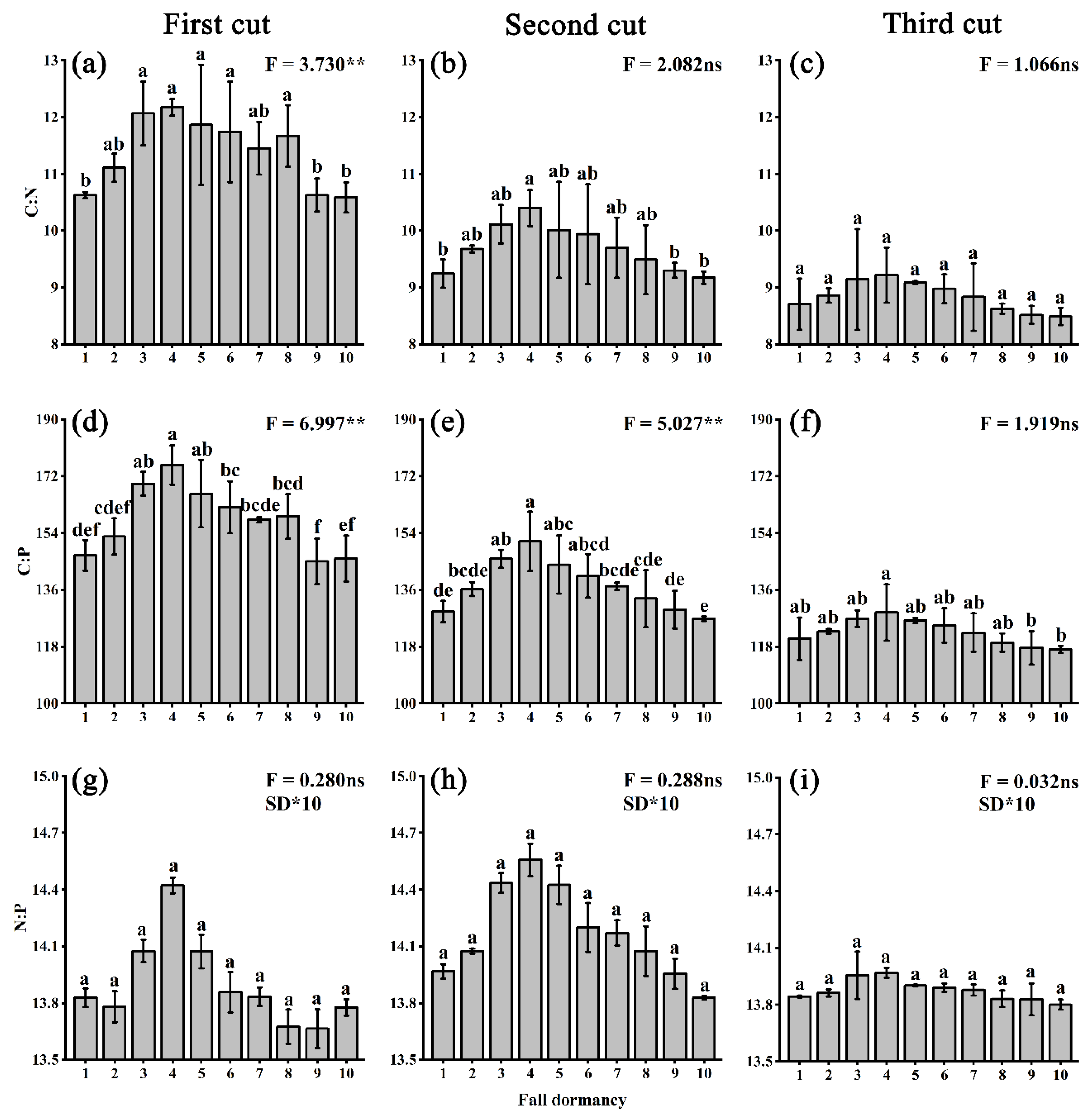
3.2. Stoichiometry Ratios of Elements in Mature Alfalfa Leaves with Different Fall-Dormancy Levels
3.3. C, N, and P Concentrations in Senescent Alfalfa Leaves with Different Fall-Dormancy Levels
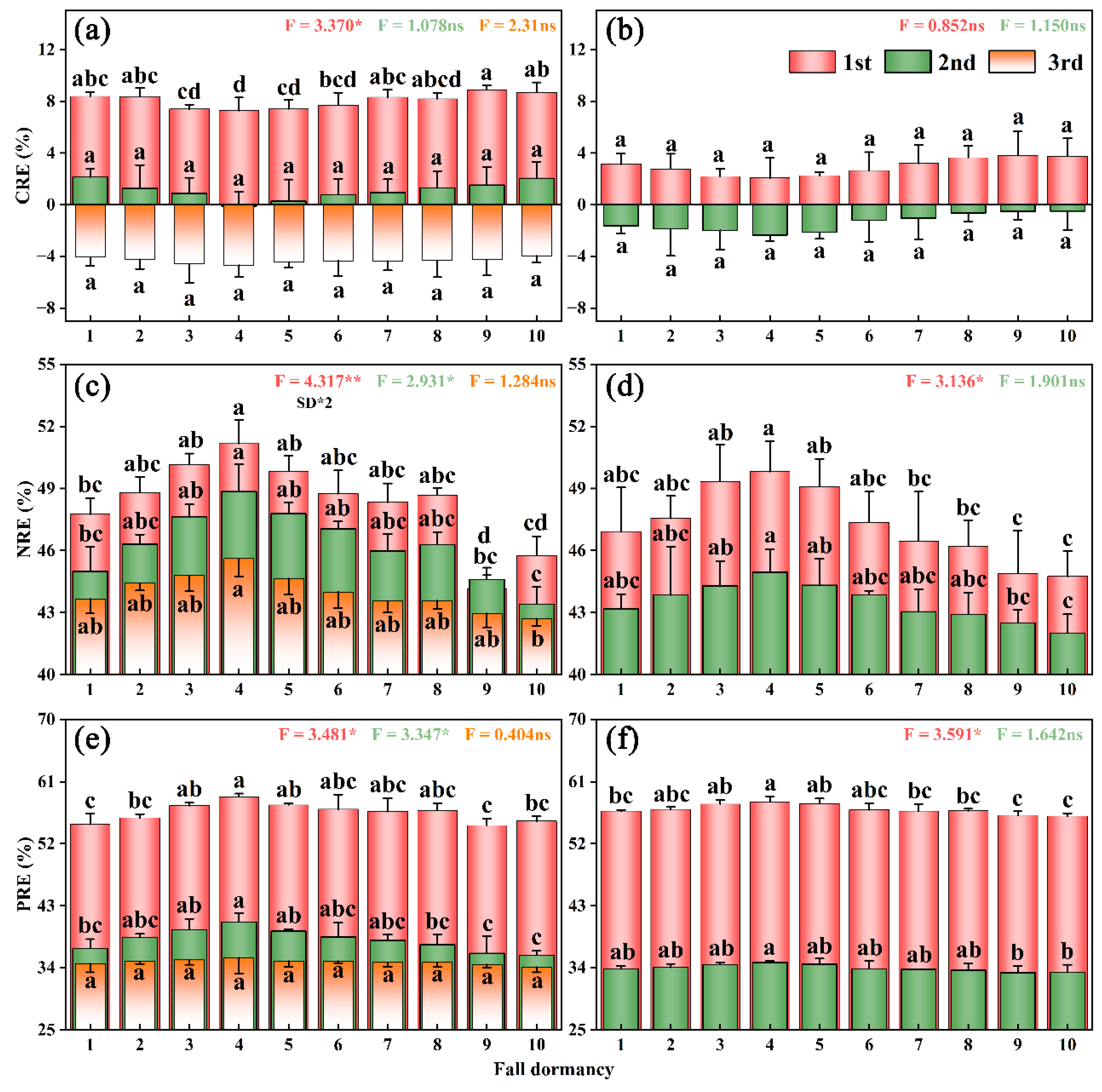
3.4. Nutrient Resorption Characteristics of Alfalfa with Different Fall-Dormancy Levels
3.5. Comparison of Survival Rates of Alfalfa Varieties with Different Fall-Dormancy Levels
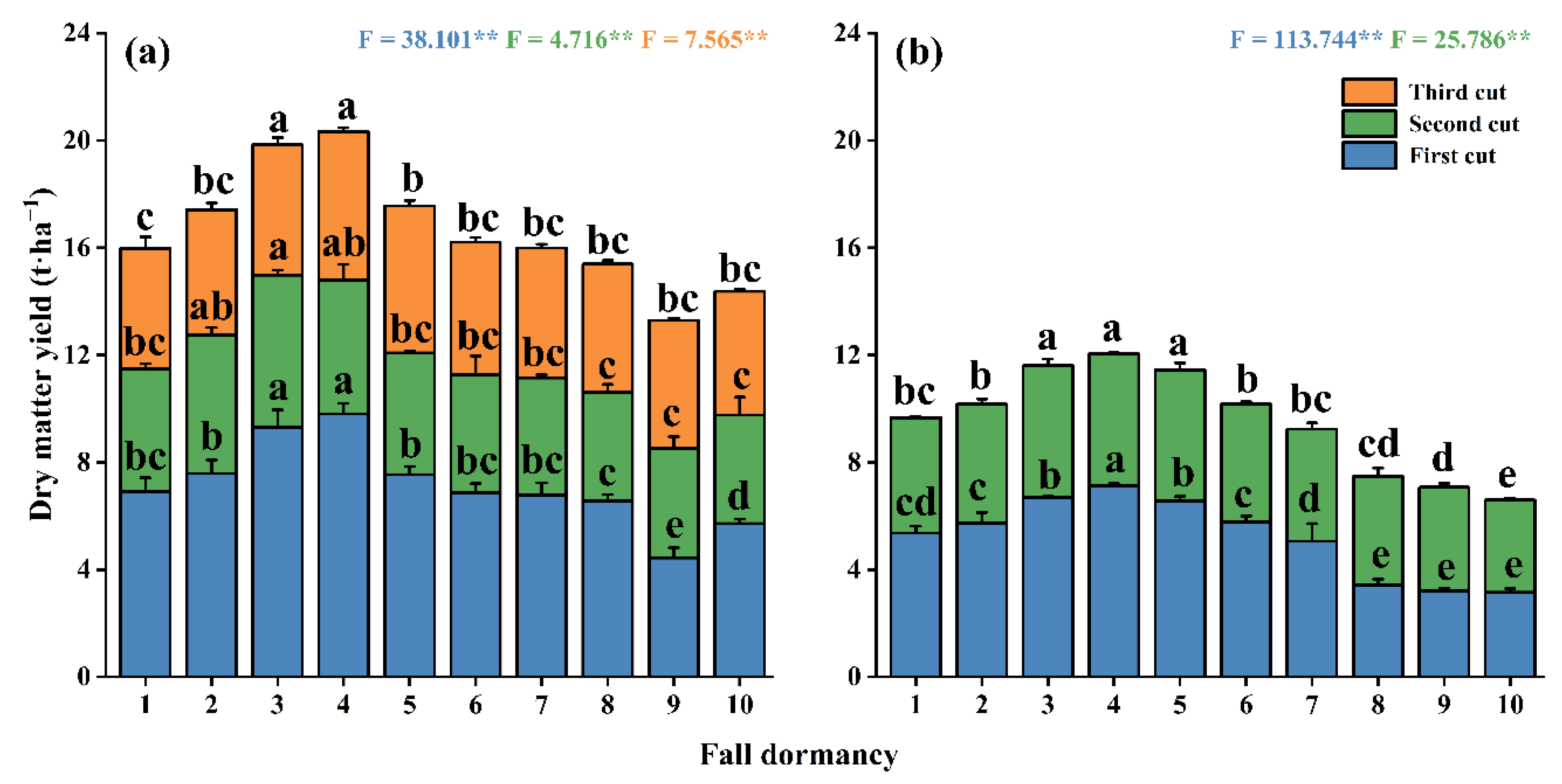
3.6. Comparison of Dry Matter Yield of Alfalfa with Different Fall-Dormancy Levels
3.7. Relationship among C, N, and P, Elemental Reabsorption, and Dry Matter Yield in Mature Leaves
4. Discussion
4.1. Effect of Fall-Dormancy Level on Stoichiometric Characteristics of Alfalfa
4.2. Effect of Fall-Dormancy Level of Alfalfa on Nutrient Reabsorption Characteristics of Leaves
4.3. Effect of Fall-Dormancy Level on Persistence and Yield of Alfalfa
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tang, Z.Y.; Xu, W.T.; Zhou, G.Y.; Bai, Y.F.; Li, J.X.; Tang, X.L.; Chen, D.M.; Liu, Q.; Ma, W.H.; Xiong, G.M.; et al. Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems. Proc. Natl. Acad. Sci. USA 2018, 115, 4033–4038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.J.; Zeng, D.H.; Mao, R.; Yu, Z.Y. Nitrogen and phosphorus resorption of artemisia scoparia, chenopodium acuminatum, cannabis sativa, and phragmites communis under nitrogen and phosphorus additions in a semiarid grassland. Plant Soil Environ. 2012, 58, 446–451. [Google Scholar] [CrossRef] [Green Version]
- Tian, D.; Yan, Z.B.; Niklas, K.; Han, W.X.; Kattge, J.; Reich, P.; Luo, Y.K.; Chen, Y.H.; Tang, Z.Y.; Hu, H.F.; et al. Global leaf nitrogen and phosphorus stoichiometry and their scaling exponent. Natl. Sci. Rev. 2018, 5, 728–739. [Google Scholar] [CrossRef] [Green Version]
- Ågren, G.I.; Wetterstedt, M.; Billberger, M. Nutrient limitation on terrestrial plant growth-modeling the interaction between nitrogen and phosphorus. New Phytol. 2012, 194, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Ågren, G.I. Stoichiometry and nutrition of plant growth in natural communities. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 153–170. [Google Scholar] [CrossRef]
- Wang, C.Z.; Ma, B.; Yan, X.B.; Han, J.F.; Guo, Y.X.; Wang, Y.H.; Li, P. Yields of alfalfa varieties with different fall-dormancy levels in a temperate environment. Agron. J. 2009, 101, 1146–1152. [Google Scholar] [CrossRef]
- Leep, R.H.; Andresen, J.A.; Jeranyama, P. Fall dormancy and snow depth effects on winterkill of alfalfa. Agron. J. 2001, 93, 1142–1148. [Google Scholar] [CrossRef]
- Ventroni, L.M.; Volenec, J.J.; Cangiano, C.A. Fall dormancy and cutting frequency impact on alfalfa yield and yield components. Field Crops Res. 2010, 119, 252–259. [Google Scholar] [CrossRef]
- Kobe, R.K.; Lepczyk, C.A.; Iyer, M. Resorption efficiency decreases with increasing green leaf nutrients in a global data set. Ecol. Monogr. 2005, 86, 2780–2792. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.N.; Lu, J.Y.; Yang, M.; Yang, H.M.; Zhang, Q.P. Stoichiometric characteristics of carbon, citrogen, and phosphorus in leaves of differently aged lucerne (Medicago sativa) stands. Front. Plant Sci. 2015, 6, 1062. [Google Scholar] [CrossRef]
- Vergutz, L.; Manzoni, S.; Porporato, A.; Novais, R.F.; Jackson, R.B. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol. Monogr. 2012, 82, 205–220. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.B.; Kim, N.; Han, W.X.; Guo, Y.L.; Han, T.S.; Du, E.Z.; Fang, J.Y. Effects of nitrogen and phosphorus supply on growth rate, leaf stoichiometry, and nutrient resorption of Arabidopsis thaliana. Plant Soil 2014, 388, 147–155. [Google Scholar] [CrossRef]
- Brant, A.N.; Chen, H.Y. Patterns and mechanisms of nutrient resorption in plants. Crit. Rev. Plant Sci. 2015, 34, 471–486. [Google Scholar] [CrossRef]
- Van, H.L.M.; Toet, S.; Aerts, R. Nitrogen and phosphorus resorption efficiency and proficiency in six sub-arctic bog species after 4 years of nitrogen fertilization. J. Ecol. 2003, 91, 1060–1070. [Google Scholar]
- Yuan, Z.Y.; Li, L.H.; Han, X.G.; Huang, J.H.; Jiang, G.M.; Wan, S.; Zhang, W.H.; Chen, Q.S. Nitrogen resorption from senescing leaves in 28 plant species in a semi-arid region of northern China. J. Arid Environ. 2005, 63, 191–202. [Google Scholar] [CrossRef]
- Lü, X.T.; Han, X.G. Nutrient resorption responses to water and nitrogen amendment in semi-arid grassland of Inner Mongolia, China. Plant Soil 2010, 327, 481–491. [Google Scholar] [CrossRef]
- Lu, J.Y.; Yang, M.; Liu, M.G.; Wang, Y.Y.; Yang, H.M. Leaf stoichiometry and resorption of N and P in Lucerne at different growth stages under different water supplies. J. Plant Nutr. 2019, 42, 501–511. [Google Scholar] [CrossRef]
- Fang, S.S. Study on Fall Dormancy and Production Performance of Different Alfalfa Cultivars. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2015. Part 3–6 (In Chinese). [Google Scholar]
- Sun, Q.Z.; Wang, Z.L.; Xun, L.J. Alfalfa in Arid Regions; China Social Sciences Press: Beijing, China, 2014; pp. 58–79. (In Chinese) [Google Scholar]
- Rimi, F.; Macolino, S.; Leinauer, B.; Lauriault, L.; Ziliotto, U. Fall dormancy and harvest stage effects on alfalfa nutritive value in a subtropical climate. Agron. J. 2012, 104, 415. [Google Scholar] [CrossRef]
- Yang, M.; Lu, J.Y.; Liu, M.G.; Lu, Y.X.; Yang, H.M. Leaf nutrient resorption in lucerne decreases with relief of relative soil nutrient limitation under phosphorus and potassium fertilization with irrigation. Sci. Rep. 2020, 10, 10525. [Google Scholar] [CrossRef]
- Koerselman, W.; Meuleman, A.F.M. The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation. J. Appl. Ecol. 1996, 33, 1441–1450. [Google Scholar] [CrossRef]
- XIE, K.Y.; LI, X.L.; Feng, H.E.; ZHANG, Y.J.; WAN, L.Q.; David, B.H.; Gamal, M.F. Effect of nitrogen fertilization on yield, N content, and nitrogen fixation of alfalfa and smooth bromegrass grown alone or in mixture in greenhouse pots. J. Integr. Agric. 2015, 14, 1864–1876. [Google Scholar] [CrossRef]
- Lu, J.Y. Effect of N and P Fertilizations on Leaf Nutrient Resorption of Alfalfa at Different Growth Stages in the Loess Plateau. Ph.D. Thesis, Lanzhou University, Lanzhou, China, 2019. Part 1–5 (In Chinese). [Google Scholar]
- Lu, J.Y.; Liu, M.G.; Yang, M.; Xie, J.H.; Yang, H.M.; Li, L.L. Leaf resorption and stoichiometry of N and P of 1, 2 and 3 year-old lucerne under one-time P fertilization. Soil Tillage Res. 2020, 197, 104481. [Google Scholar] [CrossRef]
- Yamori, W.; Masumoto, C.; Fukayama, H.; Makino, A. Rubisco activase is a key regulator of non-steady-state photosynthesis at any leaf temperature and, to a lesser extent, of steady-state photosynthesis at high temperature. Plant J. 2012, 71, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Hikosaka, K.; Ishikawa, K.; Borjigidai, A.; Muller, O.; Onoda, Y. Temperature acclimation of photosynthesis: Mechanisms involved in the changes in temperature dependence of photosynthetic rate. J. Exp. Bot. 2006, 57, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Haldimann, P.; Feller, U. Growth at moderately elevated temperature alters the physiological response of the photosynthetic apparatus to heat stress in pea (Pisum sativum L.) leaves. Plant Cell Environ. 2005, 28, 302–317. [Google Scholar] [CrossRef]
- Lamm, F.R.; Harmoney, K.R.; Aboukheira, A.A.; Johnson, S.K. Alfalfa production with subsurface drip irrigation in the Central Great Plains. Trans. ASABE 2012, 55, 1203–1212. [Google Scholar] [CrossRef]
- Malinowski, D.; Pinchak, W.; Kramp, B.; Zuo, H.; Butler, T. Supplemental irrigation and fall dormancy effects on alfalfa productivity in a semiarid, subtropical climate with a bimodal precipitation pattern. Agron. J. 2007, 99, 621–629. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, X.; Ma, C.; Zhang, Q. Effects of Nitrogen and Phosphorus Addition on Agronomic Characters, Photosynthetic Performance and Anatomical Structure of Alfalfa in Northern Xinjiang, China. Agronomy 2022, 12, 1613. [Google Scholar] [CrossRef]
- Haagenson, D.; Cunningham, S.; Volenec, J. Root physiology of less fall dormant, winter hardy alfalfa selections. Crop Sci. 2003, 43, 1441–1447. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.N.; Lu, J.Y.; Yang, H.M.; Zhang, X.; Luo, C.L.; Zhao, Y.X. Resorption of nitrogen, phosphorus and potassium from leaves of lucerne stands of different ages. Plant Soil 2014, 383, 301–312. [Google Scholar] [CrossRef]
- Rimi, F.; Macolino, S.; Leinauer, B.; Lauriault, L.; Ziliotto, U. Fall dormancy and harvest stage impact on alfalfa persistence in a subtropical climate. Agron. J. 2014, 106, 1258–1266. [Google Scholar] [CrossRef]
- Bélanger, G.; Kunelius, T.; McKenzie, D.; Papadopoulos, Y.; Thomas, B.; McRae, K.; Fillmore, S.A.E.; Christie, B. Fall cutting management affects yield and persistence of alfalfa in Atlantic Canada. Can. J. Plant Sci. 1999, 79, 57–63. [Google Scholar] [CrossRef]
- Lawati, A.A.; Pierce, C.; Murray, L.; Ray, I. Combining ability and heterosis for forage yield among elite alfalfa core collection accessions with different fall dormancy responses. Crop Sci. 2010, 50, 150–158. [Google Scholar] [CrossRef]
- Cunningham, S.M.; Gana, J.A.; Volenec, J.J. Winter hardiness, root physiology, and gene expression in successive fall dormancy selections. Crop Sci. 2001, 41, 1091–1098. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Si, D.; Sun, X.W.; Mu, L.; Zhao, H.K.; Li, Z.C.; Wei, Z.S. Effect of cutting under high temperature on regrowth and quality of five alfalfa cultivars. Pratacult. Sci. 2017, 34, 1469–1474. (In Chinese) [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Fall Dormancy Rate | Type Fall Dormancy | Source |
|---|---|---|---|
| Xinmu NO.1 | level 1 | Extreme fall-dormancy type | Institute of Grassland Research of CAAS |
| Gongnong NO.1 | level 1 | Extreme fall-dormancy type | |
| Zhaodong | level 1 | Extreme fall-dormancy type | |
| Aohan | level 2 | Fall-dormancy type | |
| Xinmu NO.2 | level 2 | Fall-dormancy type | |
| Zhongmu NO.1 | level 2 | Fall-dormancy type | |
| Zhongmu NO.2 | level 3 | Fall-dormancy type | |
| Concept | level 3 | Fall-dormancy type | |
| Zhongmu NO.3 | level 3 | Fall-dormancy type | |
| Adrenalim | level 4 | Moderate fall-dormancy type | |
| Xinjiang Daye | level 4 | Moderate fall-dormancy type | |
| Victoria | level 6 | Moderate fall-dormancy type | |
| Ghillie | level 6 | Moderate fall-dormancy type | |
| Yumu NO.1 | level 7 | Non-fall-dormancy type | |
| Gannong NO.5 | level 8 | Non-fall-dormancy type | |
| Liangmu NO.1 | level 8 | Non-fall-dormancy type | |
| WL325HQ | level 4 | Moderate fall-dormancy type | Beijing Zhengdao Ecological Technology Co. |
| WL363HQ | level 5 | Moderate fall-dormancy type | |
| WL366HQ | level 5 | Moderate fall-dormancy type | |
| WL525HQ | level 8 | Non-fall-dormancy type | |
| WL903HQ | level 9 | Extreme non-fall-dormancy type | |
| WL656HQ | level 9 | Extreme non-fall-dormancy type | |
| WL712HQ | level 10 | Extreme non-fall-dormancy type | |
| Paola | level 5 | Moderate fall-dormancy type | Tianjin Bailv International Grass Industry Co. |
| Sandili | level 6 | Moderate fall-dormancy type | |
| Sardi7 | level 7 | Non-fall-dormancy type | |
| Blue moon | level 7 | Non-fall-dormancy type | |
| Pegasis | level 9 | Extreme non-fall-dormancy type | |
| Sardi10 | level 10 | Extreme non-fall-dormancy type | |
| UC1887 | level 10 | Extreme non-fall-dormancy type |
| Index | Year | Cut | CV (%) | Linear Model | Nonlinear Model | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Y = ax + b | R2 | F | P | Y = ax2 + bx + c | R2 | F | P | ||||
| Green-leaf C | 2020 | 1 | 1.78 | y = −0.884x + 409.460 | 0.139 | 1.286 | 0.290 | y = −0.777x2 + 7.668x + 392.360 | 0.8258 | 16.586 | 0.002 |
| 2 | 1.79 | y = −1.070x + 372.150 | 0.244 | 2.574 | 0.147 | y = −0.624x2 + 5.795x + 358.430 | 0.774 | 11.991 | 0.006 | ||
| 3 | 1.48 | y = −0.664x + 359.300 | 0.146 | 1.368 | 0.276 | y = −0.548x2 + 5.361x + 347.250 | 0.783 | 12.593 | 0.005 | ||
| 2021 | 1 | 1.97 | y = −1.726x + 401.510 | 0.459 | 6.792 | 0.031 | y = −0.603x2 + 4.907x + 388.250 | 0.818 | 15.745 | 0.003 | |
| 2 | 1.37 | y = −1.091x + 374.190 | 0.426 | 5.949 | 0.041 | y = −0.378x2 + 3.062x + 365.880 | 0.753 | 10.693 | 0.007 | ||
| Green-leaf N | 2020 | 1 | 3.67 | y = 0.076x + 35.205 | 0.031 | 0.254 | 0.628 | y = 0.148x2 − 1.548x + 38.452 | 0.778 | 12.239 | 0.005 |
| 2 | 2.42 | y = 0.093x + 37.320 | 0.094 | 0.826 | 0.390 | y = 0.098x2 − 0.981x + 39.467 | 0.758 | 10.921 | 0.007 | ||
| 3 | 1.56 | y = 0.152x + 39.411 | 0.533 | 9.141 | 0.017 | y = 0.047x2 − 0.360x + 40.434 | 0.855 | 20.591 | <0.001 | ||
| 2021 | 1 | 2.66 | y = 0.227x + 35.496 | 0.495 | 7.808 | 0.023 | y = 0.066x2 − 0.502x + 36.954 | 0.765 | 11.387 | 0.006 | |
| 2 | 1.45 | y = 0.158x + 39.315 | 0.676 | 16.789 | 0.003 | y = 0.028x2 − 0.154x + 39.939 | 0.816 | 15.627 | 0.003 | ||
| Green-leaf P | 2020 | 1 | 4.90 | y = 0.011x + 2.504 | 0.076 | 0.611 | 0.457 | y = 0.013x2 − 0.134x + 2.796 | 0.727 | 9.296 | 0.011 |
| 2 | 4.02 | y = 0.008x + 2.608 | 0.025 | 1.219 | 0.302 | y = 0.015x2 − 0.159x + 2.943 | 0.539 | 12.761 | 0.005 | ||
| 3 | 1.90 | y = 0.013x + 2.832 | 0.493 | 7.766 | 0.034 | y = 0.004x2 − 0.035x + 2.927 | 0.853 | 20.335 | <0.001 | ||
| 2021 | 1 | 4.04 | y = 0.022x + 2.496 | 0.380 | 5.008 | 0.056 | y = 0.008x2 − 0.066x + 2.671 | 0.713 | 8.879 | 0.012 | |
| 2 | 1.88 | y = 0.014x + 2.827 | 0.616 | 13.986 | 0.006 | y = 0.003x2 − 0.021x + 2.898 | 0.820 | 17.086 | 0.002 | ||
| Index | Year | Cut | CV (%) | Linear Model | Non-Linear Model | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| y = ax + b | R2 | F | P | y = ax2 + bx + c | R2 | F | P | ||||
| Senesced- leaf C | 2020 | 1 | 2.39 | y = −1.160x + 378.290 | 0.157 | 1.487 | 0.257 | y = −0.930x2 + 9.065x + 357.840 | 0.801 | 14.047 | 0.004 |
| 2 | 2.48 | y = −1.241x + 369.160 | 0.175 | 1.694 | 0.229 | y = −0.914x2 + 8.807x + 349.060 | 0.781 | 12.490 | 0.005 | ||
| 3 | 1.59 | y = −0.676x + 374.300 | 0.121 | 1.103 | 0.324 | y = −0.634x2 + 6.2947x + 360.360 | 0.802 | 14.197 | 0.003 | ||
| 2021 | 1 | 2.65 | y = −2.257x + 392.850 | 0.461 | 6.840 | 0.031 | y = −0.782x2 + 6.3417x + 375.650 | 0.815 | 15.402 | 0.003 | |
| 2 | 2.00 | y = −1.784x + 382.920 | 0.521 | 8.718 | 0.018 | y = −0.499x2 + 3.704x + 371.950 | 0.782 | 12.595 | 0.005 | ||
| Senesced- leaf N | 2020 | 1 | 7.43 | y = 0.187x + 17.375 | 0.171 | 1.661 | 0.233 | y = 0.139x2 − 1.341x + 20.431 | 0.776 | 12.132 | 0.005 |
| 2 | 5.43 | y = 0.141x + 19.509 | 0.149 | 1.406 | 0.270 | y = 0.123x2 − 1.212x + 22.214 | 0.880 | 25.511 | <0.001 | ||
| 3 | 3.08 | y = 0.162x + 21.651 | 0.499 | 7.957 | 0.023 | y = 0.052x2 − 0.413x + 22.802 | 0.831 | 17.261 | 0.002 | ||
| 2021 | 1 | 5.95 | y = 0.266x + 17.933 | 0.485 | 7.540 | 0.025 | y = 0.084x2 − 0.658x + 19.780 | 0.796 | 13.632 | 0.004 | |
| 2 | 3.01 | y = 0.170x + 21.772 | 0.570 | 10.533 | 0.012 | y = 0.046x2 − 0.339x + 22.790 | 0.839 | 18.181 | 0.002 | ||
| Senesced- leaf P | 2020 | 1 | 7.72 | y = 0.007x + 1.080 | 0.052 | 0.442 | 0.525 | y = 0.009x2 − 0.094x + 1.280 | 0.706 | 8.401 | 0.014 |
| 2 | 6.40 | y = 0.015x + 1.575 | 0.182 | 1.780 | 0.219 | y = 0.011x2 − 0.105x + 1.816 | 0.807 | 14.596 | 0.003 | ||
| 3 | 2.46 | y = 0.011x + 1.833 | 0.467 | 6.996 | 0.030 | y = 0.004x2 − 0.032x + 1.919 | 0.876 | 24.819 | <0.001 | ||
| 2021 | 1 | 5.57 | y = 0.012x + 1.052 | 0.360 | 4.505 | 0.067 | y = 0.005x2 − 0.041x + 1.158 | 0.712 | 8.646 | 0.013 | |
| 2 | 2.67 | y = 0.013x + 1.845 | 0.570 | 10.605 | 0.012 | y = 0.003x2 − 0.023x + 1.916 | 0.804 | 14.347 | 0.003 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Wang, X.; Ma, C.; Zhang, Q. Characteristics of Carbon, Nitrogen and Phosphorus Stoichiometry and Nutrient Reabsorption in Alfalfa Leaves with Different Fall-Dormancy Levels in Northern Xinjiang, China. Agriculture 2022, 12, 2154. https://doi.org/10.3390/agriculture12122154
Sun Y, Wang X, Ma C, Zhang Q. Characteristics of Carbon, Nitrogen and Phosphorus Stoichiometry and Nutrient Reabsorption in Alfalfa Leaves with Different Fall-Dormancy Levels in Northern Xinjiang, China. Agriculture. 2022; 12(12):2154. https://doi.org/10.3390/agriculture12122154
Chicago/Turabian StyleSun, Yanliang, Xuzhe Wang, Chunhui Ma, and Qianbing Zhang. 2022. "Characteristics of Carbon, Nitrogen and Phosphorus Stoichiometry and Nutrient Reabsorption in Alfalfa Leaves with Different Fall-Dormancy Levels in Northern Xinjiang, China" Agriculture 12, no. 12: 2154. https://doi.org/10.3390/agriculture12122154
APA StyleSun, Y., Wang, X., Ma, C., & Zhang, Q. (2022). Characteristics of Carbon, Nitrogen and Phosphorus Stoichiometry and Nutrient Reabsorption in Alfalfa Leaves with Different Fall-Dormancy Levels in Northern Xinjiang, China. Agriculture, 12(12), 2154. https://doi.org/10.3390/agriculture12122154