Pavlovian-To-Instrumental Transfer and Alcohol Consumption in Young Male Social Drinkers: Behavioral, Neural and Polygenic Correlates

 , , , and
, , , and 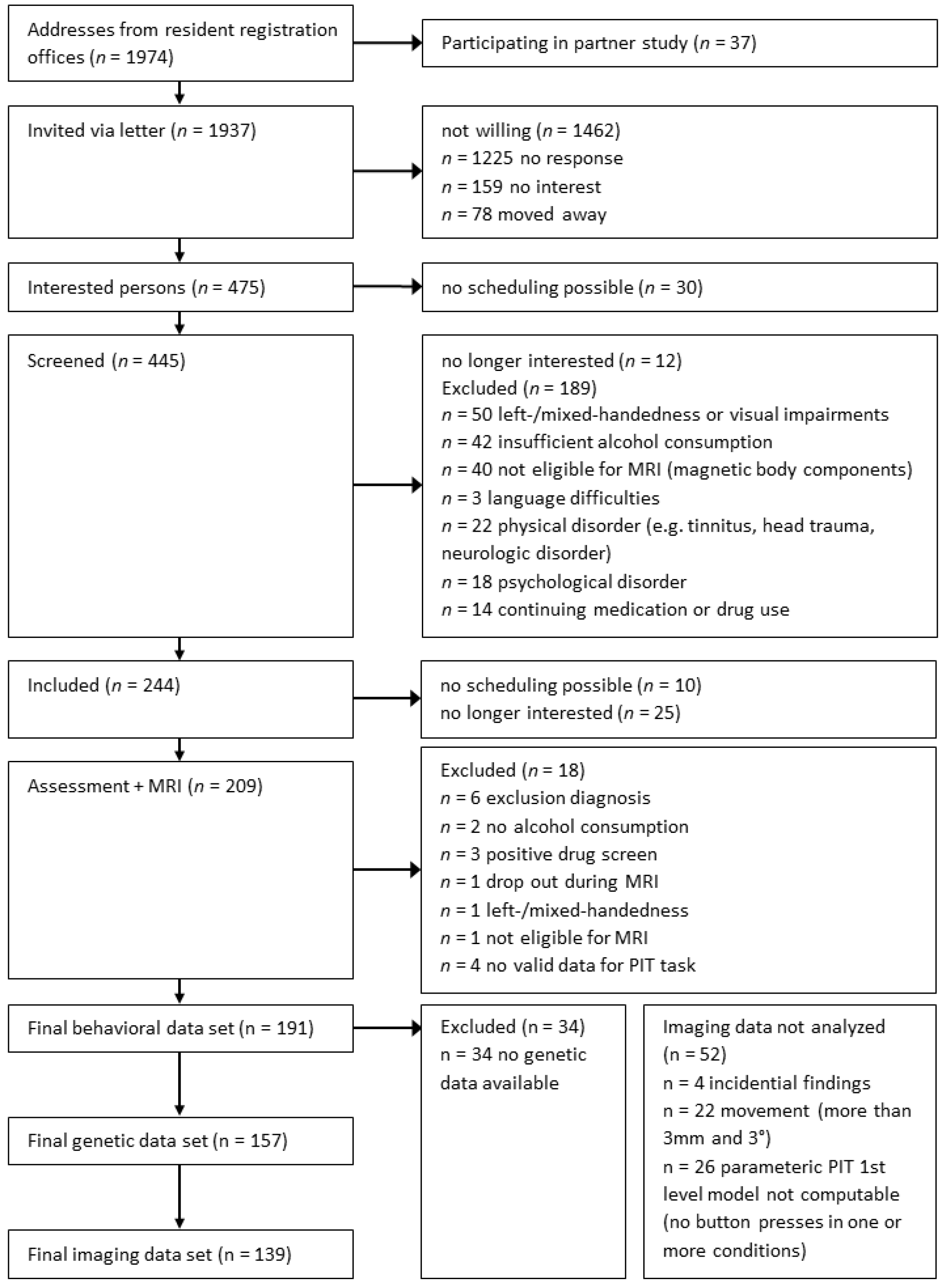{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants and Procedure
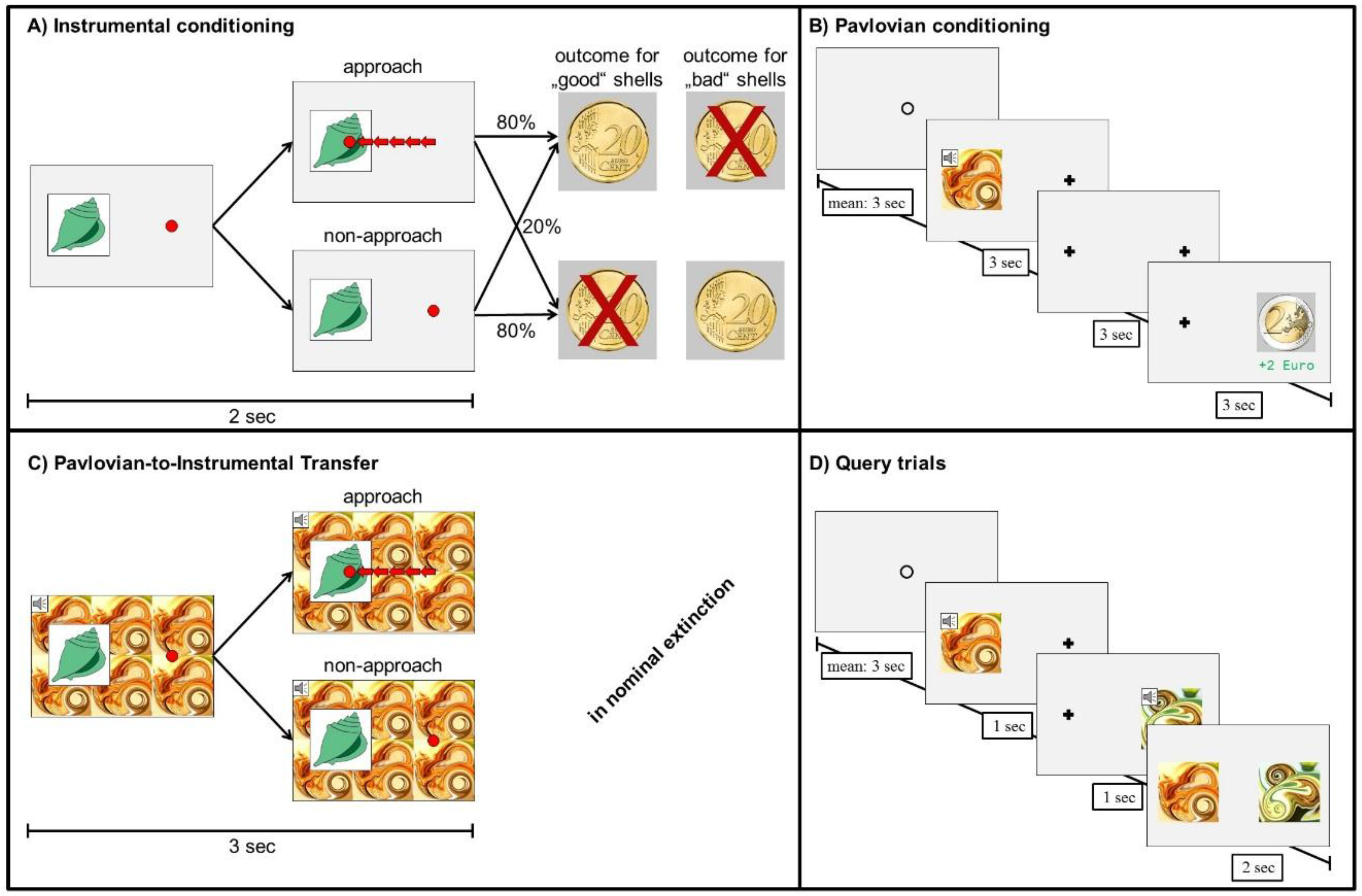
2.2. Experimental Design
2.3. Self-Reported Questionnaires
2.4. Measures of Alcohol Consumption
2.5. MRI Data Acquisition
2.6. Polygenic Risk Score
2.7. Statistical Analysis
2.8. Behavioral Analysis
2.9. Imaging Analysis
2.10. Polygenic Analyses
3. Results
3.1. Sample Characteristics by Drinking Group
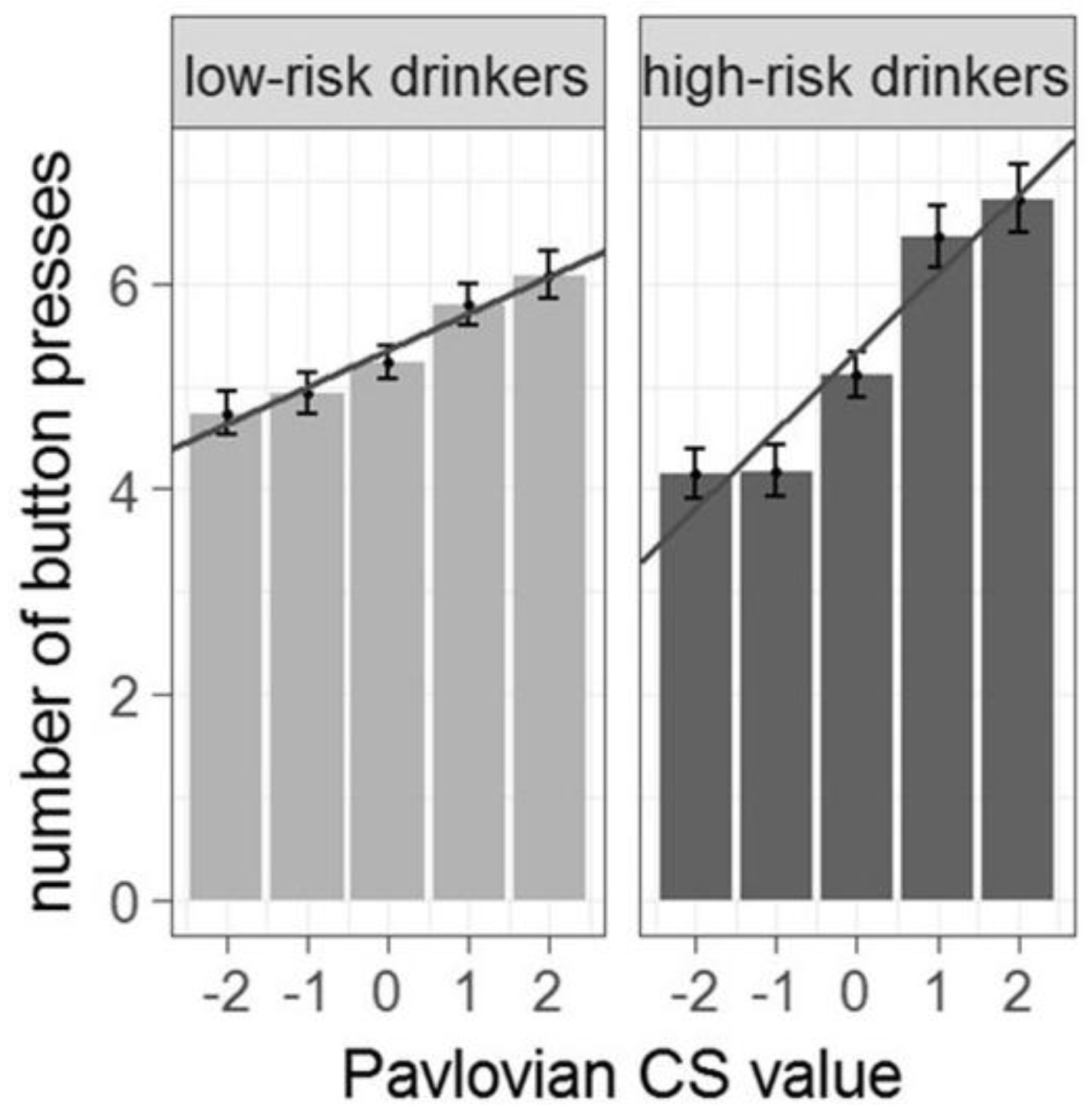
3.2. Behavioral Results
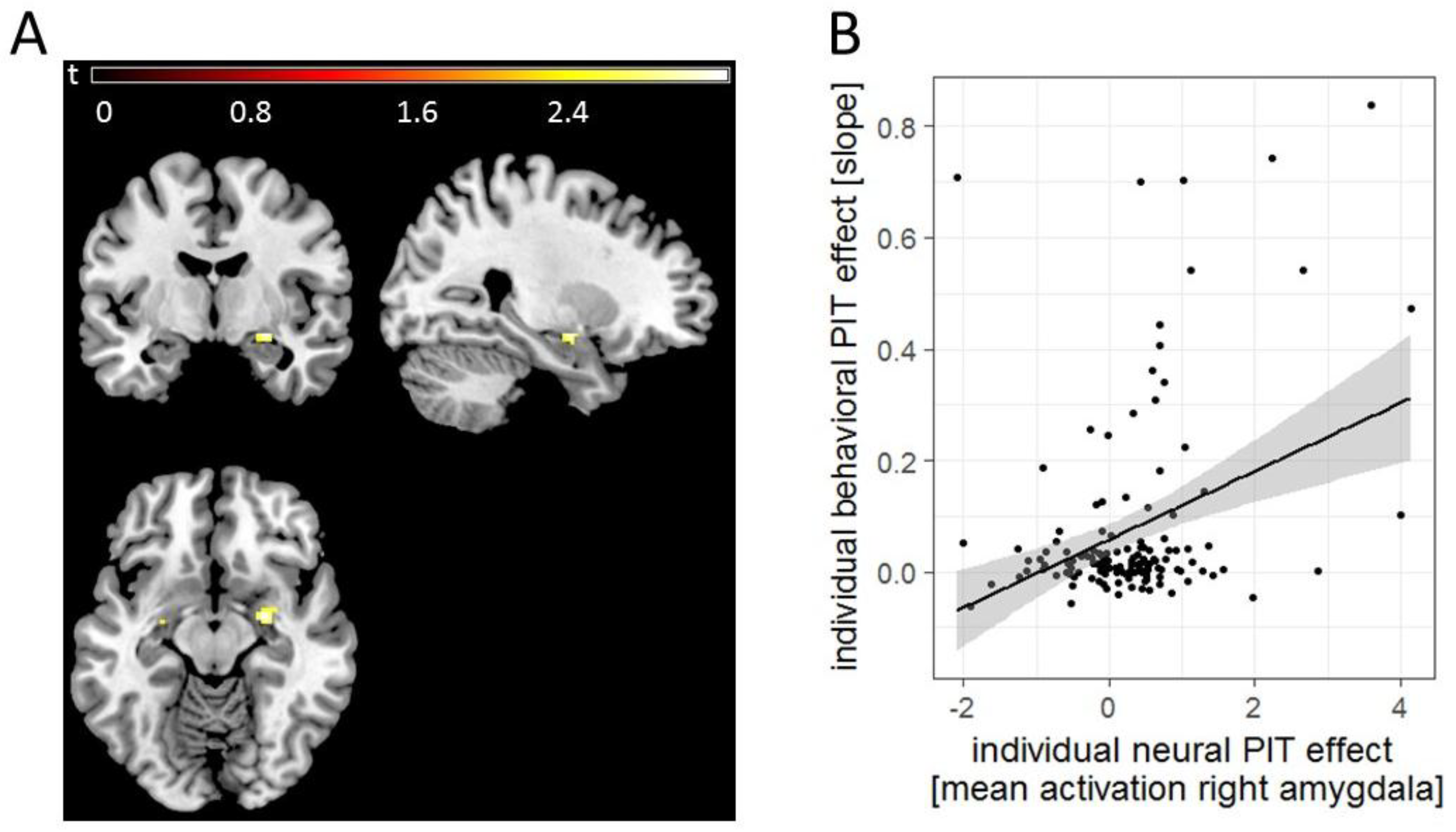
3.3. Imaging Results
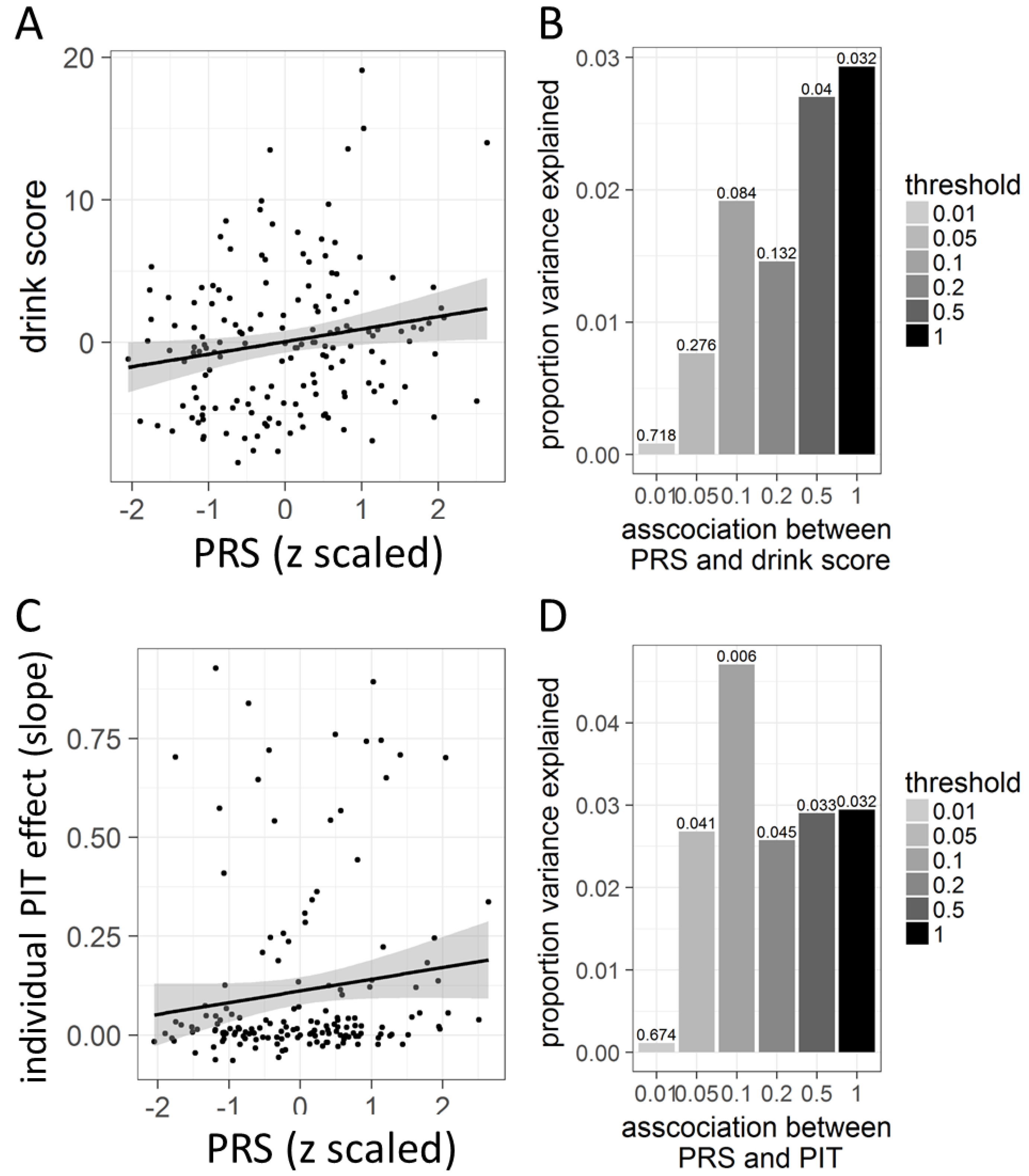
3.4. Polygenic Risk in Association with Behavioral PIT
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maggs, J.L.; Schulenberg, J.E. Initiation and course of alcohol consumption among adolescents and young adults. In Recent Developments in Alcoholism: An Official Publication of the American Medical Society on Alcoholism, the Research Society on Alcoholism, and the National Council on Alcoholism; National Institute on Alcohol Abuse and Alcoholism: Bethesda, MD, USA, 2005; Volume 17, pp. 29–47. [Google Scholar]
- Robbins, T.W.; Everitt, B.J. Drug addiction: Bad habits add up. Nature 1999, 398, 567–570. [Google Scholar] [CrossRef] [PubMed]
- Everitt, B.J.; Robbins, T.W. Neural systems of reinforcement for drug addiction: From actions to habits to compulsion. Nat. Neurosci. 2005, 8, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Everitt, B.J.; Robbins, T.W. Drug addiction: Updating actions to habits to compulsions ten years on. Annu. Rev. Psychol. 2016, 67, 23–50. [Google Scholar] [CrossRef] [PubMed]
- Garbusow, M.; Schad, D.J.; Sebold, M.; Friedel, E.; Bernhardt, N.; Koch, S.P.; Steinacher, B.; Kathmann, N.; Geurts, D.E.; Sommer, C.; et al. Pavlovian-to-instrumental transfer effects in the nucleus accumbens relate to relapse in alcohol dependence. Addict. Biol. 2016, 21, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Sommer, C.; Garbusow, M.; Jünger, E.; Pooseh, S.; Bernhardt, N.; Birkenstock, J.; Schad, D.J.; Jabs, B.; Glockler, T.; Huys, Q.M.; et al. Strong seduction: Impulsivity and the impact of contextual cues on instrumental behavior in alcohol dependence. Transl. Psychiatry 2017, 7, e1183. [Google Scholar] [CrossRef] [PubMed]
- Victorio-Estrada, A.; Mucha, R.F. The inventory of drinking situations (ids) in current drinkers with different degrees of alcohol problems. Addict. Behav. 1997, 22, 557–565. [Google Scholar] [CrossRef]
- Heinz, A.J.; Beck, A.; Meyer-Lindenberg, A.; Sterzer, P.; Heinz, A. Cognitive and neurobiological mechanisms of alcohol-related aggression. Nat. Rev. Neurosci. 2011, 12, 400–413. [Google Scholar] [CrossRef] [PubMed]
- Huys, Q.J.; Cools, R.; Golzer, M.; Friedel, E.; Heinz, A.; Dolan, R.J.; Dayan, P. Disentangling the roles of approach, activation and valence in instrumental and pavlovian responding. PLoS Comput. Biol. 2011, 7, e1002028. [Google Scholar] [CrossRef] [PubMed]
- Corbit, L.H.; Balleine, B.W. Double dissociation of basolateral and central amygdala lesions on the general and outcome-specific forms of pavlovian-instrumental transfer. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 962–970. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, K.H.; Maidment, N.T.; Ostlund, S.B. Repeated cocaine exposure facilitates the expression of incentive motivation and induces habitual control in rats. PLoS ONE 2013, 8, e61355. [Google Scholar] [CrossRef] [PubMed]
- Corbit, L.H.; Janak, P.H. Ethanol-associated cues produce general pavlovian-instrumental transfer. Alcohol. Clin. Exp. Res. 2007, 31, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Barker, J.M.; Torregrossa, M.M.; Taylor, J.R. Low prefrontal psa-ncam confers risk for alcoholism-related behavior. Nat. Neurosci. 2012, 15, 1356–1358. [Google Scholar] [CrossRef] [PubMed]
- Hogarth, L.; Retzler, C.; Munafo, M.R.; Tran, D.M.; Troisi, J.R., 2nd; Rose, A.K.; Jones, A.; Field, M. Extinction of cue-evoked drug-seeking relies on degrading hierarchical instrumental expectancies. Behav. Res. Ther. 2014, 59, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, L.; Mitchell, C.; Seabrooke, T.; Hogarth, L. Drug cue reactivity involves hierarchical instrumental learning: Evidence from a biconditional pavlovian to instrumental transfer task. Psychopharmacology 2017, 234, 1977–1984. [Google Scholar] [CrossRef] [PubMed]
- Martinovic, J.; Jones, A.; Christiansen, P.; Rose, A.K.; Hogarth, L.; Field, M. Electrophysiological responses to alcohol cues are not associated with pavlovian-to-instrumental transfer in social drinkers. PLoS ONE 2014, 9, e94605. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization; Department of Mental Health and Substance DependenceNoncommunicable Diseases. International Guide for MONITORING Alcohol Consumption and Related Harm. Technical Document who/msd/msb/00.4; World Health Organisation: Geneva, Switzerland, 2000. [Google Scholar]
- Campese, V.D.; Kim, J.; Lazaro-Munoz, G.; Pena, L.; LeDoux, J.E.; Cain, C.K. Lesions of lateral or central amygdala abolish aversive pavlovian-to-instrumental transfer in rats. Front. Behav. Neurosci. 2014, 8, 161. [Google Scholar] [CrossRef] [PubMed]
- Holland, P.C.; Hsu, M. Role of amygdala central nucleus in the potentiation of consuming and instrumental lever-pressing for sucrose by cues for the presentation or interruption of sucrose delivery in rats. Behav. Neurosci. 2014, 128, 71–82. [Google Scholar] [CrossRef]
- McCue, M.G.; LeDoux, J.E.; Cain, C.K. Medial amygdala lesions selectively block aversive pavlovian-instrumental transfer in rats. Front. Behav. Neurosci. 2014, 8, 329. [Google Scholar] [CrossRef]
- Wassum, K.M.; Ostlund, S.B.; Loewinger, G.C.; Maidment, N.T. Phasic mesolimbic dopamine release tracks reward seeking during expression of pavlovian-to-instrumental transfer. Biol. Psychiatry 2013, 73, 747–755. [Google Scholar] [CrossRef]
- Di Chiara, G.; Bassareo, V. Reward system and addiction: What dopamine does and doesn’t do. Curr. Opin. Pharmacol. 2007, 7, 69–76. [Google Scholar] [CrossRef]
- Deserno, L.; Beck, A.; Huys, Q.J.; Lorenz, R.C.; Buchert, R.; Buchholz, H.G.; Plotkin, M.; Kumakara, Y.; Cumming, P.; Heinze, H.J.; et al. Chronic alcohol intake abolishes the relationship between dopamine synthesis capacity and learning signals in the ventral striatum. Eur. J. Neurosci. 2015, 41, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wang, G.J.; Fowler, J.S.; Logan, J.; Hitzemann, R.; Ding, Y.S.; Pappas, N.; Shea, C.; Piscani, K. Decreases in dopamine receptors but not in dopamine transporters in alcoholics. Alcohol. Clin. Exp. Res. 1996, 20, 1594–1598. [Google Scholar] [CrossRef]
- Talmi, D.; Seymour, B.; Dayan, P.; Dolan, R.J. Human pavlovian-instrumental transfer. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Prevost, C.; Liljeholm, M.; Tyszka, J.M.; O’Doherty, J.P. Neural correlates of specific and general pavlovian-to-instrumental transfer within human amygdalar subregions: A high-resolution fmri study. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 8383–8390. [Google Scholar] [CrossRef] [PubMed]
- Geurts, D.E.; Huys, Q.J.; den Ouden, H.E.; Cools, R. Aversive pavlovian control of instrumental behavior in humans. J. Cogn. Neurosci. 2013, 25, 1428–1441. [Google Scholar] [CrossRef] [PubMed]
- Wiers, C.E.; Stelzel, C.; Park, S.Q.; Gawron, C.K.; Ludwig, V.U.; Gutwinski, S.; Heinz, A.; Lindenmeyer, J.; Wiers, R.W.; Walter, H.; et al. Neural correlates of alcohol-approach bias in alcohol addiction: The spirit is willing but the flesh is weak for spirits. Neuropsychopharmacology 2014, 39, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Manolio, T.A.; Collins, F.S.; Cox, N.J.; Goldstein, D.B.; Hindorff, L.A.; Hunter, D.J.; McCarthy, M.I.; Ramos, E.M.; Cardon, L.R.; Chakravarti, A.; et al. Finding the missing heritability of complex diseases. Nature 2009, 461, 747–753. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.; Simpkin, A.J.; Haycock, P.C.; Dudbridge, F.; Zuccolo, L. Exploration of a polygenic risk score for alcohol consumption: A longitudinal analysis from the alspac cohort. PLoS ONE 2016, 11, e0167360. [Google Scholar] [CrossRef]
- Kapoor, M.; Chou, Y.L.; Edenberg, H.J.; Foroud, T.; Martin, N.G.; Madden, P.A.; Wang, J.C.; Bertelsen, S.; Wetherill, L.; Brooks, A.; et al. Genome-wide polygenic scores for age at onset of alcohol dependence and association with alcohol-related measures. Transl. Psychiatry 2016, 6, e761. [Google Scholar] [CrossRef]
- Li, J.J.; Cho, S.B.; Salvatore, J.E.; Edenberg, H.J.; Agrawal, A.; Chorlian, D.B.; Porjesz, B.; Hesselbrock, V.; Dick, D.M. The impact of peer substance use and polygenic risk on trajectories of heavy episodic drinking across adolescence and emerging adulthood. Alcohol. Clin. Exp. Res. 2017, 41, 65–75. [Google Scholar] [CrossRef]
- Schumann, G.; Liu, C.; O’Reilly, P.; Gao, H.; Song, P.; Xu, B.; Ruggeri, B.; Amin, N.; Jia, T.; Preis, S.; et al. Klb is associated with alcohol drinking, and its gene product beta-klotho is necessary for fgf21 regulation of alcohol preference. Proc. Natl. Acad. Sci. USA 2016, 113, 14372–14377. [Google Scholar] [CrossRef] [PubMed]
- Livingston, M.; Room, R. Variations by age and sex in alcohol-related problematic behaviour per drinking volume and heavier drinking occasion. Drug Alcohol Depend. 2009, 101, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Jacobi, F.; Mack, S.; Gerschler, A.; Scholl, L.; Hofler, M.; Siegert, J.; Burkner, A.; Preiss, S.; Spitzer, K.; Busch, M.; et al. The design and methods of the mental health module in the german health interview and examination survey for adults (degs1-mh). Int. J. Methods Psychiatr. Res. 2013, 22, 83–99. [Google Scholar] [CrossRef] [PubMed]
- Wittchen, H.-U.; Pfister, H. Dia-x-Interviews: Manual für Screening-Verfahren und Interview; Interviewheft Längsschnittuntersuchung (Dia-x-Lifetime); Ergänzungsheft (Dia-x-Lifetime); Interviewheft Querschnittuntersuchung (Dia-x-12 Monate); Ergänzungsheft (dia-x-12 Monate); Pc-Programm zur Durchführung des Interviews (Längs- und Querschnittuntersuchung); Auswertungsprogramm; Swets & Zeitlinger: Frankfurt, Germany, 1997. [Google Scholar]
- Saß, H.; Wittchen, H.-U.; Zaudig, M.H.I. Diagnostisches und Statistisches Manual Psychischer Störungen—Textrevision—Dsm-iv-tr; Hogrefe: Göttingen, Germany, 2003. [Google Scholar]
- Sebold, M.; Nebe, S.; Garbusow, M.; Guggenmos, M.; Schad, D.J.; Beck, A.; Kuitunen-Paul, S.; Sommer, C.; Frank, R.; Neu, P.; et al. When habits are dangerous: Alcohol expectancies and habitual decision making predict relapse in alcohol dependence. Biol. Psychiatry 2017, 82, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Daw, N.D.; Gershman, S.J.; Seymour, B.; Dayan, P.; Dolan, R.J. Model-based influences on humans’ choices and striatal prediction errors. Neuron 2011, 69, 1204–1215. [Google Scholar] [CrossRef] [PubMed]
- Garbusow, M.; Schad, D.J.; Sommer, C.; Jünger, E.; Sebold, M.; Friedel, E.; Wendt, J.; Kathmann, N.; Schlagenhauf, F.; Zimmermann, U.S.; et al. Pavlovian-to-instrumental transfer in alcohol dependence: A pilot study. Neuropsychobiology 2014, 70, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Skinner, H.A.; Horn, J.L. Alcohol Dependence Scale (ads): Users Guide; Addiction Research Foundation: Toronto, ON, Canada, 1984. [Google Scholar]
- Mann, K.; Ackermann, K. Die ocds-g: Psychometrische kennwerte der deutschen version der obsessive compulsive drinking scale. Sucht 2000, 46, 90–100. [Google Scholar] [CrossRef]
- Heatherton, T.F.; Kozlowski, L.T.; Frecker, R.C.; Fagerstrom, K.O. The fagerstrom test for nicotine dependence: A revision of the fagerstrom tolerance questionnaire. Br. J. Addict. 1991, 86, 1119–1127. [Google Scholar] [CrossRef]
- Nebe, S.; Kroemer, N.B.; Schad, D.J.; Bernhardt, N.; Sebold, M.; Muller, D.K.; Scholl, L.; Kuitunen-Paul, S.; Heinz, A.; Rapp, M.A.; et al. No association of goal-directed and habitual control with alcohol consumption in young adults. Addict. Biol. 2017, 23, 379–393. [Google Scholar] [CrossRef]
- Brainard, D.H. The psychophysics toolbox. Spat. Vis. 1997, 10, 433–436. [Google Scholar] [CrossRef]
- Infinium PsychArray-24 Kit | Psychiatric Predisposition Microarray. Available online: https://www.illumina.com/products/by-type/microarray-kits/infinium-psycharray.html (accessed on 25 June 2019).
- Purcell, S.M.; Wray, N.R.; Stone, J.L.; Visscher, P.M.; O’Donovan, M.C.; Sullivan, P.F.; Sklar, P. Common polygenic variation contributes to risk of schizophrenia and bipolar disorder. Nature 2009, 460, 748–752. [Google Scholar] [PubMed]
- Matlab, version 7.12.0; The MathWorks Inc.: Natick, MA, USA, 2011.
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Statistical Parametric Mapping. Available online: http://www.fil.ion.ucl.ac.uk/spm (accessed on 7 August 2019).
- Pick Atlas Software. Available online: http://www.fmri.wfubmc.edu/download.htm (accessed on 7 August 2019).
- Wassum, K.M.; Izquierdo, A. The basolateral amygdala in reward learning and addiction. Neurosci. Biobehav. Rev. 2015, 57, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Mendelsohn, A.; Pine, A.; Schiller, D. Between thoughts and actions: Motivationally salient cues invigorate mental action in the human brain. Neuron 2014, 81, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Van Steenbergen, H.; Watson, P.; Wiers, R.W.; Hommel, B.; de Wit, S. Dissociable corticostriatal circuits underlie goal-directed vs. Cue-elicited habitual food seeking after satiation: Evidence from a multimodal mri study. Eur. J. Neurosci. 2017, 46, 1815–1827. [Google Scholar] [CrossRef] [PubMed]
- Boileau, I.; Assaad, J.M.; Pihl, R.O.; Benkelfat, C.; Leyton, M.; Diksic, M.; Tremblay, R.E.; Dagher, A. Alcohol promotes dopamine release in the human nucleus accumbens. Synapse 2003, 49, 226–231. [Google Scholar] [CrossRef]
- Belin, D.; Mar, A.C.; Dalley, J.W.; Robbins, T.W.; Everitt, B.J. High impulsivity predicts the switch to compulsive cocaine-taking. Science 2008, 320, 1352. [Google Scholar] [CrossRef]
- Robbins, T.W.; Gillan, C.M.; Smith, D.G.; de Wit, S.; Ersche, K.D. Neurocognitive endophenotypes of impulsivity and compulsivity: Towards dimensional psychiatry. Trends Cogn. Sci. 2012, 16, 81–91. [Google Scholar] [CrossRef]
- Robinson, T.E.; Berridge, K.C. The neural basis of drug craving: An incentive-sensitization theory of addiction. Brain Res. Brain Res. Rev. 1993, 18, 247–291. [Google Scholar] [CrossRef]
- Ostafin, B.D.; Palfai, T.P. When wanting to change is not enough: Automatic appetitive processes moderate the effects of a brief alcohol intervention in hazardous-drinking college students. Addict. Sci. Clin. Pract. 2012, 7, 25. [Google Scholar] [CrossRef]
- Wiers, R.W.; Bartholow, B.D.; van den Wildenberg, E.; Thush, C.; Engels, R.C.; Sher, K.J.; Grenard, J.; Ames, S.L.; Stacy, A.W. Automatic and controlled processes and the development of addictive behaviors in adolescents: A review and a model. Pharmacol. Biochem. Behav. 2007, 86, 263–283. [Google Scholar] [CrossRef]
- Wiers, R.W.; Eberl, C.; Rinck, M.; Becker, E.S.; Lindenmeyer, J. Retraining automatic action tendencies changes alcoholic patients’ approach bias for alcohol and improves treatment outcome. Psychol. Sci. 2011, 22, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Erol, A.; Karpyak, V.M. Sex and gender-related differences in alcohol use and its consequences: Contemporary knowledge and future research considerations. Drug Alcohol Depend. 2015, 156, 1–13. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garbusow, M.; Nebe, S.; Sommer, C.; Kuitunen-Paul, S.; Sebold, M.; Schad, D.J.; Friedel, E.; Veer, I.M.; Wittchen, H.-U.; Rapp, M.A.; et al. Pavlovian-To-Instrumental Transfer and Alcohol Consumption in Young Male Social Drinkers: Behavioral, Neural and Polygenic Correlates. J. Clin. Med. 2019, 8, 1188. https://doi.org/10.3390/jcm8081188
Garbusow M, Nebe S, Sommer C, Kuitunen-Paul S, Sebold M, Schad DJ, Friedel E, Veer IM, Wittchen H-U, Rapp MA, et al. Pavlovian-To-Instrumental Transfer and Alcohol Consumption in Young Male Social Drinkers: Behavioral, Neural and Polygenic Correlates. Journal of Clinical Medicine. 2019; 8(8):1188. https://doi.org/10.3390/jcm8081188
Chicago/Turabian StyleGarbusow, Maria, Stephan Nebe, Christian Sommer, Sören Kuitunen-Paul, Miriam Sebold, Daniel J. Schad, Eva Friedel, Ilya M. Veer, Hans-Ulrich Wittchen, Michael A. Rapp, and et al. 2019. "Pavlovian-To-Instrumental Transfer and Alcohol Consumption in Young Male Social Drinkers: Behavioral, Neural and Polygenic Correlates" Journal of Clinical Medicine 8, no. 8: 1188. https://doi.org/10.3390/jcm8081188