Immunogenicity and Efficacy Evaluation of Subunit Astrovirus Vaccines

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
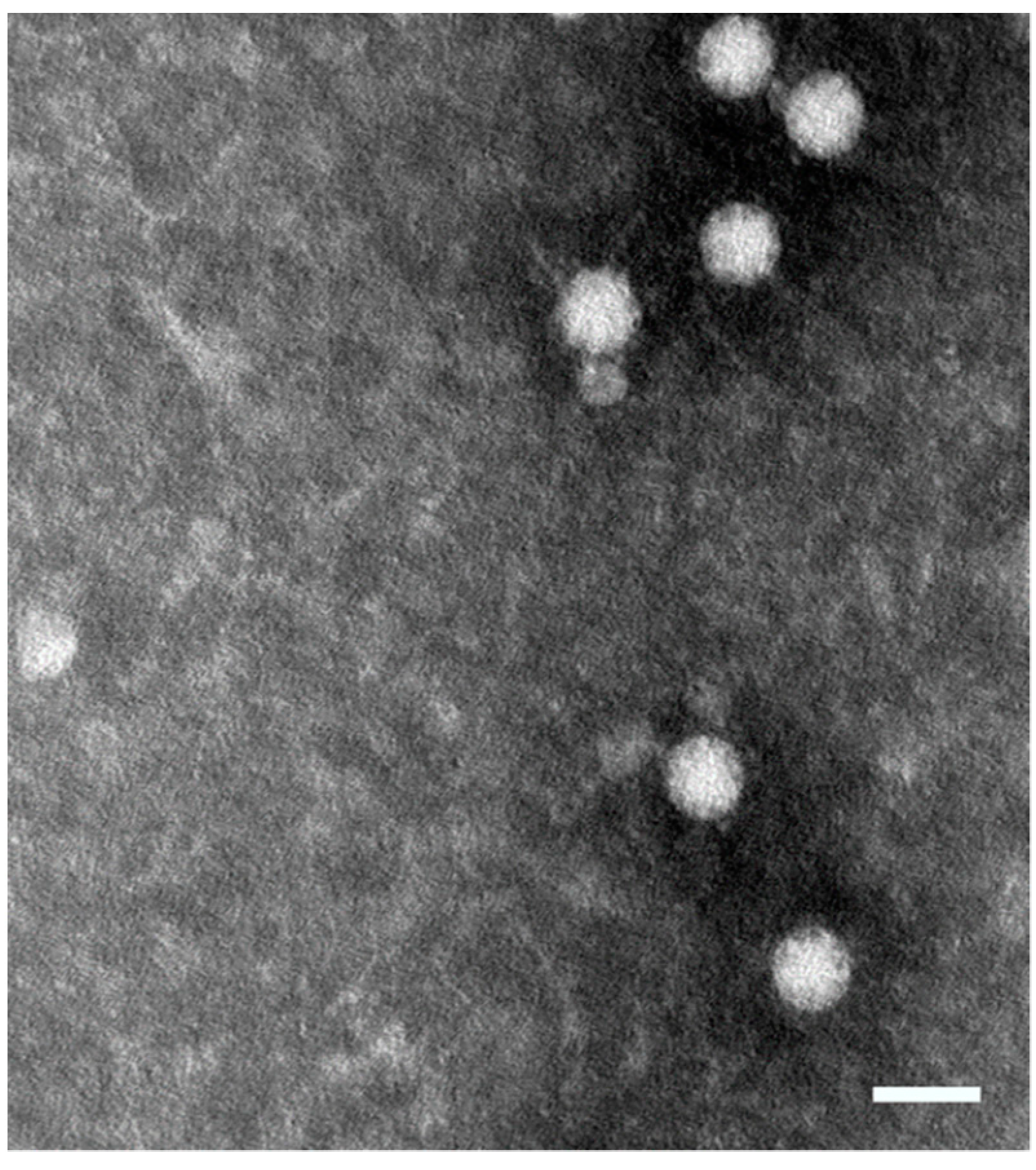
2.1. Construction and Expression of MAstV Vaccine Candidates
2.2. Immunogenicity Evaluation in Mice
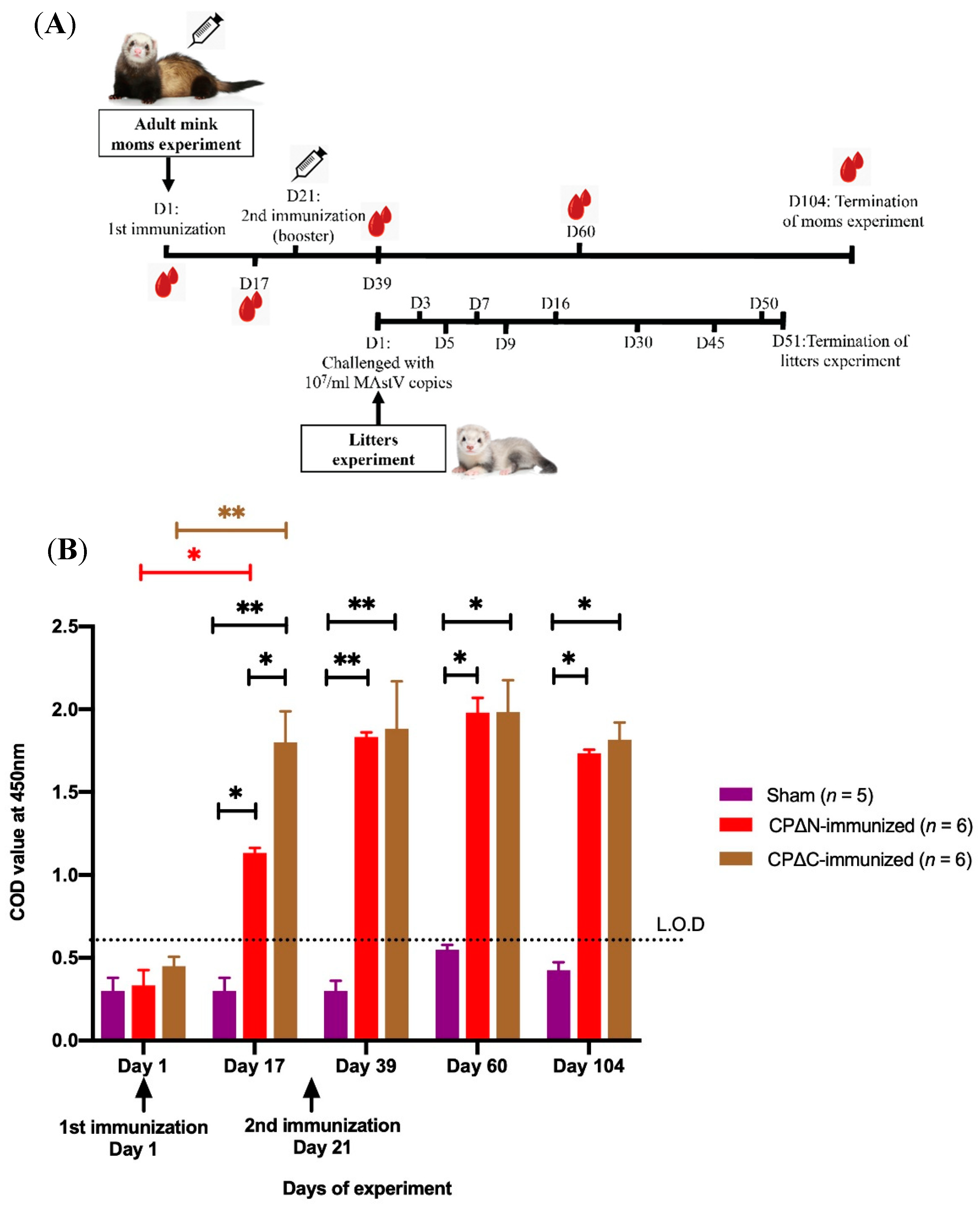
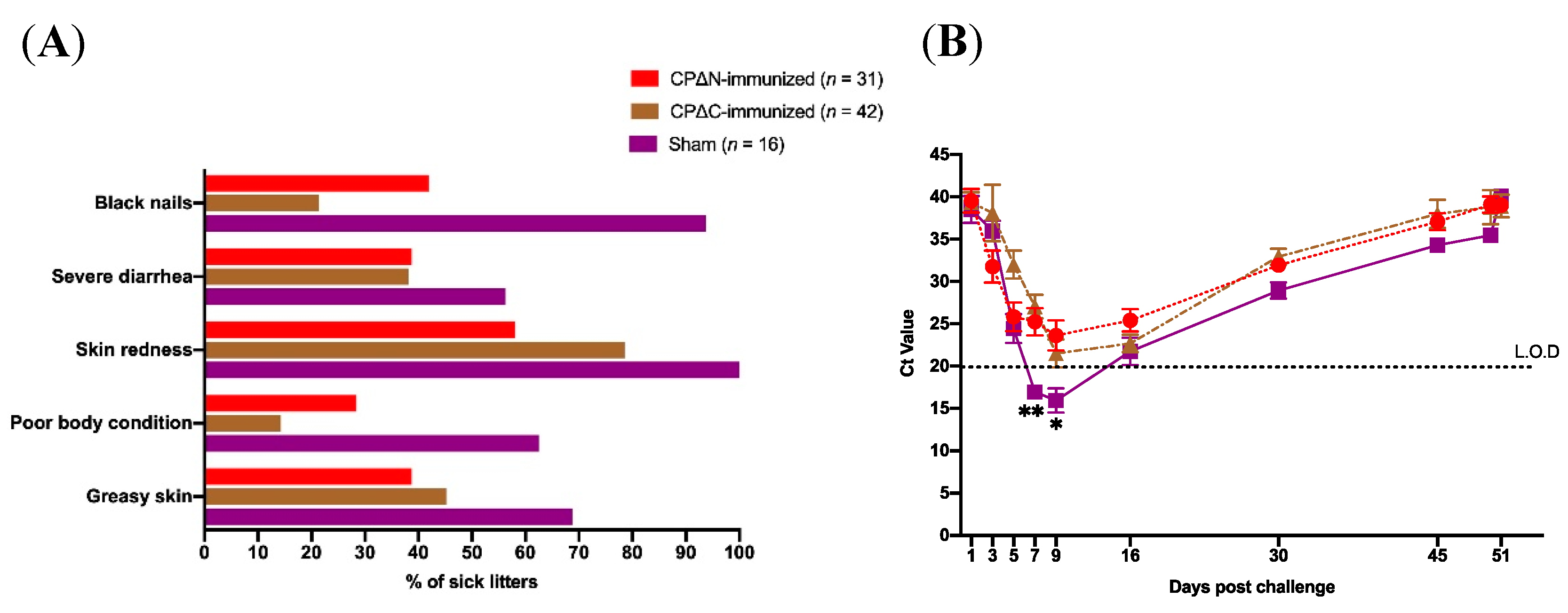
2.3. Protective Efficacy Evaluation in Mink
2.4. Indirect ELISA
2.5. Proliferation Assay
2.6. Cytokine Profiling
2.7. Statistical Analysis
3. Results
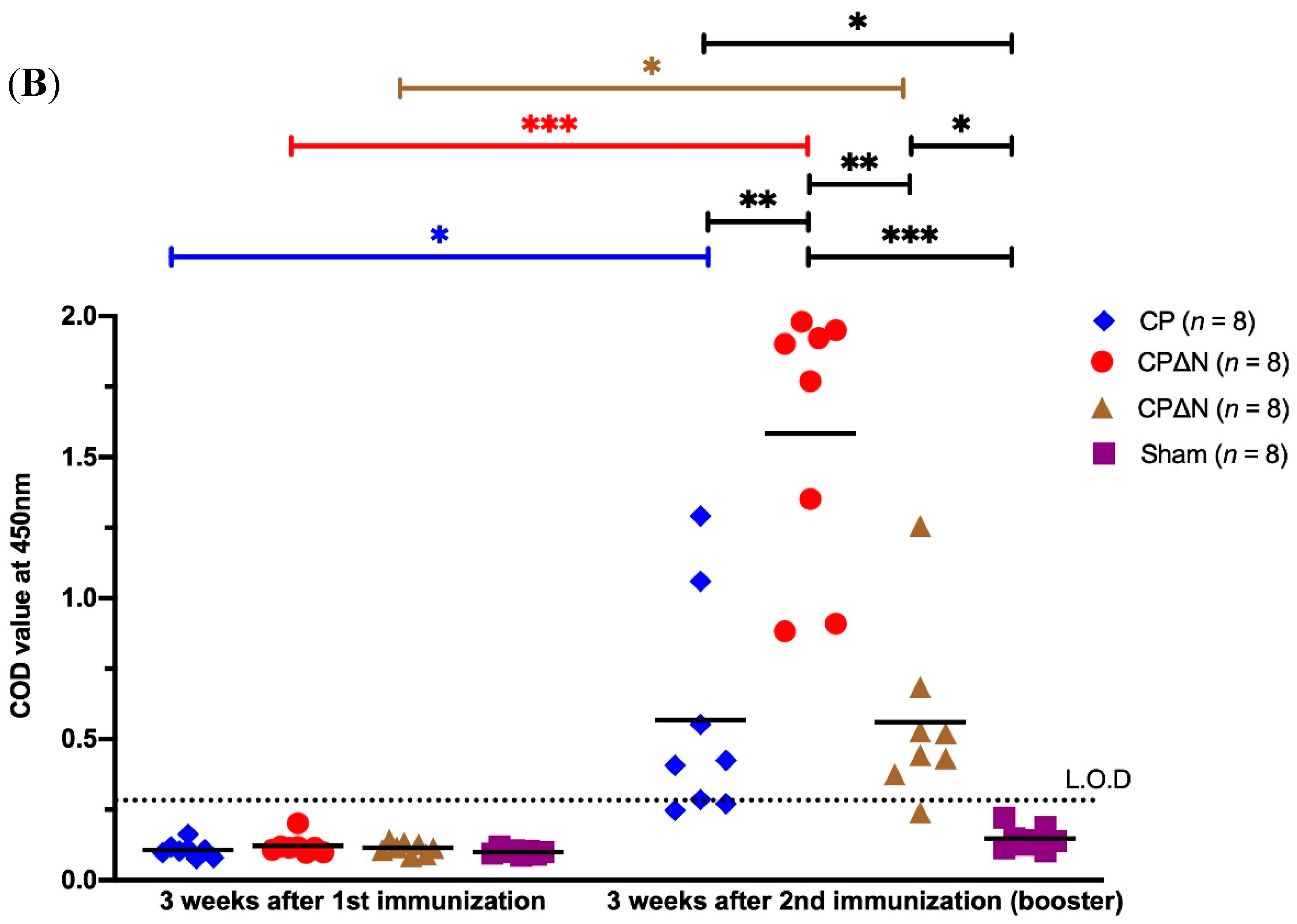
3.1. Antibody Responses to MAstV CPs in Mice
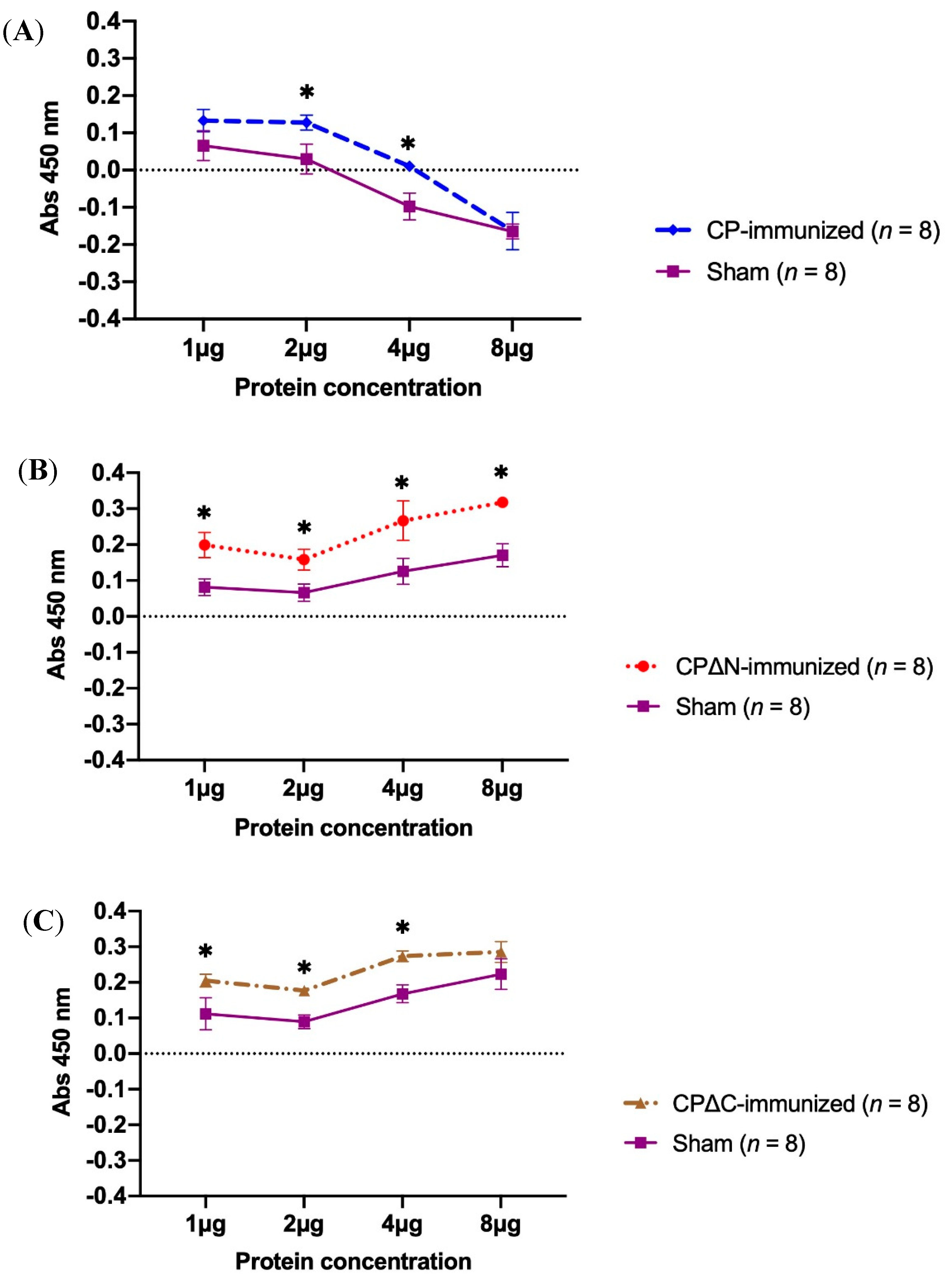
3.2. Proliferation Activity of Mice Splenocytes Immunized with MAstV CPs
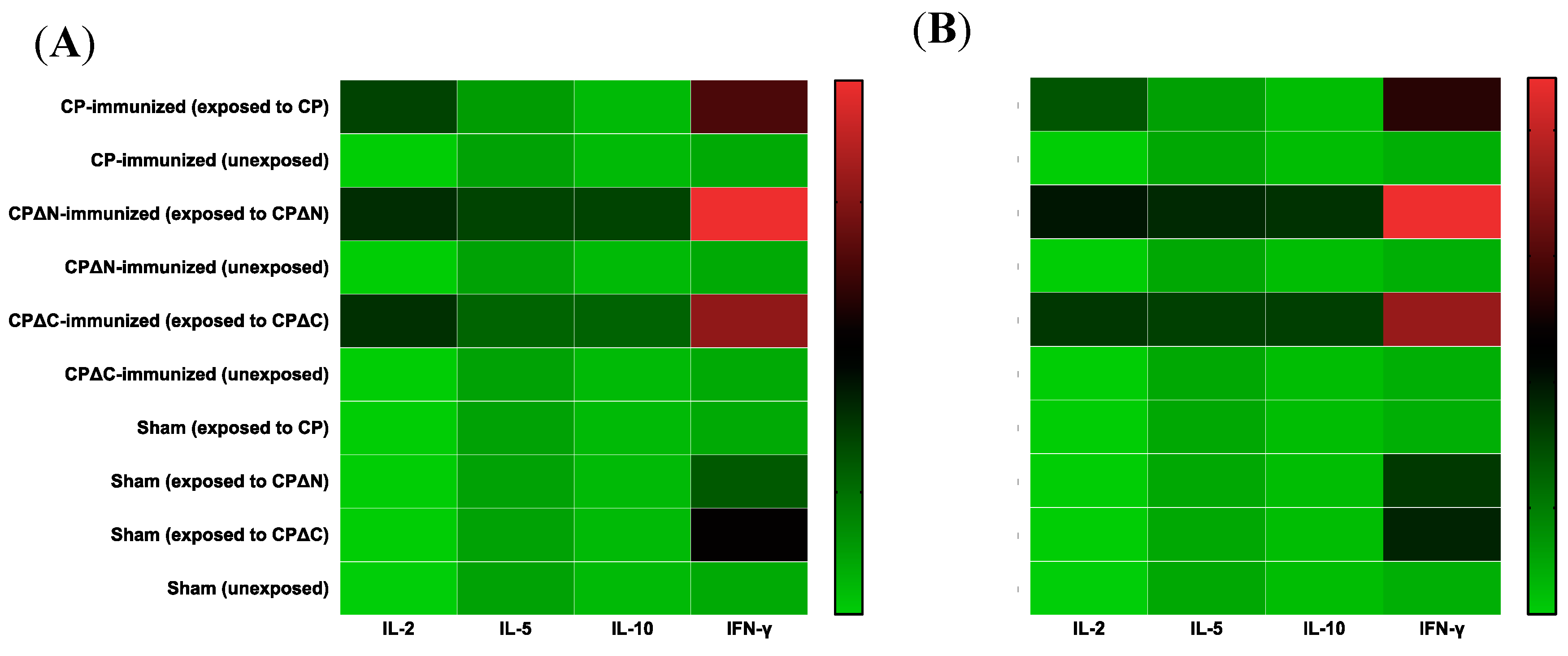
3.3. Cytokine Profiling of MAstV CPs-Immunized Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Holmgren, J.; Svennerholm, A.M. Vaccines against mucosal infections. Curr. Opin. Immunol. 2012, 24, 343–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plotkin, S.A. Vaccines, vaccination, and vaccinology. J. Infect. Dis. 2003, 187, 1349–1359. [Google Scholar] [CrossRef] [PubMed]
- Suschak, J.J.; Williams, J.A.; Schmaljohn, C.S. Advancements in DNA vaccine vectors, non-mechanical delivery methods, and molecular adjuvants to increase immunogenicity. Hum. Vaccines Immunother. 2017, 13, 2837–2848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M.; Jiang, X. Recent advancements in combination subunit vaccine development. Hum. Vaccines Immunother. 2017, 13, 180–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphreys, I.R.; Sebastian, S. Novel viral vectors in infectious diseases. Immunology 2018, 153, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.T.; Chow, Y.H.; Hsiao, K.N.; Hu, K.C.; Chiang, J.R.; Wu, S.C.; Chong, P.; Liu, C.C. Development of a full-length cDNA-derived enterovirus A71 vaccine candidate using reverse genetics technology. Antivir. Res. 2016, 132, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Englund, L.; Chriel, M.; Dietz, H.H.; Hedlund, K.O. Astrovirus epidemiologically linked to pre-weaning diarrhoea in mink. Vet. Microbiol. 2002, 85, 1–11. [Google Scholar] [CrossRef]
- Geigenmuller, U.; Chew, T.; Ginzton, N.; Matsui, S.M. Processing of nonstructural protein 1a of human astrovirus. J. Virol. 2002, 76, 2003–2008. [Google Scholar] [CrossRef]
- Mendez, E.; Salas-Ocampo, M.P.; Munguia, M.E.; Arias, C.F. Protein products of the open reading frames encoding nonstructural proteins of human astrovirus serotype 8. J. Virol. 2003, 77, 11378–11384. [Google Scholar] [CrossRef]
- Mittelholzer, C.; Hedlund, K.O.; Englund, L.; Dietz, H.H.; Svensson, L. Molecular characterization of a novel astrovirus associated with disease in mink. J. Gen. Virol. 2003, 84, 3087–3094. [Google Scholar] [CrossRef] [PubMed]
- Lewis, T.L.; Greenberg, H.B.; Herrmann, J.E.; Smith, L.S.; Matsui, S.M. Analysis of astrovirus serotype 1 RNA, identification of the viral RNA-dependent RNA polymerase motif, and expression of a viral structural protein. J. Virol. 1994, 68, 77–83. [Google Scholar] [PubMed]
- Monroe, S.S.; Jiang, B.; Stine, S.E.; Koopmans, M.; Glass, R.I. Subgenomic RNA sequence of human astrovirus supports classification of Astroviridae as a new family of RNA viruses. J. Virol. 1993, 67, 3611–3614. [Google Scholar] [Green Version]
- Willcocks, M.M.; Brown, T.D.; Madeley, C.R.; Carter, M.J. The complete sequence of a human astrovirus. J. Gen. Virol. 1994, 75 Pt 7, 1785–1788. [Google Scholar] [CrossRef]
- Mendez, E.; Aguirre-Crespo, G.; Zavala, G.; Arias, C.F. Association of the astrovirus structural protein VP90 with membranes plays a role in virus morphogenesis. J. Virol. 2007, 81, 10649–10658. [Google Scholar] [CrossRef] [PubMed]
- Toh, Y.; Harper, J.; Dryden, K.A.; Yeager, M.; Arias, C.F.; Mendez, E.; Tao, Y.J. Crystal Structure of the Human Astrovirus Capsid Protein. J. Virol. 2016, 90, 9008–9017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donelli, G.; Superti, F.; Tinari, A.; Marziano, M.L. Mechanism of astrovirus entry into Graham 293 cells. J. Med. Virol. 1992, 38, 271–277. [Google Scholar] [CrossRef]
- Bridger, J.C.; Hall, G.A.; Brown, J.F. Characterization of a calici-like virus (Newbury agent) found in association with astrovirus in bovine diarrhea. Infect. Immun. 1984, 43, 133–138. [Google Scholar] [PubMed]
- Jonassen, C.M.; Jonassen, T.T.; Sveen, T.M.; Grinde, B. Complete genomic sequences of astroviruses from sheep and turkey: Comparison with related viruses. Virus Res. 2003, 91, 195–201. [Google Scholar] [CrossRef]
- Indik, S.; Valicek, L.; Smid, B.; Dvorakova, H.; Rodak, L. Isolation and partial characterization of a novel porcine astrovirus. Vet. Microbiol. 2006, 117, 276–283. [Google Scholar] [CrossRef]
- Hoshino, Y.; Zimmer, J.F.; Moise, N.S.; Scott, F.W. Detection of astroviruses in feces of a cat with diarrhea. Brief report. Arch. Virol. 1981, 70, 373–376. [Google Scholar] [CrossRef]
- Toffan, A.; Jonassen, C.M.; De Battisti, C.; Schiavon, E.; Kofstad, T.; Capua, I.; Cattoli, G. Genetic characterization of a new astrovirus detected in dogs suffering from diarrhoea. Vet. Microbiol. 2009, 139, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Rivera, R.; Nollens, H.H.; Venn-Watson, S.; Gulland, F.M.; Wellehan, J.F., Jr. Characterization of phylogenetically diverse astroviruses of marine mammals. J. Gen. Virol. 2010, 91, 166–173. [Google Scholar] [CrossRef]
- Pantin-Jackwood, M.J.; Strother, K.O.; Mundt, E.; Zsak, L.; Day, J.M.; Spackman, E. Molecular characterization of avian astroviruses. Arch. Virol. 2011, 156, 235–244. [Google Scholar] [CrossRef]
- Koci, M.D.; Schultz-Cherry, S. Avian astroviruses. Avian Pathol. 2002, 31, 213–227. [Google Scholar] [CrossRef] [Green Version]
- Quan, P.L.; Wagner, T.A.; Briese, T.; Torgerson, T.R.; Hornig, M.; Tashmukhamedova, A.; Firth, C.; Palacios, G.; Baisre-De-Leon, A.; Paddock, C.D.; et al. Astrovirus encephalitis in boy with X-linked agammaglobulinemia. Emerg. Infect. Dis. 2010, 16, 918–925. [Google Scholar] [CrossRef]
- Vu, D.L.; Bosch, A.; Pinto, R.M.; Guix, S. Epidemiology of Classic and Novel Human Astrovirus: Gastroenteritis and Beyond. Viruses 2017, 9, 33. [Google Scholar] [CrossRef]
- Baule, C.; Bidokhti, M.R.; Chriel, M.; Czifra, G.; Dietz, H.H.; Hammer, A.S.; Sandbol, P.; Ullman, K. Recombinant Proteins as Vaccines for Protection against Disease Induced by Infection with Mink Astrovirus. Patent WO2012013191 A3, 21 June 2012. [Google Scholar]
- Pfaff, F.; Schlottau, K.; Scholes, S.; Courtenay, A.; Hoffmann, B.; Hoper, D.; Beer, M. A novel astrovirus associated with encephalitis and ganglionitis in domestic sheep. Transbound. Emerg. Dis. 2017, 64, 677–682. [Google Scholar] [CrossRef]
- Bouzalas, I.G.; Wuthrich, D.; Walland, J.; Drogemuller, C.; Zurbriggen, A.; Vandevelde, M.; Oevermann, A.; Bruggmann, R.; Seuberlich, T. Neurotropic astrovirus in cattle with nonsuppurative encephalitis in Europe. J. Clin. Microbiol. 2014, 52, 3318–3324. [Google Scholar] [CrossRef]
- Schneider, R.R.; Hunter, D.B. Mortality in mink kits from birth to weaning. Can. Vet. J. 1993, 34, 159–163. [Google Scholar]
- Gavier-Widen, D.; Brojer, C.; Dietz, H.H.; Englund, L.; Hammer, A.S.; Hedlund, K.O.; Hard af Segerstad, C.; Nilsson, K.; Nowotny, N.; Puurula, V.; et al. Investigations into shaking mink syndrome: An encephalomyelitis of unknown cause in farmed mink (Mustela vison) kits in Scandinavia. J. Vet. Diagn. Investig. 2004, 16, 305–312. [Google Scholar] [CrossRef]
- Perot, P.; Lecuit, M.; Eloit, M. Astrovirus Diagnostics. Viruses 2017, 9, 10. [Google Scholar] [CrossRef]
- Bass, D.M.; Qiu, S. Proteolytic processing of the astrovirus capsid. J. Virol. 2000, 74, 1810–1814. [Google Scholar] [CrossRef]
- Dong, J.; Dong, L.; Mendez, E.; Tao, Y. Crystal structure of the human astrovirus capsid spike. Proc. Natl. Acad. Sci. USA 2011, 108, 12681–12686. [Google Scholar] [CrossRef] [Green Version]
- Bidokhti, M.R.; Ullman, K.; Jensen, T.H.; Chriel, M.; Mottahedin, A.; Munir, M.; Andersson, A.M.; Detournay, O.; Hammer, A.S.; Baule, C. Establishment of stably transfected cells constitutively expressing the full-length and truncated antigenic proteins of two genetically distinct mink astroviruses. PLoS ONE 2013, 8, e82978. [Google Scholar] [CrossRef]
- Sellers, H.; Linneman, E.; Icard, A.H.; Mundt, E. A purified recombinant baculovirus expressed capsid protein of a new astrovirus provides partial protection to runting-stunting syndrome in chickens. Vaccine 2010, 28, 1253–1263. [Google Scholar] [CrossRef]
- Johnson, C.; Hargest, V.; Cortez, V.; Meliopoulos, V.A.; Schultz-Cherry, S. Astrovirus Pathogenesis. Viruses 2017, 9, 22. [Google Scholar] [CrossRef]
- Todd, D.; Wilkinson, D.S.; Jewhurst, H.L.; Wylie, M.; Gordon, A.W.; Adair, B.M. A seroprevalence investigation of chicken astrovirus infections. Avian Pathol. 2009, 38, 301–309. [Google Scholar] [CrossRef]
- Greenberg, H.B.; Matsui, S.M. Astroviruses and caliciviruses: Emerging enteric pathogens. Infect. Agents Dis. 1992, 1, 71–91. [Google Scholar]
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals—Molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef]
- Donato, C.; Vijaykrishna, D. The Broad Host Range and Genetic Diversity of Mammalian and Avian Astroviruses. Viruses 2017, 9, 102. [Google Scholar] [CrossRef]
- Smits, S.L.; van Leeuwen, M.; van der Eijk, A.A.; Fraaij, P.L.; Escher, J.C.; Simon, J.H.; Osterhaus, A.D. Human astrovirus infection in a patient with new-onset celiac disease. J. Clin. Microbiol. 2010, 48, 3416–3418. [Google Scholar] [CrossRef]
- Finkbeiner, S.R.; Li, Y.; Ruone, S.; Conrardy, C.; Gregoricus, N.; Toney, D.; Virgin, H.W.; Anderson, L.J.; Vinje, J.; Wang, D.; et al. Identification of a novel astrovirus (astrovirus VA1) associated with an outbreak of acute gastroenteritis. J. Virol. 2009, 83, 10836–10839. [Google Scholar] [CrossRef]
- Finkbeiner, S.R.; Le, B.M.; Holtz, L.R.; Storch, G.A.; Wang, D. Detection of newly described astrovirus MLB1 in stool samples from children. Emerg. Infect. Dis. 2009, 15, 441–444. [Google Scholar] [CrossRef]
- Marvin, S.A. The Immune Response to Astrovirus Infection. Viruses 2016, 9, 1. [Google Scholar] [CrossRef]
- Bogdanoff, W.A.; Campos, J.; Perez, E.I.; Yin, L.; Alexander, D.L.; DuBois, R.M. Structure of a Human Astrovirus Capsid-Antibody Complex and Mechanistic Insights into Virus Neutralization. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Bogdanoff, W.A.; Perez, E.I.; Lopez, T.; Arias, C.F.; DuBois, R.M. Structural Basis for Escape of Human Astrovirus from Antibody Neutralization: Broad Implications for Rational Vaccine Design. J. Virol. 2018, 92. [Google Scholar] [CrossRef]
- Gronemus, J.Q.; Hair, P.S.; Crawford, K.B.; Nyalwidhe, J.O.; Cunnion, K.M.; Krishna, N.K. Potent inhibition of the classical pathway of complement by a novel C1q-binding peptide derived from the human astrovirus coat protein. Mol. Immunol. 2010, 48, 305–313. [Google Scholar] [CrossRef]
- Bonaparte, R.S.; Hair, P.S.; Banthia, D.; Marshall, D.M.; Cunnion, K.M.; Krishna, N.K. Human astrovirus coat protein inhibits serum complement activation via C1, the first component of the classical pathway. J. Virol. 2008, 82, 817–827. [Google Scholar] [CrossRef]
- Tacket, C.O.; Sztein, M.B.; Losonsky, G.A.; Wasserman, S.S.; Estes, M.K. Humoral, mucosal, and cellular immune responses to oral Norwalk virus-like particles in volunteers. Clin. Immunol. 2003, 108, 241–247. [Google Scholar] [CrossRef]
- Wang, S.; Liu, H.; Zu, X.; Liu, Y.; Chen, L.; Zhu, X.; Zhang, L.; Zhou, Z.; Xiao, G.; Wang, W. The ubiquitin-proteasome system is essential for the productive entry of Japanese encephalitis virus. Virology 2016, 498, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Guix, S.; Perez-Bosque, A.; Miro, L.; Moreto, M.; Bosch, A.; Pinto, R.M. Type I interferon response is delayed in human astrovirus infections. PLoS ONE 2015, 10, e0123087. [Google Scholar] [CrossRef] [PubMed]
- Marvin, S.A.; Huerta, C.T.; Sharp, B.; Freiden, P.; Cline, T.D.; Schultz-Cherry, S. Type I Interferon Response Limits Astrovirus Replication and Protects against Increased Barrier Permeability In Vitro and In Vivo. J. Virol. 2016, 90, 1988–1996. [Google Scholar] [CrossRef] [PubMed]
- Sen, A.; Sharma, A.; Greenberg, H.B. Rotavirus degrades multiple type interferon receptors to inhibit IFN signaling and protects against mortality from endotoxin in suckling mice. J. Virol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Frias, A.H.; Jones, R.M.; Fifadara, N.H.; Vijay-Kumar, M.; Gewirtz, A.T. Rotavirus-induced IFN-beta promotes anti-viral signaling and apoptosis that modulate viral replication in intestinal epithelial cells. Innate Immun. 2012, 18, 294–306. [Google Scholar] [CrossRef]
- Koci, M.D.; Kelley, L.A.; Larsen, D.; Schultz-Cherry, S. Astrovirus-induced synthesis of nitric oxide contributes to virus control during infection. J. Virol. 2004, 78, 1564–1574. [Google Scholar] [CrossRef]
- Yokoyama, C.C.; Loh, J.; Zhao, G.; Stappenbeck, T.S.; Wang, D.; Huang, H.V.; Virgin, H.W.; Thackray, L.B. Adaptive immunity restricts replication of novel murine astroviruses. J. Virol. 2012, 86, 12262–12270. [Google Scholar] [CrossRef]
- Kapoor, A.; Li, L.; Victoria, J.; Oderinde, B.; Mason, C.; Pandey, P.; Zaidi, S.Z.; Delwart, E. Multiple novel astrovirus species in human stool. J. Gen. Virol. 2009, 90, 2965–2972. [Google Scholar] [CrossRef]
- Bass, D.M.; Upadhyayula, U. Characterization of human serotype 1 astrovirus-neutralizing epitopes. J. Virol. 1997, 71, 8666–8671. [Google Scholar] [Green Version]
- Meyerhoff, R.R.; Nighot, P.K.; Ali, R.A.; Blikslager, A.T.; Koci, M.D. Characterization of turkey inducible nitric oxide synthase and identification of its expression in the intestinal epithelium following astrovirus infection. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Koci, M.D.; Moser, L.A.; Kelley, L.A.; Larsen, D.; Brown, C.C.; Schultz-Cherry, S. Astrovirus induces diarrhea in the absence of inflammation and cell death. J. Virol. 2003, 77, 11798–11808. [Google Scholar] [CrossRef] [PubMed]
- Meliopoulos, V.A.; Marvin, S.A.; Freiden, P.; Moser, L.A.; Nighot, P.; Ali, R.; Blikslager, A.; Reddivari, M.; Heath, R.J.; Koci, M.D.; et al. Oral Administration of Astrovirus Capsid Protein Is Sufficient to Induce Acute Diarrhea In Vivo. MBio 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Woode, G.N.; Pohlenz, J.F.; Gourley, N.E.; Fagerland, J.A. Astrovirus and Breda virus infections of dome cell epithelium of bovine ileum. J. Clin. Microbiol. 1984, 19, 623–630. [Google Scholar] [PubMed]
- Gray, E.W.; Angus, K.W.; Snodgrass, D.R. Ultrastructure of the small intestine in astrovirus-infected lambs. J. Gen. Virol. 1980, 49, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Madhi, S.A.; Cutland, C.L.; Kuwanda, L.; Weinberg, A.; Hugo, A.; Jones, S.; Adrian, P.V.; van Niekerk, N.; Treurnicht, F.; Ortiz, J.R.; et al. Influenza vaccination of pregnant women and protection of their infants. N. Engl. J. Med. 2014, 371, 918–931. [Google Scholar] [CrossRef] [PubMed]
- Pulit-Penaloza, J.A.; Esser, E.S.; Vassilieva, E.V.; Lee, J.W.; Taherbhai, M.T.; Pollack, B.P.; Prausnitz, M.R.; Compans, R.W.; Skountzou, I. A protective role of murine langerin(+) cells in immune responses to cutaneous vaccination with microneedle patches. Sci. Rep. 2014, 4, 6094. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.H.; Chu, C.; He, C.C.; Lin, T.Y. Protection of neonatal mice from lethal enterovirus 71 infection by maternal immunization with attenuated Salmonella enterica serovar Typhimurium expressing VP1 of enterovirus 71. Microbes Infect. 2006, 8, 1671–1678. [Google Scholar] [CrossRef]
- Kim, Y.I.; Song, J.H.; Kwon, B.E.; Kim, H.N.; Seo, M.D.; Park, K.; Lee, S.; Yeo, S.G.; Kweon, M.N.; Ko, H.J.; et al. Pros and cons of VP1-specific maternal IgG for the protection of Enterovirus 71 infection. Vaccine 2015, 33, 6604–6610. [Google Scholar] [CrossRef] [PubMed]
- Munoz, F.M. Respiratory syncytial virus in infants: Is maternal vaccination a realistic strategy? Curr. Opin. Infect. Dis. 2015, 28, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Pan-Ngum, W.; Kinyanjui, T.; Kiti, M.; Taylor, S.; Toussaint, J.F.; Saralamba, S.; Van Effelterre, T.; Nokes, D.J.; White, L.J. Predicting the relative impacts of maternal and neonatal respiratory syncytial virus (RSV) vaccine target product profiles: A consensus modelling approach. Vaccine 2017, 35, 403–409. [Google Scholar] [CrossRef]
- Eberhardt, C.S.; Blanchard-Rohner, G.; Lemaitre, B.; Combescure, C.; Othenin-Girard, V.; Chilin, A.; Petre, J.; Martinez de Tejada, B.; Siegrist, C.A. Pertussis Antibody Transfer to Preterm Neonates After Second- Versus Third-Trimester Maternal Immunization. Clin. Infect. Dis. 2017, 64, 1129–1132. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, C.S.; Blanchard-Rohner, G.; Lemaitre, B.; Boukrid, M.; Combescure, C.; Othenin-Girard, V.; Chilin, A.; Petre, J.; de Tejada, B.M.; Siegrist, C.A. Maternal Immunization Earlier in Pregnancy Maximizes Antibody Transfer and Expected Infant Seropositivity Against Pertussis. Clin. Infect. Dis. 2016, 62, 829–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heath, P.T. An update on vaccination against group B streptococcus. Expert Rev. Vaccines 2011, 10, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Amirthalingam, G.; Andrews, N.; Campbell, H.; Ribeiro, S.; Kara, E.; Donegan, K.; Fry, N.K.; Miller, E.; Ramsay, M. Effectiveness of maternal pertussis vaccination in England: An observational study. Lancet 2014, 384, 1521–1528. [Google Scholar] [CrossRef]
- Dabrera, G.; Amirthalingam, G.; Andrews, N.; Campbell, H.; Ribeiro, S.; Kara, E.; Fry, N.K.; Ramsay, M. A case-control study to estimate the effectiveness of maternal pertussis vaccination in protecting newborn infants in England and Wales, 2012–2013. Clin. Infect. Dis. 2015, 60, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Janowski, A.B.; Bauer, I.K.; Holtz, L.R.; Wang, D. Propagation of astrovirus VA1, a neurotropic human astrovirus, in cell culture. J. Virol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Gattoni, A.; Parlato, A.; Vangieri, B.; Bresciani, M.; Derna, R. Interferon-gamma: Biologic functions and HCV therapy (type I/II) (1 of 2 parts). Clin. Ter. 2006, 157, 377–386. [Google Scholar]
- Molberg, O.; Nilsen, E.M.; Sollid, L.M.; Scott, H.; Brandtzaeg, P.; Thorsby, E.; Lundin, K.E. CD4+ T cells with specific reactivity against astrovirus isolated from normal human small intestine. Gastroenterology 1998, 114, 115–122. [Google Scholar] [CrossRef]
- Souza, M.; Costantini, V.; Azevedo, M.S.; Saif, L.J. A human norovirus-like particle vaccine adjuvanted with ISCOM or mLT induces cytokine and antibody responses and protection to the homologous GII.4 human norovirus in a gnotobiotic pig disease model. Vaccine 2007, 25, 8448–8459. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Wang, J.; Zhou, H.; Si, H.; Wang, M.; Song, J.; Han, B.; Shu, Y.; Ren, L.; Qu, J.; et al. Intranasal administration of a recombinant adenovirus expressing the norovirus capsid protein stimulates specific humoral, mucosal, and cellular immune responses in mice. Vaccine 2008, 26, 460–468. [Google Scholar] [CrossRef]
- Lopman, B.A.; Brown, D.W.; Koopmans, M. Human caliciviruses in Europe. J. Clin. Virol. 2002, 24, 137–160. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group of Experiment | No. of Adult Female Minks (%) | No. of Litters (%) | No. of Samples Tested by PCR (%) |
|---|---|---|---|
| Sham 1 | 5 (30) | 16 (18) | 87 (21) |
| CPΔN 2 -immunized | 6 (35) | 31 (35) | 146 (35) |
| CPΔC 3 -immunized | 6 (35) | 42 (47) | 181 (44) |
| Total | 17 (100) | 89 (100) | 414 (100) |
| Group of Experiment | No. of Litters (%) | Evaluated Clinical Signs in Litters | ||||
|---|---|---|---|---|---|---|
| Greasy Puppies (%) | Poor Body Condition (%) | Skin Redness (%) | Severe Diarrhea (%) | Black Nails (%) | ||
| Sham 1 | 16 (18) | 11 (68.7) | 10 (62.5) | 16 (100) | 9 (56.2) | 15 (90.4) |
| CPΔN 2 -immunized | 31 (35) | 12 (38.7) | 15 (48.4) | 18 (58.1) | 12 (38.7) | 13 (42) |
| CPΔC 3 -immunized | 42 (47) | 19 (45.2) | 6 (14.3) | 33 (78.6) | 16 (38.1) | 9 (21.4) |
| Group of Experiment | Ratio of Seropositivity (%) | |||
|---|---|---|---|---|
| 3 Weeks after First Immunization | 3 Weeks after Second Immunization (Booster) | |||
| Mice | Mink | Mice | Mink | |
| Sham 1 | 0/8 (0) | 0/5 (0) | 0/8 (0) | 0/5 (0) |
| CP 2 -immunized | 0/8 (0) | NT 3 | 5/8 (62.5) | NT |
| CPΔN 4 -immunized | 0/8 (0) | 5/6 (83.3) | 8/8 (100) | 6/6 (100) |
| CPΔC 5 -immunized | 0/8 (0) | 5/6 (83.3) | 7/8 (87.5) | 6/6 (100) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bidokhti, M.R.M.; Ullman, K.; Hammer, A.S.; Jensen, T.H.; Chriél, M.; Byrareddy, S.N.; Baule, C. Immunogenicity and Efficacy Evaluation of Subunit Astrovirus Vaccines. Vaccines 2019, 7, 79. https://doi.org/10.3390/vaccines7030079
Bidokhti MRM, Ullman K, Hammer AS, Jensen TH, Chriél M, Byrareddy SN, Baule C. Immunogenicity and Efficacy Evaluation of Subunit Astrovirus Vaccines. Vaccines. 2019; 7(3):79. https://doi.org/10.3390/vaccines7030079
Chicago/Turabian StyleBidokhti, Mehdi R.M., Karin Ullman, Anne Sofie Hammer, Trine Hammer Jensen, Mariann Chriél, Siddappa N. Byrareddy, and Claudia Baule. 2019. "Immunogenicity and Efficacy Evaluation of Subunit Astrovirus Vaccines" Vaccines 7, no. 3: 79. https://doi.org/10.3390/vaccines7030079
APA StyleBidokhti, M. R. M., Ullman, K., Hammer, A. S., Jensen, T. H., Chriél, M., Byrareddy, S. N., & Baule, C. (2019). Immunogenicity and Efficacy Evaluation of Subunit Astrovirus Vaccines. Vaccines, 7(3), 79. https://doi.org/10.3390/vaccines7030079