Advancing Toward HIV-1 Vaccine Efficacy through the Intersections of Immune Correlates

Abstract
:1. Introduction
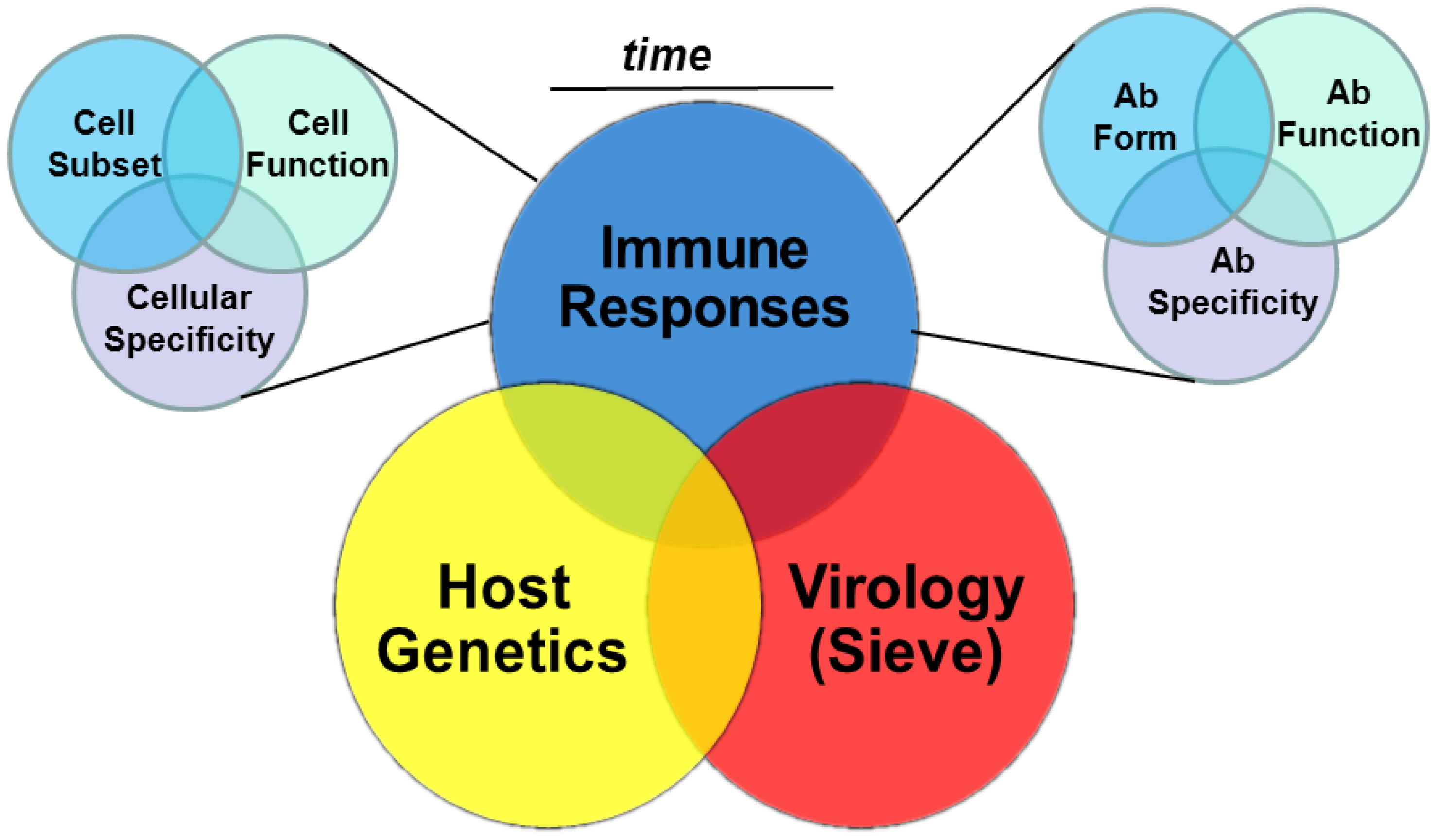
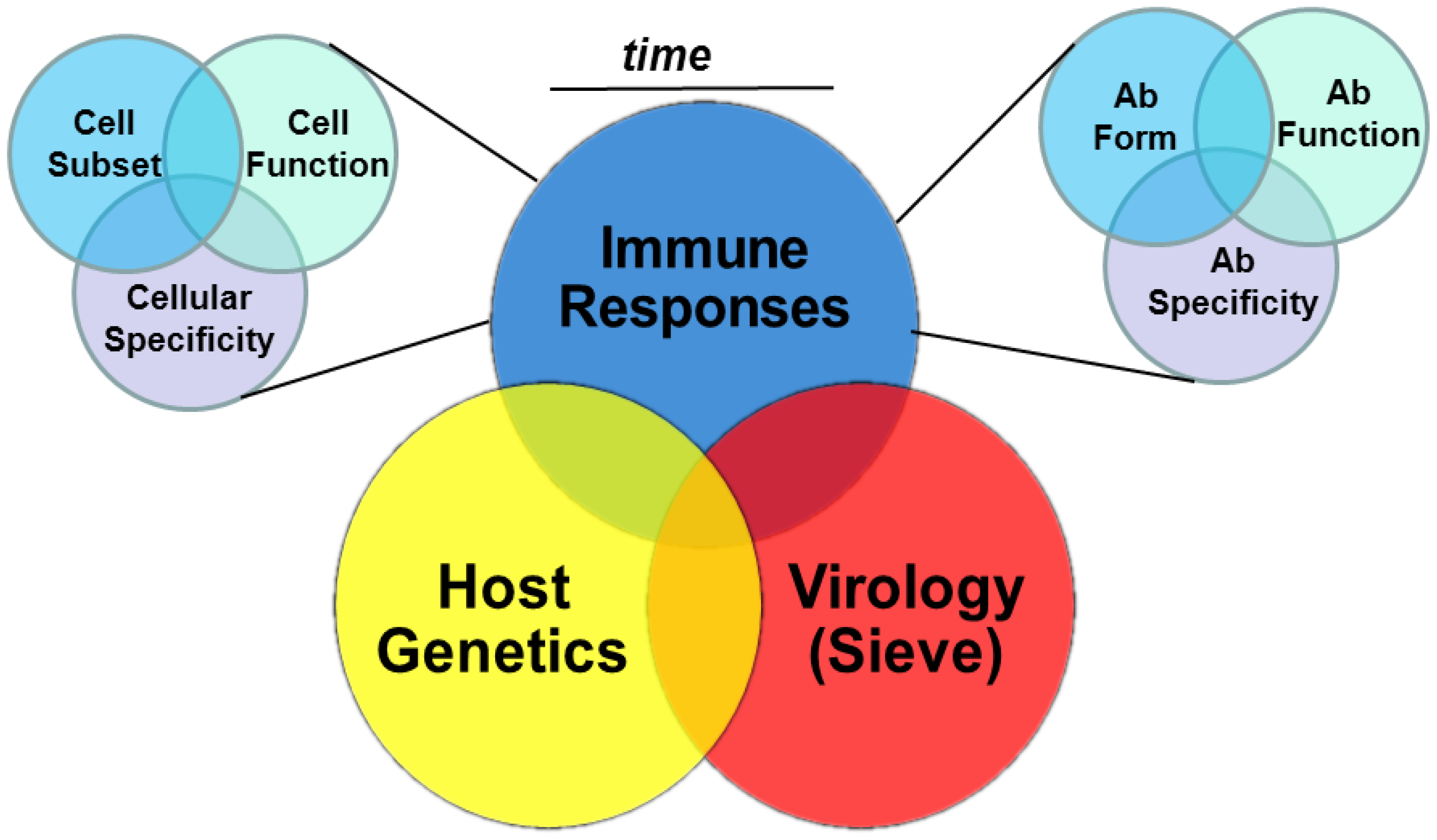
Insights from Multiple Measurements: Immune Responses, Virus Sieve Analysis and Host Genetics
2. HIV-1 Vaccine Efficacy Trials

{kind=link}
| HIV-1 Efficacy Trial | Vaccine Strategy | Risk Population | Geographic Location | Vaccine Efficacy Outcome a | Correlates of HIV-1 Risk/Incidence | Evidence of Immune Pressure (Virus Sieve) |
|---|---|---|---|---|---|---|
| 1. VAX003 [2] (Phase III) | Protein/Alum (CRF01_AE/Clade B Env) | Injection Drug Users (IDU) | Thailand | No Efficacy | No [2] | No [20] |
| 2. VAX004 [1] (Phase III) | Protein/Alum (Clade B Envs) | MSM b/High Risk Women | USA | No Efficacy | Yes [7,8,9] | No [21,22] |
| 3. Step HVTN502 [3] (Phase IIb) | Ad5 Vector (Clade B Gag/Pol Nef) | MSM/ High Risk Heterosexual Men and Women | North and South America, Australia, Caribbean | No Efficacy (Efficacy futility determined at first interim analysis after full enrollment) | Yes [10,11] | Yes [23] |
| 4. Phambili HVTN503 [4] (Phase IIb) | Ad5 Vector (Clade B Gag/Pol/Nef) | Heterosexual men and women | South Africa | No Efficacy (Vaccinations discontinued: early unblinding due to Step results) | None reported | None reported |
| 5. RV144 [5] (Phase III) | ALVAC vector (Clade B Gag/Pro + CRF01_A/E Env) + Protein/Alum (CRF01_AE/Clade B Env) | Community c | Thailand | 31.2% Efficacy | Yes [12,13,14,15] | Yes [24,25] |
| 6.HVTN505 [6] (Phase IIb) | DNA + Ad5 Vector (Clade A,B,C Env, Clade B Gag/Pol) | MSM and TG d (Ad5 seronegative, Circumcised) | USA | No Efficacy (Efficacy futility determined at first interim analysis after full enrollment ) | StudiesIn Progress | StudiesIn Progress |
3. Cellular Immunity and HIV-1 Vaccine Efficacy Trials
HVTN 502 (Step) and HVTN 503 (Phambili); Host Genetics and Virus Sieve Analyses
4. Humoral Immunity and HIV-1 Vaccine Efficacy Trials
4.1. VAX003 and VAX004: Immune Response and Host Genetics
4.2. RV144
4.2.1. Antigenicity of Immunogens
4.2.2. RV144: Intersection of Antibody Epitope Specificity, Antibody Function and Antibody Form
4.2.3. RV144 Secondary Analyses: Supporting the Primary Correlates and Further Hypotheses
4.2.4. RV144 Studies: Virus Sieve Analysis and B-Cell Repertoire
| Type of Analysis | Description of Outcome | Overlap or Intersections with Other Measurements |
|---|---|---|
| Follow-up studies to the RV144 immune correlate analysis both support and extend the original report by additional work involving the generation of monoclonal antibodies (mABs), virology (virus sieve analysis) and host genetics. | ||
| V2 Virus Sieve Analysis [24] | Vaccine efficacy was higher against viruses matching the vaccine at position 169 in Env (another signature at position 181 that mismatched the vaccine was also identified as unrelated to the V2 site). | To date, five follow-up studies support the identification of V1V2 IgG correlate [12], and the virus sieve analysis in the V2 loop are consistent with these findings. (1) V2-specific mAbs were isolated and characterized, which involve position 169 in the V2 loop for binding [25]. (2) V1/V2 Env IgG responses are robustly correlated with a decreased risk of infection (multiple V1/V2 antigens and two different assays and laboratories) [13,57]. (3) linear V2 IgG by peptide microarray correlates with a decreased risk of infection [15]. (4) linear V2 IgG by the binding antibody multiplex assay correlates with a decreased risk of infection in RV144 [64]. (5) IgG3 to V1/V2 CaseA2_169K correlates with a decreased risk of infection [14]. |
| V3 Virus Sieve Analysis [65] | Vaccine efficacy was higher against viruses matching the vaccine in the Env V3 loop. | V3 IgG correlates with a decreased risk of infection [15]. |
| Host Genetics: Human Leukocyte Antigen (HLA) Class I [66] | There was an association of the HLA Class I allele (A* 02) with HIV-1 infection risk and the two identified antibody correlates in RV144 vaccines. | This analysis of host genetics for HLA Class I alleles intersects with both the V1/V2 IgG response and the Env IgA response. (1) Vaccine efficacy for viruses with a lysine at position 169 was higher in those with the HLA A* 02 allele; (2) there was a direct correlation between plasma C1 Env IgA response and infection rate in the A* 02 (−) subgroup, but not the A* 02 (+) subgroup. |
| Host Genetics/HLA Class II [67] | There was an association of the HLA Class II alleles with HIV-1 infection risk and the plasma Env IgA correlate in RV144. | HLA II allele, DQB1* 06, had a significant interaction with the plasma Env IgA responses, such that DQB1* 06 had a significant effect on HIV-1 infection among the high IgA responders. |
| Host Genetics/FcγRIIC [68] | There was an association of FcγRIIC with vaccine efficacy. | This analysis of host genetics for HLA Class I alleles intersects with both V2 sieve analysis and the Env IgA response. (1) Vaccine efficacy for viruses with a lysine at position 169 was higher in those with at least one FcγRIIC-118l allele; (2) direct correlation of plasma Env IgA with infection risk was only when FcγRIIC-118l was present. |
| RV144 Immune Measurement | Description of Outcome | Overlap or Intersections with other Measurements |
|---|---|---|
| Interaction Models of Primary Correlate Analysis The six primary immune measurements evaluated in the RV144 correlate analysis were chosen in part based on each one representing distinct immunological space. Two of these measurements (V1/V2 IgG and Env IgA) significantly correlated with the risk of HIV-1 infection as reported in Haynes et al [12]. Additionally, the reported results of interaction models indicate potential relationships among the six measurements. Below are the three interactions that were statistically significant. | ||
| 1. Interaction of Plasma Env IgA and IgG Env Avidity [12] | In the presence of low plasma Env IgA, avidity to vaccine strain Env correlated with a decreased risk of infection. | HIV-1-Specific Plasma IgA Env Breadth Score. IgG Env avidity. |
| 2. Interaction of Plasma Env IgA and antibody-dependent cellular cytotoxicity (ADCC) [12,54,60] | In the presence of low plasma Env IgA, ADCC correlated with a decreased risk of infection. | HIV-1-Specific Plasma IgA Env Breadth Score antibody-dependent cellular cytotoxicity (ADCC). Follow-up mechanistic studies demonstrated that conformational C1-specific IgA antibody can block IgG-mediated ADCC (NK effectors) [54] . |
| 3. Interaction of Plasma Env IgA and NAb [12] | In the presence of low plasma Env IgA, Nab correlated with a decreased risk of infection. | HIV-1-Specific Plasma IgA Env Breadth Score. Neutralizing Antibody (NAb) Score. |
| Secondary Immune Measurements A series of immune measurements, in addition to the preselected primary six variables, were evaluated for correlations with the risk of HIV-1 infection. Of all the secondary immune variables, eight of these had a p-value of < 0.05. Three of these are related to the direct correlation with plasma Env IgA; two are related to the indirect correlation with Env IgG responses, and three are related to cellular responses (i.e., peripheral blood mononuclear cell (PBMC) derived cytokines). | ||
| 1. IgA CRF01 AE. C1 peptide [12,54] | IgA binding to this C1 peptide significantly correlated with increased risk of infection among vaccinees (decreased vaccine efficacy). | Follow-up analysis of the C1 Env IgA/IgG ratio significantly correlated with decreased vaccine efficacy [54]. |
| 2. IgA A.con.env03 gp140 CF [12,54] | IgA binding to this individual Env protein correlated with increased risk of infection among vaccinees (decreased vaccine efficacy). | This measurement was part of the Plasma Env IgA Breadth Score, which was a primary variable in RV144. Follow-up analysis of the Env IgA/IgG ratio significantly correlated with decreased vaccine efficacy [54]. |
| 3. IgA A.OOMSA gp140 CF [12,54] | IgA binding to this individual Env protein correlated with increased risk of infection among vaccinees (decreased vaccine efficacy). | This measurement was part of the Plasma Env IgA Breadth Score, which was a primary variable in RV144. |
| Secondary Immune Measurements A series of immune measurements, in addition to the preselected primary six variables, were evaluated for correlations with the risk of HIV-1 infection. Of all the secondary immune variables, eight of these had a p-value of < 0.05. Three of these are related to the direct correlation with plasma Env IgA; two are related to the indirect correlation with Env IgG responses, and three are related to cellular responses (i.e., PBMC derived cytokines). | ||
| 4. IgG A244gD-293T gp120 [12] | IgG binding to vaccine strain Env correlated with a decreased vaccine efficacy. | This supports IgA/Avidity interaction model, where avidity includes binding to A244 Env. Follow-up analysis of the Env IgA/IgG ratio significantly correlated with decreased vaccine efficacy [54]. |
| 5. Composite V2 microarray (hotspot) [12,15]. | IgG binding to the V2 composite score from the peptide microarray correlated with a decreased risk of infection. | This supports the V1/V2 IgG primary immune measurement that correlated with a decreased risk of infection [12], the V2 virus sieve analysis [24] and the follow-up analysis to examine linear V2 by the binding antibody multiplex assay. |
| 6. PBMC Luminex Score [12] | The cytokine score from Env stimulated PBMC supernatants correlated with a decreased risk of HIV-1 infection. | Individual cytokines (IL-10 and IL-13), as indicated below, correlated with a decreased risk of HIV-1 infection. |
| 7. IL-10 [12] and 8. IL-13 [12] | These individual cytokine responses from Env stimulated PBMC supernatants correlated with a decreased risk of HIV-1 infection. | Single cell transcriptomics demonstrated that RV144 vaccine-specific T-cells produced this cytokine [69]. |
4.2.5. Multiple Antibody Specificities and Function: Vaccine Efficacy
5. Time-Dependent Immune Measurements and Vaccine Efficacy
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Flynn, N.M.; Forthal, D.N.; Harro, C.D.; Judson, F.N.; Mayer, K.H.; Para, M.F.; rgp120 HIV Vaccine Study Group. Placebo-controlled phase 3 trial of a recombinant glycoprotein 120 vaccine to prevent HIV-1 infection. J. Infect. Dis. 2005, 191, 654–665. [Google Scholar] [CrossRef]
- Pitisuttithum, P.; Gilbert, P.; Gurwith, M.; Heyward, W.; Martin, M.; van Griensven, F.; Hu, D.; Tappero, J.W.; Choopanya, K.; Bangkok Vaccine Evaluation Group. Randomized, double-blind, placebo-controlled efficacy trial of a bivalent recombinant glycoprotein 120 HIV-1 vaccine among injection drug users in Bangkok, Thailand. J. Infect. Dis. 2006, 194, 1661–1671. [Google Scholar] [CrossRef]
- Buchbinder, S.P.; Mehrotra, D.V.; Duerr, A.; Fitzgerald, D.W.; Mogg, R.; Li, D.; Gilbert, P.B.; Lama, J.R.; Marmor, M.; del Rio, C.; et al. Efficacy assessment of a cell-mediated immunity HIV-1 vaccine (the step study): A double-blind, randomised, placebo-controlled, test-of-concept trial. Lancet 2008, 372, 1881–1893. [Google Scholar] [CrossRef]
- Gray, G.E.; Allen, M.; Moodie, Z.; Churchyard, G.; Bekker, L.G.; Nchabeleng, M.; Mlisana, K.; Metch, B.; de Bruyn, G.; Latka, M.H.; et al. Safety and efficacy of the HVTN 503/Phambili study of a clade-b-based HIV-1 vaccine in South Africa: A double-blind, randomised, placebo-controlled test-of-concept phase 2b study. Lancet Infect. Dis. 2011, 11, 507–515. [Google Scholar] [CrossRef]
- Rerks-Ngarm, S.; Pitisuttithum, P.; Nitayaphan, S.; Kaewkungwal, J.; Chiu, J.; Paris, R.; Premsri, N.; Namwat, C.; de Souza, M.; Adams, E.; et al. Vaccination with ALVAC and AIDSVAX to prevent HIV-1 infection in Thailand. N. Engl. J. Med. 2009, 361, 2209–2220. [Google Scholar] [CrossRef]
- Hammer, S.M.; Sobieszczyk, M.E.; Janes, H.; Karuna, S.T.; Mulligan, M.J.; Grove, D.; Koblin, B.A.; Buchbinder, S.P.; Keefer, M.C.; Tomaras, G.D.; et al. Efficacy trial of a DNA/rAd5 HIV-1 preventive vaccine. N. Engl. J. Med. 2013, 369, 2083–2092. [Google Scholar]
- Gilbert, P.B.; Peterson, M.L.; Follmann, D.; Hudgens, M.G.; Francis, D.P.; Gurwith, M.; Heyward, W.L.; Jobes, D.V.; Popovic, V.; Self, S.G.; et al. Correlation between immunologic responses to a recombinant glycoprotein 120 vaccine and incidence of HIV-1 infection in a phase 3 HIV-1 preventive vaccine trial. J. Infect. Dis. 2005, 191, 666–677. [Google Scholar] [CrossRef]
- Forthal, D.N.; Gilbert, P.B.; Landucci, G.; Phan, T. Recombinant gp120 vaccine-induced antibodies inhibit clinical strains of HIV-1 in the presence of Fc receptor-bearing effector cells and correlate inversely with HIV infection rate. J. Immunol. 2007, 178, 6596–6603. [Google Scholar]
- Forthal, D.N.; Gabriel, E.E.; Wang, A.; Landucci, G.; Phan, T.B. Association of fcgamma receptor IIIa genotype with the rate of HIV infection after gp120 vaccination. Blood 2012, 120, 2836–2842. [Google Scholar] [CrossRef]
- Huang, Y.; Gilbert, P.B. Comparing biomarkers as principal surrogate endpoints. Biometrics 2011, 67, 1442–1451. [Google Scholar] [CrossRef]
- Huang, Y.; Duerr, A.; Moodie, Z.; Frahm, N.; DeRosa, S.C.; McElrath, J.; Gilbert, P. Immune-correlates analysis of the step HIV vaccine efficacy trial, a post-hoc analysis of HIV-specific and non-specific cellular immune responses. In AIDS Vaccine, Barcelona, Spain, 7–10 October 2013.
- Haynes, B.F.; Gilbert, P.B.; McElrath, M.J.; Zolla-Pazner, S.; Tomaras, G.D.; Alam, S.M.; Evans, D.T.; Montefiori, D.C.; Karnasuta, C.; Sutthent, R.; et al. Immune-correlates analysis of an HIV-1 vaccine efficacy trial. N. Engl. J. Med. 2012, 366, 1275–1286. [Google Scholar] [CrossRef]
- Zolla-Pazner, S.; DeCamp, A.; Gilbert, P.; Williams, C.; Yates, N.L.; Williams, W.; Howington, R.; Fong, Y.; Morris, D.E.; Soderberg, K.E.; et al. Vaccine-induced IgG antibodies to V1V2 regions of multiple HIV-1 subtypes correlate with decreased risk of HIV-1 infection. PLoS One 2013, in press. [Google Scholar]
- Yates, N.L.; Liao, H.-X.; Fong, Y.; DeCamp, A.; Vandergrift, N.; Williams, W.T.; Alam, M.; Ferrari, G.; Yang, Z.Y.; Seaton, K.; et al. Spectrum of Vaccine-Elicited Humoral Responses. In HIV-1 Vaccines, Keystone Symposia, Keystone, CO, 10–15 February 2013.
- Gottardo, R.; Bailer, R.T.; Korber, B.T.; Gnanakaran, S.; Phillips, J.; Shen, X.; Tomaras, G.D.; Turk, E.; Imholte, G.; Eckler, L.; et al. Plasma IgG to linear epitopes in the V2 and V3 regions of HIV-1 gp120 correlate with a reduced risk of infection in the RV144 vaccine efficacy trial. PLoS One 2013, 8, e75665. [Google Scholar]
- Rolland, M.; Gilbert, P. Evaluating immune correlates in HIV type 1 vaccine efficacy trials: What RV144 may provide. AIDS Res. Hum. Retroviruses 2012, 28, 400–404. [Google Scholar] [CrossRef]
- Qin, L.; Gilbert, P.B.; Corey, L.; McElrath, M.J.; Self, S.G. A framework for assessing immunological correlates of protection in vaccine trials. J. Infect. Dis. 2007, 196, 1304–1312. [Google Scholar] [CrossRef]
- Plotkin, S.A. Vaccines: Correlates of vaccine-induced immunity. Clin. Infect. Dis. 2008, 47, 401–409. [Google Scholar] [CrossRef]
- Plotkin, S.A.; Gilbert, P.B. Nomenclature for immune correlates of protection after vaccination. Clin. Infect. Dis. 2012, 54, 1615–1617. [Google Scholar] [CrossRef]
- Gilbert, P.; Vaccine and Infectious Disease and Public Health Sciences Divisions, Fred Hutchinson Cancer Research Center, and the Department of Biostatistics, University of Washington, Seattle, WA, USA. Unpublished observations. 2013.
- Gilbert, P.B.; McKeague, I.W.; Sun, Y. The 2-sample problem for failure rates depending on a continuous mark: An application to vaccine efficacy. Biostatistics 2008, 9, 263–276. [Google Scholar] [CrossRef]
- Gilbert, P.B.; Wu, C.; Jobes, D.V. Genome scanning tests for comparing amino acid sequences between groups. Biometrics 2008, 64, 198–207. [Google Scholar] [CrossRef]
- Rolland, M.; Tovanabutra, S.; deCamp, A.C.; Frahm, N.; Gilbert, P.B.; Sanders-Buell, E.; Heath, L.; Magaret, C.A.; Bose, M.; Bradfield, A.; et al. Genetic impact of vaccination on breakthrough HIV-1 sequences from the step trial. Nat. Med. 2011, 17, 366–371. [Google Scholar] [CrossRef]
- Rolland, M.; Edlefsen, P.T.; Larsen, B.B.; Tovanabutra, S.; Sanders-Buell, E.; Hertz, T.; deCamp, A.C.; Carrico, C.; Menis, S.; Magaret, C.A.; et al. Increased HIV-1 vaccine efficacy against viruses with genetic signatures in Env V2. Nature 2012, 490, 417–420. [Google Scholar] [CrossRef]
- Liao, H.X.; Bonsignori, M.; Alam, S.M.; McLellan, J.S.; Tomaras, G.D.; Moody, M.A.; Kozink, D.M.; Hwang, K.K.; Chen, X.; Tsao, C.Y.; et al. Vaccine induction of antibodies against a structurally heterogeneous site of immune pressure within HIV-1 envelope protein variable regions 1 and 2. Immunity 2013, 38, 176–186. [Google Scholar]
- Koup, R.A.; Safrit, J.T.; Cao, Y.; Andrews, C.A.; McLeod, G.; Borkowsky, W.; Farthing, C.; Ho, D.D. Temporal association of cellular immune responses with the initial control of viremia in primary human immunodeficiency virus type 1 syndrome. J. Virol. 1994, 68, 4650–4655. [Google Scholar]
- Borrow, P.; Lewicki, H.; Wei, X.; Horwitz, M.S.; Peffer, N.; Meyers, H.; Nelson, J.A.; Gairin, J.E.; Hahn, B.H.; Oldstone, M.B.; et al. Antiviral pressure exerted by HIV-1-specific cytotoxic T lymphocytes (CTLs) during primary infection demonstrated by rapid selection of ctl escape virus. Nat. Med. 1997, 3, 205–211. [Google Scholar] [CrossRef]
- Goonetilleke, N.; Liu, M.K.; Salazar-Gonzalez, J.F.; Ferrari, G.; Giorgi, E.; Ganusov, V.V.; Keele, B.F.; Learn, G.H.; Turnbull, E.L.; Salazar, M.G.; et al. The first T cell response to transmitted/founder virus contributes to the control of acute viremia in HIV-1 infection. J. Exp. Med. 2009, 206, 1253–1272. [Google Scholar] [CrossRef]
- Ferrari, G.; Korber, B.; Goonetilleke, N.; Liu, M.K.; Turnbull, E.L.; Salazar-Gonzalez, J.F.; Hawkins, N.; Self, S.; Watson, S.; Betts, M.R.; et al. Relationship between functional profile of HIV-1 specific CD8 T cells and epitope variability with the selection of escape mutants in acute HIV-1 infection. PLoS Pathog. 2011, 7, e1001273. [Google Scholar] [CrossRef]
- Freel, S.A.; Picking, R.A.; Ferrari, G.; Ding, H.; Ochsenbauer, C.; Kappes, J.C.; Kirchherr, J.L.; Soderberg, K.A.; Weinhold, K.J.; Cunningham, C.K.; et al. Initial HIV-1 antigen-specific CD8+ T cells in acute HIV-1 infection inhibit transmitted/founder virus replication. J. Virol. 2012, 86, 6835–6846. [Google Scholar] [CrossRef]
- McMichael, A.J.; Borrow, P.; Tomaras, G.D.; Goonetilleke, N.; Haynes, B.F. The immune response during acute HIV-1 infection: Clues for vaccine development. Nat. Rev. 2010, 10, 11–23. [Google Scholar]
- Betts, M.R.; Nason, M.C.; West, S.M.; De Rosa, S.C.; Migueles, S.A.; Abraham, J.; Lederman, M.M.; Benito, J.M.; Goepfert, P.A.; Connors, M.; et al. HIV nonprogressors preferentially maintain highly functional HIV-specific CD8+ T cells. Blood 2006, 107, 4781–4789. [Google Scholar] [CrossRef]
- Migueles, S.A.; Connors, M. Long-term nonprogressive disease among untreated HIV-infected individuals: Clinical implications of understanding immune control of HIV. JAMA 2010, 304, 194–201. [Google Scholar] [CrossRef]
- Freel, S.A.; Lamoreaux, L.; Chattopadhyay, P.K.; Saunders, K.; Zarkowsky, D.; Overman, R.G.; Ochsenbauer, C.; Edmonds, T.G.; Kappes, J.C.; Cunningham, C.K.; et al. Phenotypic and functional profile of HIV-inhibitory CD8 T cells elicited by natural infection and heterologous prime/boost vaccination. J. Virol. 2010, 84, 4998–5006. [Google Scholar]
- Freel, S.A.; Saunders, K.O.; Tomaras, G.D. CD8(+)T-cell-mediated control of HIV-1 and SIV infection. Immunol. Res. 2011, 49, 135–146. [Google Scholar] [CrossRef]
- Walker, B.D.; Yu, X.G. Unravelling the mechanisms of durable control of HIV-1. Nat. Rev. Immunol. 2013, 13, 487–498. [Google Scholar] [CrossRef]
- Saez-Cirion, A.; Pancino, G. HIV controllers: A genetically determined or inducible phenotype? Immunol. Rev. 2013, 254, 281–294. [Google Scholar] [CrossRef]
- Hansen, S.G.; Sacha, J.B.; Hughes, C.M.; Ford, J.C.; Burwitz, B.J.; Scholz, I.; Gilbride, R.M.; Lewis, M.S.; Gilliam, A.N.; Ventura, A.B.; et al. Cytomegalovirus vectors violate CD8+ T cell epitope recognition paradigms. Science 2013, 340, 1237874. [Google Scholar]
- Hansen, S.G.; Ford, J.C.; Lewis, M.S.; Ventura, A.B.; Hughes, C.M.; Coyne-Johnson, L.; Whizin, N.; Oswald, K.; Shoemaker, R.; Swanson, T.; et al. Profound early control of highly pathogenic siv by an effector memory T-cell vaccine. Nature 2011, 473, 523–527. [Google Scholar]
- Hansen, S.G.; Vieville, C.; Whizin, N.; Coyne-Johnson, L.; Siess, D.C.; Drummond, D.D.; Legasse, A.W.; Axthelm, M.K.; Oswald, K.; Trubey, C.M.; et al. Effector memory T cell responses are associated with protection of rhesus monkeys from mucosal simian immunodeficiency virus challenge. Nat. Med. 2009, 15, 293–299. [Google Scholar] [CrossRef]
- Fukazawa, Y.; Park, H.; Cameron, M.J.; Lefebvre, F.; Lum, R.; Coombes, N.; Mahyari, E.; Hagen, S.I.; Bae, J.Y.; Reyes, M.D., 3rd; et al. Lymph node T cell responses predict the efficacy of live attenuated siv vaccines. Nat. Med. 2012, 18, 1673–1681. [Google Scholar] [CrossRef]
- Reardon, S. HIV vaccine raised infection risk. Nature 2013. [Google Scholar] [CrossRef]
- Frahm, N.; DeCamp, A.C.; Friedrich, D.P.; Carter, D.K.; Defawe, O.D.; Kublin, J.G.; Casimiro, D.R.; Duerr, A.; Robertson, M.N.; Buchbinder, S.P.; et al. Human adenovirus-specific T cells modulate HIV-specific T cell responses to an ad5-vectored HIV-1 vaccine. J. Clin. Invest. 2012, 122, 359–367. [Google Scholar] [CrossRef]
- McElrath, M.J.; De Rosa, S.C.; Moodie, Z.; Dubey, S.; Kierstead, L.; Janes, H.; Defawe, O.D.; Carter, D.K.; Hural, J.; Akondy, R.; et al. HIV-1 vaccine-induced immunity in the test-of-concept step study: A case-cohort analysis. Lancet 2008, 372, 1894–1905. [Google Scholar] [CrossRef]
- Gray, G.; Buchbinder, S.; Duerr, A. Overview of step and phambili trial results: Two phase IIb test-of-concept studies investigating the efficacy of mrk adenovirus type 5 gag/pol/nef subtype b HIV vaccine. Curr. Opin. HIV AIDS 2010, 5, 357–361. [Google Scholar]
- Duerr, A.; Huang, Y.; Buchbinder, S.; Coombs, R.W.; Sanchez, J.; del Rio, C.; Casapia, M.; Santiago, S.; Gilbert, P.; Corey, L.; et al. Extended follow-up confirms early vaccine-enhanced risk of HIV acquisition and demonstrates waning effect over time among participants in a randomized trial of recombinant adenovirus HIV vaccine (step study). J. Infect. Dis. 2012, 206, 258–266. [Google Scholar] [CrossRef]
- Hertz, T.; Ahmed, H.; Friedrich, D.P.; Casimiro, D.R.; Self, S.G.; Corey, L.; McElrath, M.J.; Buchbinder, S.; Horton, H.; Frahm, N.; et al. HIV-1 vaccine-induced T-cell responses cluster in epitope hotspots that differ from those induced in natural infection with HIV-1. PLoS Pathog. 2013, 9, e1003404. [Google Scholar] [CrossRef]
- Fitzgerald, D.W.; Janes, H.; Robertson, M.; Coombs, R.; Frank, I.; Gilbert, P.; Loufty, M.; Mehrotra, D.; Duerr, A.; Step Study Protocol Team. An ad5-vectored HIV-1 vaccine elicits cell-mediated immunity but does not affect disease progression in HIV-1-infected male subjects: Results from a randomized placebo-controlled trial (the step study). J. Infect. Dis. 2011, 203, 765–772. [Google Scholar] [CrossRef]
- Migueles, S.A.; Rood, J.E.; Berkley, A.M.; Guo, T.; Mendoza, D.; Patamawenu, A.; Hallahan, C.W.; Cogliano, N.A.; Frahm, N.; Duerr, A.; et al. Trivalent adenovirus type 5 HIV recombinant vaccine primes for modest cytotoxic capacity that is greatest in humans with protective HLA class I alleles. PLoS Pathog. 2011, 7, e1002002. [Google Scholar] [CrossRef]
- Janes, H.; Friedrich, D.P.; Krambrink, A.; Smith, R.J.; Kallas, E.; Horton, H.; Casimiro, D.R.; Carrington, M.; Geraghty, D.; Gilbert, P.; et al. Vaccine-induced Gag-specific T cells are associated with reduced viremia after HIV infection. J. Infect. Dis. 2013. [Google Scholar] [CrossRef]
- Janes, H.; Frahm, N.; DeCamp, A.; Rolland, M.; Gabriel, E.; Wolfson, J.; Hertz, T.; Kallas, E.; Goepfert, P.; Friedrich, D.P.; et al. MrkAd5 HIV-1 gag/pol/nef vaccine-induced T-cell responses inadequately predict distance of breakthrough HIV-1 sequences to the vaccine or viral load. PLoS One 2012, 7, e43396. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, P.; Wang, M.; Wrin, T.; Petropoulos, C.; Gurwith, M.; Sinangil, F.; D’Souza, P.; Rodriguez-Chavez, I.R.; DeCamp, A.; Giganti, M.; et al. Magnitude and breadth of a nonprotective neutralizing antibody response in an efficacy trial of a candidate HIV-1 gp120 vaccine. J. Infect. Dis. 2010, 202, 595–605. [Google Scholar] [CrossRef]
- Montefiori, D.C.; Karnasuta, C.; Huang, Y.; Ahmed, H.; Gilbert, P.; de Souza, M.S.; McLinden, R.; Tovanabutra, S.; Laurence-Chenine, A.; Sanders-Buell, E.; et al. Magnitude and breadth of the neutralizing antibody response in the RV144 and VAX003 HIV-1 vaccine efficacy trials. J. Infect. Dis. 2012, 206, 431–441. [Google Scholar] [CrossRef]
- Tomaras, G.D.; Ferrari, G.; Shen, X.; Alam, S.M.; Liao, H.X.; Pollara, J.; Bonsignori, M.; Moody, M.A.; Fong, Y.; Chen, X.; et al. Vaccine-induced plasma iga specific for the C1 region of the HIV-1 envelope blocks binding and effector function of IgG. Proc. Natl. Acad. Sci. USA 2013, 110, 9019–9024. [Google Scholar] [CrossRef]
- De Souza, M.S.; Ratto-Kim, S.; Chuenarom, W.; Schuetz, A.; Chantakulkij, S.; Nuntapinit, B.; Valencia-Micolta, A.; Thelian, D.; Nitayaphan, S.; Pitisuttithum, P.; et al. The Thai phase III trial (RV144) vaccine regimen induces T cell responses that preferentially target epitopes within the V2 region of HIV-1 envelope. J. Immunol. 2012, 188, 5166–5176. [Google Scholar] [CrossRef]
- Alam, S.M.; Liao, H.X.; Tomaras, G.D.; Bonsignori, M.; Tsao, C.Y.; Hwang, K.K.; Chen, H.; Lloyd, K.E.; Bowman, C.; Sutherland, L.; et al. Antigenicity and immunogenicity of RV144 vaccine AIDSVAX clade E envelope immunogen is enhanced by a gp120 N-terminal deletion. J. Virol. 2013, 87, 1554–1568. [Google Scholar] [CrossRef]
- Zolla-Pazner, S.; deCamp, A.C.; Cardozo, T.; Karasavvas, N.; Gottardo, R.; Williams, C.; Morris, D.E.; Tomaras, G.; Rao, M.; Billings, E.; et al. Analysis of V2 antibody responses induced in vaccinees in the ALVAC/AIDSVAX HIV-1 vaccine efficacy trial. PloS One 2013, 8, e53629. [Google Scholar] [CrossRef]
- Karasavvas, N.; Billings, E.; Rao, M.; Williams, C.; Zolla-Pazner, S.; Bailer, R.T.; Koup, R.A.; Madnote, S.; Arworn, D.; Shen, X.; et al. The Thai phase III HIV type 1 vaccine trial (RV144) regimen induces antibodies that target conserved regions within the V2 loop of gp120. AIDS Res. Hum. Retroviruses 2012, 28, 1444–1457. [Google Scholar] [CrossRef]
- Ferrari, G.; Pollara, J.; Kozink, D.; Harms, T.; Drinker, M.; Freel, S.; Moody, M.A.; Alam, S.M.; Tomaras, G.D.; Ochsenbauer, C.; et al. An HIV-1 gp120 envelope human monoclonal antibody that recognizes a c1 conformational epitope mediates potent antibody-dependent cellular cytotoxicity (ADCC) activity and defines a common ADCC epitope in human HIV-1 serum. J. Virol. 2011, 85, 7029–7036. [Google Scholar] [CrossRef]
- Bonsignori, M.; Pollara, J.; Moody, M.A.; Alpert, M.D.; Chen, X.; Hwang, K.K.; Gilbert, P.B.; Huang, Y.; Gurley, T.C.; Kozink, D.M.; et al. Antibody-dependent cellular cytotoxicity-mediating antibodies from an HIV-1 vaccine efficacy trial target multiple epitopes and preferentially use the VH1 gene family. J. Virol. 2012, 86, 11521–11532. [Google Scholar] [CrossRef]
- Tomaras, G.D.; Yates, N.L.; Liu, P.; Qin, L.; Fouda, G.G.; Chavez, L.L.; Decamp, A.C.; Parks, R.J.; Ashley, V.C.; Lucas, J.T.; et al. Initial B-cell responses to transmitted human immunodeficiency virus type 1: Virion-binding immunoglobulin IgM and IgG antibodies followed by plasma anti-gp41 antibodies with ineffective control of initial viremia. J. Virol. 2008, 82, 12449–12463. [Google Scholar] [CrossRef]
- Haynes, B.F.; Korber, B.; Liao, H.; Tomaras, G.; Duke Human Vaccine Institute, Duke University, Durham, NC, USA and Los Alamos National Laboratory, Theoretical Division, Los Alamos, NM, USA. Unpublished observations. 2013.
- Liu, P.; Yates, N.L.; Shen, X.; Bonsignori, M.; Moody, M.A.; Liao, H.X.; Fong, Y.; Alam, S.M.; Overman, R.G.; Denny, T.; et al. Infectious virion capture by HIV-1 gp120-specific IgG from RV144 vaccinees. J. Virol. 2013, 87, 7828–7836. [Google Scholar] [CrossRef]
- Tomaras, G.; Shen, X.; Seaton, K.; Janes, H.; Grove, D.; DeCamp, A.; Fong, Y.; Liao, H.; Yang, Z.Y.; Xu, T.; et al. Vaccine induced antibody responses in HVTN 505, a phase IIb HIV-1 efficacy trial. In AIDS Vaccine, Barcelona, Spain, 7–10 October 2013.
- Rolland, M.; Edlefsen, P.T.; Gottardo, R.; Montefiori, D.; Zolla-Pazner, S.; Moody, M.A.; Liao, H.; Liu, P.; Tomaras, G.; Haynes, B.F.; et al. Genetic and immunological evidence for a role of Env-V3 antibodies in the RV144 trial. In AIDS Vaccine, Barcelona, Spain, 7–10 October 2013.
- Hertz, T.; Gartland, A.; Janes, H.; Li, S.S.; Fong, Y.; Tomaras, G.; Morris, D.E.; Geraghty, D.; Kijak, G.H.; Edlefsen, P.T.; et al. T-cell based sieve analysis ties HLA A*02 to vaccine efficacy and iga-c1 immune correlate in RV144 Thai trial. Retrovirology 2012, 9, O61. [Google Scholar] [CrossRef]
- Prentice, H.; Geraghty, D.; Tomaras, G.; Fong, Y.; Nelson, W.; Kijak, G.H.; Zolla-Pazner, S.; Nitayaphan, S.; Rerks-Ngarm, S.; Kaewkungwal, J.; et al. HLA class II genes interact with the immune correlates from the RV144 vaccine efficacy trial and impact HIV-1 acquisition. In AIDS Vaccine, Barcelona, Spain, 7–10 October 2013.
- Li, S.; Gilbert, P.; Tomaras, G.; Kijak, G.H.; Ferrari, G.; Thomas, R.; Zolla-Pazner, S.; Evans, D.T.; Li, Y.; Gottardo, R.; et al. Association of FcRIIc polymorphism with vaccine efficacy and correlates of HIV-1 infection risk in RV144. In AIDS Vaccine, Barcelona, Spain, 7–10 October 2013.
- Roederer, M. Single cell transcriptomics: Measuring quality and quantity. In HIV-1 Vaccines/B Cell Development and Function, Keystone Symposia, Keystone, CO, USA, 10–15 February 2013.
- Robb, M.L.; Rerks-Ngarm, S.; Nitayaphan, S.; Pitisuttithum, P.; Kaewkungwal, J.; Kunasol, P.; Khamboonruang, C.; Thongcharoen, P.; Morgan, P.; Benenson, M.; et al. Risk behaviour and time as covariates for efficacy of the HIV vaccine regimen ALVAC-HIV (vcp1521) and AIDSVAX B/E: A post-hoc analysis of the Thai phase 3 efficacy trial RV144. Lancet Infect. Dis. 2012, 12, 531–537. [Google Scholar] [CrossRef]
- Shea, P.R.; Shianna, K.V.; Carrington, M.; Goldstein, D.B. Host genetics of HIV acquisition and viral control. Annu. Rev. Med. 2013, 64, 203–217. [Google Scholar] [CrossRef]
- Carrington, M.; Walker, B.D. Immunogenetics of spontaneous control of HIV. Annu. Rev. Med. 2012, 63, 131–145. [Google Scholar]
- Siegrist, C. Vaccine Immunology. In Vaccines; Plotkin, S.A., Orenstein, W.A., Offit, P.A., Eds.; Elsevier: Philadelphia, PA, USA, 2008; pp. 17–36. [Google Scholar]
- Amanna, I.J.; Slifka, M.K. Contributions of humoral and cellular immunity to vaccine-induced protection in humans. Virology 2011, 411, 206–215. [Google Scholar] [CrossRef]
- Lambert, P.H.; Liu, M.; Siegrist, C.A. Can successful vaccines teach us how to induce efficient protective immune responses? Nat. Med. 2005, 11, S54–S62. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tomaras, G.D.; Haynes, B.F. Advancing Toward HIV-1 Vaccine Efficacy through the Intersections of Immune Correlates. Vaccines 2014, 2, 15-35. https://doi.org/10.3390/vaccines2010015
Tomaras GD, Haynes BF. Advancing Toward HIV-1 Vaccine Efficacy through the Intersections of Immune Correlates. Vaccines. 2014; 2(1):15-35. https://doi.org/10.3390/vaccines2010015
Chicago/Turabian StyleTomaras, Georgia D., and Barton F. Haynes. 2014. "Advancing Toward HIV-1 Vaccine Efficacy through the Intersections of Immune Correlates" Vaccines 2, no. 1: 15-35. https://doi.org/10.3390/vaccines2010015