Tumor-Associated Glycans and Immune Surveillance

Abstract
:1. Introduction
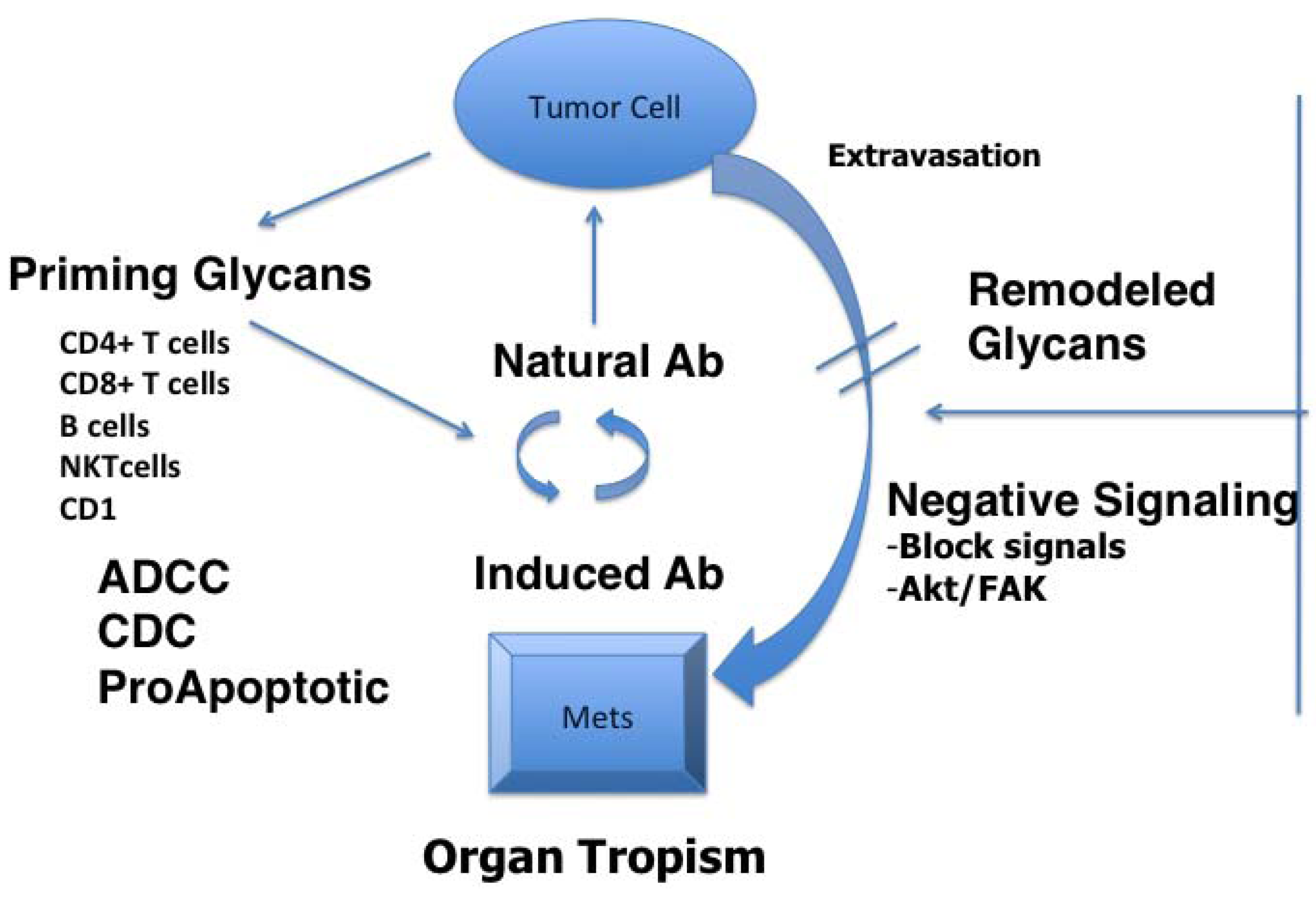
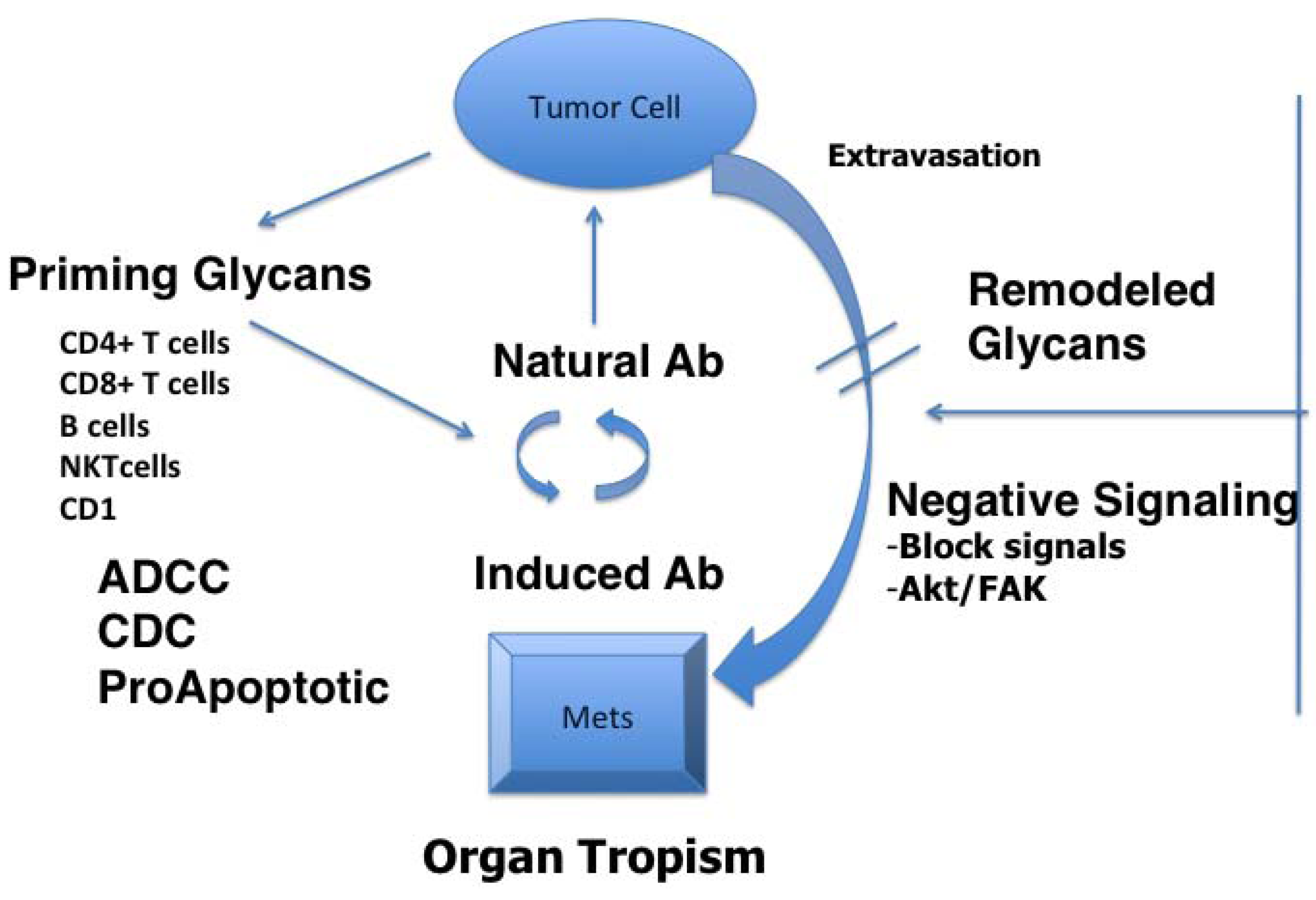
2. Glycans as Tumor Antigens

{kind=link}
| Carbohydrate antigens | Structure |
| Polysialic acid (PSA) | α2,8-/α2,9 NeuAc |
| Tn | GalNAcSer/Thr |
| sialyl Tn | Neu5Acα2-6GalNAcaSer/Thr |
| T antigen | Galβ1-3GalNAcαSer/Thr |
| Globo-H | Fucα1-2Galβ1-3GalNAcβ1-3Galα1-4Galβ1-4Glc |
| LeY | Fucα1-2Galβ1-4(Fucα1-3)GlcNAc-Galβ1-3(Fucα1-4)GlcNAc |
| SLeX | Neu5Acα2-3Galβ1-4(Fucα1-3)GlcNAc |
| SLeA | Neu5Acα2-3Galβ1-3(Fucα1-4)GlcNAc |
3. Innate Recognition of Glycans That Lends to Tumor Suppression
3.1. The Case for TACA-Directed Antibody-Mediated Tissue Rejection
3.2. The Case for Glycan-Directed T-Cell Mediated Tissue Rejection
3.3. The Case for NK Cell-Mediated Rejection
3.4. Remodeling Glycan Signatures
4. Augmenting Responses to TACA
4.1. Taking Advantage of Natural Antibodies
4.2. Bridging Humoral and Cellular Responses
5. Conclusions
Conflict of Interest
References
- Swann, J.B.; Smyth, M.J. Immune surveillance of tumors. J. Clin. Invest. 2007, 117, 1137–1146. [Google Scholar] [CrossRef]
- Burnet, F.M. The concept of immunological surveillance. Prog. Exp. Tumor Res. 1970, 13, 1–27. [Google Scholar]
- Soloski, M.J. Recognition of tumor cells by the innate immune system. Curr. Opin. Immunol. 2001, 13, 154–162. [Google Scholar] [CrossRef]
- Wang, E.; Monaco, A.; Monsurro, V.; Sabatino, M.; Pos, Z.; Uccellini, L.; Wang, J.; Worschech, A.; Stroncek, D.F.; Marincola, F.M. Antitumor vaccines, immunotherapy and the immunological constant of rejection. IDrugs 2009, 12, 297–301. [Google Scholar]
- Wang, E.; Worschech, A.; Marincola, F.M. The immunologic constant of rejection. Trends Immunol. 2008, 29, 256–262. [Google Scholar] [CrossRef]
- Houghton, A.N.; Guevara-Patiño, J.A. Immune recognition of self in immunity against cancer. J. Clin. Invest. 2004, 114, 468–471. [Google Scholar]
- Schreiber, T.H.; Raez, L.; Rosenblatt, J.D.; Podack, E.R. Tumor immunogenicity and responsiveness to cancer vaccine therapy: The state of the art. Semin. Immunol. 2010, 22, 105–112. [Google Scholar] [CrossRef]
- Hakomori, S. Tumor-associated carbohydrate antigens defining tumor malignancy: Basis for development of anti-cancer vaccines. Adv. Exp. Med. Biol. 2001, 491, 369–402. [Google Scholar] [CrossRef]
- Xu, Y.; Sette, A.; Sidney, J.; Gendler, S.J.; Franco, A. Tumor-associated carbohydrate antigens: A possible avenue for cancer prevention. Immunol. Cell Biol. 2005, 83, 440–448. [Google Scholar] [CrossRef]
- Ono, M.; Hakomori, S. Glycosylation defining cancer cell motility and invasiveness. Glycoconj. J. 2004, 20, 71–78. [Google Scholar]
- Sakamoto, S.; Kyprianou, N. Targeting anoikis resistance in prostate cancer metastasis. Mol. Aspects Med. 2010, 31, 205–214. [Google Scholar] [CrossRef]
- Zhong, X.; Rescorla, F.J. Cell surface adhesion molecules and adhesion-initiated signaling: Understanding of anoikis resistance mechanisms and therapeutic opportunities. Cell Signal. 2012, 24, 393–401. [Google Scholar] [CrossRef]
- Kornberg, L.J. Focal adhesion kinase and its potential involvement in tumor invasion and metastasis. Head Neck 1998, 20, 745–752. [Google Scholar] [CrossRef]
- Hauck, C.R.; Hsia, D.A.; Schlaepfer, D.D. The focal adhesion kinase—A regulator of cell migration and invasion. IUBMB Life 2002, 53, 115–119. [Google Scholar] [CrossRef]
- Sawai, H.; Okada, Y.; Funahashi, H.; Matsuo, Y.; Takahashi, H.; Takeyama, H.; Manabe, T. Activation of focal adhesion kinase enhances the adhesion and invasion of pancreatic cancer cells via extracellular signal-regulated kinase-1/2 signaling pathway activation. Mol. Cancer 2005, 4, 37. [Google Scholar] [CrossRef]
- Danker, K.; Reutter, W.; Semini, G. Glycosidated phospholipids: Uncoupling of signalling pathways at the plasma membrane. Br. J. Pharmacol. 2010, 160, 36–47. [Google Scholar] [CrossRef]
- Zhang, D.; Wei, J.; Wang, J.; Liu, S.; Wang, X.; Yan, Q. Difucosylated oligosaccharide Lewis Y is contained within integrin alphavbeta3 on RL95–2 cells and required for endometrial receptivity. Fertil. Steril. 2011, 95, 1446–1451. [Google Scholar]
- Valentino, L.A.; Ladisch, S. Tumor gangliosides enhance alpha2 beta1 integrin-dependent platelet activation. Biochim. Biophys. Acta 1996, 1316, 19–28. [Google Scholar] [CrossRef]
- Chen, Y.X.; Chen, X.W.; Li, C.G.; Yue, L.J.; Mai, H.R.; Wen, F.Q. Effect of tumor gangliosides on tyrosine phosphorylation of p125FAK in platelet adhesion to collagen. Oncol. Rep. 2013, 29, 343–348. [Google Scholar]
- Ohkawa, Y.; Miyazaki, S.; Hamamura, K.; Kambe, M.; Miyata, M.; Tajima, O.; Ohmi, Y.; Yamauchi, Y.; Furukawa, K. Ganglioside GD3 enhances adhesion signals and augments malignant properties of melanoma cells by recruiting integrins to glycolipid-enriched microdomains. J. Biol. Chem. 2010, 285, 27213–27223. [Google Scholar] [CrossRef]
- Aixinjueluo, W.; Furukawa, K.; Zhang, Q.; Hamamura, K.; Tokuda, N.; Yoshida, S.; Ueda, R. Mechanisms for the apoptosis of small cell lung cancer cells induced by anti-GD2 monoclonal antibodies: Roles of anoikis. J. Biol. Chem. 2005, 280, 29828–29836. [Google Scholar]
- Christiansen, D.; Vaughan, H.A.; Milland, J.; Dodge, N.; Mouhtouris, E.; Smyth, M.J.; Godfrey, D.I.; Sandrin, M.S. Antibody responses to glycolipid-borne carbohydrates require CD4+ T cells but not CD1 or NKT cells. Immunol. Cell Biol. 2011, 89, 502–510. [Google Scholar] [CrossRef]
- Zajonc, D.M.; Kronenberg, M. Carbohydrate specificity of the recognition of diverse glycolipids by natural killer T cells. Immunol. Rev. 2009, 230, 188–200. [Google Scholar] [CrossRef]
- Freire, T.; Zhang, X.; Deriaud, E.; Ganneau, C.; Vichier-Guerre, S.; Azria, E.; Launay, O.; Lo-Man, R.; Bay, S.; Leclerc, C. Glycosidic Tn-based vaccines targeting dermal dendritic cells favor germinal center B-cell development and potent antibody response in the absence of adjuvant. Blood 2010, 116, 3526–3536. [Google Scholar] [CrossRef]
- Ragupathi, G.; Liu, N.X.; Musselli, C.; Powell, S.; Lloyd, K.; Livingston, P.O. Antibodies against tumor cell glycolipids and proteins, but not mucins, mediate complement-dependent cytotoxicity. J. Immunol. 2005, 174, 5706–5712. [Google Scholar]
- Lavrsen, K.; Madsen, C.B.; Rasch, M.G.; Woetmann, A.; Odum, N.; Mandel, U.; Clausen, H.; Pedersen, A.E.; Wandall, H.H. Aberrantly glycosylated MUC1 is expressed on the surface of breast cancer cells and a target for antibody-dependent cell-mediated cytotoxicity. Glycoconj. J. 2013, 30, 227–236. [Google Scholar] [CrossRef]
- Vollmers, H.P.; Brandlein, S. Natural antibodies and cancer. N. Biotechnol. 2009, 25, 294–298. [Google Scholar] [CrossRef]
- Champion, E.; Andre, I.; Moulis, C.; Boutet, J.; Descroix, K.; Morel, S.; Monsan, P.; Mulard, L.A.; Remaud-Simeon, M. Design of alpha-transglucosidases of controlled specificity for programmed chemoenzymatic synthesis of antigenic oligosaccharides. J. Am. Chem. Soc. 2009, 131, 7379–7389. [Google Scholar] [CrossRef]
- Pon, R.A.; Biggs, N.J.; Jennings, H.J. Polysialic acid bioengineering of neuronal cells by N-acyl sialic acid precursor treatment. Glycobiology 2007, 17, 249–260. [Google Scholar]
- Bertozzi, C.R.; Kiessling, L.L. Chemical glycobiology. Science 2001, 291, 2357–2364. [Google Scholar] [CrossRef]
- Livingston, P.O.; Zhang, S.; Lloyd, K.O. Carbohydrate vaccines that induce antibodies against cancer. 1. Rationale. Cancer Immunol. Immunother. 1997, 45, 1–9. [Google Scholar] [CrossRef]
- Nakagoe, T.; Fukushima, K.; Tanaka, K.; Sawai, T.; Tsuji, T.; Jibiki, M.; Nanashima, A.; Yamaguchi, H.; Yasutake, T.; Ayabe, H.; Arisawa, K. Evaluation of sialyl Lewis(a), sialyl Lewis(x), and sialyl Tn antigens expression levels as predictors of recurrence after curative surgery in node-negative colorectal cancer patients. J. Exp. Clin. Cancer Res. 2002, 21, 107–113. [Google Scholar]
- Walz, G.; Aruffo, A.; Kolanus, W.; Bevilacqua, M.; Seed, B. Recognition by ELAM-1 of the sialyl-Lex determinant on myeloid and tumor cells. Science 1990, 250, 1132–1135. [Google Scholar]
- Gout, S.; Tremblay, P.L.; Huot, J. Selectins and selectin ligands in extravasation of cancer cells and organ selectivity of metastasis. Clin. Exp. Metastasis 2008, 25, 335–344. [Google Scholar] [CrossRef]
- Doekhie, F.S.; Morreau, H.; de Bock, G.H.; Speetjens, F.M.; Dekker-Ensink, N.G.; Putter, H.; van de Velde, C.J.; Tollenaar, R.A.; Kuppen, P.J. Sialyl Lewis X expression and lymphatic microvessel density in primary tumors of node-negative colorectal cancer patients predict disease recurrence. Cancer Microenviron. 2008, 1, 141–151. [Google Scholar] [CrossRef]
- Shimodaira, K.; Nakayama, J.; Nakamura, N.; Hasebe, O.; Katsuyama, T.; Fukuda, M. Carcinoma-associated expression of core 2 beta-1,6-N-acetylglucosaminyltransferase gene in human colorectal cancer: Role of O-glycans in tumor progression. Cancer Res. 1997, 57, 5201–5206. [Google Scholar]
- Dennis, J.W.; Pawling, J.; Cheung, P.; Partridge, E.; Demetriou, M. UDP-N-acetylglucosamine:alpha-6-D-mannoside beta1,6 N-acetylglucosaminyltransferase V (Mgat5) deficient mice. Biochim. Biophys. Acta 2002, 1573, 414–422. [Google Scholar]
- Yousefi, S.; Higgins, E.; Daoling, Z.; Pollex-Kruger, A.; Hindsgaul, O.; Dennis, J.W. Increased UDP-GlcNAc:Gal beta 1-3GaLNAc-R (GlcNAc to GaLNAc) beta-1, 6-N-acetylglucosaminyltransferase activity in metastatic murine tumor cell lines. Control of polylactosamine synthesis. J. Biol. Chem. 1991, 266, 1772–1782. [Google Scholar]
- Hagisawa, S.; Ohyama, C.; Takahashi, T.; Endoh, M.; Moriya, T.; Nakayama, J.; Arai, Y.; Fukuda, M. Expression of core 2 beta1,6-N-acetylglucosaminyltransferase facilitates prostate cancer progression. Glycobiology 2005, 15, 1016–1024. [Google Scholar] [CrossRef]
- Tsuboi, S.; Fukuda, M. Roles of O-linked oligosaccharides in immune responses. Bioessays 2001, 23, 46–53. [Google Scholar] [CrossRef]
- Fukuda, M. Roles of mucin-type O-glycans in cell adhesion. Biochim. Biophys. Acta 2002, 1573, 394–405. [Google Scholar] [CrossRef]
- Pang, P.C.; Tissot, B.; Drobnis, E.Z.; Morris, H.R.; Dell, A.; Clark, G.F. Analysis of the human seminal plasma glycome reveals the presence of immunomodulatory carbohydrate functional groups. J. Proteome Res. 2009, 8, 4906–4915. [Google Scholar] [CrossRef]
- Okamoto, T.; Yoneyama, M.S.; Hatakeyama, S.; Mori, K.; Yamamoto, H.; Koie, T.; Saitoh, H.; Yamaya, K.; Funyu, T.; Fukuda, M.; et al. Core2 O-glycan-expressing prostate cancer cells are resistant to NK cell immunity. Mol. Med. Rep. 2013, 7, 359–364. [Google Scholar]
- Suzuki, Y.; Sutoh, M.; Hatakeyama, S.; Mori, K.; Yamamoto, H.; Koie, T.; Saitoh, H.; Yamaya, K.; Funyu, T.; Habuchi, T.; et al. MUC1 carrying core 2 O-glycans functions as a molecular shield against NK cell attack, promoting bladder tumor metastasis. Int. J. Oncol. 2012, 40, 1831–1838. [Google Scholar]
- Tsuboi, S.; Sutoh, M.; Hatakeyama, S.; Hiraoka, N.; Habuchi, T.; Horikawa, Y.; Hashimoto, Y.; Yoneyama, T.; Mori, K.; Koie, T.; et al. A novel strategy for evasion of NK cell immunity by tumours expressing core2 O-glycans. EMBO J. 2011, 30, 3173–3185. [Google Scholar] [CrossRef]
- Tsuboi, S. Tumor defense systems using O-glycans. Biol. Pharm. Bull. 2012, 35, 1633–1636. [Google Scholar] [CrossRef]
- Galli-Stampino, L.; Meinjohanns, E.; Frische, K.; Meldal, M.; Jensen, T.; Werdelin, O.; Mouritsen, S. T-cell recognition of tumor-associated carbohydrates: The nature of the glycan moiety plays a decisive role in determining glycopeptide immunogenicity. Cancer Res. 1997, 57, 3214–3222. [Google Scholar]
- Snijdewint, F.G.; von Mensdorff-Pouilly, S.; Karuntu-Wanamarta, A.H.; Verstraeten, A.A.; Livingston, P.O.; Hilgers, J.; Kenemans, P. Antibody-dependent cell-mediated cytotoxicity can be induced by MUC1 peptide vaccination of breast cancer patients. Int. J. Cancer 2001, 93, 97–106. [Google Scholar] [CrossRef]
- Pashov, A.; Monzavi-Karbassi, B.; Chow, M.; Cannon, M.; Kieber-Emmons, T. Immune surveillance as a rationale for immunotherapy? Hum. Vaccin. 2007, 3, 224–228. [Google Scholar]
- Van Kooyk, Y.; Rabinovich, G.A. Protein-glycan interactions in the control of innate and adaptive immune responses. Nat. Immunol. 2008, 9, 593–601. [Google Scholar] [CrossRef]
- Pashov, A.; Monzavi-Karbassi, B.; Raghava, G.P.; Kieber-Emmons, T. Bridging innate and adaptive antitumor immunity targeting glycans. J. Biomed. Biotechnol. 2010. [Google Scholar] [CrossRef]
- Freire, T.; Osinaga, E. The sweet side of tumor immunotherapy. Immunotherapy 2012, 4, 719–734. [Google Scholar] [CrossRef]
- Olive, C. Pattern recognition receptors: Sentinels in innate immunity and targets of new vaccine adjuvants. Expert Rev. Vaccines 2012, 11, 237–256. [Google Scholar] [CrossRef]
- Krishnaswamy, J.K.; Chu, T.; Eisenbarth, S.C. Beyond pattern recognition: NOD-like receptors in dendritic cells. Trends Immunol. 2013, 34, 224–233. [Google Scholar] [CrossRef]
- Davicino, R.C.; Elicabe, R.J.; di Genaro, M.S.; Rabinovich, G.A. Coupling pathogen recognition to innate immunity through glycan-dependent mechanisms. Int. Immunopharmacol. 2011, 11, 1457–1463. [Google Scholar] [CrossRef]
- Schwartz-Albiez, R. Naturally occurring antibodies directed against carbohydrate tumor antigens. Adv. Exp. Med. Biol. 2012, 750, 27–43. [Google Scholar] [CrossRef]
- Rodriguez-Zhurbenko, N.; Martinez, D.; Blanco, R.; Rondon, T.; Grinan, T.; Hernandez, A.M. Human antibodies reactive to NeuGcGM3 ganglioside have cytotoxic anti-tumor properties. Eur. J. Immunol. 2013, 14, 201242693. [Google Scholar]
- Monzavi-Karbassi, B.; Artaud, C.; Jousheghany, F.; Hennings, L.; Carcel-Trullols, J.; Shaaf, S.; Korourian, S.; Kieber-Emmons, T. Reduction of spontaneous metastases through induction of carbohydrate cross-reactive apoptotic antibodies. J. Immunol. 2005, 174, 7057–7065. [Google Scholar]
- Farag, S.S.; Caligiuri, M.A. Human natural killer cell development and biology. Blood Rev. 2006, 20, 123–137. [Google Scholar] [CrossRef]
- Koch, J.; Steinle, A.; Watzl, C.; Mandelboim, O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol. 2013, 34, 182–191. [Google Scholar] [CrossRef]
- Porgador, A. Natural cytotoxicity receptors: Pattern recognition and involvement of carbohydrates. ScientificWorldJournal 2005, 5, 151–154. [Google Scholar] [CrossRef]
- Disis, M.L. Immune regulation of cancer. J. Clin. Oncol. 2010, 28, 4531–4538. [Google Scholar] [CrossRef]
- Backer, A.E.; Holgersson, J.; Samuelsson, B.E.; Karlsson, H. Rapid and sensitive GC/MS characterization of glycolipid released Galalpha1,3Gal-terminated oligosaccharides from small organ specimens of a single pig. Glycobiology 1998, 8, 533–545. [Google Scholar] [CrossRef]
- Hallberg, E.C.; Holgersson, J.; Samuelsson, B.E. Glycosphingolipid expression in pig aorta: Identification of possible target antigens for human natural antibodies. Glycobiology 1998, 8, 637–649. [Google Scholar] [CrossRef]
- Bouhours, D.; Liaigre, J.; Lemoine, J.; Mayer-Posner, F.; Bouhours, J.F. Two novel isoneolacto-undecaglycosylceramides carrying Galalpha1-->3Lewis(x) on the 6-linked antenna and N-acetylneuraminic acidalpha2-->3 or Galactose alpha1-->3 on the 3-linked antenna, expressed in porcine kidney. Glycoconj. J. 1998, 15, 1001–1016. [Google Scholar]
- Macher, B.A.; Galili, U. The Galalpha1,3Galbeta1,4GlcNAc-R (alpha-Gal) epitope: A carbohydrate of unique evolution and clinical relevance. Biochim. Biophys. Acta 2008, 1780, 75–88. [Google Scholar] [CrossRef]
- Stussi, G.; Mueller, R.J.; Passweg, J.; Schanz, U.; Rieben, R.; Seebach, J.D. ABO blood group incompatible haematopoietic stem cell transplantation and xenograft rejection. Swiss Med. Wkly. 2007, 137, 101S–108S. [Google Scholar]
- Gates, M.A.; Wolpin, B.M.; Cramer, D.W.; Hankinson, S.E.; Tworoger, S.S. ABO blood group and incidence of epithelial ovarian cancer. Int. J. Cancer 2011, 128, 482–486. [Google Scholar] [CrossRef]
- Hakomori, S. Antigen structure and genetic basis of histo-blood groups A, B and O: Their changes associated with human cancer. Biochim. Biophys. Acta 1999, 1473, 247–266. [Google Scholar] [CrossRef]
- Lin, S.S.; Parker, W.; Everett, M.L.; Platt, J.L. Differential recognition by proteins of alpha-galactosyl residues on endothelial cell surfaces. Glycobiology 1998, 8, 433–443. [Google Scholar] [CrossRef]
- Korourian, S.; Siegel, E.; Kieber-Emmons, T.; Monzavi-Karbassi, B. Expression analysis of carbohydrate antigens in ductal carcinoma in situ of the breast by lectin histochemistry. BMC Cancer 2008, 8, 136. [Google Scholar] [CrossRef]
- Preiss, S.; Kammertoens, T.; Lampert, C.; Willimsky, G.; Blankenstein, T. Tumor-induced antibodies resemble the response to tissue damage. Int. J. Cancer 2005, 115, 456–462. [Google Scholar] [CrossRef]
- Spalter, S.H.; Kaveri, S.V.; Bonnin, E.; Mani, J.C.; Cartron, J.P.; Kazatchkine, M.D. Normal human serum contains natural antibodies reactive with autologous ABO blood group antigens. Blood 1999, 93, 4418–4424. [Google Scholar]
- Galili, U. The alpha-gal epitope and the anti-Gal antibody in xenotransplantation and in cancer immunotherapy. Immunol. Cell Biol. 2005, 83, 674–686. [Google Scholar] [CrossRef]
- Galili, U. Immune response, accommodation, and tolerance to transplantation carbohydrate antigens. Transplantation 2004, 78, 1093–1098. [Google Scholar] [CrossRef]
- Apostolopoulos, V.; Osinski, C.; McKenzie, I.F. MUC1 cross-reactive Gal alpha(1,3)Gal antibodies in humans switch immune responses from cellular to humoral. Nat. Med. 1998, 4, 315–320. [Google Scholar] [CrossRef]
- Abdel-Motal, U.M.; Wigglesworth, K.; Galili, U. Mechanism for increased immunogenicity of vaccines that form in vivo immune complexes with the natural anti-Gal antibody. Vaccine 2009, 27, 3072–3082. [Google Scholar] [CrossRef]
- Cobb, B.A.; Kasper, D.L. Coming of age: Carbohydrates and immunity. Eur. J. Immunol. 2005, 35, 352–356. [Google Scholar] [CrossRef]
- Jackson, S.; Folks, T.M.; Wetterskog, D.L.; Kindt, T.J. A rabbit helper T cell clone reactive against group-specific streptococcal carbohydrate. J. Immunol. 1984, 133, 1553–1557. [Google Scholar]
- Petersen, J.; Purcell, A.W.; Rossjohn, J. Post-translationally modified T cell epitopes: Immune recognition and immunotherapy. J. Mol. Med. 2009, 87, 1045–1051. [Google Scholar] [CrossRef]
- Szabo, T.G.; Palotai, R.; Antal, P.; Tokatly, I.; Tothfalusi, L.; Lund, O.; Nagy, G.; Falus, A.; Buzas, E.I. Critical role of glycosylation in determining the length and structure of T cell epitopes. Immun. Res. 2009, 5, 4. [Google Scholar] [CrossRef]
- Zhao, X.J.; Cheung, N.K. GD2 oligosaccharide: Target for cytotoxic T lymphocytes. J. Exp. Med. 1995, 182, 67–74. [Google Scholar] [CrossRef]
- Tzianabos, A.O.; Finberg, R.W.; Wang, Y.; Chan, M.; Onderdonk, A.B.; Jennings, H.J.; Kasper, D.L. T cells activated by zwitterionic molecules prevent abscesses induced by pathogenic bacteria. J. Biol. Chem. 2000, 275, 6733–6740. [Google Scholar]
- Cobb, B.A.; Kasper, D.L. Characteristics of carbohydrate antigen binding to the presentation protein HLA-DR. Glycobiology 2008, 18, 707–718. [Google Scholar] [CrossRef]
- Haurum, J.S.; Arsequell, G.; Lellouch, A.C.; Wong, S.Y.; Dwek, R.A.; McMichael, A.J.; Elliott, T. Recognition of carbohydrate by major histocompatibility complex class I-restricted, glycopeptide-specific cytotoxic T lymphocytes. J. Exp. Med. 1994, 180, 739–744. [Google Scholar] [CrossRef]
- Haurum, J.S.; Hoier, I.B.; Arsequell, G.; Neisig, A.; Valencia, G.; Zeuthen, J.; Neefjes, J.; Elliott, T. Presentation of cytosolic glycosylated peptides by human class I major histocompatibility complex molecules in vivo. J. Exp. Med. 1999, 190, 145–150. [Google Scholar] [CrossRef]
- Haurum, J.S.; Tan, L.; Arsequell, G.; Frodsham, P.; Lellouch, A.C.; Moss, P.A.; Dwek, R.A.; McMichael, A.J.; Elliott, T. Peptide anchor residue glycosylation: Effect on class I major histocompatibility complex binding and cytotoxic T lymphocyte recognition. Eur. J. Immunol. 1995, 25, 3270–3276. [Google Scholar] [CrossRef]
- Carbone, F.R.; Gleeson, P.A. Carbohydrates and antigen recognition by T cells. Glycobiology 1997, 7, 725–730. [Google Scholar] [CrossRef]
- Madsen, C.B.; Petersen, C.; Lavrsen, K.; Harndahl, M.; Buus, S.; Clausen, H.; Pedersen, A.E.; Wandall, H.H. Cancer associated aberrant protein O-glycosylation can modify antigen processing and immune response. PLoS One 2012, 7, e50139. [Google Scholar]
- Muthukkumar, S.; Stein, K.E. Immunization with meningococcal polysaccharide-tetanus toxoid conjugate induces polysaccharide-reactive T cells in mice. Vaccine 2004, 22, 1290–1299. [Google Scholar] [CrossRef]
- Glithero, A.; Tormo, J.; Haurum, J.S.; Arsequell, G.; Valencia, G.; Edwards, J.; Springer, S.; Townsend, A.; Pao, Y.L.; Wormald, M.; et al. Crystal structures of two H-2Db/glycopeptide complexes suggest a molecular basis for CTL cross-reactivity. Immunity 1999, 10, 63–74. [Google Scholar] [CrossRef]
- Speir, J.A.; Abdel-Motal, U.M.; Jondal, M.; Wilson, I.A. Crystal structure of an MHC class I presented glycopeptide that generates carbohydrate-specific CTL. Immunity 1999, 10, 51–61. [Google Scholar] [CrossRef]
- Stepensky, D.; Tzehoval, E.; Vadai, E.; Eisenbach, L. O-glycosylated versus non-glycosylated MUC1-derived peptides as potential targets for cytotoxic immunotherapy of carcinoma. Clin. Exp. Immunol. 2006, 143, 139–149. [Google Scholar] [CrossRef]
- Bohm, C.M.; Mulder, M.C.; Zennadi, R.; Notter, M.; Schmitt-Graff, A.; Finn, O.J.; Taylor-Papadimitriou, J.; Stein, H.; Clausen, H.; Riecken, E.O.; et al. Carbohydrate recognition on MUC1-expressing targets enhances cytotoxicity of a T cell subpopulation. Scand. J. Immunol. 1997, 46, 27–34. [Google Scholar] [CrossRef]
- Abdel-Motal, U.M.; Berg, L.; Rosen, A.; Bengtsson, M.; Thorpe, C.J.; Kihlberg, J.; Dahmen, J.; Magnusson, G.; Karlsson, K.A.; Jondal, M. Immunization with glycosylated Kb-binding peptides generates carbohydrate-specific, unrestricted cytotoxic T cells. Eur. J. Immunol. 1996, 26, 544–551. [Google Scholar] [CrossRef]
- Qiu, J.; Luo, P.; Wasmund, K.; Steplewski, Z.; Kieber-Emmons, T. Towards the development of peptide mimotopes of carbohydrate antigens as cancer vaccines. Hybridoma 1999, 18, 103–112. [Google Scholar] [CrossRef]
- Luo, P.; Canziani, G.; Cunto-Amesty, G.; Kieber-Emmons, T. A molecular basis for functional peptide mimicry of a carbohydrate antigen. J. Biol. Chem. 2000, 275, 16146–16154. [Google Scholar]
- Pashov, A.; Perry, M.; Dyar, M.; Chow, M.; Kieber-Emmons, T. Carbohydrate mimotopes in the rational design of cancer vaccines. Curr. Top. Med. Chem. 2005, 5, 1171–1185. [Google Scholar] [CrossRef]
- Monzavi-Karbassi, B.; Cunto-Amesty, G.; Luo, P.; Kieber-Emmons, T. Peptide mimotopes as surrogate antigens of carbohydrates in vaccine discovery. Trends Biotechnol. 2002, 20, 207–214. [Google Scholar] [CrossRef]
- Monzavi-Karbassi, B.; Cunto-Amesty, G.; Luo, P.; Shamloo, S.; Blaszcyk-Thurin, M.; Kieber-Emmons, T. Immunization with a carbohydrate mimicking peptide augments tumor-specific cellular responses. Int. Immunol. 2001, 13, 1361–1371. [Google Scholar] [CrossRef]
- Monzavi-Karbassi, B.; Luo, P.; Jousheghany, F.; Torres-Quinones, M.; Cunto-Amesty, G.; Artaud, C.; Kieber-Emmons, T. A mimic of tumor rejection antigen-associated carbohydrates mediates an antitumor cellular response. Cancer Res. 2004, 64, 2162–2166. [Google Scholar] [CrossRef]
- Wondimu, A.; Zhang, T.; Kieber-Emmons, T.; Gimotty, P.; Sproesser, K.; Somasundaram, R.; Ferrone, S.; Tsao, C.Y.; Herlyn, D. Peptides mimicking GD2 ganglioside elicit cellular, humoral and tumor-protective immune responses in mice. Cancer Immunol. Immunother. 2008, 57, 1079–1089. [Google Scholar] [CrossRef]
- Wierzbicki, A.; Gil, M.; Ciesielski, M.; Fenstermaker, R.A.; Kaneko, Y.; Rokita, H.; Lau, J.T.; Kozbor, D. Immunization with a mimotope of GD2 ganglioside induces CD8+ T cells that recognize cell adhesion molecules on tumor cells. J. Immunol. 2008, 181, 6644–6653. [Google Scholar]
- Monzavi-Karbassi, B.; Hennings, L.J.; Artaud, C.; Liu, T.; Jousheghany, F.; Pashov, A.; Murali, R.; Hutchins, L.F.; Kieber-Emmons, T. Preclinical studies of carbohydrate mimetic peptide vaccines for breast cancer and melanoma. Vaccine 2007, 25, 3022–3031. [Google Scholar] [CrossRef]
- Saijo, N.; Ozaki, A.; Beppu, Y.; Takahashi, K.; Fujita, J.; Sasaki, Y.; Nomori, H.; Kimata, M.; Shimizu, E.; Hoshi, A. Analysis of metastatic spread and growth of tumor cells in mice with depressed natural killer activity by anti-asialo GM1 antibody or anticancer agents. J. Cancer Res. Clin. Oncol. 1984, 107, 157–163. [Google Scholar] [CrossRef]
- Wiltrout, R.H.; Herberman, R.B.; Zhang, S.R.; Chirigos, M.A.; Ortaldo, J.R.; Green, K.M., Jr.; Talmadge, J.E. Role of organ-associated NK cells in decreased formation of experimental metastases in lung and liver. J. Immunol. 1985, 134, 4267–4275. [Google Scholar]
- Bezouska, K.; Yuen, C.T.; O’Brien, J.; Childs, R.A.; Chai, W.; Lawson, A.M.; Drbal, K.; Fiserova, A.; Pospisil, M.; Feizi, T. Oligosaccharide ligands for NKR-P1 protein activate NK cells and cytotoxicity. Nature 1994, 372, 150–157. [Google Scholar] [CrossRef]
- Brown, M.G.; Scalzo, A.A. NK gene complex dynamics and selection for NK cell receptors. Semin. Immunol. 2008, 20, 361–368. [Google Scholar] [CrossRef]
- Cheent, K.; Khakoo, S.I. Natural killer cells: Integrating diversity with function. Immunology 2009, 126, 449–457. [Google Scholar] [CrossRef]
- Biassoni, R. Natural killer cell receptors. Adv. Exp. Med. Biol. 2008, 640, 35–52. [Google Scholar] [CrossRef]
- Lanier, L.L. NK cell recognition. Annu. Rev. Immunol. 2005, 23, 225–274. [Google Scholar] [CrossRef]
- Fauriat, C.; Ivarsson, M.A.; Ljunggren, H.G.; Malmberg, K.J.; Michaelsson, J. Education of human natural killer cells by activating killer cell immunoglobulin-like receptors. Blood 2010, 115, 1166–1174. [Google Scholar] [CrossRef]
- Foley, B.; de Santis, D.; Lathbury, L.; Christiansen, F.; Witt, C. KIR2DS1-mediated activation overrides NKG2A-mediated inhibition in HLA-C C2-negative individuals. Int. Immunol. 2008, 20, 555–563. [Google Scholar] [CrossRef]
- Omidvar, N.; Wang, E.C.; Brennan, P.; Longhi, M.P.; Smith, R.A.; Morgan, B.P. Expression of glycosylphosphatidylinositol-anchored CD59 on target cells enhances human NK cell-mediated cytotoxicity. J. Immunol. 2006, 176, 2915–2923. [Google Scholar]
- Tarek, N.; Le Luduec, J.B.; Gallagher, M.M.; Zheng, J.; Venstrom, J.M.; Chamberlain, E.; Modak, S.; Heller, G.; Dupont, B.; Cheung, N.K.; et al. Unlicensed NK cells target neuroblastoma following anti-GD2 antibody treatment. J. Clin. Invest. 2012, 122, 3260–3270. [Google Scholar] [CrossRef]
- Cheung, N.K.; Kushner, B.H.; Cheung, I.Y.; Kramer, K.; Canete, A.; Gerald, W.; Bonilla, M.A.; Finn, R.; Yeh, S.J.; Larson, S.M. Anti-G(D2) antibody treatment of minimal residual stage 4 neuroblastoma diagnosed at more than 1 year of age. J. Clin. Oncol. 1998, 16, 3053–3060. [Google Scholar]
- Modak, S.; Cheung, N.K. Neuroblastoma: Therapeutic strategies for a clinical enigma. Cancer Treat. Rev. 2010, 36, 307–317. [Google Scholar] [CrossRef]
- Yu, A.L.; Gilman, A.L.; Ozkaynak, M.F.; London, W.B.; Kreissman, S.G.; Chen, H.X.; Smith, M.; Anderson, B.; Villablanca, J.G.; Matthay, K.K.; et al. Anti-GD2 antibody with GM-CSF, interleukin-2, and isotretinoin for neuroblastoma. N. Engl. J. Med. 2010, 363, 1324–1334. [Google Scholar] [CrossRef]
- Alderson, K.L.; Luangrath, M.; Elsenheimer, M.M.; Gillies, S.D.; Navid, F.; Rakhmilevich, A.L.; Sondel, P.M. Enhancement of the anti-melanoma response of Hu14.18K322A by alphaCD40+ CpG. Cancer Immunol. Immunother. 2012, 15, 15. [Google Scholar]
- Esser, R.; Muller, T.; Stefes, D.; Kloess, S.; Seidel, D.; Gillies, S.D.; Aperlo-Iffland, C.; Huston, J.S.; Uherek, C.; Schonfeld, K.; et al. NK cells engineered to express a GD2-specific antigen receptor display built-in ADCC-like activity against tumour cells of neuroectodermal origin. J. Cell. Mol. Med. 2012, 16, 569–581. [Google Scholar] [CrossRef]
- Hombach, A.A.; Holzinger, A.; Abken, H. The weal and woe of costimulation in the adoptive therapy of cancer with chimeric antigen receptor (CAR)-redirected T cells. Curr. Mol. Med. 2012, 31, 31. [Google Scholar]
- Albertsson, P.A.; Basse, P.H.; Hokland, M.; Goldfarb, R.H.; Nagelkerke, J.F.; Nannmark, U.; Kuppen, P.J. NK cells and the tumour microenvironment: Implications for NK-cell function and anti-tumour activity. Trends Immunol. 2003, 24, 603–609. [Google Scholar] [CrossRef]
- Parkhurst, M.R.; Riley, J.P.; Dudley, M.E.; Rosenberg, S.A. Adoptive transfer of autologous natural killer cells leads to high levels of circulating natural killer cells but does not mediate tumor regression. Clin. Cancer Res. 2011, 17, 6287–6297. [Google Scholar] [CrossRef]
- Kariya, Y.; Kawamura, C.; Tabei, T.; Gu, J. Bisecting GlcNAc residues on laminin-332 down-regulate galectin-3-dependent keratinocyte motility. J. Biol. Chem. 2009, 285, 3330–3340. [Google Scholar] [CrossRef]
- Chung, T.W.; Kim, K.S.; Kang, S.K.; Lee, J.W.; Song, E.Y.; Chung, T.H.; Yeom, Y.I.; Kim, C.H. Remodeling of the major mouse xenoantigen, Galalpha1-3Galbeta1-4GlcNAc-R, by N-acetylglucosaminyltransferase-III. Mol. Cells 2003, 16, 343–353. [Google Scholar]
- Hershkovitz, O.; Jivov, S.; Bloushtain, N.; Zilka, A.; Landau, G.; Bar-Ilan, A.; Lichtenstein, R.G.; Campbell, K.S.; van Kuppevelt, T.H.; Porgador, A. Characterization of the recognition of tumor cells by the natural cytotoxicity receptor, NKp44. Biochemistry 2007, 46, 7426–7436. [Google Scholar] [CrossRef]
- Byrd, A.; Hoffmann, S.C.; Jarahian, M.; Momburg, F.; Watzl, C. Expression analysis of the ligands for the Natural Killer cell receptors NKp30 and NKp44. PLoS One 2007, 2, e1339. [Google Scholar] [CrossRef]
- Higai, K.; Ichikawa, A.; Matsumoto, K. Binding of sialyl Lewis X antigen to lectin-like receptors on NK cells induces cytotoxicity and tyrosine phosphorylation of a 17-kDa protein. Biochim. Biophys. Acta 2006, 1760, 1355–1363. [Google Scholar] [CrossRef]
- Ito, K.; Higai, K.; Shinoda, C.; Sakurai, M.; Yanai, K.; Azuma, Y.; Matsumoto, K. Unlike natural killer (NK) p30, natural cytotoxicity receptor NKp44 binds to multimeric alpha2,3-NeuNAc-containing N-glycans. Biol. Pharm. Bull. 2012, 35, 594–600. [Google Scholar] [CrossRef]
- Kovalenko, E.I.; Abakushina, E.; Telford, W.; Kapoor, V.; Korchagina, E.; Khaidukov, S.; Molotkovskaya, I.; Sapozhnikov, A.; Vlaskin, P.; Bovin, N. Clustered carbohydrates as a target for natural killer cells: A model system. Histochem. Cell. Biol. 2007, 127, 313–326. [Google Scholar] [CrossRef]
- Bezouska, K.; Kren, V.; Kieburg, C.; Lindhorst, T.K. GlcNAc-terminated glycodendrimers form defined precipitates with the soluble dimeric receptor of rat natural killer cells, sNKR-P1A. FEBS Lett. 1998, 426, 243–247. [Google Scholar] [CrossRef]
- Benson, V.; Grobarova, V.; Richter, J.; Fiserova, A. Glycosylation regulates NK cell-mediated effector function through PI3K pathway. Int. Immunol. 2010, 22, 167–177. [Google Scholar] [CrossRef]
- Grayson, G.; Ladisch, S. Immunosuppression by human gangliosides. II. Carbohydrate structure and inhibition of human NK activity. Cell. Immunol. 1992, 139, 18–29. [Google Scholar] [CrossRef]
- Nicoll, G.; Avril, T.; Lock, K.; Furukawa, K.; Bovin, N.; Crocker, P.R. Ganglioside GD3 expression on target cells can modulate NK cell cytotoxicity via siglec-7-dependent and -independent mechanisms. Eur. J. Immunol. 2003, 33, 1642–1648. [Google Scholar] [CrossRef]
- Kawasaki, Y.; Ito, A.; Withers, D.A.; Taima, T.; Kakoi, N.; Saito, S.; Arai, Y. Ganglioside DSGb5, preferred ligand for Siglec-7, inhibits NK cell cytotoxicity against renal cell carcinoma cells. Glycobiology 2010, 20, 1373–1379. [Google Scholar] [CrossRef]
- Belisle, J.A.; Horibata, S.; Jennifer, G.A.; Petrie, S.; Kapur, A.; Andre, S.; Gabius, H.J.; Rancourt, C.; Connor, J.; Paulson, J.C.; et al. Identification of Siglec-9 as the receptor for MUC16 on human NK cells, B cells, and monocytes. Mol. Cancer 2010, 9, 118. [Google Scholar]
- Yarema, K.J.; Bertozzi, C.R. Chemical approaches to glycobiology and emerging carbohydrate-based therapeutic agents. Curr. Opin. Chem. Biol. 1998, 2, 49–61. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, Q. Recent development in carbohydrate-based cancer vaccines. Curr. Opin. Chem. Biol. 2009, 13, 608–617. [Google Scholar] [CrossRef]
- Costa, C.; Zhao, L.; Burton, W.V.; Bondioli, K.R.; Williams, B.L.; Hoagland, T.A.; Ditullio, P.A.; Ebert, K.M.; Fodor, W.L. Expression of the human alpha1,2-fucosyltransferase in transgenic pigs modifies the cell surface carbohydrate phenotype and confers resistance to human serum-mediated cytolysis. FASEB J. 1999, 13, 1762–1773. [Google Scholar]
- Artrip, J.H.; Kwiatkowski, P.; Michler, R.E.; Wang, S.F.; Tugulea, S.; Ankersmit, J.; Chisholm, L.; McKenzie, I.F.; Sandrin, M.S.; Itescu, S. Target cell susceptibility to lysis by human natural killer cells is augmented by alpha(1,3)-galactosyltransferase and reduced by alpha(1,2)-fucosyltransferase. J. Biol. Chem. 1999, 274, 10717–10722. [Google Scholar] [CrossRef]
- Horvath-Arcidiacono, J.A.; Porter, C.M.; Bloom, E.T. Human NK cells can lyse porcine endothelial cells independent of their expression of Galalpha(1,3)-Gal and killing is enhanced by activation of either effector or target cells. Xenotransplantation 2006, 13, 318–327. [Google Scholar] [CrossRef]
- Dennis, J.W. N-linked oligosaccharide processing and tumor cell biology. Semin. Cancer Biol. 1991, 2, 411–420. [Google Scholar]
- Humphries, M.J.; Matsumoto, K.; White, S.L.; Olden, K. Inhibition of experimental metastasis by castanospermine in mice: Blockage of two distinct stages of tumor colonization by oligosaccharide processing inhibitors. Cancer Res. 1986, 46, 5215–5222. [Google Scholar]
- Chen, L.; Sundback, J.; Olofsson, S.; Jondal, M. Interference with O-glycosylation in RMA lymphoma cells leads to a reduced in vivo growth of the tumor. Int. J. Cancer 2006, 119, 1495–1500. [Google Scholar] [CrossRef]
- Andre, S.; Sanchez-Ruderisch, H.; Nakagawa, H.; Buchholz, M.; Kopitz, J.; Forberich, P.; Kemmner, W.; Bock, C.; Deguchi, K.; Detjen, K.M.; et al. Tumor suppressor p16INK4a—Modulator of glycomic profile and galectin-1 expression to increase susceptibility to carbohydrate-dependent induction of anoikis in pancreatic carcinoma cells. FEBS J. 2007, 274, 3233–3256. [Google Scholar]
- Wen, Y.J.; Mancino, A.; Pashov, A.; Whitehead, T.; Stanley, J.; Kieber-Emmons, T. Antigen binding of human IgG Fabs mediate ERK-associated proliferation of human breast cancer cells. DNA Cell Biol. 2005, 24, 73–84. [Google Scholar] [CrossRef]
- Nobumoto, A.; Oomizu, S.; Arikawa, T.; Katoh, S.; Nagahara, K.; Miyake, M.; Nishi, N.; Takeshita, K.; Niki, T.; Yamauchi, A.; et al. Galectin-9 expands unique macrophages exhibiting plasmacytoid dendritic cell-like phenotypes that activate NK cells in tumor-bearing mice. Clin. Immunol. 2009, 130, 322–330. [Google Scholar] [CrossRef]
- Irie, R.F.; Morton, D.L. Regression of cutaneous metastatic melanoma by intralesional injection with human monoclonal antibody to ganglioside GD2. Proc. Natl. Acad. Sci. USA 1986, 83, 8694–8698. [Google Scholar] [CrossRef]
- Takahashi, T.; Johnson, T.D.; Nishinaka, Y.; Morton, D.L.; Irie, R.F. IgM anti-ganglioside antibodies induced by melanoma cell vaccine correlate with survival of melanoma patients. J. Invest. Dermatol. 1999, 112, 205–209. [Google Scholar] [CrossRef]
- Perez, C.A.; Ravindranath, M.H.; Soh, D.; Gonzales, A.; Ye, W.; Morton, D.L. Serum anti-ganglioside IgM antibodies in soft tissue sarcoma: Clinical prognostic implications. Cancer J. 2002, 8, 384–394. [Google Scholar] [CrossRef]
- Kawashima, I.; Yoshida, Y.; Taya, C.; Shitara, H.; Yonekawa, H.; Karasuyama, H.; Tada, N.; Furukawa, K.; Tai, T. Expansion of natural killer cells in mice transgenic for IgM antibody to ganglioside GD2: Demonstration of prolonged survival after challenge with syngeneic tumor cells. Int. J. Oncol. 2003, 23, 381–388. [Google Scholar]
- Bao, Y.; Han, Y.; Chen, Z.; Xu, S.; Cao, X. IFN-alpha-producing PDCA-1+ Siglec-H-B cells mediate innate immune defense by activating NK cells. Eur. J. Immunol. 2011, 41, 657–668. [Google Scholar] [CrossRef]
- Hsueh, E.C.; Gupta, R.K.; Qi, K.; Morton, D.L. Correlation of specific immune responses with survival in melanoma patients with distant metastases receiving polyvalent melanoma cell vaccine. J. Clin. Oncol. 1998, 16, 2913–2920. [Google Scholar]
- Baumgarth, N.; Herman, O.C.; Jager, G.C.; Brown, L.E.; Herzenberg, L.A.; Chen, J. B-1 and B-2 cell-derived immunoglobulin M antibodies are nonredundant components of the protective response to influenza virus infection. J. Exp. Med. 2000, 192, 271–280. [Google Scholar] [CrossRef]
- Diamond, M.S.; Sitati, E.M.; Friend, L.D.; Higgs, S.; Shrestha, B.; Engle, M. A critical role for induced IgM in the protection against West Nile virus infection. J. Exp. Med. 2003, 198, 1853–1862. [Google Scholar] [CrossRef]
- Brandlein, S.; Pohle, T.; Ruoff, N.; Wozniak, E.; Muller-Hermelink, H.K.; Vollmers, H.P. Natural IgM antibodies and immunosurveillance mechanisms against epithelial cancer cells in humans. Cancer Res. 2003, 63, 7995–8005. [Google Scholar]
- Vollmers, H.P.; Brandlein, S. The “early birds”: Natural IgM antibodies and immune surveillance. Histol. Histopathol. 2005, 20, 927–937. [Google Scholar]
- Yamada, K.; Kinoshita, M.; Hayakawa, T.; Nakaya, S.; Kakehi, K. Comparative studies on the structural features of O-glycans between leukemia and epithelial cell lines. J. Proteome Res. 2009, 8, 521–537. [Google Scholar] [CrossRef]
- Xin, X.; Higai, K.; Imaizumi, Y.; Suzuki, C.; Ito, K.; Itoh, A.; Matsumoto, S.; Azuma, Y.; Matsumoto, K. Natural killer group 2A (NKG2A) and natural killer group 2C (NKG2C) bind to sulfated glycans and alpha2,3-NeuAc-containing glycoproteins. Biol. Pharm. Bull. 2011, 34, 480–485. [Google Scholar] [CrossRef]
- Hennings, L.; Artaud, C.; Jousheghany, F.; Monzavi-Karbassi, B.; Pashov, A.; Kieber-Emmons, T. Carbohydrate mimetic peptides augment carbohydrate-reactive immune responses in the absence of immune pathology. Cancers 2011, 3, 4151–4169. [Google Scholar] [CrossRef]
- Hirohashi, S.; Clausen, H.; Yamada, T.; Shimosato, Y.; Hakomori, S. Blood group A cross-reacting epitope defined by monoclonal antibodies NCC-LU-35 and -81 expressed in cancer of blood group O or B individuals: Its identification as Tn antigen. Proc. Natl. Acad. Sci. USA 1985, 82, 7039–7043. [Google Scholar]
- Galili, U.; Buehler, J.; Shohet, S.B.; Macher, B.A. The human natural anti-Gal IgG. III. The subtlety of immune tolerance in man as demonstrated by crossreactivity between natural anti-Gal and anti-B antibodies. J. Exp. Med. 1987, 165, 693–704. [Google Scholar] [CrossRef]
- Sandrin, M.S.; Vaughan, H.A.; Xing, P.X.; McKenzie, I.F. Natural human anti-Gal alpha(1,3)Gal antibodies react with human mucin peptides. Glycoconj. J. 1997, 14, 97–105. [Google Scholar] [CrossRef]
- Manches, O.; Plumas, J.; Lui, G.; Chaperot, L.; Molens, J.P.; Sotto, J.J.; Bensa, J.C.; Galili, U. Anti-Gal-mediated targeting of human B lymphoma cells to antigen-presenting cells: A potential method for immunotherapy using autologous tumor cells. Haematologica 2005, 90, 625–634. [Google Scholar]
- Corbiere, V.; Chapiro, J.; Stroobant, V.; Ma, W.; Lurquin, C.; Lethe, B.; van Baren, N.; van den Eynde, B.J.; Boon, T.; Coulie, P.G. Antigen spreading contributes to MAGE vaccination-induced regression of melanoma metastases. Cancer Res. 2011, 71, 1253–1262. [Google Scholar] [CrossRef]
- Butschak, G.; Karsten, U. Isolation and characterization of thomsen-friedenreich-specific antibodies from human serum. Tumour Biol. 2002, 23, 113–122. [Google Scholar] [CrossRef]
- Andre, F.; Dieci, M.V.; Dubsky, P.; Sotiriou, C.; Curigliano, G.; Denkert, C.; Loi, S. Molecular pathways: Involvement of immune pathways in the therapeutic response and outcome in breast cancer. Clin. Cancer Res. 2013, 19, 28–33. [Google Scholar] [CrossRef]
- Kurtenkov, O.; Klaamas, K.; Mensdorff-Pouilly, S.; Miljukhina, L.; Shljapnikova, L.; Chuzmarov, V. Humoral immune response to MUC1 and to the Thomsen-Friedenreich (TF) glycotope in patients with gastric cancer: Relation to survival. Acta Oncol. 2007, 46, 316–323. [Google Scholar] [CrossRef]
- Smorodin, E.P.; Kurtenkov, O.A.; Sergeyev, B.L.; Kodar, K.E.; Chuzmarov, V.I.; Afanasyev, V.P. Postoperative change of anti-Thomsen-Friedenreich and Tn IgG level: The follow-up study of gastrointestinal cancer patients. World J. Gastroenterol. 2008, 14, 4352–4358. [Google Scholar] [CrossRef]
- Ragupathi, G.; Gathuru, J.; Livingston, P. Antibody inducing polyvalent cancer vaccines. Cancer Treat. Res. 2005, 123, 157–180. [Google Scholar] [CrossRef]
- Johnston, D.; Bystryn, J.C. Heterogeneous antibody response to polyvalent melanoma vaccines in syngeneic mice. Cancer Immunol. Immunother. 2005, 54, 345–350. [Google Scholar] [CrossRef]
- Liu, C.C.; Ye, X.S. Carbohydrate-based cancer vaccines: Target cancer with sugar bullets. Glycoconj. J. 2012, 29, 259–271. [Google Scholar] [CrossRef]
- Yin, Z.; Huang, X. Recent development in carbohydrate based anti-cancer vaccines. J. Carbohydr. Chem. 2012, 31, 143–186. [Google Scholar]
- Hevey, R.; Ling, C.C. Recent advances in developing synthetic carbohydrate-based vaccines for cancer immunotherapies. Future Med. Chem. 2012, 4, 545–584. [Google Scholar] [CrossRef]
- Zhu, J.; Warren, J.D.; Danishefsky, S.J. Synthetic carbohydrate-based anticancer vaccines: The Memorial Sloan-Kettering experience. Expert Rev. Vaccines 2009, 8, 1399–1413. [Google Scholar] [CrossRef]
- Xu, Y.; Gendler, S.J.; Franco, A. Designer glycopeptides for cytotoxic T cell-based elimination of carcinomas. J. Exp. Med. 2004, 199, 707–716. [Google Scholar] [CrossRef]
- Singh, S.K.; Stephani, J.; Schaefer, M.; Kalay, H.; Garcia-Vallejo, J.J.; den Haan, J.; Saeland, E.; Sparwasser, T.; van Kooyk, Y. Targeting glycan modified OVA to murine DC-SIGN transgenic dendritic cells enhances MHC class I and II presentation. Mol. Immunol. 2009, 47, 164–174. [Google Scholar] [CrossRef]
- Singh, S.K.; Streng-Ouwehand, I.; Litjens, M.; Kalay, H.; Saeland, E.; van Kooyk, Y. Tumour-associated glycan modifications of antigen enhance MGL2 dependent uptake and MHC class I restricted CD8 T cell responses. Int. J. Cancer. 2011, 128, 1371–1383. [Google Scholar] [CrossRef]
- Denda-Nagai, K.; Aida, S.; Saba, K.; Suzuki, K.; Moriyama, S.; Oo-Puthinan, S.; Tsuiji, M.; Morikawa, A.; Kumamoto, Y.; Sugiura, D.; et al. Distribution and function of macrophage galactose-type C-type lectin 2 (MGL2/CD301b): Efficient uptake and presentation of glycosylated antigens by dendritic cells. J. Biol. Chem. 2010, 285, 19193–19204. [Google Scholar] [CrossRef]
- Lakshminarayanan, V.; Thompson, P.; Wolfert, M.A.; Buskas, T.; Bradley, J.M.; Pathangey, L.B.; Madsen, C.S.; Cohen, P.A.; Gendler, S.J.; Boons, G.J. Immune recognition of tumor-associated mucin MUC1 is achieved by a fully synthetic aberrantly glycosylated MUC1 tripartite vaccine. Proc. Natl. Acad. Sci. USA 2012, 109, 261–266. [Google Scholar] [CrossRef]
- Samsen, A.; Bogoevska, V.; Klampe, B.; Bamberger, A.M.; Lucka, L.; Horst, A.K.; Nollau, P.; Wagener, C. DC-SIGN and SRCL bind glycans of carcinoembryonic antigen (CEA) and CEA-related cell adhesion molecule 1 (CEACAM1): Recombinant human glycan-binding receptors as analytical tools. Eur. Cell Biol. 2010, 89, 87–94. [Google Scholar] [CrossRef]
- Saeland, E.; van Vliet, S.J.; Backstrom, M.; van den Berg, V.C.; Geijtenbeek, T.B.; Meijer, G.A.; van Kooyk, Y. The C-type lectin MGL expressed by dendritic cells detects glycan changes on MUC1 in colon carcinoma. Cancer Immunol. Immunother. 2007, 56, 1225–1236. [Google Scholar] [CrossRef]
- Sato, K.; Imai, Y.; Higashi, N.; Kumamoto, Y.; Mukaida, N.; Irimura, T. Redistributions of macrophages expressing the macrophage galactose-type C-type lectin (MGL) during antigen-induced chronic granulation tissue formation. Int. Immunol. 2005, 17, 559–568. [Google Scholar] [CrossRef]
- Chun, K.H.; Imai, Y.; Higashi, N.; Irimura, T. Migration of dermal cells expressing a macrophage C-type lectin during the sensitization phase of delayed-type hypersensitivity. J. Leukoc. Biol. 2000, 68, 471–478. [Google Scholar]
- Becker, Y. Molecular immunological approaches to biotherapy of human cancers—A review, hypothesis and implications. Anticancer Res. 2006, 26, 1113–1134. [Google Scholar]
- Botella-Estrada, R.; Escudero, M.; O’Connor, J.E.; Nagore, E.; Fenollosa, B.; Sanmartin, O.; Requena, C.; Guillen, C. Cytokine production by peripheral lymphocytes in melanoma. Eur. Cytokine Netw. 2005, 16, 47–55. [Google Scholar]
- Knutson, K.L.; Disis, M.L. Tumor antigen-specific T helper cells in cancer immunity and immunotherapy. Cancer Immunol. Immunother. 2005, 54, 721–728. [Google Scholar] [CrossRef]
- Van Die, I.; Cummings, R.D. Glycan gimmickry by parasitic helminths: A strategy for modulating the host immune response? Glycobiology 2009, 20, 2–12. [Google Scholar] [CrossRef]
- Minkis, K.; Kavanagh, D.G.; Alter, G.; Bogunovic, D.; O’Neill, D.; Adams, S.; Pavlick, A.; Walker, B.D.; Brockman, M.A.; Gandhi, R.T.; et al. Type 2 Bias of T cells expanded from the blood of melanoma patients switched to type 1 by IL-12p70 mRNA-transfected dendritic cells. Cancer Res. 2008, 68, 9441–9450. [Google Scholar] [CrossRef]
- Monzavi-Karbassi, B.; Shamloo, S.; Kieber-Emmons, M.; Jousheghany, F.; Luo, P.; Lin, K.Y.; Cunto-Amesty, G.; Weiner, D.B.; Kieber-Emmons, T. Priming characteristics of peptide mimotopes of carbohydrate antigens. Vaccine 2003, 21, 753–760. [Google Scholar] [CrossRef]
- Pashov, A.D.; Plaxco, J.; Kaveri, S.V.; Monzavi-Karbassi, B.; Harn, D.; Kieber-Emmons, T. Multiple antigenic mimotopes of HIV carbohydrate antigens: Relating structure and antigenicity. J. Biol. Chem. 2006, 281, 29675–29683. [Google Scholar]
- Hermsen, B.B.; Verheijen, R.H.; Menko, F.H.; Gille, J.J.; van Uffelen, K.; Blankenstein, M.A.; Meijer, S.; van Diest, P.J.; Kenemans, P.; von Mensdorff-Pouilly, S. Humoral immune responses to MUC1 in women with a BRCA1 or BRCA2 mutation. Eur. J. Cancer 2007, 43, 1556–1563. [Google Scholar] [CrossRef]
- Klinman, N.R. The mechanism of antigenic stimulation of primary and secondary clonal precursor cells. J. Exp. Med. 1972, 136, 241–257. [Google Scholar] [CrossRef]
- Joncker, N.T.; Fernandez, N.C.; Treiner, E.; Vivier, E.; Raulet, D.H. NK cell responsiveness is tuned commensurate with the number of inhibitory receptors for self-MHC class I: The rheostat model. J. Immunol. 2009, 182, 4572–4580. [Google Scholar] [CrossRef]
- Joncker, N.T.; Shifrin, N.; Delebecque, F.; Raulet, D.H. Mature natural killer cells reset their responsiveness when exposed to an altered MHC environment. J. Exp. Med. 2010, 207, 2065–2072. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Monzavi-Karbassi, B.; Pashov, A.; Kieber-Emmons, T. Tumor-Associated Glycans and Immune Surveillance. Vaccines 2013, 1, 174-203. https://doi.org/10.3390/vaccines1020174
Monzavi-Karbassi B, Pashov A, Kieber-Emmons T. Tumor-Associated Glycans and Immune Surveillance. Vaccines. 2013; 1(2):174-203. https://doi.org/10.3390/vaccines1020174
Chicago/Turabian StyleMonzavi-Karbassi, Behjatolah, Anastas Pashov, and Thomas Kieber-Emmons. 2013. "Tumor-Associated Glycans and Immune Surveillance" Vaccines 1, no. 2: 174-203. https://doi.org/10.3390/vaccines1020174