Effect of Yeast Probiotic on Growth, Antioxidant Enzyme Activities and Malondialdehyde Concentration of Broiler Chickens

 ,
,
Abstract
:1. Introduction
2. Experimental Section
2.1. Experimental Design and Animal Management

{kind=link}
{kind=link}
| Ingredients | Starter | Finisher | Starter Nutrient levels | Finisher Nutrient levels | ||
|---|---|---|---|---|---|---|
| Maize | N/S | N/S | ME, Kcal/Kg | 2800 | ME, Kcal/kg | 2900 |
| Soybean | N/S | N/S | CP, % | 20 | CP, % | 19 |
| Palm kernel cake | N/S | N/S | Ca, % | 1.0 | Ca, % | 1.0 |
| Wheat offal | N/S | N/S | Available P, % | 0.45 | Available P, % | 0.40 |
| Fish meal | N/S | N/S | CF, % | 9 | CF, % | 10 |
| Blood meal | N/S | N/S | Fat, % | 10 | Fat, % | 10 |
| Bone meal | N/S | N/S | ||||
| Oyster shell | N/S | N/S | ||||
| l-Lysine | N/S | N/S | ||||
| l-Methionine | N/S | N/S | ||||
| Vit/Min premix | N/S | N/S | ||||
| Salt | N/S | N/S | ||||
| Total | N/S | N/S | ||||
| Ingredient (%) | Starter | Finisher |
|---|---|---|
| Dry matter | 93.87 | 94.86 |
| Crude protein | 23.69 | 21.20 |
| Crude fibre | 4.87 | 6.14 |
| Oil | 6.04 | 5.31 |
| Ash | 7.86 | 7.88 |
| NFE | 57.54 | 59.47 |
2.2. Body Weight Measurement
2.3. Carcass and Organ Weight Measurement
2.4. Enzymatic Measurements
2.5. Statistical Analysis
3. Results and Discussion
3.1. Body Weight, Feed Intake and Feed Conversion Ratio of Broiler Chickens
| Week | C | T1 0.25 mL | T2 0.5 mL | T3 1.0 mL | p-values |
|---|---|---|---|---|---|
| 1 | 90.97 ± 1.98 | 91.63 ± 1.14 | 67.63 ± 2.45 *** | 89.77 ± 1.61 *** | p < 0.0001 |
| 2 | 185.4 ± 5.25 | 199 ± 3.84 | 147.9 ± 6.34 *** | 185.4 ± 5.29 *** | p < 0.0001 |
| 3 | 402.7 ± 12.55 | 425.3 ± 10.13 | 355.4 ± 16.57 * | 407.9 ± 12.48 * | 0.0015 |
| 4 | 647.4 ± 17.29 | 733.6 ± 13.93 ** | 611.6 ± 23.92 *** | 544.7 ± 18.15 ** | p < 0.0001 |
| 5 | 886.0 ± 30.77 | 926.5 ± 18.59 | 862.1 ± 38.19 | 804 ± 32.50 * | 0.0322 |
| 6 | 1157 ± 49.55 | 1269 ± 31.17 | 1141 ± 43.29 | 1143 ± 46.48 | 0.1307 |
| Week | C | T1 0.25 mL | T2 0.5 mL | T3 1.0 mL | p-values |
|---|---|---|---|---|---|
| 1 | 424.3 ± 68 | 574.3 ± 116 | 365.7 ± 33 | 557.4 ± 108 | 0.0395 |
| 2 | 1161.0 ± 91 | 1175.0 ± 103 | 1097.0 ± 97 | 1089.0 ± 91 | p < 0.0001 |
| 3 | 2149.0 ± 222 | 2206.0 ± 205 | 1771.0 ± 139 * | 1805.0 ± 137 * | p < 0.0001 |
| 4 | 2958.0 ± 133 | 3007.0 ± 132 | 2559.0 ± 107 *** | 2579.0 ± 105 *** | p < 0.0001 |
| 5 | 5379.0 ± 424 | 5521.0 ± 343 | 4342.0 ± 273 *** | 4377.0 ± 271 *** | p < 0.0001 |
| 6 | 7283.0 ± 116 | 7570.0 ± 173 | 5856.0 ± 65 *** | 6159.0 ± 107 *** | p < 0.0001 |
| Week | C | T1 0.25 mL | T2 0.5 mL | T3 1.0 mL |
|---|---|---|---|---|
| 1 | 0.16 | 0.21 | 0.24 | 0.22 |
| 2 | 0.33 | 0.24 | 0.39 | 0.31 |
| 3 | 0.27 | 0.21 | 0.26 | 0.22 |
| 4 | 0.33 | 0.22 | 0.30 | 0.52 |
| 5 | 0.66 | 0.64 | 0.53 | 0.48 |
| 6 | 0.79 | 0.49 | 0.64 | 0.53 |
3.2. Carcass and Organ Weights of Broiler Chickens
| Parameters | C | T1 0.25 mL | T2 0.5 mL | T3 1.0 mL | p-values |
|---|---|---|---|---|---|
| Live weight (g) | 1382.0 ± 37.95 | 1678.0 ± 64.34 ** | 1482.0 ± 34.69 | 1515.0 ± 56.08 | 0.0034 |
| Carcass weight | 903.5 ± 35.47 | 1094.0 ± 46.68 ** | 987.5 ± 26.82 | 996.7 ± 31.77 | 0.0096 |
| % | 65.38 | 65.20 | 66.63 | 65.79 | |
| Thigh (g) | 270.0 ± 8.65 | 318.6 ± 14.49 * | 298.9 ± 8.11 | 303.8 ± 10.91 | 0.0424 |
| % | 29.88 | 29.12 | 30.27 | 30.48 | |
| Drum stick (g) | 153.9 ± 4.68 | 178.5 ± 7.73 * | 171.5 ± 5.55 | 170.7 ± 3.56 | 0.0368 |
| % | 17.03 | 16.32 | 17.37 | 17.13 | |
| Abdominal fat | 9.10 ± 0.18 | 6.60 ± 0.16 *** | 7.60 ± 0.16 *** | 7.10 ± 0.23 *** | p < 0.0001 |
| % | 1.01 | 0.60 | 0.77 | 0.71 | |
| Gizzard (g) | 45.00 ± 4.79 | 57.50 ± 2.32 | 54.80 ± 2.34 | 56.30 ± 3.89 | 0.1050 |
| % | 4.98 | 5.26 | 5.55 | 5.65 | |
| Liver (g) | 44.50 ± 1.92 | 45.80 ± 1.98 | 47.10 ± 1.96 | 50.00 ± 1.76 | 0.1455 |
| % | 4.93 | 4.19 | 4.77 | 5.02 | |
| Heart (g) | 9.40 ± 0.45 | 10.40 ± 0.52 | 9.40 ± 0.31 | 10.90 ± 0.32 | 0.0376 |
| % | 1.04 | 0.95 | 0.95 | 1.09 | |
| Lungs (g) | 10.70 ± 0.42 | 11.80 ± 0.49 | 10.80 ± 0.51 | 11.60 ± 0.43 | 0.2633 |
| % | 1.18 | 1.08 | 1.09 | 1.16 | |
| Intestine (g) | 151.7 ± 8.23 | 177.6 ± 7.64 | 157.3 ± 4.64 | 168.0 ± 7.11 | 0.0590 |
| % | 16.79 | 16.23 | 15.93 | 16.86 |
3.3. Anti-Oxidant Enzyme Activities and Malondialdehyde Concentration of Broiler Chickens
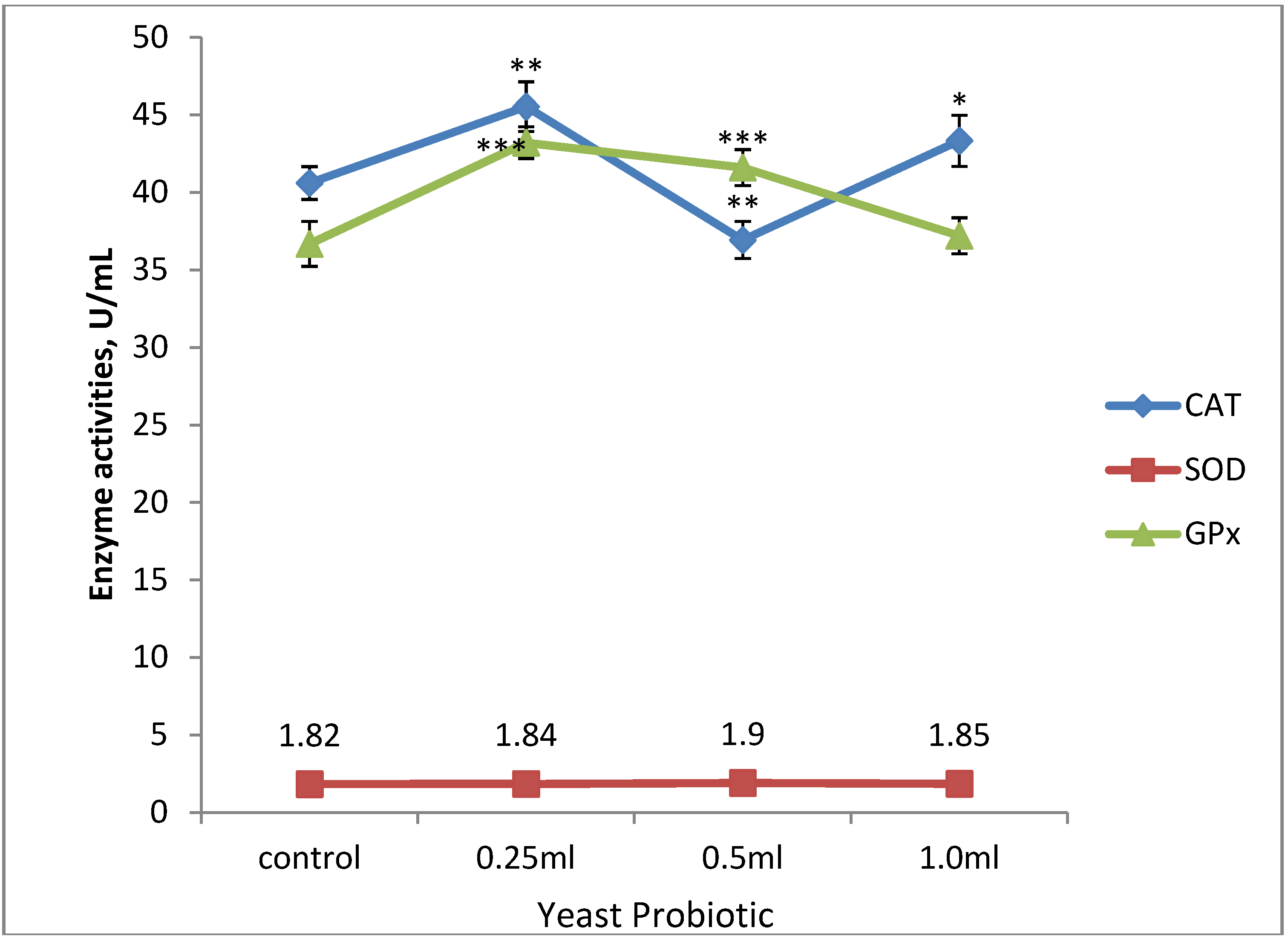
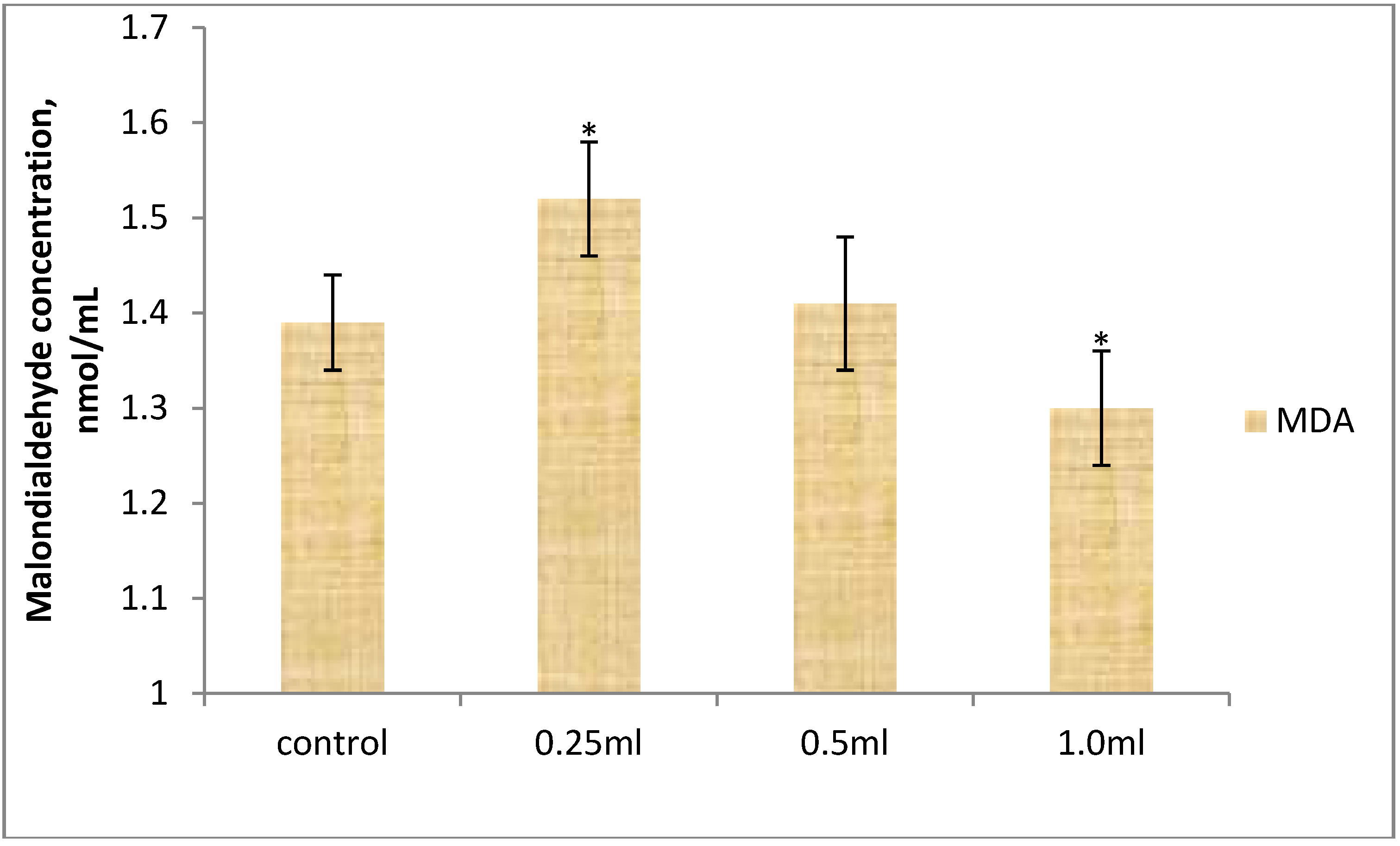
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Breves, G.; Walter, C.; Burmeister, M.; Shroder, B. In vitro studies on the effects of Saccharomyces boulardii and Bacillus cereus var. toyoi on nutrient transport in pig jejunum. J. Anim. Physiol. Anim. Nutr. 2000, 84, 9–20. [Google Scholar] [CrossRef]
- Simon, O.; Vahjen, W.; Scharek, L. Microorganisms as Feed Additive-Probiotics. In Proceedings of the 9th International Symposium on Digestive Physiology in Pigs, Banff, Alberta, Canada, 14–17 May 2003; Volume 1, pp. 295–318.
- O’dea, E.E.; Fasenko, G.M.; Allison, G.E.; Korver, D.R.; Tannock, G.W.; Guan, L.L. Investigating the effects of commercial probiotics on broiler chick quality and production efficiency. Poult. Sci. 2006, 85, 1855–1863. [Google Scholar] [CrossRef] [PubMed]
- Peric, L.; Milosevic, N.; Zikic, D.; Bjedov, S.; Cvetkovic, D.; Markov, S.; Mohnl, M.; Steiner, T. Effect of probiotic and phytobiotic products on performance, gut morphology and cecal microflora of broiler chicks. Arch. Tierz. 2010, 53, 350–359. [Google Scholar]
- Zhang, A.W.; Lee, B.D.; Lee, S.K.; Lee, K.W.; An, G.H.; Song, K.B.; Lee, C.H. Effects of yeast (Saccharomyces cerevisiae) cell components on growth performance, meat quality and ileal mucosa development of broiler chicks. Poult. Sci. 2005, 84, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Cowey, C.B. The role of nutritional factors in the prevention of peroxidative damage to tissues. Fish Physiol. Biochem. 1986, 2, 171–178. [Google Scholar] [CrossRef]
- Sies, H. Oxidative Stress: Oxidants and Antioxidants; Academic Press: New York, NY, USA, 1991. [Google Scholar]
- Jones, D.P. Redefining oxidative stress. Antioxid. Redox Signal. 2006, 8, 1865–1879. [Google Scholar] [CrossRef]
- Halliwell, B. Reactive species and antioxidants redox biology is a fundamental theme of aerobic life. Plant Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef]
- Buetler, T.M.; Krauskopf, A.; Ruegg, U.T. Role of superoxide as a signalling molecule. News Physiol. Sci. 2004, 19, 120–123. [Google Scholar] [PubMed]
- Day, C.P. Is necro-inflammation a prerequisite for fibrosis? Hepatogastroenterology 1996, 43, 104–120. [Google Scholar] [PubMed]
- Wang, Y.Z.; Xu, C.L.; An, Z.H.; Liu, J.-X.; Feng, J. Effect of dietary bovine lactoferin on performance and antioxidant status of piglets. Anim. Feed Sci. Technol. 2008, 140, 326–336. [Google Scholar] [CrossRef]
- Gasch, A.P.; Spellman, P.T.; Kao, C.M.; Carmel-Harel, O.; Eisen, M.B.; Storz, G.; Botstein, D.; Brown, P.O. Genomic expression programs in the response of yeast cells to environmental changes. Mol. Biol. Cell. 2000, 11, 4241–4257. [Google Scholar] [CrossRef]
- Ikner, A.; Shiozaki, K. Yeast signalling pathways in the oxidative stress response. Mutat. Res. 2005, 569, 13–27. [Google Scholar] [CrossRef]
- Temple, M.D.; Perrone, G.G.; Dawes, I.W. Complex cellular responses to reactive species. Trends Cell Biol. 2005, 15, 319–326. [Google Scholar] [CrossRef]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar] [PubMed]
- Martin, J.P., Jr.; Dailey, M.; Sugarman, E. Negative and positive assays of superoxide dismutase based on haematoxylin auto-oxidation. Arch. Biochem. Biophys. 1987, 255, 329–336. [Google Scholar] [CrossRef]
- Beers, R.F., Jr.; Sizer, I.W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by Catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [PubMed]
- Placer, Z.A.; Cushman, L.L.; Johnson, B.C. Estimation of lipid peroxidation, malondialdehyde in biochemical system. Ann. Biochem. 1996, 16, 359–367. [Google Scholar]
- Gohain, A.K.; Sapcota, D. Effect of probiotics feeding on the performance of broilers. Indian J. Poult. Sci. 1998, 33, 101–105. [Google Scholar]
- Morales-Lopez, R.; Auclair, E.; van Immerseel, F.; Ducatelle, R.; Garcia, F.; Brufau, J. Effects of different yeast cell wall supplements added to maize- or wheat-based diets for broiler chickens. Br. Poult. Sci. 2010, 51, 399–408. [Google Scholar] [CrossRef]
- Santin, E.; Maiorka, A.; Macari, M.; Grecco, M.; Sanchez, J.C.; Okada, T.M.; Myasaka, A.M. Performance and intestinal mucosa development of broiler chickens fed diets containing Saccharomyces cerevisiae cell wall. J. Appl. Poult. Res. 2001, 10, 236–244. [Google Scholar] [CrossRef]
- Bansal, G.R.; Singh, V.P.; Sachan, N. Effect of probiotic supplementation on the performance of broilers. Asian J. Anim. Sci. 2011, 5, 277–284. [Google Scholar] [CrossRef]
- Safalaoh, A.C.L. Body weight gain, dressing percentage, abdominal fat and serum cholesterol of broilers supplemented with a microbial preparation. Afr. J. Food Agric. Nutr. Dev. 2006, 6, 1–10. [Google Scholar]
- Sanchez, R.; Ayaya, J.A. Effect of MOS on Broiler Performance under Field Conditions; Alltech’s INC.: Selden, NY, USA, 1998. [Google Scholar]
- Yeo, J.; Kyu-il, K. Effect of feeding diets containing an antibiotic, a probiotic, or yucca extract on growth and intestinal urease activity in broiler chicks. Poult. Sci. 1997, 76, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Sims, M.D.; Sefton, A.E. Comparative Effects of Mannan-Oligosaccharide and an Antibiotic Growth Promoter on Performance of Commercial Broilers. In Presented at the 50th North Central Avian Disease Conference, Vancouver, BC, Canada, 1999.
- Jin, L.Z.; Ho, Y.W.; Abdullahi, N.; Ali, M.A.; Jalaludin, S. Effects of adherent Lactobacillus cultures on growth, weight of organs and intestinal microflora and volatile fatty acids in broilers. Anim. Feed Sci. 1998, 70, 197–209. [Google Scholar] [CrossRef]
- Szymczyk, B.; Pisulewski, M.P.; Szczurek, S. Effects of conjugated linoleic acid on growth performance, feed conversion efficiency, and subsequent carcass quality in broiler chickens. Br. J. Nutr. 2001, 85, 465–473. [Google Scholar] [CrossRef]
- Santoso, U.; Tanaka, K.; Ohtani, S. Effect of dried Bacillus subtilis culture on growth, body composition and hepatic lipogenic enzyme activity in female broiler chicks. Br. J. Nutr. 1995, 74, 523–529. [Google Scholar] [CrossRef]
- Gaya, L.G.; Mourao, G.B.; Rzende, F.M.; Chicaronide Matos, E.; Filho, T.M.; Figueiredo, L.G.G.; Ferraz, J.B.S.; Eler, J.P. Genetic trends of abdominal fat content in a male broiler chicken line. Genet. Mol. Res. 2005, 4, 760–764. [Google Scholar] [PubMed]
- Homma, H.; Shinohara, T. Effects of probiotic Bacillus cereus toyoi on abdominal fat accumulation in Japanese quail (Coturnix japonica). Anim. Sci. J. 2004, 75, 37–41. [Google Scholar] [CrossRef]
- Anjum, M.L.; Khan, A.G.; Azim, A.; Afzal, M. Effect of dietary supplementation of multi-strain probiotic on broiler growth performance. Pak. Vet. J. 2005, 25, 25–29. [Google Scholar]
- Bozkurt, M.; Kucukyilmaz, K.; Cath, A.U.; Cinar, M. Growth Performance and Carcass Yield of Broiler Chickens Given Antibiotic, Mannan Oligosaccharide and Dextran Oligosaccharide Supplemented Diets. In Nutritional Biotechnology in the Feed and Food Industries, Proceedings of Alltech’s 21st Annual Symposium, Lexington, KY, USA, 22–25 May 2005; pp. 5–13.
- Alcicek, A.; Bozkurt, M.; Cabuk, M. The effects of a mixture of herbal essential oil, an organic acid or a probiotic on broiler performance. S. Afr. J. Anim. Sci. 2004, 34, 217–222. [Google Scholar]
- Bozkurt, M.; Kucukyilmaz, K.; Cath, A.U.; Cinar, M. The effect of single or combined dietary supplementation of prebiotics, organic acid and probiotics on performance and slaughter characteristics of broilers. S. Afr. J. Anim. Sci. 2009, 39, 197–205. [Google Scholar]
- Gutowicz, M.; Cholojczyk, M.; Pyrzanowska, R.M.; Cholojczyk, M.; Pyrzanowska, J.; Widy-Tyszkiewicz, E.; Baranczyk-Kuzma, A. Effect of curcumin on antioxidant and detoxification mechanisms in the livers of aging rats. Med. Weter. 2008, 64, 955–957. (in Polish). [Google Scholar]
- Lecewicz, A.; Jankowski, J.; Zdunczyk, Z.; Juskiewicz, J. Selected factors stimulating the development of some gastrointestinal parts in turkeys. Med. Weter. 2008, 64, 1184–1186. [Google Scholar]
- Ognik, K.; Krauze, M. Dietary supplementation of mannanoligosaccharides to turkey hens on their growth performance and antioxidant status in blood. S. Afr. J. Anim. Sci. 2012, 42, 379–388. [Google Scholar]
- Krizkova, L.; Durackova, Z.; Sandula, J.; Sasinkova, V.; Krajcovic, J. Antioxidative and antimutagenic activity of yeast cell wall mannans in vitro. Mutat. Res. 2001, 497, 213–222. [Google Scholar] [CrossRef]
- Kogan, G.; Pajtinka, M.; Babincova, M.; Miadokova, E.; Rauko, P.; Slamenova, D.; Korolenko, T.A. Yesat cell wall polysaccharides as antioxidants and antimutagens: Can they fight cancer? Neoplasma 2008, 55, 387–393. [Google Scholar] [PubMed]
- Milinkovic-Tur, S.; Aladrovic, J.; Ljubic, B.B.; Poljicak-Milas, N. Age-related antioxidant enzyme activities and lipid peroxidation in heart muscles of broiler chickens fed with supplementary organic selenium. Vet. Arhiv. 2009, 79, 481–489. [Google Scholar]
- Ibrahim, W.; Lee, U.S.; Yen, H.C.; Siclair, D.K.; Chow, C.K. Antioxidant and oxidative stress status in tissues of manganese superoxide dismutase transgenic mice. Free Radic. Biol. Med. 2000, 3, 397–402. [Google Scholar]
- Milinkovic-Tur, S.; Stojevic, Z.; Pirsljin, J.; Zdelar-Tuk, M.; Poljicak-Milas, N.; Beer Ljubic, B.; Gradinski-Vrbanac, B. Effect of fasting and refeeding on the antioxidant system in cockerels and pullets. Acta Vet. Hung. 2007, 55, 181–189. [Google Scholar] [CrossRef]
- Kuricova, S.; Boldizarova, K.; Geresakova, L.; Bobcek, R.; Levkut, M.; Leng, L. Chicken selenium status when fed a diet supplemented with Se-yeast. Acta Vet. Brno 2003, 72, 339–346. [Google Scholar] [CrossRef]
- Pirsljin, J.; Milinkovic-Tur, S.; Beer Ljubic, B.; Zdelar-Tuk, M. The effect of organic selenium supplementation on the antioxidative characteristics and lipid peroxidation of chicken blood during fattening and after fasting. Vet. Arhiv. 2008, 78, 187–196. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C. Free Radical Biology and Medicine, 3rd ed.; Oxford University Press: Oxford, UK, 1999; pp. 23–27. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Aluwong, T.; Kawu, M.; Raji, M.; Dzenda, T.; Govwang, F.; Sinkalu, V.; Ayo, J. Effect of Yeast Probiotic on Growth, Antioxidant Enzyme Activities and Malondialdehyde Concentration of Broiler Chickens. Antioxidants 2013, 2, 326-339. https://doi.org/10.3390/antiox2040326
Aluwong T, Kawu M, Raji M, Dzenda T, Govwang F, Sinkalu V, Ayo J. Effect of Yeast Probiotic on Growth, Antioxidant Enzyme Activities and Malondialdehyde Concentration of Broiler Chickens. Antioxidants. 2013; 2(4):326-339. https://doi.org/10.3390/antiox2040326
Chicago/Turabian StyleAluwong, Tagang, Mohammed Kawu, Moshood Raji, Tavershima Dzenda, Felix Govwang, Victor Sinkalu, and Joseph Ayo. 2013. "Effect of Yeast Probiotic on Growth, Antioxidant Enzyme Activities and Malondialdehyde Concentration of Broiler Chickens" Antioxidants 2, no. 4: 326-339. https://doi.org/10.3390/antiox2040326
APA StyleAluwong, T., Kawu, M., Raji, M., Dzenda, T., Govwang, F., Sinkalu, V., & Ayo, J. (2013). Effect of Yeast Probiotic on Growth, Antioxidant Enzyme Activities and Malondialdehyde Concentration of Broiler Chickens. Antioxidants, 2(4), 326-339. https://doi.org/10.3390/antiox2040326