Valuing the Endangered Species Antirrhinum lopesianum: Neuroprotective Activities and Strategies for in vitro Plant Propagation

 ,
,
Abstract
:
1. Introduction
2. Experimental Section
2.1. Plant Material
2.2. Chemical Characterization
2.2.1. Extraction of Plant Phytomolecules
2.2.2. Fractionation by Solid Phase Extraction
2.2.3. Total Phenolic Content
2.2.4. Total Flavonoid Content
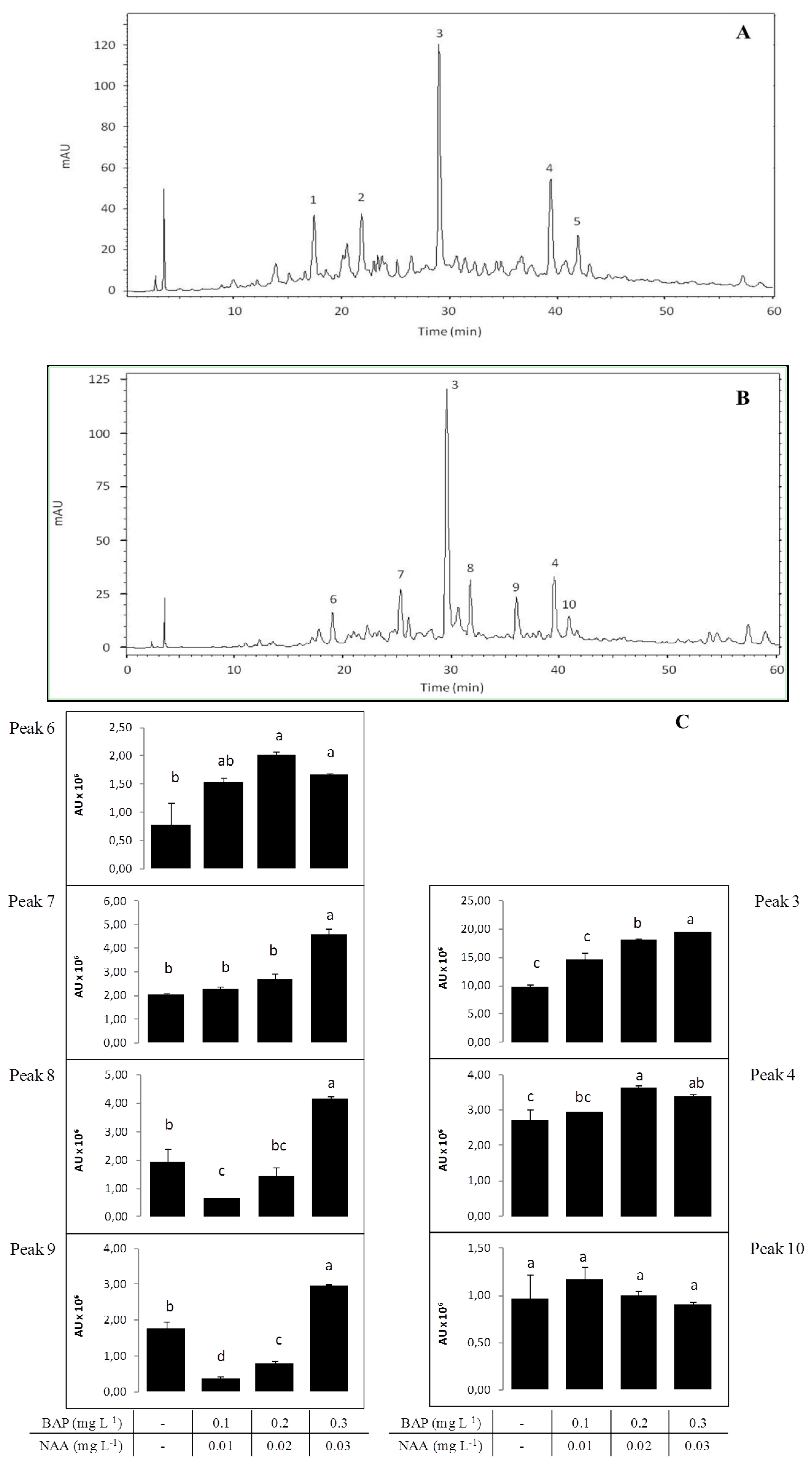
2.2.5. HPLC Profile
2.3. Bioactivities Assessment
2.3.1. Antioxidant Capacity
2.3.2. Acetylcholinesterase (AChE) Inhibitory Assay
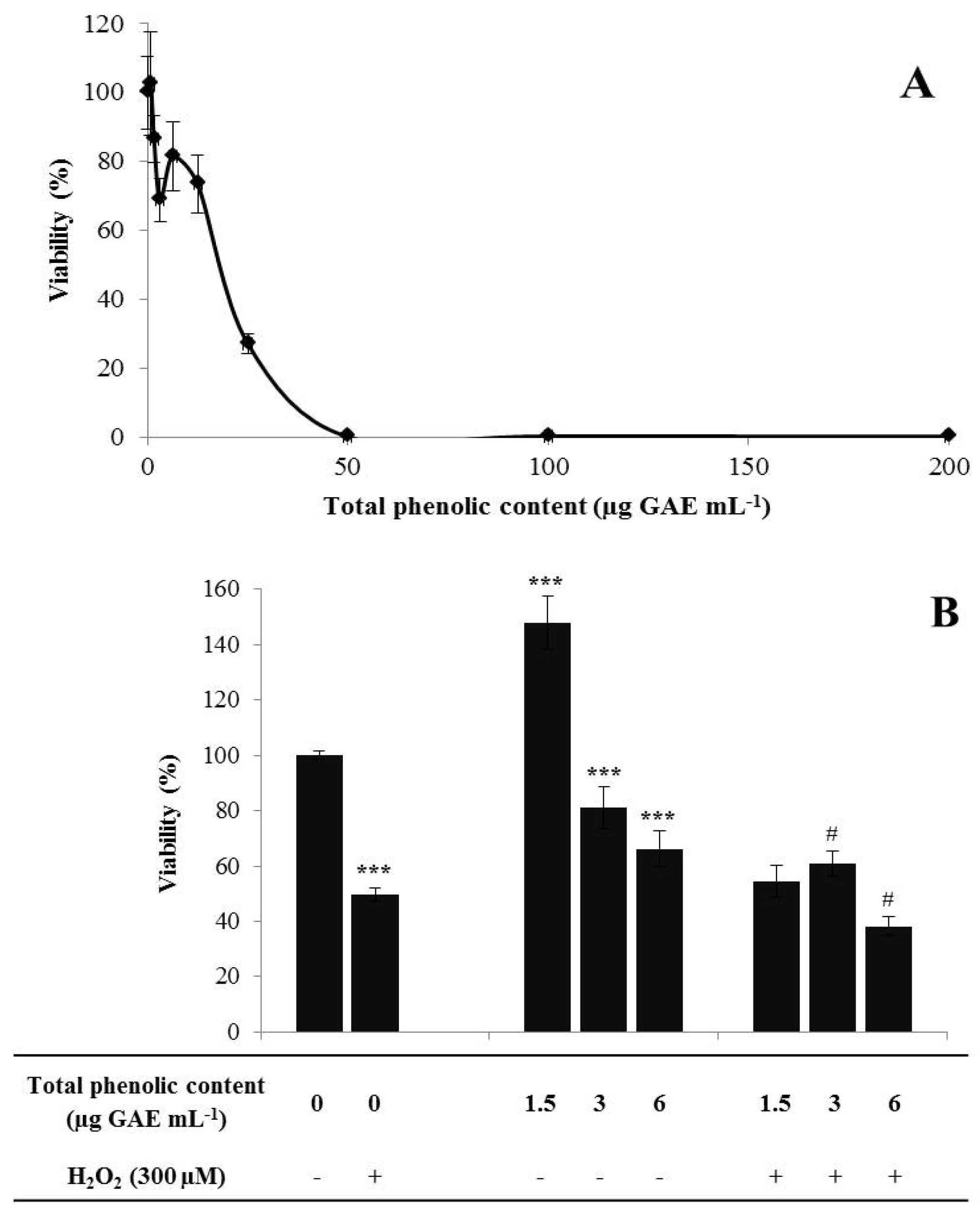
2.3.3. Cytotoxicity Profile
2.3.4. Neuroprotective Effect
2.4. In vitro Establishment and Propagation
2.4.1. Seed Disinfection and Germination
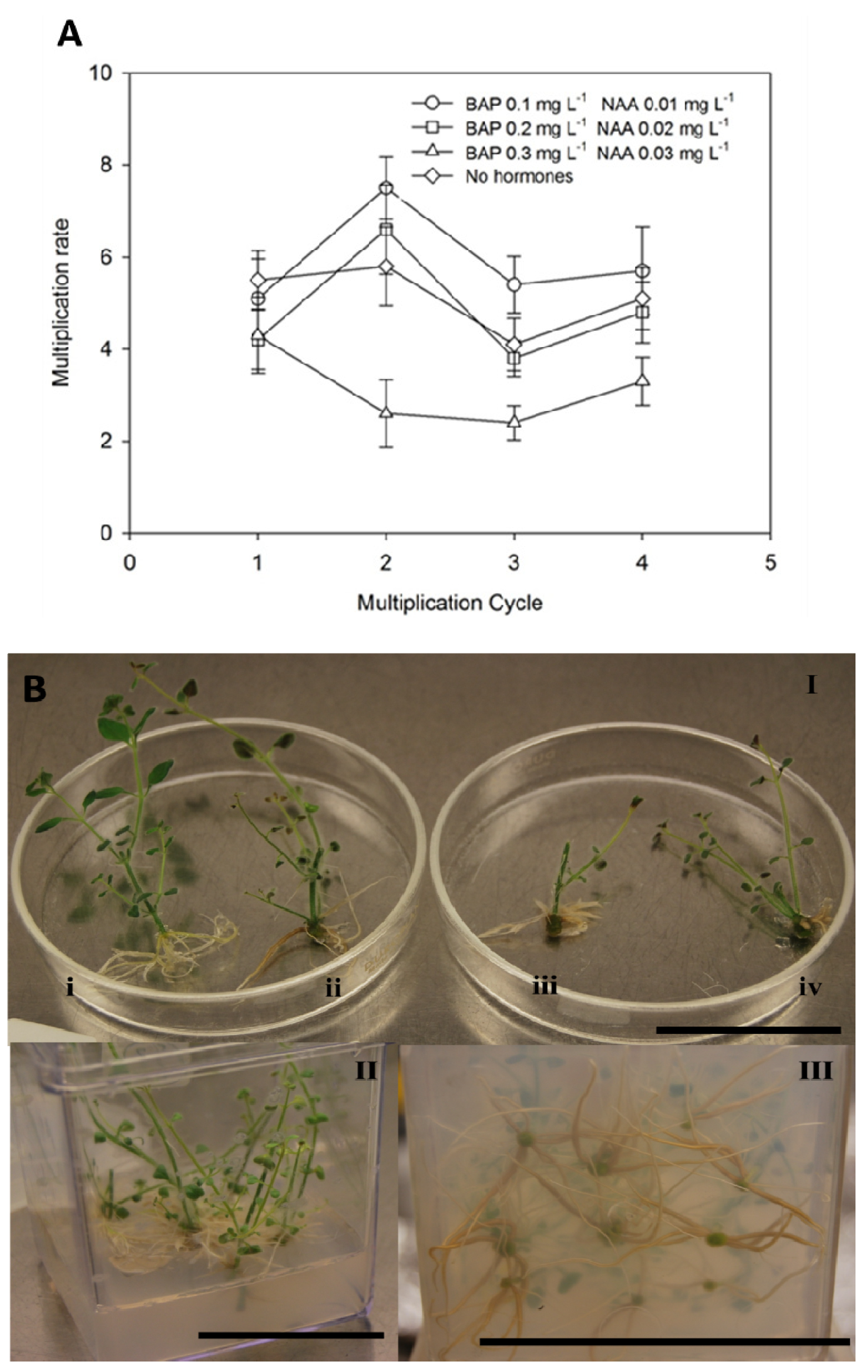
2.4.2. Shoot Multiplication
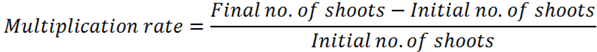
2.4.3. Rooting and Acclimatization
2.5. Statistical Analysis
3. Results and Discussion
3.1. Chemical Characterization and Antioxidant Capacity of Wild A. lopesianum

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Parameters | I | II |
|---|---|---|
| Total phenolic content (mg GAE g−1 dw) | 5.545 ± 0.20 | 6.132 ± 0.56 |
| Flavonoid content (mg CE g−1 dw) | 0.673 ± 0.04 | 0.705 ± 0.04 |
| Antioxidant capacity (nmol TE 100 g−1 dw) | 29.115 ± 3.72 | 11.956 ± 1.16 ** |
3.2. Bioactivities Assessment
| Hydroethanolic extract | Polyphenol-enriched fraction | |
|---|---|---|
| % AChE inhibition (2 mg mL−1) | 37.48 ± 9.04 | 63.66 ± 7.11 |
| % AChE inhibition (1 mg mL−1) | 25.00 ± 9.70 | 32.57 ± 9.02 |
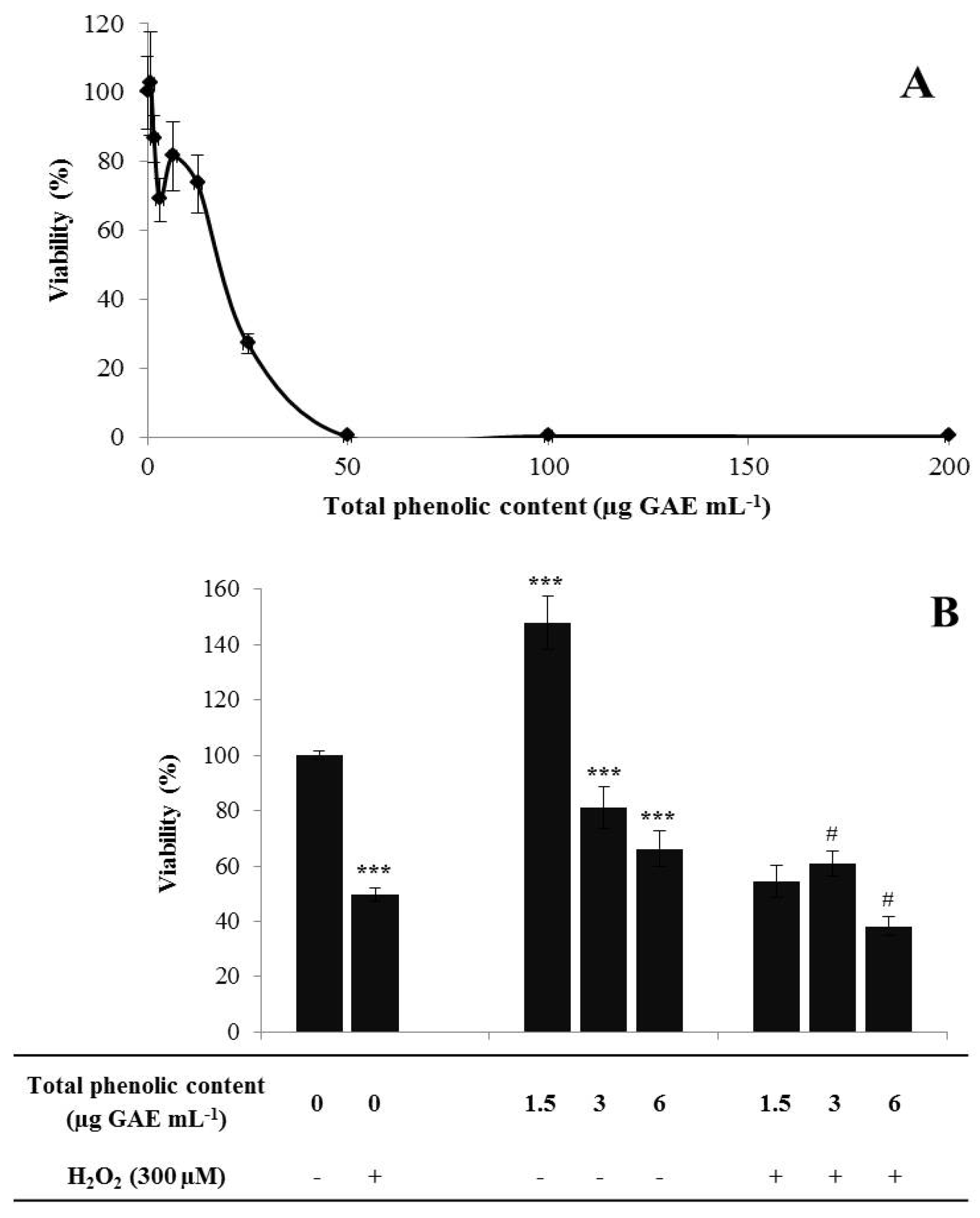
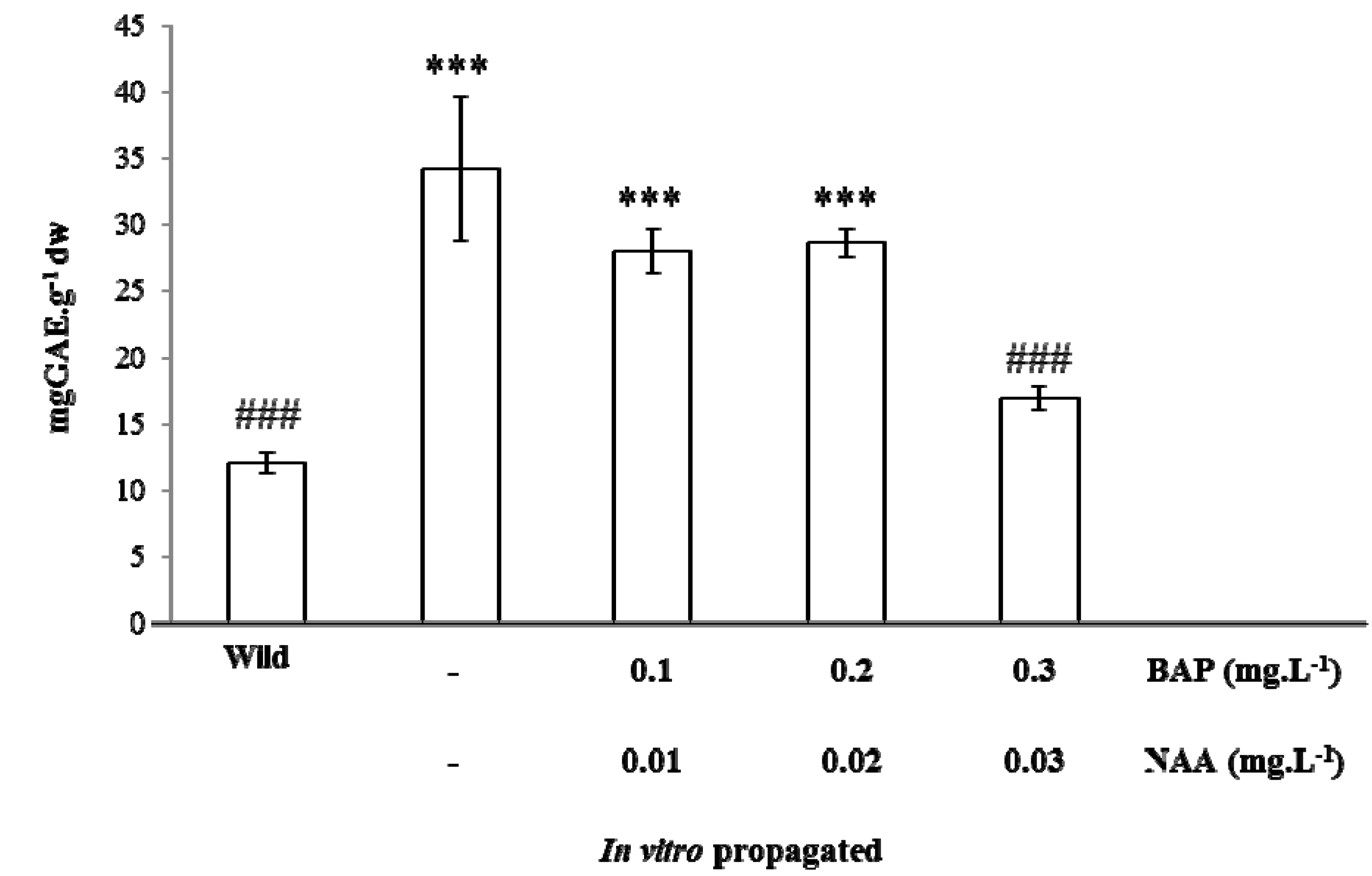
3.3. Ex situ Conservation and in vitro Propagation
Chemical Characterization and Antioxidant Capacity of in vitro Propagated Plants


4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M.; Snader, K.M. Natural products as sources of new drugs over the period 1981–2002. J. Nat. Prod. 2003, 66, 1022–1037. [Google Scholar] [CrossRef]
- Yuliana, N.D.; Khatib, A.; Choi, Y.H.; Verpoorte, R. Metabolomics for bioactivity assessment of natural products. Phytother. Res. 2011, 25, 157–169. [Google Scholar] [PubMed]
- Saklani, A.; Kutty, S.K. Plant-derived compounds in clinical trials. Drug Discov. Today 2008, 13, 161–171. [Google Scholar] [CrossRef]
- Lee, D.H.; Iwanski, G.B.; Thoennissen, N.H. Cucurbitacin: Ancient compound shedding new light on cancer treatment. Sci. World J. 2010, 10, 413–418. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the last 25 years. J. Nat. Prod. 2007, 70, 461–477. [Google Scholar] [CrossRef]
- Mandel, S.; Amit, T.; Reznichenko, L.; Weinreb, O.; Youdim, M.B.H. Green tea catechins as brain-permeable, natural iron chelators-antioxidants for the treatment of neurodegenerative disorders. Mol. Nutr. Food. Res. 2006, 50, 229–234. [Google Scholar] [CrossRef]
- Ramassamy, C. Emerging role of polyphenolic compounds in the treatment of neurodegenerative diseases: A review of their intracellular targets. Eur. J. Pharmacol. 2006, 545, 51–64. [Google Scholar] [CrossRef]
- Williams, P.; Sorribas, A.; Howes, M.-J.R. Natural products as a source of Alzheimer’s drug leads. Nat. Prod. Rep. 2011, 28, 48–77. [Google Scholar] [CrossRef]
- Beking, K.; Vieira, A. Flavonoid intake and disability-adjusted life years due to Alzheimer’s and related dementias: A population-based study involving twenty-three developed countries. Public Health Nutr. 2010, 13, 1403–1409. [Google Scholar] [CrossRef]
- Wimo, A.; Jonsson, L.; Gustavsson, A.; McDaid, D.; Ersek, K.; Georges, J.; Gulacsi, L.; Karpati, K.; Kenigsberg, P.; Valtonen, H. The economic impact of dementia in Europe in 2008-cost estimates from the Eurocode project. Int. J. Geriatr. Psychiatry 2011, 26, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.K. Oxidative stress in neurodegeneration: Cause or consequence? Nat. Med. 2004, 10, S18–S25. [Google Scholar] [CrossRef]
- Bachurin, S.O. Medicinal chemistry approaches for the treatment and prevention of Alzheimer’s disease. Med. Chem. Res. 2003, 23, 48–88. [Google Scholar]
- Thanvi, B.R.; Lo, T.C.N. Update on myasthenia gravis. Postgrad. Med. J. 2004, 80, 690–700. [Google Scholar] [CrossRef]
- Soreq, H.; Seidman, S. Acetylcholinesterase—New roles for an old actor. Nat. Rev. Neurosci. 2001, 2, 294–302. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Kumar, V.; Mal, M.; Houghton, P.J. Acetylcholinesterase inhibitors from plants. Phytomedicine 2007, 14, 289–300. [Google Scholar] [CrossRef]
- Nordberg, A.; Svensson, A.L. Cholinesterase inhibitors in the treatment of Alzheimer’s disease: A comparison of tolerability and pharmacology. Drug Saf. 1998, 19, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Perry, G.; Nunomura, A.; Hirai, K.; Zhu, X.; Perez, M.; Avila, J.; Castellani, R.J.; Atwood, C.S.; Aliev, G.; Sayre, L.M.; et al. Is oxidative damage the fundamental pathogenic mechanism of Alzheimer’s and other neurodegenerative diseases? Free Radic. Biol. Med. 2002, 33, 1475–1479. [Google Scholar] [CrossRef] [PubMed]
- Vargas, P.; Carrió, E.; Guzmán, B.; Amat, E.; Güemes, J. A geographical pattern of Antirrhinum (Scrophulariaceae) speciation since the Pliocene based on plastid and nuclear DNA polymorphisms. J. Biogeogr. 2009, 36, 1297–1312. [Google Scholar] [CrossRef]
- Amich García, F.; Bernardos Hernández, S.; González Talaván, A.; Barreto Caldas, F.; Alves, P. IUCN Red List of Threatened Species, Version 2013.1. >Antirrhinum lopesianum. Available online: http://www.iucnredlist.org (accessed on 15 July 2013).
- Amich, F.; Rodríguez, J.A.S.; Gallego, F.; Anta, M.S. Antirrhinum lopesianum Rothm., novedad para la flora española. Bol. Soc. Brot. 1989, 62, 231–237. [Google Scholar]
- Coutinho, A.X.P. Apontamentos para o estudo da flora transmontana. Bol. Soc. Brot. 1883, 2, 129–163. [Google Scholar]
- Lopes, J.M.M. A flora do concelho de Vimioso e arredores. Bol. Soc. Brot. 1926, 4, 234–278. [Google Scholar]
- Rothmaler, W. Taxonomische Monographie der Gattung Antirrhinum; Akademie Verlag: Berlin, Germany, 1956. [Google Scholar]
- Aguiar, C. Distribuição Geográfica e Estatuto de Ameaça das espécies da Flora a Proteger; ESAB: Bragança, Portugal, 1996. [Google Scholar]
- Bernardos, S.; Amado, A.; Aguiar, C.; Santos, C.; Fernández-Diez, J.; González-Talaván, A.; Amich, F. Conservation status of the threatened Iberian Peninsula narrow endemic Antirrhinum lopesianum Rothm. (Scrophulariaceae). Plant Biosyst. 2006, 140, 2–9. [Google Scholar] [CrossRef]
- Anonymous. Directive 92/43 of the Council of the European Community on the Conservation of Habitats and Wild Fauna and Flora; European Community: Brussels, Belgium, 1992. [Google Scholar]
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R.V. European Red List of Vascular Plants; Publications Office of the European Union: Luxembourg, Luxembourg, 2011. [Google Scholar]
- Almeida, R.; Gonçalves, S.; Romano, A. In vitro micropropagation of endangered Rhododendron ponticum L. subsp. baeticum (Boissier & Reuter) Handel-Mazzetti. Biodivers. Conserv. 2005, 14, 1059–1069. [Google Scholar] [CrossRef]
- Guo, B.; Gao, M.; Liu, C.-Z. In vitro propagation of an endangered medicinal plant Saussurea involucrata Kar. et Kir. Plant Cell Rep. 2007, 26, 261–265. [Google Scholar] [CrossRef]
- Ket, N.V.; Hahn, E.J.; Park, S.Y.; Chakrabarty, D.; Paek, K.Y. Micropropagation of an endangered orchid Anoectochilus formosanus. Biol. Plant. 2004, 48, 339–344. [Google Scholar] [CrossRef]
- Mao, A.; Kaliamoorthy, S.; Ranyaphi, R.; Das, J.; Gupta, S.; Athili, J.; Yumnam, J.; Chanu, L. In vitro micropropagation of three rare, endangered, and endemic rhododendron species of Northeast India. In Vitro Cell Dev.Biol. Plant 2011, 47, 674–681. [Google Scholar] [CrossRef]
- Piovan, A.; Cassina, G.; Filippini, R. Crambe tataria: Actions for ex situ conservation. Biodivers. Conserv. 2011, 20, 359–371. [Google Scholar] [CrossRef]
- Remya, M.; Narmatha Bai, V.; Mutharaian, V. In vitro regeneration of Aristolochia tagala and production of artificial seeds. Biol. Plant. 2013, 57, 210–218. [Google Scholar] [CrossRef]
- Tavares, L.; Fortalezas, S.; Carrilho, C.; McDougall, G.J.; Stewart, D.; Ferreira, R.B.; Santos, C.N. Antioxidant and antiproliferative properties of strawberry tree tissues. J. Berry Res. 2010, 1, 3–12. [Google Scholar]
- Fortalezas, S.; Tavares, L.; Pimpão, R.; Tyagi, M.; Pontes, V.; Alves, P.M.; McDougall, G.; Stewart, D.; Ferreira, R.B.; Santos, C.N. Antioxidant properties and neuroprotective capacity of strawberry tree fruit (Arbutus unedo). Nutrients 2010, 2, 214–229. [Google Scholar] [CrossRef] [PubMed]
- Tavares, L.; Carrilho, D.; Tyagi, M.; Barata, D.; Serra, A.T.; Duarte, C.M.M.; Duarte, R.O.; Feliciano, R.P.; Bronze, M.R.; Chicau, P.; et al. Antioxidant capacity of Macaronesian traditional medicinal plants. Molecules 2010, 15, 2576–2592. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V.J.; Feather-Stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Tavares, L.; Fortalezas, S.; Tyagi, M.; Barata, D.; Serra, A.; Duarte, C.; Duarte, R.; Feliciano, R.; Bronze, M.; Espírito-Santo, M.; et al. Bioactive compounds from endemic plants of Southwest Portugal: Inhibition of acetylcholinesterase and radical scavenging activities. Pharma Biol. 2011, 50, 239–246. [Google Scholar]
- Tavares, L.; McDougall, G.J.; Fortalezas, S.; Stewart, D.; Ferreira, R.B.; Santos, C.N. The neuroprotective potential of phenolic-enriched fractions from four Juniperus species found in Portugal. Food Chem. 2012, 135, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Nacif de Abreu, I.; Mazzafera, P. Effect of water and temperature stress on the content of active constituents of Hypericum brasiliense choisy. Plant Physiol. Biochem. 2005, 43, 241–248. [Google Scholar] [CrossRef]
- Lin, K.-H.; Chao, P.-Y.; Yang, C.-M.; Cheng, W.-C.; Lo, H.-F.; Chang, T.-R. The effects of flooding and drought stresses on the antioxidant constituents in sweet potato leaves. Bot. Stud. 2006, 47, 417–426. [Google Scholar]
- Ross, H.A.; McDougall, G.J.; Stewart, D. Antiproliferative activity is predominantly associated with ellagitannins in raspberry extracts. Phytochemistry 2007, 68, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Bammidi, S.R.; Volluri, S.S.; Chippada, S.C.; Avanigadda, S.; Vangalapati, M. A review on pharmacological studies of Bacopa monniera. J. Chem. Biol. Phys. Sci. 2011, 1, 250–259. [Google Scholar]
- Kolak, U.; Boga, M.; Urusak, E.A.; Ulubelen, A. Constituents of Plantago major subsp. intermedia with antioxidant and anticholinesterase capacities. Turk. J. Chem. 2011, 35, 637–645. [Google Scholar]
- Chung, Y.K.; Heo, H.J.; Kim, E.K.; Kim, H.K.; Huh, T.L.; Lim, Y.; Kim, S.K.; Shin, D.H. Inhibitory effect of ursolic acid purified from Origanum majorana L. on the acetylcholinesterase. Mol. Cells 2001, 11, 137–143. [Google Scholar] [PubMed]
- El-Hassan, A.; El-Sayed, M.; Hamed, A.I.; Rhee, I.K.; Ahmed, A.A.; Zeller, K.P.; Verpoorte, R. Bioactive constituents of Leptadenia arborea. Fitoterapia 2003, 74, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, S.; Halliwell, B. Do polyphenols enter the brain and does it matter? Some theoretical and practical considerations. Genes Nutr. 2012, 7, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Genebank Standards; Food and Agriculture Organization of the United Nations, International Plant Genetic Resources Institute: Rome, Italy, 1994.
- Atkinson, N.J.; Ford-Lloyd, B.V.; John Newbury, H. Regeneration of plants from Antirrhinum majus L. callus. Plant Cell Tissue Org. 1989, 17, 59–70. [Google Scholar]
- Gonzalez-Benito, M.E.; Tapia, J.; Rodriguez, N.; Iriondo, J.M. Micropropagation of commercial and wild genotypes of snapdragon (Antirrhinum spp.). J. Hortic. Sci. 1996, 71, 11–15. [Google Scholar]
- Sangwan, R.S.; Detrez, C.; Sangwan-Norreel, B.S. In vitro culture of shoot-tip meristems in some higher plants. Acta Hortic. (ISHS) 1987, 212, 661–666. [Google Scholar]
- Arnold, S.V.; Eriksson, T. Induction of adventitious buds on embryos of Norway spruce grown in vitro. Physiol. Plant 1978, 44, 283–287. [Google Scholar] [CrossRef]
- Qiu, J.; Xue, X.; Chen, F.; Li, C.; Bolat, N.; Wang, X.; Baima, Y.; Zhao, Q.; Zhao, D.; Ma, F. Quality evaluation of snow lotus (Saussurea): quantitative chemical analysis and antioxidant activity assessment. Plant Cell Rep. 2010, 29, 1325–1337. [Google Scholar] [CrossRef] [PubMed]
- Danova, K.; Čellárová, E.; Macková, A.; Daxnerová, Z.; Kapchina-Toteva, V. In vitro culture of Hypericum rumeliacum Boiss. and production of phenolics and flavonoids. In Vitro Cell Dev. Biol. Plant 2010, 46, 422–429. [Google Scholar] [CrossRef]
- Robards, K.; Antolovich, M. Analytical chemistry of fruit bioflavonoids—A review. Analyst 1997, 122, 11R–34R. [Google Scholar] [CrossRef]
- Amat, A.; Clementi, C.; De Angelis, F.; Sgamellotti, A.; Fantacci, S. Absorption and emission of the apigenin and luteolin flavonoids: A TDDFT investigation. J. Phys. Chem. A 2009, 113, 15118–15126. [Google Scholar] [CrossRef] [PubMed]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gomes, A.; Fortalezas, S.; Pimpão, R.; Figueira, I.; Maroco, J.; Aguiar, C.; Ferreira, R.B.; Miguel, C.; Santos, C.N. Valuing the Endangered Species Antirrhinum lopesianum: Neuroprotective Activities and Strategies for in vitro Plant Propagation. Antioxidants 2013, 2, 273-292. https://doi.org/10.3390/antiox2040273
Gomes A, Fortalezas S, Pimpão R, Figueira I, Maroco J, Aguiar C, Ferreira RB, Miguel C, Santos CN. Valuing the Endangered Species Antirrhinum lopesianum: Neuroprotective Activities and Strategies for in vitro Plant Propagation. Antioxidants. 2013; 2(4):273-292. https://doi.org/10.3390/antiox2040273
Chicago/Turabian StyleGomes, Andreia, Sofia Fortalezas, Rui Pimpão, Inês Figueira, João Maroco, Carlos Aguiar, Ricardo B. Ferreira, Célia Miguel, and Cláudia N. Santos. 2013. "Valuing the Endangered Species Antirrhinum lopesianum: Neuroprotective Activities and Strategies for in vitro Plant Propagation" Antioxidants 2, no. 4: 273-292. https://doi.org/10.3390/antiox2040273