Hippocampal Physiology, Structure and Function and the Neuroscience of Schizophrenia: A Unified Account of Declarative Memory Deficits, Working Memory Deficits and Schizophrenic Symptoms

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Review—Memory, the Hippocampus and Schizophrenia
2.1. Hippocampal Function, Anatomy and Physiology as It Relates to Schizophrenia
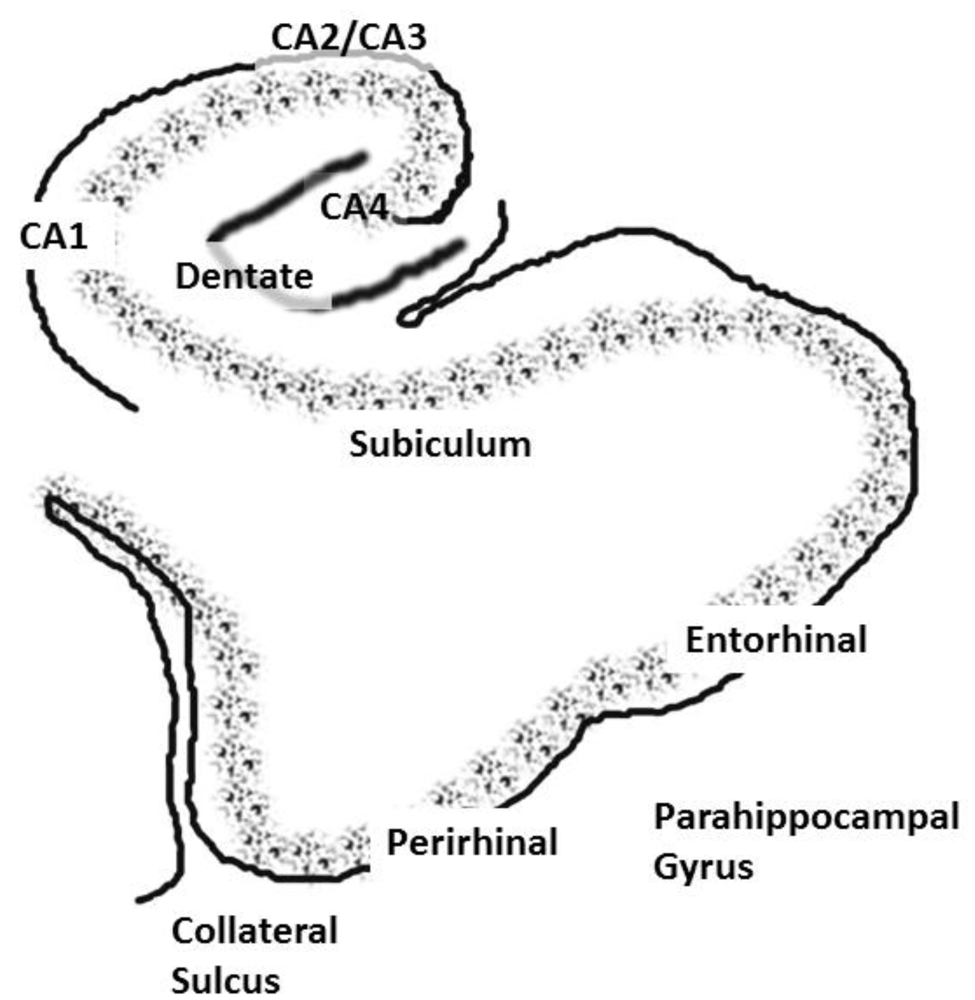
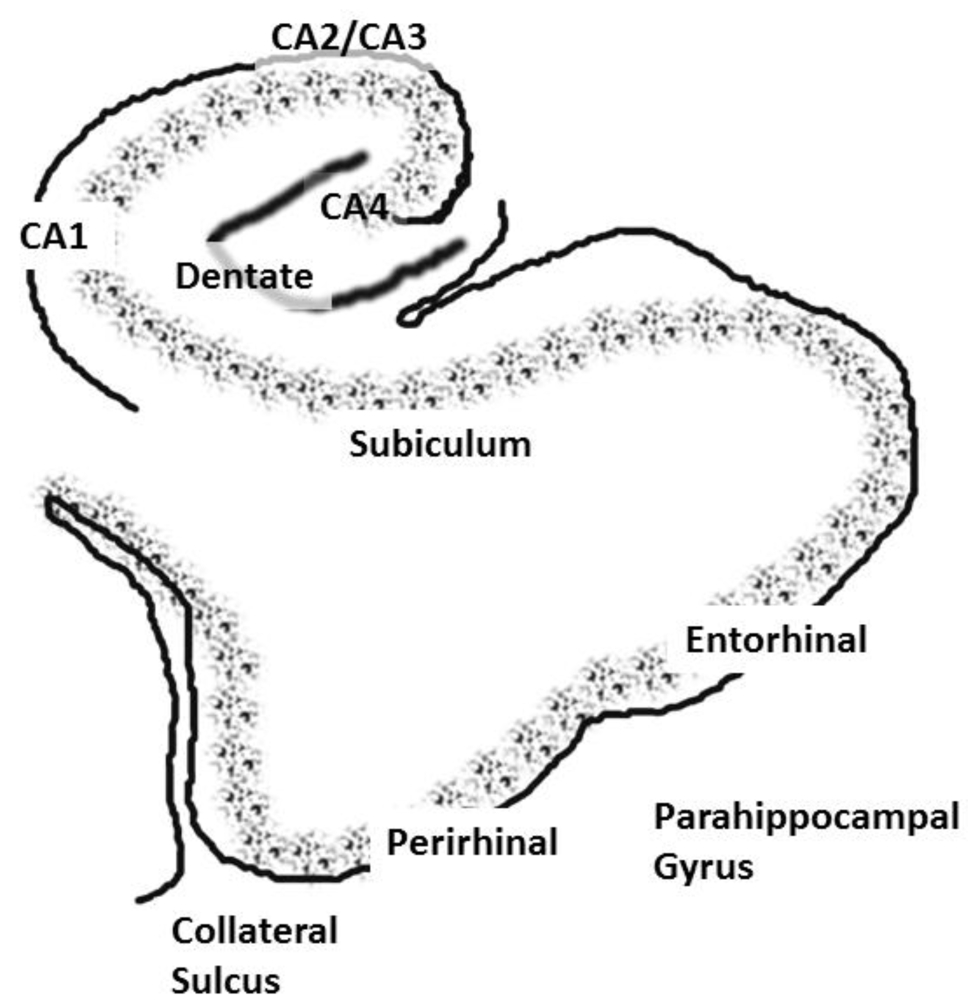
2.2. The Interconnectivity of the Hippocampal System: Functional Consequences and Implications for Schizophrenia
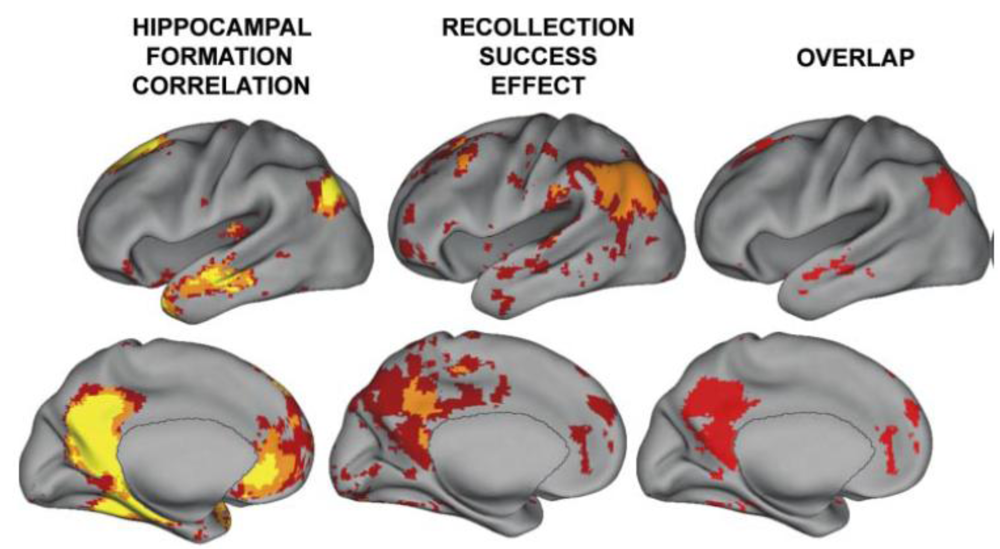
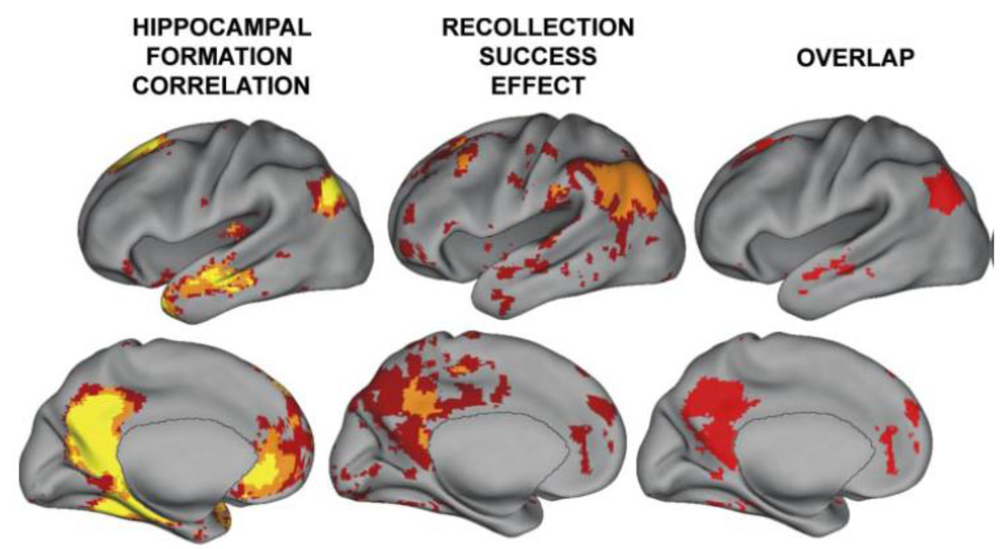
2.3. Hippocampal—TPJ Interaction and the Symptoms of Schizophrenia
3. Summary and Conclusions
Conflict of Interest
Acknowledgements
References
- Cirillo, M.A.; Seidman, L.J. Verbal declarative memory dysfunction in schizophrenia: from clinical assessment to genetics and brain mechanisms. Neuropsychol. Rev. 2003, 13, 43–77. [Google Scholar]
- Goldman-Rakic, P.S. Working memory dysfunction in schizophrenia. J. Neuropsychiatry Clin. Neurosci. 1994, 6, 348–357. [Google Scholar]
- Warren, D.E.; Duff, M.C.; Jensen, U.; Tranel, D.; Cohen, N.J. Hiding in plain view: Lesions of the medial temporal lobe impair online representation. Hippocampus 2012, 22, 1577–1588. [Google Scholar]
- Watanabe, T.; Niki, H. Hippocampal unit activity and delayed response in the monkey. Brain Res. 1985, 325, 241–254. [Google Scholar]
- Hannula, D.E.; Tranel, D.; Cohen, N.J. The long and the short of it: Relational memory impairments in amnesia, Even at short lags. J. Neurosci. 2006, 26, 8352–8359. [Google Scholar]
- Wible, C.G.; Shenton, M.E.; McCarley, R.W. Functional neuroanatomy of the limbic system and planum temporale. In Brain Imaging in Clinical Psychiatry; Krishnan, R.R., Doraiswamy, P.M., Eds.; Marcel Dekker: New York, NY, USA, 1997; pp. 63–101. [Google Scholar]
- Small, S.A.; Schobel, S.A.; Buxton, R.B.; Witter, M.P.; Barnes, C.A. A pathophysiological framework of hippocampal dysfunction in ageing and disease. Nat. Rev. Neurosci. 2011, 12, 585–601. [Google Scholar]
- Wible, C.G. Hippocampal temporal-parietal junction interaction in the production of psychotic pymptoms: A framework for understanding the schizophrenic syndrome. Front. Neurosci. 2012, 6, 1–30. [Google Scholar]
- Saykin, A.J.; Gur, R.C.; Gur, R.E.; Mozley, P.D.; Mozley, L.H.; Resnick, S.M.; Kester, D.B.; Stafiniak, P. Neuropsychological function in schizophrenia. Selective impairment in memory and learning. Arch. Gen. Psychiatr. 1991, 48, 618–624. [Google Scholar]
- Aleman, A.; Hijman, R.; de Haan, E.H.; Kahn, R.S. Memory impairment in schizophrenia: A meta-analysis. Am. J. Psychiatr. 1999, 156, 1358–1366. [Google Scholar]
- Eichenbaum, H.C.N.; Otto, T.; Wible, C.G. Memory representation in the hippocampus: Functional domain and functional organization. In Memory: Organization and Locus of Change; Squire, L.R., Weinberger, N.M., McGaugh, J.L., Eds.; Oxford University Press: New York, NY, USA, 1991. [Google Scholar]
- Seidman, L.J.; Pantelis, C.; Keshavan, M.S.; Faraone, S.V.; Goldstein, J.M.; Horton, N.J.; Makris, N.; Falkai, P.; Caviness, V.S.; Tsuang, M.T. A review and new report of medial temporal lobe dysfunction as a vulnerability indicator for schizophrenia: A magnetic resonance imaging morphometric family study of the parahippocampal gyrus. Schizophr. Bull. 2003, 29, 803–830. [Google Scholar]
- Heckers, S. Neuroimaging studies of the hippocampus in schizophrenia. Hippocampus 2001, 11, 520–528. [Google Scholar]
- Velakoulis, D.; Wood, S.J.; Wong, M.T.; McGorry, P.D.; Yung, A.; Phillips, L.; Smith, D.; Brewer, W.; Proffitt, T.; Desmond, P.; et al. Hippocampal and amygdala volumes according to psychosis stage and diagnosis: A magnetic resonance imaging study of chronic schizophrenia, First-episode psychosis, And ultra-high-risk individuals. Arch. Gen. Psychiatr. 2006, 63, 139–149. [Google Scholar]
- Heckers, S.; Konradi, C. Hippocampal neurons in schizophrenia. J. Neural. Transm. 2002, 109, 891–905. [Google Scholar]
- Heckers, S.; Rauch, S.L.; Goff, D.; Savage, C.R.; Schacter, D.L.; Fischman, A.J.; Alpert, N.M. Impaired recruitment of the hippocampus during conscious recollection in schizophrenia. Nat. Neurosci. 1998, 1, 318–323. [Google Scholar]
- Cohen, N.J.; Ryan, J.; Hunt, C.; Romine, L.; Wszalek, T.; Nash, C. Hippocampal system and declarative (relational) memory: Summarizing the data from functional neuroimaging studies. Hippocampus 1999, 9, 83–98. [Google Scholar]
- Amaral, D.G. Emerging principles of intrinsic hippocampal organization. Curr. Opin. Neurobiol. 1993, 3, 225–229. [Google Scholar]
- Insausti, R.; Amaral, D.G.; Cowan, W.M. The entorhinal cortex of the monkey: II. Cortical afferents. J. Comp. Neurol. 1987, 264, 356–395. [Google Scholar] [CrossRef]
- Suzuki, W.A.; Amaral, D.G. Perirhinal and parahippocampal cortices of the macaque monkey: Cortical afferents. J. Comp. 1994, 350, 497–533. [Google Scholar]
- Witter, M.P.; Amaral, D.G. Entorhinal cortex of the monkey: V. Projections to the dentate gyrus, hippocampus, and subicular complex. J. Comp. Neurol. 1991, 307, 437–459. [Google Scholar] [CrossRef]
- Benes, F.M. Myelination of cortical-hippocampal relays during late adolescence. Schizophr. Bull. 1989, 15, 585–593. [Google Scholar]
- Suzuki, M.; Hagino, H.; Nohara, S.; Zhou, S.Y.; Kawasaki, Y.; Takahashi, T.; Matsui, M.; Seto, H.; Ono, T.; Kurachi, M. Male-specific volume expansion of the human hippocampus during adolescence. Cereb. Cortex. 2005, 15, 187–193. [Google Scholar]
- Lavenex, P.; Amaral, D.G. Hippocampal-neocortical interaction: A hierarchy of associativity. Hippocampus 2000, 10, 420–430. [Google Scholar]
- Lavenex, P.; Suzuki, W.A.; Amaral, D.G. Perirhinal and parahippocampal cortices of the macaque monkey: Projections to the neocortex. J. Neurol. 2002, 447, 394–420. [Google Scholar]
- Suzuki, W.A.; Amaral, D.G. Perirhinal and parahippocampal cortices of the macaque monkey: Cortical afferents. J. Comp. Neurol. 1994, 350, 497–533. [Google Scholar]
- Sloviter, R.S.; Lomo, T. Updating the lamellar hypothesis of hippocampal organization. Front. Neural Circuit. 2012, 6, 102. [Google Scholar]
- Insausti, R.; Tunon, T.; Sobreviela, T.; Insausti, A.M.; Gonzalo, L.M. The human entorhinal cortex: A cytoarchitectonic analysis. J. Comp. Neurol. 1995, 355, 171–198. [Google Scholar]
- Kartsounis, L.D.; Rudge, P.; Stevens, J.M. Bilateral lesions of CA1 and CA2 fields of the hippocampus are sufficient to cause a severe amnesic syndrome in humans. J. Neurol. Neurosurg. Psychiatr. 1995, 59, 95–98. [Google Scholar]
- Rempel-Clower, N.L.; Zola, S.M.; Squire, L.R.; Amaral, D.G. Three cases of enduring memory impairment after bilateral damage limited to the hippocampal formation. J. Neurosci. 1996, 16, 5233–5255. [Google Scholar]
- Zola-Morgan, S.; Squire, L.R.; Amaral, D.G. Human amnesia and the medial temporal region: Enduring memory impairment following a bilateral lesion limited to field CA1 of the hippocampus. J. Neurosci. 1986, 6, 2950–2967. [Google Scholar]
- Zola-Morgan, S.; Squire, L.R.; Rempel, N.L.; Clower, R.P.; Amaral, D.G. Enduring memory impairment in monkeys after ischemic damage to the hippocampus. J. Neurosci. 1992, 12, 2582–2596. [Google Scholar]
- Reed, J.M.; Squire, L.R. Retrograde amnesia for facts and events: findings from four new cases. J. Neurosci. 1998, 18, 3943–3954. [Google Scholar]
- Epp, J.R.; Chow, C.; Galea, L.A. Hippocampus-dependent learning influences hippocampal neurogenesis. Front. Neurosci. 2013, 7, 57. [Google Scholar]
- Cotman, C.W.; Monaghan, D.T. Anatomical organization of excitatory amino acid receptors and their properties. Adv. Exp. Med. 1986, 203, 237–252. [Google Scholar]
- Bliss, T.V.; Lomo, T. Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J. Physiol. 1973, 232, 331–356. [Google Scholar]
- Schobel, S.A.; Lewandowski, N.M.; Corcoran, C.M.; Moore, H.; Brown, T.; Malaspina, D.; Small, S.A. Differential targeting of the CA1 subfield of the hippocampal formation by schizophrenia and related psychotic disorders. Arch. Gen. Psychiatr. 2009, 66, 938–946. [Google Scholar]
- Eichenbaum, H. Hippocampus: Cognitive processes and neural representations that underlie declarative memory. Neuron 2004, 44, 109–120. [Google Scholar]
- Fried, I.; Cameron, K.A.; Yashar, S.; Fong, R.; Morrow, J.W. Inhibitory and excitatory responses of single neurons in the human medial temporal lobe during recognition of faces and objects. Cereb. Cortex. 2002, 12, 575–584. [Google Scholar]
- Halgren, E.; Babb, T.L.; Crandall, P.H. Activity of human hippocampal formation and amygdala neurons during memory testing. Electroencephalogr. Clin. Neurophysiol. 1978, 45, 585–601. [Google Scholar]
- Quiroga, R.Q.; Kreiman, G.; Koch, C.; Fried, I. Sparse but not ‘grandmother-cell’ coding in the medial temporal lobe. Trends Cogn. Sci. 2008, 12, 87–91. [Google Scholar]
- Quiroga, R.Q.; Reddy, L.; Kreiman, G.; Koch, C.; Fried, I. Invariant visual representation by single neurons in the human brain. Nature 2005, 435, 1102–1107. [Google Scholar]
- O'Keefe, J. A review of the hippocampal place cells. Prog. Neurobiol. 1979, 13, 419–439. [Google Scholar]
- Wible, C.G.; Findling, R.L.; Shapiro, M.; Lang, E.J.; Crane, S.; Olton, D.S. Mnemonic correlates of unit activity in the hippocampus. Brain Res. 1986, 399, 97–110. [Google Scholar]
- Riches, I.P.; Wilson, F.A.; Brown, M.W. The effects of visual stimulation and memory on neurons of the hippocampal formation and the neighboring parahippocampal gyrus and inferior temporal cortex of the primate. J. Neurosci. 1991, 11, 1763–1779. [Google Scholar]
- Weiss, C.; Bouwmeester, H.; Power, J.M.; Disterhoft, J.F. Hippocampal lesions prevent trace eyeblink conditioning in the freely moving rat. Behav. Brain Res. 1999, 99, 123–132. [Google Scholar]
- Thompson, L.T.; Best, P.J. Long-term stability of the place-field activity of single units recorded from the dorsal hippocampus of freely behaving rats. Brain Res. 1990, 509, 299–308. [Google Scholar]
- Muller, R.U.; Kubie, J.L.; Ranck, J.B., Jr. Spatial firing patterns of hippocampal complex-spike cells in a fixed environment. J. Neurosci. 1987, 7, 1935–1950. [Google Scholar]
- Cohen, N.J.; Poldrack, R.A.; Eichenbaum, H. Memory for items and memory for relations in the procedural/declarative memory framework. Memory 1997, 5, 131–178. [Google Scholar]
- Williams, L.E.; Must, A.; Avery, S.; Woolard, A.; Woodward, N.D.; Cohen, N.J.; Heckers, S. Eye-movement behavior reveals relational memory impairment in schizophrenia. Biol. Psychiatr. 2010, 68, 617–624. [Google Scholar]
- Buonomano, D.V.; Merzenich, M.M. Cortical plasticity: from synapses to maps. Annu. Rev. Neurosci. 1998, 21, 149–186. [Google Scholar]
- Javitt, D.C. When doors of perception close: Bottom-up models of disrupted cognition in schizophrenia. Annu. Rev. Clin. Psychol. 2009, 5, 249–275. [Google Scholar]
- Wiebe, S. Epidemiology of temporal lobe epilepsy. In Can. J. Neurol. Sci. 2000; 27 (Suppl. 1), pp. S6–S10; discussion S20–S21. [Google Scholar]
- Griesemer, D.; Mautes, A.M. Closed head injury causes hyperexcitability in rat hippocampal CA1 but not in CA3 pyramidal cells. J. Neurotrauma. 2007, 24, 1823–1832. [Google Scholar]
- Villanueva, V.; Serratosa, J.M. Temporal lobe epilepsy: Clinical semiology and age at onset. Epileptic Disord. 2005, 7, 83–90. [Google Scholar]
- Sapolsky, R.M.; Uno, H.; Rebert, C.S.; Finch, C.E. Hippocampal damage associated with prolonged glucocorticoid exposure in primates. J. Neurosci. 1990, 10, 2897–2902. [Google Scholar]
- Sandi, C. Glucocorticoids act on glutamatergic pathways to affect memory processes. Trends Neurosci. 2011, 34, 165–176. [Google Scholar] [CrossRef]
- Clarke, M.C.; Tanskanen, A.; Huttunen, M.O.; Clancy, M.; Cotter, D.R.; Cannon, M. Evidence for shared susceptibility to epilepsy and psychosis: A population-based family study. Biol. Psychiatr. 2012, 71, 836–839. [Google Scholar] [CrossRef]
- Briellmann, R.S.; Kalnins, R.M.; Hopwood, M.J.; Ward, C.; Berkovic, S.F.; Jackson, G.D. TLE patients with postictal psychosis: mesial dysplasia and anterior hippocampal preservation. Neurology 2000, 55, 1027–1030. [Google Scholar]
- Nishida, T.; Kudo, T.; Inoue, Y.; Nakamura, F.; Yoshimura, M.; Matsuda, K.; Yagi, K.; Fujiwara, T. Postictal mania versus postictal psychosis: Differences in clinical features, Epileptogenic zone, And brain functional changes during postictal period. Epilepsia 2006, 47, 2104–2114. [Google Scholar] [CrossRef]
- Tarulli, A.; Devinsky, O.; Alper, K. Progression of postictal to interictal psychosis. Epilepsia 2001, 42, 1468–1471. [Google Scholar]
- Elliott, B.; Joyce, E.; Shorvon, S. Delusions, Illusions and hallucinations in epilepsy: 2. Complex phenomena and psychosis. Epilepsy Res. 2009, 85, 172–186. [Google Scholar] [CrossRef]
- Verhoeven, W.M.; Egger, J.I.; Gunning, W.B.; Bevers, M.; de Pont, B.J. Recurrent schizophrenia-like psychosis as first manifestation of epilepsy: A diagnostic challenge in neuropsychiatry. Neuropsychiatr. Dis. Treat. 2010, 6, 227–231. [Google Scholar]
- Woo, T.U.; Spencer, K.; McCarley, R.W. Gamma oscillation deficits and the onset and early progression of schizophrenia. Harv Rev Psychiatr. 2010, 18, 173–189. [Google Scholar] [CrossRef]
- Li, K.X.; Lu, Y.M.; Xu, Z.H.; Zhang, J.; Zhu, J.M.; Zhang, J.M.; Cao, S.X.; Chen, X.J.; Chen, Z.; Luo, J.H.; et al. Neuregulin 1 regulates excitability of fast-spiking neurons through Kv1.1 and acts in epilepsy. Nat. Neurosci. 2011, 15, 267–273. [Google Scholar] [CrossRef]
- Allen, P.; Laroi, F.; McGuire, P.K.; Aleman, A. The hallucinating brain: A review of structural and functional neuroimaging studies of hallucinations. Neurosci. Biobehav. Rev. 2008, 32, 175–191. [Google Scholar] [CrossRef]
- Woodward, N.D.; Rogers, B.; Heckers, S. Functional resting-state networks are differentially affected in schizophrenia. Schizophr. Res. 2011, 130, 86–93. [Google Scholar] [CrossRef]
- Diederen, K.M.; Neggers, S.F.; Daalman, K.; Blom, J.D.; Goekoop, R.; Kahn, R.S.; Sommer, I.E. Deactivation of the parahippocampal gyrus preceding auditory hallucinations in schizophrenia. Am. J. Psychiatr. 2010, 167, 427–435. [Google Scholar] [CrossRef]
- Jardri, R.; Pouchet, A.; Pins, D.; Thomas, P. Cortical activations during auditory verbal hallucinations in schizophrenia: A coordinate-based meta-analysis. Am. J. Psychiatr. 2011, 168, 73–81. [Google Scholar] [CrossRef]
- Arzy, S.; Mohr, C.; Michel, C.M.; Blanke, O. Duration and not strength of activation in temporo-parietal cortex positively correlates with schizotypy. Neuroimage 2007, 35, 326–333. [Google Scholar] [CrossRef]
- Ffytche, D.H.; Howard, R.J.; Brammer, M.J.; David, A.; Woodruff, P.; Williams, S. The anatomy of conscious vision: An fMRI study of visual hallucinations. Nat. Neurosci. 1998, 1, 738–742. [Google Scholar] [CrossRef]
- Santhouse, A.M.; Howard, R.J.; ffytche, D.H. Visual hallucinatory syndromes and the anatomy of the visual brain. Brain 2000, 123, 2055–2064. [Google Scholar] [CrossRef]
- Kahn, I.; Andrews-Hanna, J.R.; Vincent, J.L.; Snyder, A.Z.; Buckner, R.L. Distinct cortical anatomy linked to subregions of the medial temporal lobe revealed by intrinsic functional connectivity. J. Neurophysiol. 2008, 100, 129–139. [Google Scholar] [CrossRef]
- Vincent, J.L.; Snyder, A.Z.; Fox, M.D.; Shannon, B.J.; Andrews, J.R.; Raichle, M.E.; Buckner, R.L. Coherent spontaneous activity identifies a hippocampal-parietal memory network. J. Neurophysiol. 2006, 96, 3517–3531. [Google Scholar] [CrossRef]
- Buckner, R.L.; Sepulcre, J.; Talukdar, T.; Krienen, F.M.; Liu, H.; Hedden, T.; Andrews-Hanna, J.R.; Sperling, R.A.; Johnson, K.A. Cortical hubs revealed by intrinsic functional connectivity: Mapping, assessment of stability, and relation to Alzheimer's disease. J. Neurosci. 2009, 29, 1860–1873. [Google Scholar] [CrossRef]
- Hutchinson, J.B.; Uncapher, M.R.; Wagner, A.D. Posterior parietal cortex and episodic retrieval: Convergent and divergent effects of attention and memory. Learn. Mem. 2009, 16, 343–356. [Google Scholar] [CrossRef]
- Clower, D.M.; West, R.A.; Lynch, J.C.; Strick, P.L. The inferior parietal lobule is the target of output from the superior colliculus, hippocampus, and cerebellum. J. Neurosci. 2001, 21, 6283–6291. [Google Scholar]
- Rockland, K.S.; Van Hoesen, G.W. Some temporal and parietal cortical connections converge in CA1 of the primate hippocampus. Cereb. Cortex. 1999, 9, 232–237. [Google Scholar] [CrossRef]
- Ghent, L.; Mishkin, M.; Teuber, H.L. Short-term memory after frontal-lobe injury in man. J. Comp. Physiol. Psychol. 1962, 55, 705–709. [Google Scholar] [CrossRef]
- Paulesu, E.; Frith, C.D.; Frackowiak, R.S. The neural correlates of the verbal component of working memory. Nature 1993, 362, 342–345. [Google Scholar] [CrossRef]
- Jonides, J.; Schumacher, E.H.; Smith, E.E.; Koeppe, R.A.; Awh, E.; Reuter-Lorenz, P.A.; Marshuetz, C.; Willis, C.R. The role of parietal cortex in verbal working memory. J. Neurosci. 1998, 18, 5026–5034. [Google Scholar]
- Lee, J.; Park, S. Working memory impairments in schizophrenia: A meta-analysis. J. Abnorm. Psychol. 2005, 114, 599–611. [Google Scholar] [CrossRef]
- Manoach, D.S. Prefrontal cortex dysfunction during working memory performance in schizophrenia: Reconciling discrepant findings. Schizophr. Res. 2003, 60, 285–298. [Google Scholar] [CrossRef]
- Frisk, V.; Milner, B. The role of the left hippocampal region in the acquisition and retention of story content. Neuropsychologia 1990, 28, 349–359. [Google Scholar] [CrossRef]
- Petrides, M.; Alivisatos, B.; Evans, A.C.; Meyer, E. Dissociation of human mid-dorsolateral from posterior dorsolateral frontal cortex in memory processing. Proc. Natl. Acad. Sci. USA 1993, 90, 873–877. [Google Scholar] [CrossRef]
- Postle, B.R. Working memory as an emergent property of the mind and brain. Neuroscience 2006, 139, 23–38. [Google Scholar] [CrossRef]
- Postle, B.R.; Ferrarelli, F.; Hamidi, M.; Feredoes, E.; Massimini, M.; Peterson, M.; Alexander, A.; Tononi, G. Repetitive transcranial magnetic stimulation dissociates working memory manipulation from retention functions in the prefrontal, but not posterior parietal, cortex. J. Cogn. Neurosci. 2006, 18, 1712–1722. [Google Scholar] [CrossRef]
- Hamidi, M.; Tononi, G.; Postle, B.R. Evaluating frontal and parietal contributions to spatial working memory with repetitive transcranial magnetic stimulation. Brain Res. 2008, 1230, 202–210. [Google Scholar] [CrossRef]
- Buchsbaum, B.R.; D'Esposito, M. The search for the phonological store: From loop to convolution. J. Cogn. Neurosci. 2008, 20, 762–778. [Google Scholar] [CrossRef]
- Owen, A.M.; Morris, R.G.; Sahakian, B.J.; Polkey, C.E.; Robbins, T.W. Double dissociations of memory and executive functions in working memory tasks following frontal lobe excisions, temporal lobe excisions or amygdalo-hippocampectomy in man. Brain 1996, 119, 1597–1615. [Google Scholar] [CrossRef]
- D'Esposito, M.; Postle, B.R. The dependence of span and delayed-response performance on prefrontal cortex. Neuropsychologia 1999, 37, 1303–1315. [Google Scholar] [CrossRef]
- Manoach, D.S.; Greve, D.N.; Lindgren, K.A.; Dale, A.M. Identifying regional activity associated with temporally separated components of working memory using event-related functional MRI. Neuroimage 2003, 20, 1670–1684. [Google Scholar] [CrossRef]
- Curtis, C.E.; D'Esposito, M. Persistent activity in the prefrontal cortex during working memory. Trends Cogn. Sci. 2003, 7, 415–423. [Google Scholar] [CrossRef]
- Wible, C.G.; Preus, A.P.; Hashimoto, R. A cognitive neuroscience view of schizophrenic symptoms: Abnormal activation of a system for social perception and communication. Brain Imaging Behav. 2009, 3, 85–110. [Google Scholar] [CrossRef]
- Wible, C.G. Schizophrenia as a Disorder of Social Communication. Schizophr. Res. Treat. 2012, 2012. Article ID 920485. [Google Scholar]
- Wible, C.G. The Brain Bases of Phantom Auditory Phenomena: From Tinnitus to Hearing Voices. Semin. Hear. 2012, 33, 295–304. [Google Scholar] [CrossRef]
- Levine, D.N.; Finklestein, S. Delayed psychosis after right temporoparietal stroke or trauma: Relation to epilepsy. Neurology 1982, 32, 267–273. [Google Scholar] [CrossRef]
- Suzuki, K.; Takei, N.; Toyoda, T.; Iwata, Y.; Hoshino, R.; Minabe, Y.; Mori, N. Auditory hallucinations and cognitive impairment in a patient with a lesion restricted to the hippocampus. Schizophr. Res. 2003, 64, 87–89. [Google Scholar] [CrossRef]
- Stefanacci, L.; Buffalo, E.A.; Schmolck, H.; Squire, L.R. Profound amnesia after damage to the medial temporal lobe: A neuroanatomical and neuropsychological profile of patient E.P. J. Neurosci. 2000, 20, 7024–7036. [Google Scholar]
- Anderson, S.W.; Damasio, H.; Jones, R.D.; Tranel, D. Wisconsin Card Sorting Test performance as a measure of frontal lobe damage. J. Clin. Exp. Neuropsychol. 1991, 13, 909–922. [Google Scholar] [CrossRef]
- Teuber, H.L.; Battersby, W.S.; Bender, M.B. Performance of complex visual tasks after cerebral lesions. J. Nerv. Ment. Dis. 1951, 114, 413–429. [Google Scholar]
- Ishii, R.; Canuet, L.; Iwase, M.; Kurimoto, R.; Ikezawa, K.; Robinson, S.E.; Ukai, S.; Shinosaki, K.; Hirata, M.; Yoshimine, T.; et al. Right parietal activation during delusional state in episodic interictal psychosis of epilepsy: A report of two cases. Epilepsy Behav. 2006, 9, 367–372. [Google Scholar] [CrossRef]
- Redcay, E. The superior temporal sulcus performs a common function for social and speech perception: Implications for the emergence of autism. Neurosci. Biobehav. Rev. 2008, 32, 123–142. [Google Scholar] [CrossRef]
- Nummenmaa, L.; Passamonti, L.; Rowe, J.; Engell, A.D.; Calder, A.J. Connectivity analysis reveals a cortical network for eye gaze perception. Cereb. Cortex. 2010, 20, 1780–1787. [Google Scholar] [CrossRef]
- Redcay, E.; Dodell-Feder, D.; Pearrow, M.J.; Mavros, P.L.; Kleiner, M.; Gabrieli, J.D.; Saxe, R. Live face-to-face interaction during fMRI: A new tool for social cognitive neuroscience. NeuroImage 2010, 50, 1639–1647. [Google Scholar] [CrossRef] [Green Version]
- Saxe, R.; Wexler, A. Making sense of another mind: The role of the right temporo-parietal junction. Neuropsychologia 2005, 43, 1391–1399. [Google Scholar] [CrossRef]
- Saxe, R.; Xiao, D.K.; Kovacs, G.; Perrett, D.I.; Kanwisher, N. A region of right posterior superior temporal sulcus responds to observed intentional actions. Neuropsychologia 2004, 42, 1435–1446. [Google Scholar] [CrossRef]
- Pelphrey, K.A.; Morris, J.P.; Michelich, C.R.; Allison, T.; McCarthy, G. Functional anatomy of biological motion perception in posterior temporal cortex: An FMRI study of eye, mouth and hand movements. Cereb. Cortex. 2005, 15, 1866–1876. [Google Scholar] [CrossRef]
- Perrett, D.I.; Smith, P.A.; Mistlin, A.J.; Chitty, A.J.; Head, A.S.; Potter, D.D.; Broennimann, R.; Milner, A.D.; Jeeves, M.A. Visual analysis of body movements by neurones in the temporal cortex of the macaque monkey: A preliminary report. Behav. Brain Res. 1985, 16, 153–170. [Google Scholar] [CrossRef]
- Blanke, O.; Arzy, S. The out-of-body experience: Disturbed self-processing at the temporo-parietal junction. Neuroscientist 2005, 11, 16–24. [Google Scholar] [CrossRef]
- Perrett, D.I.; Hietanen, J.K.; Oram, M.W.; Benson, P.J. Organization and functions of cells responsive to faces in the temporal cortex. Phil. Trans. Roy. Soc. Lond. B Biol. Sci. 1992, 335, 23–30. [Google Scholar] [CrossRef]
- Perrett, D.I.; Smith, P.A.; Potter, D.D.; Mistlin, A.J.; Head, A.S.; Milner, A.D.; Jeeves, M.A. Visual cells in the temporal cortex sensitive to face view and gaze direction. Proc. Roy. Soc. Lond. B Biol. Sci. 1985, 223, 293–317. [Google Scholar] [CrossRef]
- Perrett, D.I.; Xiao, D.; Barraclough, N.E.; Keysers, C.; Oram, M.W. Seeing the future: Natural image sequences produce “anticipatory” neuronal activity and bias perceptual report. Q. J. Exp. Psychol. (Colchester) 2009, 62, 2081–2104. [Google Scholar] [CrossRef]
- Arzy, S.; Seeck, M.; Ortigue, S.; Spinelli, L.; Blanke, O. Induction of an illusory shadow person. Nature 2006, 443, 287. [Google Scholar] [CrossRef]
- Brugger, P.; Blanke, O.; Regard, M.; Bradford, D.T.; Landis, T. Polyopic heautoscopy: Case report and review of the literature. Cortex 2006, 42, 666–674. [Google Scholar] [CrossRef]
- Hashimoto, R.I.; Lee, K.; Preus, A.; McCarley, R.W.; Wible, C.G. An fMRI Study of Functional Abnormalities in the verbal working memory system and the relationship to clinical symptoms in chronic schizophrenia. Cereb. Cortex. 2010, 20, 46–60. [Google Scholar] [CrossRef]
- Ayhan, Y.; Abazyan, B.; Nomura, J.; Kim, R.; Ladenheim, B.; Krasnova, I.N.; Sawa, A.; Margolis, R.L.; Cadet, J.L.; Mori, S.; et al. Differential effects of prenatal and postnatal expressions of mutant human DISC1 on neurobehavioral phenotypes in transgenic mice: Evidence for neurodevelopmental origin of major psychiatric disorders. Mol. Psychiatr. 2011, 16, 293–306. [Google Scholar] [CrossRef]
- Sachdev, P. Schizophrenia-like psychosis and epilepsy: The status of the association. Am. J. Psychiatry. 1998, 155, 325–336. [Google Scholar]
- Stewart, I. Environmental risk factors for temporal lobe epilepsy—is prenatal exposure to the marine algal neurotoxin domoic acid a potentially preventable cause? Med. Hypotheses 2010, 74, 466–481. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wible, C.G. Hippocampal Physiology, Structure and Function and the Neuroscience of Schizophrenia: A Unified Account of Declarative Memory Deficits, Working Memory Deficits and Schizophrenic Symptoms. Behav. Sci. 2013, 3, 298-315. https://doi.org/10.3390/bs3020298
Wible CG. Hippocampal Physiology, Structure and Function and the Neuroscience of Schizophrenia: A Unified Account of Declarative Memory Deficits, Working Memory Deficits and Schizophrenic Symptoms. Behavioral Sciences. 2013; 3(2):298-315. https://doi.org/10.3390/bs3020298
Chicago/Turabian StyleWible, Cynthia G. 2013. "Hippocampal Physiology, Structure and Function and the Neuroscience of Schizophrenia: A Unified Account of Declarative Memory Deficits, Working Memory Deficits and Schizophrenic Symptoms" Behavioral Sciences 3, no. 2: 298-315. https://doi.org/10.3390/bs3020298