
Extensive Intestinal Resection Triggers Behavioral Adaptation, Intestinal Remodeling and Microbiota Transition in Short Bowel Syndrome

 ,
,
Abstract
:
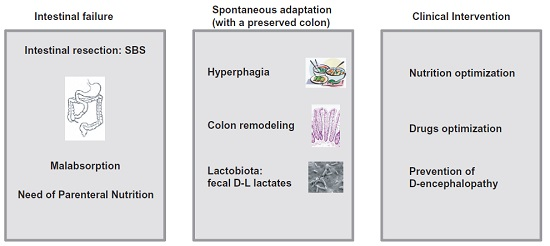{kind=link}
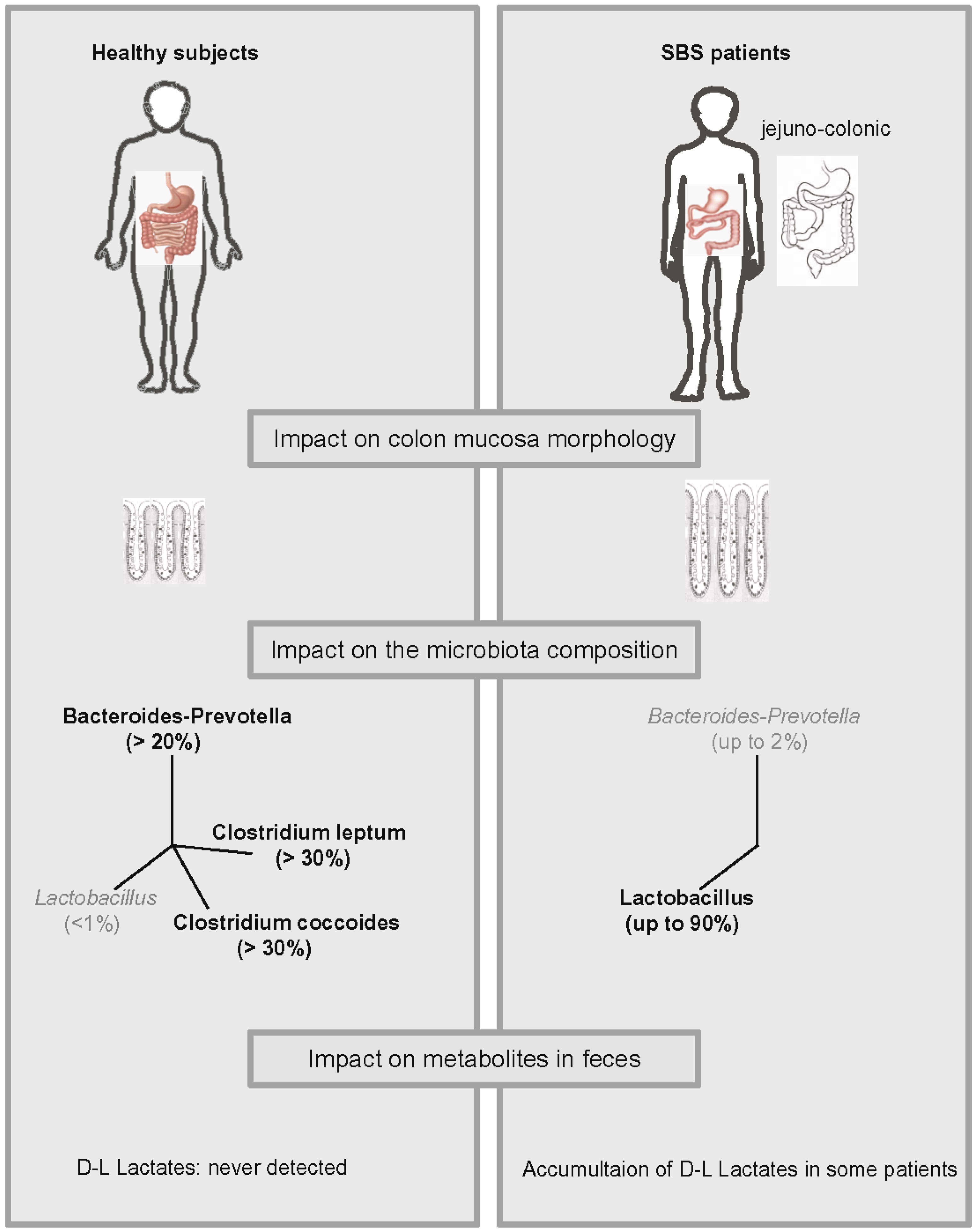{kind=link}
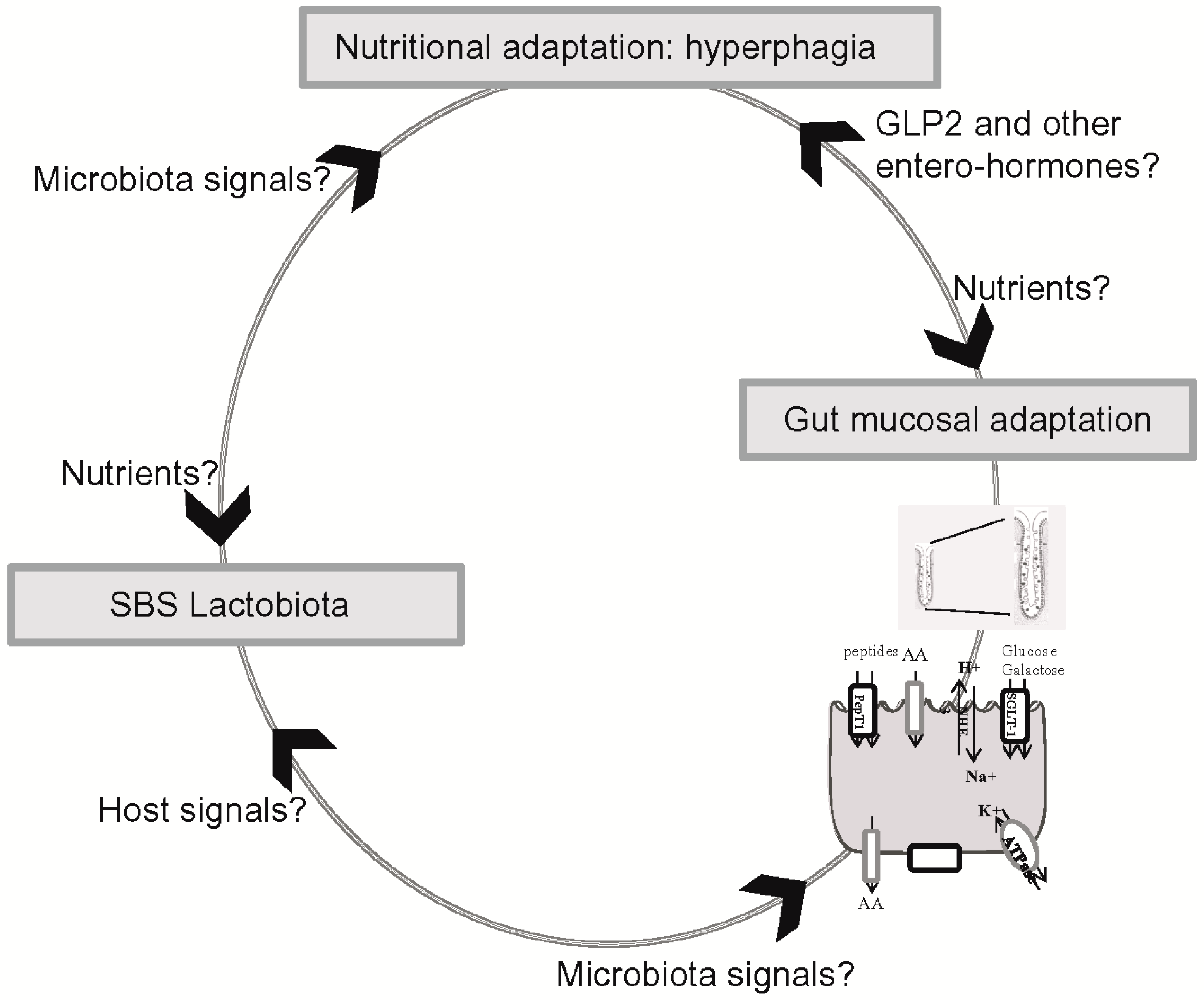{kind=link}
1. Introduction
2. Intestinal Failure and Short Bowel Syndrome
3. Endocrine Functions in Adult SBS
4. Hyperphagia in Adult SBS
5. Morphological Adaptation of Colon Mucosa in Adult SBS
6. Transporter Adaptation in Adult SBS
7. Colon Microbiota Composition and Its Metabolic Functions in Adult SBS
8. Fecal d-and l-Lactate and Clinical Risk for d-Encephalopathy in Adult SBS with Colon in Continuity
9. Conclusions
Conflicts of Interest
References
- Pironi, L.; Arends, J.; Baxter, J.; Bozzetti, F.; Pelaez, R.B.; Cuerda, C.; Forbes, A.; Gabe, S.; Gillanders, L.; Holst, M.; et al. ESPEN endorsed recommendations. Definition and classification of intestinal failure in adults. Clin. Nutr. 2015, 34, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Joly, F.; Dray, X.; Corcos, O.; Barbot, L.; Kapel, N.; Messing, B. Tube feeding improves intestinal absorption in short bowel syndrome patients. Gastroenterology 2009, 136, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Messing, B.; Blethen, S.; Dibaise, J.K.; Matarese, L.E.; Steiger, E. Treatment of adult short bowel syndrome with recombinant human growth hormone: A review of clinical studies. J. Clin. Gastroenterol. 2006, 40 (Suppl. 2), S75–S84. [Google Scholar] [CrossRef] [PubMed]
- Messing, B.; Lemann, M.; Landais, P.; Gouttebel, M.C.; Gerard-Boncompain, M.; Saudin, F.; Vangossum, A.; Beau, P.; Guedon, C.; Barnoud, D.; et al. Prognosis of patients with nonmalignant chronic intestinal failure receiving long-term home parenteral nutrition. Gastroenterology 1995, 108, 1005–1010. [Google Scholar] [CrossRef]
- Cherbuy, C.; Honvo-Houeto, E.; Bruneau, A.; Bridonneau, C.; Mayeur, C.; Duee, P.H.; Langella, P.; Thomas, M. Microbiota matures colonic epithelium through a coordinated induction of cell cycle-related proteins in gnotobiotic rat. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G348–G357. [Google Scholar] [CrossRef] [PubMed]
- Kaiko, G.E.; Stappenbeck, T.S. Host-microbe interactions shaping the gastrointestinal environment. Trends Immunol. 2014, 35, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Neish, A.S. Microbes in gastrointestinal health and disease. Gastroenterology 2009, 136, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Tomas, J.; Reygner, J.; Mayeur, C.; Ducroc, R.; Bouet, S.; Bridonneau, C.; Cavin, J.B.; Thomas, M.; Langella, P.; Cherbuy, C. Early colonizing Escherichia coli elicits remodeling of rat colonic epithelium shifting toward a new homeostatic state. ISME J. 2015, 9, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Tomas, J.; Wrzosek, L.; Bouznad, N.; Bouet, S.; Mayeur, C.; Noordine, M.L.; Honvo-Houeto, E.; Langella, P.; Thomas, M.; Cherbuy, C. Primocolonization is associated with colonic epithelial maturation during conventionalization. FASEB J. 2013, 27, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Tap, J.; Mondot, S.; Levenez, F.; Pelletier, E.; Caron, C.; Furet, J.P.; Ugarte, E.; Munoz-Tamayo, R.; Paslier, D.L.; Nalin, R.; et al. Towards the human intestinal microbiota phylogenetic core. Environ. Microbiol. 2009, 11, 2574–2584. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Jumpertz, R.; Le, D.S.; Turnbaugh, P.J.; Trinidad, C.; Bogardus, C.; Gordon, J.I.; Krakoff, J. Energy-balance studies reveal associations between gut microbes, caloric load, and nutrient absorption in humans. Am. J. Clin. Nutr. 2011, 94, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Lepage, P.; Hasler, R.; Spehlmann, M.E.; Rehman, A.; Zvirbliene, A.; Begun, A.; Ott, S.; Kupcinskas, L.; Dore, J.; Raedler, A.; et al. Twin study indicates loss of interaction between microbiota and mucosa of patients with ulcerative colitis. Gastroenterology 2011, 141, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Corcelles, R.; Daigle, C.; Schauer, P. Management of endocrine disease: Metabolic effects of bariatric surgery. Eur. J. Endocrinol. 2016, 174, R19–R28. [Google Scholar] [CrossRef] [PubMed]
- Buchman, A.L.; Scolapio, J.; Fryer, J. AGA technical review on short bowel syndrome and intestinal transplantation. Gastroenterology 2003, 124, 1111–1134. [Google Scholar] [CrossRef]
- Messing, B.; Crenn, P.; Beau, P.; Boutron-Ruault, M.C.; Rambaud, J.C.; Matuchansky, C. Long-term survival and parenteral nutrition dependence in adult patients with the short bowel syndrome. Gastroenterology 1999, 117, 1043–1050. [Google Scholar] [CrossRef]
- Corcos, O.; Cazals-Hatem, D.; Durand, F.; Kapel, N.; Guinhut, M.; Stefanescu, C.; Treton, X.; Bondjemah, V.; Attar, A.; Marmuse, J.P.; et al. Intestinal failure after bariatric surgery. Lancet 2013, 382, 742. [Google Scholar] [CrossRef]
- Jeppesen, P.B. Teduglutide, a novel glucagon-like peptide 2 analog, in the treatment of patients with short bowel syndrome. Therap. Adv. Gastroenterol. 2012, 5, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, P.B. Spectrum of short bowel syndrome in adults: Intestinal insufficiency to intestinal failure. JPEN J. Parenter. Enteral. Nutr. 2014, 38 (Suppl. 1), S8–S13. [Google Scholar] [CrossRef] [PubMed]
- Goulet, O.; Joly, F. Intestinal microbiota in short bowel syndrome. Gastroenterol. Clin. Biol. 2010, 34 (Suppl. 1), S37–S43. [Google Scholar] [CrossRef]
- Tappenden, K.A. Intestinal adaptation following resection. JPEN J. Parenter. Enteral Nutr. 2014, 38 (Suppl. 1), S23–S31. [Google Scholar] [CrossRef] [PubMed]
- Amiot, A.; Messing, B.; Corcos, O.; Panis, Y.; Joly, F. Determinants of home parenteral nutrition dependence and survival of 268 patients with non-malignant short bowel syndrome. Clin. Nutr. 2013, 32, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Crenn, P.; Coudray-Lucas, C.; Thuillier, F.; Cynober, L.; Messing, B. Postabsorptive plasma citrulline concentration is a marker of absorptive enterocyte mass and intestinal failure in humans. Gastroenterology 2000, 119, 1496–1505. [Google Scholar] [CrossRef] [PubMed]
- Remington, M.; Malagelada, J.R.; Zinsmeister, A.; Fleming, C.R. Abnormalities in gastrointestinal motor activity in patients with short bowels: Effect of a synthetic opiate. Gastroenterology 1983, 85, 629–636. [Google Scholar] [PubMed]
- Rodrigues, C.A.; Lennard-Jones, J.E.; Thompson, D.G.; Farthing, M.J. Energy absorption as a measure of intestinal failure in the short bowel syndrome. Gut 1989, 30, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.P.; Sarna, S.K.; Zhu, Y.R.; Buchmann, E.; Bonham, L.; Telford, G.L.; Roza, A.M.; Adams, M.B. Delayed gastroduodenal emptying is an important mechanism for control of intestinal transit in short-gut syndrome. Am. J. Surg. 1996, 171, 90–95; discussion 95–96. [Google Scholar] [CrossRef]
- Jeppesen, P.B.; Hartmann, B.; Thulesen, J.; Hansen, B.S.; Holst, J.J.; Poulsen, S.S.; Mortensen, P.B. Elevated plasma glucagon-like peptide 1 and 2 concentrations in ileum resected short bowel patients with a preserved colon. Gut 2000, 47, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Shin, C.E.; Falcone, R.A., Jr.; Kemp, C.J.; Erwin, C.R.; Litvak, D.A.; Evers, B.M.; Warner, B.W. Intestinal adaptation and enterocyte apoptosis following small bowel resection is p53 independent. Am. J. Physiol. 1999, 277 Pt 1, G717–G724. [Google Scholar] [PubMed]
- Martin, G.R.; Wallace, L.E.; Hartmann, B.; Holst, J.J.; Demchyshyn, L.; Toney, K.; Sigalet, D.L. Nutrient-stimulated GLP-2 release and crypt cell proliferation in experimental short bowel syndrome. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G431–G438. [Google Scholar] [CrossRef] [PubMed]
- Sigalet, D.L.; Martin, G.R.; Butzner, J.D.; Buret, A.; Meddings, J.B. A pilot study of the use of epidermal growth factor in pediatric short bowel syndrome. J. Pediatr. Surg. 2005, 40, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Nightingale, J.M.; Kamm, M.A.; van der Sijp, J.R.; Ghatei, M.A.; Bloom, S.R.; Lennard-Jones, J.E. Gastrointestinal hormones in short bowel syndrome. Peptide YY may be the “colonic brake” to gastric emptying. Gut 1996, 39, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, D.; Basseri, B.; Low, K.; Lezcano, S.; Soffer, E.E.; Conklin, J.L.; Mathur, R.; Pimentel, M. Efficacy of the glucagon-like peptide-1 agonist exenatide in the treatment of short bowel syndrome. Neurogastroenterol. Motil. 2011, 23, 739–e328. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J.; Erlich, P.; Asa, S.L.; Brubaker, P.L. Induction of intestinal epithelial proliferation by glucagon-like peptide 2. Proc. Natl. Acad. Sci. USA 1996, 93, 7911–7916. [Google Scholar] [CrossRef] [PubMed]
- Madsen, K.B.; Askov-Hansen, C.; Naimi, R.M.; Brandt, C.F.; Hartmann, B.; Holst, J.J.; Mortensen, P.B.; Jeppesen, P.B. Acute effects of continuous infusions of glucagon-like peptide (GLP)-1, GLP-2 and the combination (GLP-1 + GLP-2) on intestinal absorption in short bowel syndrome (SBS) patients. A placebo-controlled study. Regul. Pept. 2013, 184, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, P.B.; Hartmann, B.; Thulesen, J.; Graff, J.; Lohmann, J.; Hansen, B.S.; Tofteng, F.; Poulsen, S.S.; Madsen, J.L.; Holst, J.J.; et al. Glucagon-like peptide 2 improves nutrient absorption and nutritional status in short-bowel patients with no colon. Gastroenterology 2001, 120, 806–815. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, S.J. Nutritional Issues in the Short Bowel Syndrome—Total Parenteral Nutrition, Enteral Nutrition and the Role of Transplantation. Nestle Nutr. Inst. Workshop Ser. 2015, 82, 75–90. [Google Scholar] [PubMed]
- Jeppesen, P.B.; Sanguinetti, E.L.; Buchman, A.; Howard, L.; Scolapio, J.S.; Ziegler, T.R.; Gregory, J.; Tappenden, K.A.; Holst, J.; Mortensen, P.B. Teduglutide (ALX-0600), a dipeptidyl peptidase IV resistant glucagon-like peptide 2 analogue, improves intestinal function in short bowel syndrome patients. Gut 2005, 54, 1224–1231. [Google Scholar] [CrossRef] [PubMed]
- Seidner, D.L.; Joly, F.; Youssef, N.N. Effect of Teduglutide, a Glucagon-Like Peptide 2 Analog, on Citrulline Levels in Patients With Short Bowel Syndrome in Two Phase III Randomized Trials. Clin. Transl. Gastroenterol. 2015, 6, e93. [Google Scholar] [CrossRef] [PubMed]
- Mayeur, C.; Gratadoux, J.J.; Bridonneau, C.; Chegdani, F.; Larroque, B.; Kapel, N.; Corcos, O.; Thomas, M.; Joly, F. Faecal d/l lactate ratio is a metabolic signature of microbiota imbalance in patients with short bowel syndrome. PLoS ONE 2013, 8, e54335. [Google Scholar] [CrossRef] [PubMed]
- Joly, F.; Mayeur, C.; Messing, B.; Lavergne-Slove, A.; Cazals-Hatem, D.; Noordine, M.L.; Cherbuy, C.; Duee, P.H.; Thomas, M. Morphological adaptation with preserved proliferation/transporter content in the colon of patients with short bowel syndrome. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G116–G123. [Google Scholar] [CrossRef] [PubMed]
- Messing, B.; Pigot, F.; Rongier, M.; Morin, M.C.; Ndeindoum, U.; Rambaud, J.C. Intestinal absorption of free oral hyperalimentation in the very short bowel syndrome. Gastroenterology 1991, 100, 1502–1508. [Google Scholar] [PubMed]
- Compher, C.W.; Kinosian, B.P.; Metz, D.C. Ghrelin does not predict adaptive hyperphagia in patients with short bowel syndrome. JPEN J. Parenter. Enteral Nutr. 2009, 33, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Crenn, P.; Morin, M.C.; Joly, F.; Penven, S.; Thuillier, F.; Messing, B. Net digestive absorption and adaptive hyperphagia in adult short bowel patients. Gut 2004, 53, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Hanson, W.R.; Osborne, J.W.; Sharp, J.G. Compensation by the residual intestine after intestinal resection in the rat. II. Influence of postoperative time interval. Gastroenterology 1977, 72 Pt 1, 701–705. [Google Scholar] [PubMed]
- Feldman, E.J.; Dowling, R.H.; McNaughton, J.; Peters, T.J. Effects of oral versus intravenous nutrition on intestinal adaptation after small bowel resection in the dog. Gastroenterology 1976, 70 Pt 1, 712–719. [Google Scholar] [PubMed]
- Buchman, A.L.; Moukarzel, A.A.; Bhuta, S.; Belle, M.; Ament, M.E.; Eckhert, C.D.; Hollander, D.; Gornbein, J.; Kopple, J.D.; Vijayaroghavan, S.R. Parenteral nutrition is associated with intestinal morphologic and functional changes in humans. JPEN J. Parenter. Enteral Nutr. 1995, 19, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Dowling, R.H.; Booth, C.C. Functional compensation after small-bowel resection in man. Demonstration by direct measurement. Lancet 1966, 2, 146–147. [Google Scholar] [PubMed]
- Gouttebel, M.C.; Saint Aubert, B.; Colette, C.; Astre, C.; Monnier, L.H.; Joyeux, H. Intestinal adaptation in patients with short bowel syndrome. Measurement by calcium absorption. Dig. Dis. Sci. 1989, 34, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Lardy, H.; Thomas, M.; Noordine, M.L.; Bruneau, A.; Cherbuy, C.; Vaugelade, P.; Philippe, C.; Colomb, V.; Duee, P.H. Changes induced in colonocytes by extensive intestinal resection in rats. Dig. Dis. Sci. 2006, 51, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, T.R.; Fernandez-Estivariz, C.; Gu, L.H.; Bazargan, N.; Umeakunne, K.; Wallace, T.M.; Diaz, E.E.; Rosado, K.E.; Pascal, R.R.; Galloway, J.R.; et al. Distribution of the H+/peptide transporter PepT1 in human intestine: Up-regulated expression in the colonic mucosa of patients with short-bowel syndrome. Am. J. Clin. Nutr. 2002, 75, 922–930. [Google Scholar] [PubMed]
- Musch, M.W.; Bookstein, C.; Rocha, F.; Lucioni, A.; Ren, H.; Daniel, J.; Xie, Y.; McSwine, R.L.; Rao, M.C.; Alverdy, J.; et al. Region-specific adaptation of apical Na/H exchangers after extensive proximal small bowel resection. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, G975–G985. [Google Scholar] [CrossRef] [PubMed]
- Hines, O.J.; Bilchik, A.J.; Zinner, M.J.; Skotzko, M.J.; Moser, A.J.; McFadden, D.W.; Ashley, S.W. Adaptation of the Na+/glucose cotransporter following intestinal resection. J. Surg. Res. 1994, 57, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, C.W.; Qandeel, H.G.; Zheng, Y.; Duenes, J.A.; Sarr, M.G. Mechanisms of ileal adaptation for glucose absorption after proximal-based small bowel resection. J. Gastrointest. Surg. 2008, 12, 1854–1864; discussion 1864–1865. [Google Scholar] [CrossRef] [PubMed]
- Tsujikawa, T.; Itoh, A.; Fukunaga, T.; Satoh, J.; Yasuoka, T.; Fujiyama, Y. Alteration of aquaporin mRNA expression after small bowel resection in the rat residual ileum and colon. J. Gastroenterol. Hepatol. 2003, 18, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Zoetendal, E.G.; Akkermans, A.D.; de Vos, W.M. Temperature gradient gel electrophoresis analysis of 16S rRNA from human fecal samples reveals stable and host-specific communities of active bacteria. Appl. Environ. Microbiol. 1998, 64, 3854–3859. [Google Scholar] [PubMed]
- Kaneko, T.; Bando, Y.; Kurihara, H.; Satomi, K.; Nonoyama, K.; Matsuura, N. Fecal microflora in a patient with short-bowel syndrome and identification of dominant lactobacilli. J. Clin. Microbiol. 1997, 35, 3181–3185. [Google Scholar] [PubMed]
- Joly, F.; Mayeur, C.; Bruneau, A.; Noordine, M.L.; Meylheuc, T.; Langella, P.; Messing, B.; Duee, P.H.; Cherbuy, C.; Thomas, M. Drastic changes in fecal and mucosa-associated microbiota in adult patients with short bowel syndrome. Biochimie 2010, 92, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Lapthorne, S.; Pereira-Fantini, P.M.; Fouhy, F.; Wilson, G.; Thomas, S.L.; Dellios, N.L.; Scurr, M.; O’Sullivan, O.; Ross, R.P.; Stanton, C.; et al. Gut microbial diversity is reduced and is associated with colonic inflammation in a piglet model of short bowel syndrome. Gut Microbes 2013, 4, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Macfarlane, S. Fermentation in the human large intestine: Its physiologic consequences and the potential contribution of prebiotics. J. Clin. Gastroenterol. 2011, 45, S120–S127. [Google Scholar] [CrossRef] [PubMed]
- McNeil, N.I. The contribution of the large intestine to energy supplies in man. Am. J. Clin. Nutr. 1984, 39, 338–342. [Google Scholar] [PubMed]
- Briet, F.; Flourie, B.; Achour, L.; Maurel, M.; Rambaud, J.C.; Messing, B. Bacterial adaptation in patients with short bowel and colon in continuity. Gastroenterology 1995, 109, 1446–1453. [Google Scholar] [CrossRef]
- Nordgaard, I.; Hansen, B.S.; Mortensen, P.B. Importance of colonic support for energy absorption as small-bowel failure proceeds. Am. J. Clin. Nutr. 1996, 64, 222–231. [Google Scholar] [PubMed]
- Nordgaard, I.; Hansen, B.S.; Mortensen, P.B. Colon as a digestive organ in patients with short bowel. Lancet 1994, 343, 373–376. [Google Scholar] [CrossRef]
- Royall, D.; Wolever, T.M.; Jeejeebhoy, K.N. Evidence for colonic conservation of malabsorbed carbohydrate in short bowel syndrome. Am. J. Gastroenterol. 1992, 87, 751–756. [Google Scholar] [PubMed]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.P.; Lee, S.; Kang, S.K. d-lactic acidosis in humans: Review of update. Electrolyte Blood Press. 2006, 4, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.S.; Phelps, K.R.; Traube, M.; Barbosa-Saldivar, J.L.; Boxhill, C.; Carroll, H.J. d-lactic acidosis in a man with the short-bowel syndrome. N. Engl. J. Med. 1979, 301, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Halperin, M.L.; Kamel, K.S. d-lactic acidosis: Turning sugar into acids in the gastrointestinal tract. Kidney Int. 1996, 49, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kowlgi, N.G.; Chhabra, L. d-lactic acidosis: An underrecognized complication of short bowel syndrome. Gastroenterol. Res. Pract. 2015, 2015, 476215. [Google Scholar] [CrossRef] [PubMed]
- Davidovics, Z.H.; Vance, K.; Etienne, N.; Hyams, J.S. Fecal Transplantation Successfully Treats Recurrent d-Lactic Acidosis in a Child With Short Bowel Syndrome. JPEN J. Parenter. Enteral Nutr. 2015. [Google Scholar] [PubMed]
- Belenguer, A.; Holtrop, G.; Duncan, S.H.; Anderson, S.E.; Calder, A.G.; Flint, H.J.; Lobley, G.E. Rates of production and utilization of lactate by microbial communities from the human colon. FEMS Microbiol. Ecol. 2011, 77, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Tamayo, R.; Laroche, B.; Walter, E.; Dore, J.; Duncan, S.H.; Flint, H.J.; Leclerc, M. Kinetic modelling of lactate utilization and butyrate production by key human colonic bacterial species. FEMS Microbiol. Ecol. 2011, 76, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Bourriaud, C.; Robins, R.J.; Martin, L.; Kozlowski, F.; Tenailleau, E.; Cherbut, C.; Michel, C. Lactate is mainly fermented to butyrate by human intestinal microfloras but inter-individual variation is evident. J. Appl. Microbiol. 2005, 99, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Abeysekara, S.; Naylor, J.M.; Wassef, A.W.; Isak, U.; Zello, G.A. d-Lactic acid-induced neurotoxicity in a calf model. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E558–E565. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayeur, C.; Gillard, L.; Le Beyec, J.; Bado, A.; Joly, F.; Thomas, M. Extensive Intestinal Resection Triggers Behavioral Adaptation, Intestinal Remodeling and Microbiota Transition in Short Bowel Syndrome. Microorganisms 2016, 4, 16. https://doi.org/10.3390/microorganisms4010016
Mayeur C, Gillard L, Le Beyec J, Bado A, Joly F, Thomas M. Extensive Intestinal Resection Triggers Behavioral Adaptation, Intestinal Remodeling and Microbiota Transition in Short Bowel Syndrome. Microorganisms. 2016; 4(1):16. https://doi.org/10.3390/microorganisms4010016
Chicago/Turabian StyleMayeur, Camille, Laura Gillard, Johanne Le Beyec, André Bado, Francisca Joly, and Muriel Thomas. 2016. "Extensive Intestinal Resection Triggers Behavioral Adaptation, Intestinal Remodeling and Microbiota Transition in Short Bowel Syndrome" Microorganisms 4, no. 1: 16. https://doi.org/10.3390/microorganisms4010016