Defoliation of Soybean Expressing Cry1Ac by Lepidopteran Pests
by
 ,
,
María G. Murúa
1,* ,
,
Martín A. Vera
2,
María I. Herrero
1,
Sofia V. Fogliata
1 and
Andrew Michel
3 1
Instituto de Tecnología Agroindustrial del Noroeste Argentino (ITANOA), Estación Experimental Agroindustrial Obispo Colombres (EEAOC), Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Av. William Cross 3150, Las Talitas T4104AUD, Tucumán, Argentina
2
EEAOC, Las Talitas 4001, Tucumán, Argentina
3
Department of Entomology, Ohio Agricultural Research and Development Center, The Ohio State University, Wooster OH 44691, USA
*
Author to whom correspondence should be addressed.
Insects 2018, 9(3), 93; https://doi.org/10.3390/insects9030093
Submission received: 23 May 2018
/
Revised: 18 June 2018
/
Accepted: 27 June 2018
/
Published: 1 August 2018
Abstract
:Lepidoptera, stink bugs, and weevils are important pests in soybean. For lepidopteran control, insecticides and seed treatments are used. As an alternative, Bt soybean was developed to control primary pests of Lepidoptera such as Rachiplusia nu (Guenée) (Noctuidae), Chrysodeixis includens (Walker) (Noctuidae), Anticarsia gemmatalis Hübner (Erebidae), Helicoverpa gelotopoeon (Dyar) (Noctuidae), and Crocidosema aporema (Walsingham) (Tortricidae). However, the use of transgenic plants, and the resulting reduction of insecticide against target pests, may allow other pest species to become more prevalent in agricultural environments. Soybean expressing Cry1Ac against different lepidopteran nontarget and target insect pests was evaluated, and its performance was compared with non-Bt soybean with seed treatment. The treatments were Bt soybean, non-Bt soybean with seed treatment (Fortenza® diamide insecticide, Syngenta, Buenos Aires, Argentina), and non-Bt soybean without seed treatment. Larvae of H. gelotopoeon, Spodoptera albula (Walker) (Noctuidae), Spodoptera cosmiodes (Walker) (Noctuidae), Spodoptera eridania (Stoll) (Noctuidae), and Spodoptera frugiperda (J. E. Smith) (Lep.: Noctuidae) were used. The plants of each treatment were infested with larvae of each species, and the percentage of leaf damage produced by each species was recorded. The results showed that Bt soybean provided control of H. gelotopoeon and had a suppressive effect on S. frugiperda and S. albula. However, S. eridania and S. cosmiodes were not susceptible to the Cry1Ac protein in MON 87701 × MON 89788 soybean when evaluated by greenhouse infestation. Considering the performance of each species using non-Bt soybean without seed treatment, S. eridania would represent a potential risk in soybean crops.
1. Introduction
Soybean (Glycine max (L.) Merrill) is an oilseed plant of the family Fabaceae that is widely planted in various countries worldwide [1]. Argentina is the third-major soybean producer in the world, covering an area of 19.2 million hectares [2,3]. Lepidoptera, stink bugs (Hem.: Pentatomidae), and weevils (Col.: Curculionidae) are important pests that inflict the major yield loss of this crop [1,4,5,6,7,8]. For Lepidoptera in soybean, insecticides and seed treatments are used, but Bacillus thuringiensis (Berliner 1915) soybean (Bt soybean) is a new alternative method to control several species [5,6,7,9,10]. Bt soybean is a stacked varietal line developed by Monsanto that combines the transformation events MON 87701 (expressing Cry1Ac protein) and MON 89788 (glyphosate tolerance) [11]. At present, Bt soybean provides control for the primary Lepidopteran pests of soybean such as Rachiplusia nu (Guenée) (Lep.: Noctuidae), Chrysodeixis includens (Walker) (Lep.: Noctuidae), Anticarsia gemmatalis Hübner (Lep.: Erebidae), Helicoverpa gelotopoeon (Dyar) (Lep.: Noctuidae), Crocidosema aporema (Walsingham) (Lep.: Tortricidae), Colias lesbia (F.) (Lep.: Pieridae), Spilosoma virginica (F.) (Lep.: Arctiidae), Chloridea virescens (F.) (Lep.: Noctuidae), and Achyra bifidalis (F.) (Lep.: Crambidae). This technology can also suppress populations of Spodoptera frugiperda (J. E. Smith) (Lep.: Noctuidae), Elasmopalpus lignosellus (Zeller) (Lep.: Noctuidae), and Helicoverpa zea (Boddie) (Lep.: Noctuidae) [11]. However, the use of Bt plants and the resulting reduction of insecticide use against target pests may allow other pest species to become more relevant in agricultural environments [9]. Field trials conducted in northern China showed that mirid bugs (Het.: Miridae) have progressively increased population sizes and acquired pest status in cotton and multiple other crops, in association with a regional increase in Bt cotton adoption. Bt cotton has become a source of mirid bugs, and their population increases are related to drops in insecticide use in this crop [12]. On the other hand, the emergence of Striacosta albicosta (Smith) (Lep.: Noctuidae), as a potential pest of corn in South Dakota (USA) may be related to the widespread planting of Cry1Ab Bt corn hybrids. Continuous planting of Cry1Ab Bt corn hybrids over large areas favors this species by effectively eliminating competition from Ostrinia nubilalis (Hübner) (Lep.: Crambidae) [13]. Therefore, research on the interaction of nontarget pest species with this Bt technology is of great theoretical and practical importance.
In recent years, the occurrence of caterpillars from the Spodoptera genus (Lep. Noctuidae) has increased and caused damage to soybean crops in Brazil [14,15,16,17,18] and Argentina [5,6,7,19]. Within this complex, S. cosmiodes (Walker), S. eridania (Stoll), and S. frugiperda are prominent in causing damage [7,16,20]. It is important to highlight that the damage produced by S. frugiperda in soybean crops is common in farms with grasses and pasture weeds before soybean sowing. Some of these grasses and various pasture weeds [Conyza bonariensis (L.) Cronquist (Asteraceae), Brassica campestris (L.) Metzg. (Brassicaceae), and Sphaeralcea bonariensis (Cav.) Griseb (Malvaceae)] play an important role as reservoirs for insect pests, which move from one to another species of grasses or crops [21,22]. The presence of S. albula (Walker) in soybean crops is occasional and they were also detected in farms with grasses and pasture weeds.
The bioecological characteristics of the Spodoptera complex (polyphagia, great voracity in feeding, high reproductive capacity, migration behavior, host races) [23,24,25,26,27,28,29,30,31] added to some precedents of insecticide resistance and the natural tolerance of Spodoptera spp. to the Cry1A protein [32,33], making it necessary to evaluate their behavior with Bt soybean. Previous studies that evaluated the susceptibility and tolerance of different pests suggested that the most susceptible species to the Cry1Ac protein are A. gemmatalis, R. nu, and C. includens, and the most tolerant species are S. frugiperda and S. cosmioides [34,35]. While S. frugiperda has developed resistance to Cry1F proteins [36,37,38], S. eridania and S. albula are tolerant to several chemical insecticides [39,40,41,42,43,44] and to the B. thuringiensis Cry1Ac gene [45,46].
Given the agronomic challenges of soybean production combined with the emerging Spodoptera pest complex, evaluations of additional control strategies are needed. Although various aspects of Spodoptera spp. performance have been studied using different crops or artificial diet through field and laboratory studies (Table 1), many of the field studies used natural infestations where the specific identity of each species was not considered. This is due to the fact that the first larval instars of this genus, in general, present similar characters which make it impossible to identify [46,47,48,49]. This has led to much speculation about the damage of this complex in the field. The knowledge about the defoliation produced by each species will help to determine the real individual defoliation in soybean plants.
Helicoverpa gelotopoeon is another major pest of soybean. Larvae of this species cause damage in the vegetative and reproductive plant growth stages. In Tucumán and other provinces of Argentina, this species causes severe damage to soybean and chickpea (Cicer arietinum L. (Fabaceae)) crops and can be difficult to control with insecticides [50,51,52,53]. Some other species of Helicoverpa, such as H. armigera Hübner and H. zea, have also developed resistance to insecticides and Cry proteins [54,55,56,57,58,59].
Previous studies were conducted under field conditions considering the Helicoverpa genus in general [7,8,19]. This has led to much speculation, given the coexistence of H. gelotopoeon and H. armigera in soybean crops in northwestern Argentina [52]. In this genus, species can only be separated by morphological characters of the adults [50]. Thus, the study of H. gelotopoeon performance is important to know the real potential of defoliation in soybean crops.
The great potential for defoliation of soybean plants [16,20] and damage to flowers and pods [14] by Spodoptera spp. and H. gelotopoeon [51,52] requires the adoption of control tactics to prevent yield loss. Control is achieved with insecticides, often indirectly as result of sprays for A. gemmatalis, C. includens, and R. nu [5,6,7,8,60].
The rapid adoption of Bt soybean in Argentina [2] and other countries in South America [61] has increased the need to know the behavior of nontarget and target pests of Bt soybean. The objective of this study was to evaluate the performance of soybean expressing Cry1Ac against different lepidopteran nontarget and target insect pests and to compare its performance with that of a non-Bt soybean with seed treatment. We addressed this study by evaluating the defoliation produced by different lepidopteran pests recorded at eight days after infestation.
2. Materials and Methods
2.1. Larval Collection
Larvae of H. gelotopoeon, S. albula, S. cosmiodes, S. eridania, and S. frugiperda were collected from January to March 2014 in commercial soybean fields in La Cocha (Dpto La Cocha) and Overo Pozo (Dpto Cruz Alta) counties in Tucumán province (Argentina). A minimum of 250 larvae (from 3rd to 6th instars) of each species were collected using a vertical beat sheet [62]. Then, each larva was placed in a glass tube (12 cm H and 1.5 cm D) with leaves of soybean and transported to our laboratories at the Estación Experimental Agroindustrial Obispo Colombres (EEAOC). The collected larvae were placed in growth chambers under controlled conditions (27 ± 2 °C, 70 ± 5% RH, 14:10 h L:D) until adult emergence. Late larval instars and adults were examined using morphological characters both to confirm the identity of species [47,49,63,64] and to establish pure cultures for each species in the laboratory. Voucher specimens for each species were deposited in the collection of Sección Zoología Agrícola, (EEAOC) Tucumán, Argentina.
2.2. Insect Rearing
Each species colony was maintained in the same chamber under identically controlled conditions at 27 ± 2 °C, 70 ± 5% RH, 14:10 h L:D. Colonies were reared according to the methodology described by Murúa et al. [65] and Herrero et al. [53]. Twenty-five pairs per cage and per species (25 females and 25 males) (N = 6) were used. Adults were maintained in cylindrical oviposition cages made out of plastic mesh (20 cm high and 15 cm in diameter) lined with polyethylene bags as an oviposition substrate. For aeration, both ends of the cage were covered with a nylon cloth. The food was provided via a cotton plug saturated with a mixture of honey and water (1:1 v/v) which was replaced every day. Cages were checked daily for oviposition and adult mortality. Eggs were collected daily with a moistened brush and deposited in Petri dishes lined with moistened filter paper. Once emerged, neonate larvae were placed in 20-cm diameter, 800-mL containers with an artificial larval diet that included bean flour (Grandiet®, Buenos Aires, Argentina), wheat germ (Grandiet®, Buenos Aires, Argentina), brewer’s yeast (Calsa®, Tucumán, Argentina), vitamin C (Anedra®, Buenos Aires, Argentina), sorbic acid (Anedra®, Buenos Aires, Argentina), vitamin supplement amino acids (Ruminal®, Buenos Aires, Argentina), and methylparaben (Todo Droga®, Córdoba, Argentina). Diet was replaced every five days. As larvae pupated, pupae were sexed and placed in cup containers with moistened filter paper until adult’s emergence. Adults were used to initiate a new generation. After establishing a colony for each species, individuals from the 2nd generation (F2) were used for the evaluation of soybean Cry1Ac against different pest species in a greenhouse.
2.3. Greenhouse Studies
Soybean seeds of maturity group 7 were used (recommended for cultivation in northwestern Argentina). The treatments evaluated were Bt soybean Cry1Ac (T1), non-Bt soybean plus seed treatment (T2) (Fortenza® diamide insecticide, Syngenta, Argentina) according to the dose/rate recommended by the company (36 g a.i./100 Kg/seed), and non-Bt soybean without seed treatment as an untreated control (T3). The three treatments were planted in different pots (15 cm D, 600 mL) using sterilized soil. One seed per pot was planted and each plant was labeled to distinguish both the species and treatment. The plants were maintained under greenhouse conditions under ambient lighting at approximately 33 ± 4 °C, 80 ± 10% RH, 14:10 h L:D.
The experimental design was completely randomized with three replicates per treatment, where each replicate consisted of 20 soybean plants. A total of 60 plants for each pest species (5) per treatment (3) were evaluated, resulting in 900 plants.
Each plant for each treatment was inoculated at the V1 stage [66] (approximately 14 days after planting) with 10 larvae (L1) of a pest species. The evaluations of defoliation produced by each species were recorded at 8 days after inoculation (DAI), according the visual estimation of defoliation scale described by Kogan and Turnipseed [67].
Expression of the Cry1Ac protein in the soybean plants was confirmed using qualitative ELISA Quickstix lateral flow detection strips (Envirologix, Portland, ME, USA).
2.4. Data Analysis
To meet parametric assumptions, percentage data on defoliation damage were transformed to arcsine square root prior to analysis [68]; nevertheless, untransformed means (±SE) are shown in the figure. The percentage of defoliation damage produced by each species in the different treatments and the performance of these species using non-Bt soybean without seed treatment were analyzed using a one-way ANOVA, and means were separated using Tukey’s tests (p < 0.05) with InfoStat [69].
3. Results
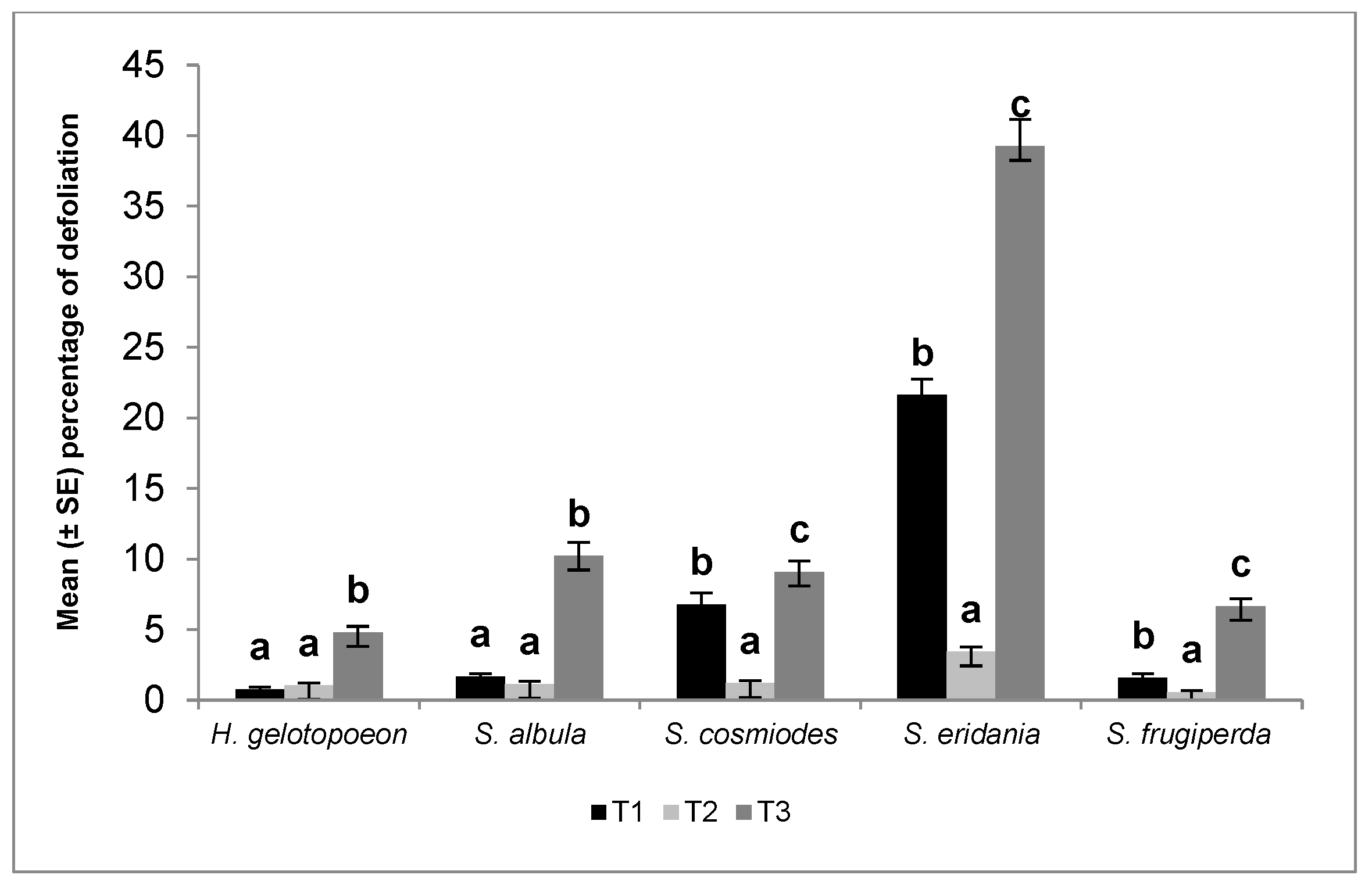
All results are shown in Figure 1. Significant differences were found in the average defoliation damage produced by H. gelotopoeon among different treatments (F = 64.3; df = 2,177; p < 0.0001). Bt soybean (T1) and non-Bt soybean plus Fortenza® (T2) presented similar values, but significant differences were found when they were compared with the control (T3). The average defoliation damage produced by S. albula was lower in the Bt soybean and non-Bt soybean plus Fortenza® treatments compared to the control (Figure 1) (F = 111.0; df = 2,177; p < 0.0001). For S. cosmiodes, lower feeding damage was recorded in the non-Bt soybean plus Fortenza® treatment, compared to the Bt soybean and control plants (F = 76.0; df = 2,176; p < 0.0001). The best control of S. eridania was with the non-Bt soybean plus Fortenza® treatment, registering a lower percentage of defoliation compared to Bt soybean and the control plants (F = 269.1; df = 2,177; p < 0.0001). Finally, significant differences were found in the percentage of defoliation produced by S. frugiperda in the different treatments (F = 94.7; df = 2,177; p < 0.0001). The lowest defoliation was obtained with non-Bt soybean plus Fortenza® (0.57 ± 0.12), whereas the control plants showed the highest defoliation.
A comparison of the potential risk of these species on untreated plants (non-Bt soybean without seed treatment) showed significant differences (F = 100.9; df = 4,293; p < 0.0001). The highest defoliation damage was produced by S. eridania larvae (39.3 ± 1.89), followed by S. albula (10.2 ± 0.97), S. cosmiodes (9.1 ± 0.79), S. frugiperda (6.7 ± 0.54), and H. gelotopoeon (4.8 ± 0.43).
4. Discussion
This study was conducted to characterize the efficacy of soybean expressing Cry1Ac against H. gelotopoeon, S. albula, S. cosmiodes, S. eridania, and S. frugiperda and compare its performance with that of a non-Bt soybean with seed treatment by assessing defoliation (Figure 1). On the other hand, the performance analysis made among the species using non-Bt soybean without seed treatment showed which of these species would represent a potential risk for soybean crops. For both analyses, the defoliation and potential risk were calculated considering the identity for each species in greenhouse conditions (Table 1).
Our results showed that Bt soybean provided variable control of some pests evaluated. This new technology had the best control of H. gelotopoeon and a suppressive effect on S. frugiperda and S. albula.
Non-Bt soybean with seed treatment provided good control of the five species. Nevertheless, this control will be only for the early plant growth stage, when plants have more insecticide active ingredient in their system. S. cosmiodes, S. eridania, S. frugiperda, and S. albula feeding resulted in the lowest percentage of defoliation compared with the other treatments.
In the case of S. albula, the defoliation recorded in T1 was similar to that recorded in T2 and the differences were not significant.
These results are consistent with field studies evaluating Bt soybean in Brazil and Argentina. A study of diversity, composition, and population dynamics of arthropods in non-Bt soybean and Bt soybean showed that A. gemmatalis, C. includens, C. virescens, and S. frugiperda were significantly controlled by Bt soybean. However, other species of the Spodoptera complex were not controlled by this technology [70]. Other studies reported that S. cosmiodes was the most abundant species attacking Bt soybean [71] and that development and reproduction of this species were not affected by the Cry1Ac protein [20]. A field study showed that the lepidopteran pests recorded during the soybean cycle were R. nu, A. gemmatalis, H. gelotopoeon, S. frugiperda, and S. cosmioides [72] and the population abundance of the last two species was similar in Bt and non-Bt soybean. In northwestern Argentina, a trial conducted to evaluate Bt soybean against different insect pest and natural predators found that this new technology provided good control of target pests such as A. gemmatalis, C. includens, and R. nu. The levels of leaf damage observed with Bt soybean were lowest and significantly different to those obtained with non-Bt soybean [4]. On the other hand, bioassays and greenhouse studies found that S. cosmiodes, S. eridania, and S. frugiperda exhibited low to no susceptibility to MON 87701 × MON 89788 soybean containing the protein Cry1Ac, and these species showed higher tolerance to the Cry1Ac protein than other Lepidoptera species, such as C. includens, C. virescens, and H. zea [60]. In the same study, a moderate larval incidence of S. eridania and S. frugiperda on MON 87701 x MON 89788 soybean and the respective near-isogenic negative check was found. No significant differences in larval incidence and defoliation by S. eridania were found with Bt soybean and the near-isogenic negative checks for both maturity groups. Similarly, larval incidence of S. frugiperda on Bt soybean did not differ significantly from the respective near-isogenic negative checks for both maturity groups. However, defoliation by S. frugiperda on Bt soybean was significantly lower than on the near-isogenic, being the defoliation registered, similar to those reported in this study in Bt soybean. The defoliation recorded for S. eridania was different to those reported in greenhouse studies in Brazil [60], but in both studies, defoliation was high in non-Bt soybean. These differences could be due to the number of larvae used to evaluate the defoliation per plants.
The non-Bt soybean with seed treatment showed the lowest defoliation of S. frugiperda, S. albula, S. cosmiodes, and S. eridania. These results are similar to another study [73], where diamides had good potential to control different lepidopteran pests such as S. eridania, S. cosmiodes, S. albula, A. gemmatalis, and S. frugiperda.
Considering the performance of each species using non-Bt soybean without seed treatment, H. gelotopoeon showed the lowest defoliation. This species is one of the most important pests in soybean crops, but its larvae prefer the reproductive plant growth stages [50,74], which explains the low defoliation recorded in this study. As mentioned, Spodoptera spp. are polyphagous, but show preferences to different host plants, such as the preference of S. frugiperda to corn (Zea mays L. (Poaceae)) over soybean and other crops [75]. Spodoptera cosmiodes and S. albula presented similar levels of defoliation in our test, although S. cosmiodes is the most frequent Spodoptera spp. affecting soybean crops during the vegetative and reproductive stages [5,6,14,16,20,50]. Another study suggested that S. cosmioides may be more adapted to chemical compounds of soybean and cotton (Gossypium hirsutum L. (Malvaceae)), given their faster development and higher survival rates on these hosts [76]. Larvae of S. albula feed on a wide variety of host plants and they also exhibit some preference for several weeds (Boerhavia erecta (L.) (Nyctaginaceae); Echinochloa colonum (L.) Link (Poaceae), from which they can migrate to cultivated plants [27,29]. The highest defoliation damage was produced by S. eridania. This species had been cited as infesting a large number of crops in various regions of the Americas [49,74,77]. Additionally, this species has been reported from outbreaks under different conditions [15,18,28]. This species develops on weeds, which generally constitute a primary source of cultivated plant infestations such as those of soybean [15,26,28]. Considering the defoliation recorded in this study, S. eridania would represent a potential risk to soybean crops. According to these results, this species may be more adapted to chemical compounds of soybean given their faster development and higher survival rates [76].
5. Conclusions
Our results suggest that H. gelotopoeon, S. frugiperda, and S. albula were susceptible to the Cry1Ac protein. However, S. eridania and S. cosmiodes were not susceptible to the Cry1Ac protein in MON 87701 × MON 89788 soybean when evaluated by greenhouse infestation, supporting the idea that the level of activity of this protein against these species is low. Consequently, other control tactics, such as seed treatment, must be used in combination with MON 87701 × MON 89788 soybean in the field for the efficient management of Spodoptera species.
Our results contribute to determining the defoliation and potential risk of these pests in soybean crops. Considering the increasing importance of S. cosmioides and S. eridania in the region, future studies should be focused toward understanding survival, population dynamics, and infestation of these species during all growth stages of soybean plants, and their biology, including host adaptation. On the other hand, determination of the combined action of Bt with the seed treatment in the defoliation in the field will be important, like other control tactics. Therefore, the use of Bt soybean as a tool for integrated pest management should be planned according to the major insect problems in each area.
Author Contributions
M.G.M. and M.A.V. conceived and designed the experiment; M.G.M. and M.A.V. performed the experiments; M.G.M. analyzed the data; M.I.H., S.V.F., A.M., and M.G.M. contributed reagents/materials/analysis tools; M.G.M. and A.M. wrote the paper.
Funding
This research was funded by [Agencia Nacional de Promoción Científica de Argentina through the Fondo Nacional de Ciencia y Tecnología, Ministerio de Ciencia, Tecnología e Innovación Productiva] grant number [PICTO 2016 EEAOC N°0112, Préstamo BID].
Acknowledgments
We thank David González and José Olmos at Estación Experimental Agroindustrial Obispo Colombres for their assistance in the collection of material and excellent technical support in the greenhouse and laboratory, Lic. Eduardo Willink (Estación Experimental Agroindustrial Obispo Colombres) for his constructive comments on an earlier draft of the manuscript, and Fernando Navarro (Consejo Nacional de Investigaciones Científicas y Técnicas, Instituto Superior de Entomología Abraham Willink, Facultad de Ciencias Naturales, Universidad Nacional de Tucumán) for the taxonomic identification of the Spodoptera complex and H. gelotopoeon, for his unconditional support, and for sharing memorable moments. This study was supported by the Agencia Nacional de Promoción Científica de Argentina through the Fondo Nacional de Ciencia y Tecnología, Ministerio de Ciencia, Tecnología e Innovación Productiva, Grant PICTO 2016 EEAOC N°0112, Préstamo BID, awarded to MGM and Consejo de Investigaciones de la Universidad Nacional de Tucumán, Grant CIUNT G638/1.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Silva, G.V.; Pasini, A.; Bueno, A.D.F.; Bortolotto, O.C.; Caselato Barbosa, G.; Stopa Cruz, Y.K. No impact of Bt soybean that express Cry1Ac protein on biological traits of Euschistus heros (Hemiptera, Pentatomidae) and its egg parasitoid Telenomus podisi (Hymenoptera, Platygastridae). Rev. Bras. Entomol. 2014, 58, 285–290. [Google Scholar] [CrossRef]
- Argenbio. Gráfico de Evolución de las Superficies Sembradas con OGM en la Argentina, en Porcentajes. Available online: http://www.argenbio.org/adc/uploads/imagenes_doc/planta_stransgenicas/1_Grafico_de_evolucion_de_superficie_cultivada_OGM_en_Arg_en_porcentaje.pdf (accessed on 22 May 2018).
- Bolsa de Cereales. Panorama Agrícola Semanal. Estimaciones Agrícolas. Relevamiento al 29/03/2018. Available online: http://www.bolsadecereales.com.ar (accessed on 22 May 2018).
- Cazado, L.E.; Murúa, M.G.; Casmuz, A.S.; Socías, M.G.; Vera, M.T.; O´Brien, C.W.; Gastaminza, G. Geographical distribution and new associations of Rhyssomatus subtilis (Coleoptera: Curculionidae) in Argentina. Fla. Entomol. 2013, 96, 663–669. [Google Scholar] [CrossRef]
- Casmuz, A.S.; Cazado, L.E.; Scalora, F.S.; Tuzza, M.F.; Fernández, R.A.; Fadda, C.; Fadda, L.A.; Dami, L.; Colledani Toranzo, A.; Gómez, C.H.; et al. Evaluación de diferentes alternativas para el control del complejo de plagas del cultivo de soja. In El Cultivo de la Soja en el Noroeste Argentino: Campaña 2013–2014; Devani, M.R., Ledesma, F., Sánchez, J.R., Eds.; Publicación Especial Soja EEAOC Nº 50; EEAOC: San Miguel de Tucumán, Argentina, 2014; pp. 137–142. Available online: http://www.eeaoc.org.ar/publicaciones/categoria/16/461/10Plagas.html (accessed on 22 May 2018).
- Casmuz, A.S.; Cazado, L.E.; Fadda, L.A.; Tuzza, M.F.; Fadda, C.; Jadur, A.; Merlini, A.; Orce, J.D.; Vera, A.; Marín, M.N.; et al. Momento de aplicación de los insecticidas y su relación con el control ejercido sobre el complejo de orugas defoliadoras de la soja. In El Cultivo de la Soja en el Noroeste Argentino: Campaña 2014–2015; Devani, M.R., Ledesma, F., Sánchez, J.R., Eds.; Publicación Especial Soja EEAOC Nº 51, EEAOC: San Miguel de Tucumán, Argentina, 2015; pp. 120–125. Available online: http://www.eeaoc.org.ar/publicaciones/categoria/16/539/62-Zoologia-621Defoliadoras.html (accessed on 22 May 2018).
- Casmuz, A.S.; Fadda, L.A.; Tuzza, M.F.; Jadur, A.; Fernández, C.; Díaz Arnijas, G.; Vera, M.A.; Cazado, L.E.; Orce, J.D.; Murúa, M.G.; et al. Manejo de plagas en soja Intacta y RR1. In El Cultivo de la Soja en el Noroeste Argentino: Campaña 2015–2016; Devani, M.R., Ledesma, F., Sánchez, J.R., Eds.; Publicación Especial Soja EEAOC Nº 52, EEAOC: San Miguel de Tucumán, Argentina, 2016; pp. 134–142. Available online: http://www.eeaoc.org.ar/publicaciones/categoria/16/645/Capitulo-5.html (accessed on 22 May 2018).
- Casmuz, A.; Fadda, L.; Vera, M.A.; Jadur, A.; Díaz Arnijas, G.; Ramos, L.; Tarulli, L.; Bayos, N.; Fernández Alvarez, C.; Murúa, M.G.; et al. Estrategias para la prevención de la resistencia de insectos en soja Bt: Manejo de plagas en el refugio. In El Cultivo de la Soja en el Noroeste Argentino: Campaña 2016–2017; Devani, M.R., Ledesma, F., Sánchez, J.R., Eds.; Publicación Especial Soja EEAOC Nº 53, EEAOC: San Miguel de Tucumán, Argentina, 2017; pp. 115–121. Available online: http://www.eeaoc.org.ar/publicaciones/categoria/16/738/Capitulo-5---2017.html (accessed on 22 May 2018).
- Wu, K.; Um, W.; Liang, G.; Gou, Y. Regional reversion of insecticide resistance in Helicoverpa armigera (Lepidoptera: Noctuidae) is associated with the use of Bt cotton in northern China. Pest Manag. Sci. 2005, 61, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Cazado, L.E.; Casmuz, A.S.; Riley, D.G.; Scalora, F.S.; Gastaminza, G.A.; Murúa, M.G. Rhyssomatus subtilis (Coleoptera: Curculionidae) impact in soybean plan stands. J. Entomol. Sci. 2016, 51, 69–78. [Google Scholar] [CrossRef]
- Monsanto. Available online: http://www.intactarr2pro.com.ar/ (accessed on 23 May 2018).
- Lu, Y.; Wu, K.; Jiang, Y.; Xia, B.; Li, P.; Feng, H.; Wyckhuys, K.A.G.; Guo, Y. Mirid bug outbreaks in multiple crops correlated with wide-scale adoption of Bt cotton in China. Science 2010, 328, 1151–1154. [Google Scholar] [CrossRef] [PubMed]
- Catangui, M.A.; Berg, R.K. Western bean cutworm, Striacosta albicosta (Smith) (Lepidoptera: Noctuidae), as a potential pest of transgenic Cry1Ab Bacillus thuringiensis corn hybrids in South Dakota. Environ. Entomol. 2006, 35, 1439–1452. [Google Scholar] [CrossRef]
- Hoffmann-Campo, C.B.; Moscardi, F.; Corrêa-Ferreira, B.; Oliveira, L.J.; Sosa-Gómez, D.R.; Panizzi, A.R.; Corso, I.C.; Gazzoni, D.L.; Oliveira, E.B. Pragas da Soja no Brasil e seu Manejo Integrado; Circular Técnica; Embrapa e Centro Nacional de Pesquisa de Soja: Londrina, Brazil, 2000; Volume 30, p. 70. ISSN 1516-7860. [Google Scholar]
- Santos, K.B.; Meneguim, A.M.; Neves, P.M.O.J. Biology and consumption of Spodoptera eridania (Cramer) (Lepidoptera: Noctuidae) in different hosts. Neotrop. Entomol. 2005, 4, 903–910. [Google Scholar] [CrossRef]
- Bueno, R.C.O.F.; Bueno, A.F.; Moscardi, F.; Parra, J.R.P.; Hoffmann-Campo, C.B. Lepidopteran larva consumption of soybean foliage: Basis for developing multiple-species economic thresholds for pest management decisions. Pest Manag. Sci. 2011, 67, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Sosa-Gómez, D.R.; Correa-Ferreira, B.S.; Hoffmann-Campo, C.B.; Corso, I.C.; Oliveira, L.J.; Moscardi, F.; Panizzi, A.R.; de Freitas Bueno, A.; Hirose, E.; Roggia, S. Manual de Identificação de Insetos e Outros Invertebrados da Cultura de Soja; Embrapa Soja: Londrina, Brazil, 2014; p. 100. ISSN 1516-781X. [Google Scholar]
- Favetti, B.M.; Butnariu, A.R.; Foerster, L.A. Biology and reproductive capacity of Spodoptera eridania (Cramer) (Lepidoptera, Noctuidae) in different soybean cultivars. Rev. Bras. Entomol. 2015, 59, 89–95. [Google Scholar] [CrossRef]
- Cazado, L.E.; Casmuz, A.S.; Scalora, F.S.; Fadda, C.; Fernández, R.A.; Tuzza, M.F.; Fadda, L.A.; Dami, L.; Colledani Toranzo, A.; Jadur, A.; et al. Comportamiento de la soja Bt frente a las principales plagas insectiles y depredadores en el Noroeste Argentino (NOA). In El Cultivo de la Soja en el Noroeste Argentino: Campaña 2013–2014; Devani, M.R., Ledesma, F., Sánchez, J.R., Eds.; Publicación Especial Soja EEAOC Nº 50, EEAOC: San Miguel de Tucumán, Argentina, 2014; pp. 153–161. Available online: http://www.eeaoc.org.ar/publicaciones/categoria/16/463/12Plagas-vs-Bt.html (accessed on 22 May 2018).
- Silva, V.G.; Bueno, F.A.; Bortolottoa, C.O.; Santos, C.A.; Fernandes, P.A. Biological characteristics of black armyworm Spodoptera cosmioides on genetically modified soybean and corn crops that express insecticide Cry proteins. Rev. Bras. Entomol. 2016, 60, 255–259. [Google Scholar] [CrossRef]
- Bentancourt, C.M.; Scatoni, I.B. Lepidópteros de importancia económica. In Reconocimiento, Biología y Daños de las Plagas Agrícolas y Forestales; Hemisferio Sur: Montevideo, Uruguay, 1996; p. 129. ISBN 9974-556-96-6. [Google Scholar]
- Casmuz, A.; Juárez, M.L.; Socías, M.G.; Murúa, M.G.; Prieto, S.; Medina, S.; Willink, E.; Gastaminza, G. Revisión de los hospederos del gusano cogollero del maíz, Spodoptera frugiperda (Lepidoptera: Noctuidae). Rev. Soc. Entomol. Argent. 2010, 69, 209–231. [Google Scholar]
- Nagoshi, R.N.; Meagher, R.L. Behavior and distribution of the two fall armyworm host strains in Florida. Fla. Entomol. 2004, 87, 440–449. [Google Scholar] [CrossRef]
- Murúa, M.G.; Vera, M.T.; Abraham, S.; Juaréz, M.L.; Prieto, S.; Head, G.P.; Willink, E. Fitness and mating compatibility of Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) populations from different host plant species and regions in Argentina. Entomol. Soc. Am. 2008, 101, 639–649. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Murúa, M.G.; Hay-Roe, M.; Juárez, M.L.; Willink, E.; Meagher, R.L. Genetic characterization of fall armyworm (Lepidoptera: Noctuidae) host strains in Argentina. J. Econ. Entomol. 2012, 105, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Barros, N.M. Biotic potential and reproductive parameters of Spodoptera eridania (Stoll) (Lepidoptera, Noctuidae) in the laboratory. Rev. Bras. Entomol. 2013, 57, 340–345. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Bortolin, T.M.; Fronza, E.; Pezzi, P.; Luz, P.C.; Barros, N.M. Immature stages of Spodoptera albula (Walker) (Lepidoptera: Noctuidae): Developmental parameters and host plants. An. Acad. Bras. Cienc. 2013, 85, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Barros, N.M. Immature stages of Spodoptera eridania (Lepidoptera: Noctuidae): Developmental parameters and host plants. J. Insect Sci. 2014, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Bortolin, T.M.; Fronza, E.; Pezzi, P.; Luz, P.C.; Barros, N.M. Biotic potential, fertility and life table of Spodoptera albula (Walker) (Lepidoptera: Noctuidae), under controlled conditions. An. Acad. Bras. Cienc. 2014, 86, 723–732. [Google Scholar] [CrossRef]
- Specht, A.; Roque-Specht, V.F. Immature stages of Spodoptera cosmiodes (Lepidoptera: Noctuidae): Developmental parameters and host plants. Zoologia 2016. [Google Scholar] [CrossRef]
- Specht, A.; Montezano, D.G.; Sosa-Gómez, D.R.; Paula-Moraes, S.V.; Roque-Specht, V.F.; Barros, N.M. Reproductive potential of Spodoptera eridania (Stoll) (Lepidoptera: Noctuidae) in the laboratory: Effect of multiple couples and the size. Braz. J. Biol. 2016, 76, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Luttrell, R.G.; Wan, L.; Knighten, K. Variation in susceptibility of Noctuid (Lepidoptera) larvae attacking cotton and soybean to purified endotoxin proteins and commercial formulations of Bacillus thuringiensis. J. Econ. Entomol. 1999, 92, 21–32. [Google Scholar] [CrossRef]
- Sivasupramaniam, S.; Moar, W.J.; Ruschke, L.G.; Osborn, J.A.; Jiang, C.; Sebaugh, J.L.; Brown, G.R.; Shappley, Z.W.; Oppenhuizen, M.E.; Mullins, J.W.; et al. Toxicity and characterization of cotton expressing Bacillus thuringiensis Cry1Ac and Cry2Ab2 proteins for control of lepidopteran pests. J. Econ. Entomol. 2008, 101, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Yano, S.A.C.; Santana, G.K.S.; Neiva, M.M.; Moscardi, F.; Martinelli, S.; Sosa Gómez, D.R. Tolerância de Anticarsia gemmatalis Hübner, Pseudoplusia includens (Walker) e Rachiplusia nu (Guenée) à proteína Cry1Ac. In Proceedings of the 6th Congresso Brasileiro de Soja, Cuiabá, Brazil, 11–14 June 2012; pp. 1–5. [Google Scholar]
- Sosa-Gómez, D.; Omoto, C. Resistencia a insecticidas e otros agentes de controle em artrópodes asociados a cultura da soja. In Soja: Manejo Integrado de Insetos e Outros Artópodes-Praga; Hoffmann-Campo, C.B., Correa-Ferreira, B.S., Moscardi, F., Eds.; Embrapa Press: Brasilia, Brazil, 2012; pp. 673–724. ISBN 978-85-7035-139-5. [Google Scholar]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Farias, J.R.; Andow, D.A.; Horikoshi, R.J.; Sorgatto, R.J.; Fresia, P.; Dos Santos, A.C.; Omoto, C. Field evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop Prot. 2014, 64, 150–158. [Google Scholar] [CrossRef]
- Chandrasena, D.; Signorini, A.; Abratti, G.; Storer, N.P.; Lopez Olaciregui, M.; Alves, A.P.; Pilcher, C.D. Characterization of field-evolved resistance to Bacillus thuringiensis- derived Cry1F δ-endotoxin in Spodoptera frugiperda populations from Argentina. Pest Manag. Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- González, B.J.B. Aspectos Importantes sobre la Evolución y Combate de las Plagas del Algodonero en Colombia. Rev. Peru. Entomol. 1966, 9, 145–155. [Google Scholar]
- Aziz, S.A. Toxicity of certain insecticide standards against the Southern armyworm. J. Econ. Entomol. 1973, 66, 68–70. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, P.A.; Vasquez, C.H. Prueba de laboratorio con 30 insecticidas en larvas de Prodenia eridania (Cramer) (Lepidoptera: Noctuidae) en alfalfa. Idesia 1974, 3, 133–140. [Google Scholar]
- Gloria, B.R. Control químico del gusano ejercito Prodenia sunia (G.) en tomateira. Rev. Peru. Entomol. 1975, 18, 120–123. [Google Scholar]
- Campos, L.E. Toxicidad de cuatro insecticidas contra Spodoptera frugiperda (J.E. Smith) y Prodenia eridania (Cram.) del Valle de Lluta, Chile. Idesia 1982, 6, 13–19. [Google Scholar]
- Savoie, K.L. Alimentación selectiva por especies de Spodoptera (Lepidoptera: Noctuidae) en un campo de frijol con labranza mínima. Turrialba 1988, 38, 67–70. [Google Scholar]
- Zenner de Polanía, I.; Álvarez Rodriguez, J.A.; Arévalo Maldonado, H.A.; Mejía Cruz, R.; Bayona, M.A. Susceptibilidad de cuatro noctuidos plaga (Lepidoptera) al gene Cry1Ac del Bacillus thuringiensis incorporado al algodonero. Rev. Colomb. Entomol. 2008, 34, 41–50. [Google Scholar]
- Santos-Amaya, O.; Delgado-Restrepo, O.; Argüelles, J.; Aguilera-Garramuño, E. Evaluación del comportamiento del complejo Spodoptera con la introducción de algodón transgénico al Tolima, Colombia. Corpoica cienc. Tecnol. Agropecu. 2009, 10, 24–32. [Google Scholar] [CrossRef]
- Levy, R.; Habeck, D.H. Descriptions of the larvae of Spodoptera sunia and S. latifascia with a key to the mature Spodoptera larvae of the Eastern United States. Ann. Entomol. Soc. Am. 1976, 69, 585–588. [Google Scholar] [CrossRef]
- Passoa, S. Color identification of economically important Spodoptera larvae in Honduras (Lepidoptera: Noctuidae). Insecta Mundi. 1991, 5, 185–195. [Google Scholar]
- Pogue, G.M. A world revision of the genus Spodoptera Guenée (Lepidoptera: Noctuidae). Mem. Am. Entomol. Soc. 2002, 43, 1–202. [Google Scholar]
- Navarro, F.R.; Saini, E.D.; Leiva, P.D. Clave pictórica de polillas de interés agrícola, agrupadas por relación de semejanza; INTA-EEA Pergamino: Buenos Aires, Argentina, 2009; p. 100. ISBN 978-987-521-344-9. [Google Scholar]
- Scalora, F.; Casmuz, A.; Cazado, L.; Socías, G.; Tolosa, G.; Aralde, M.; Guchea, M.A.; Fadda, L.; Gómez, M.; Gómez, H.; et al. Evaluación de diferentes insecticidas para el control de la oruga bolillera, Helicoverpa gelotopoeon Dyar (Lepidoptera: Noctuidae). In El Cultivo de la Soja en el Noroeste Argentino: Campaña 2011/2012; Devani, M.R., Ledesma, F., Sánchez, J.R., Eds.; Publicación Especial EEAOC nº 45. EEAOC: San Miguel de Tucumán, Argentina, 2012; pp. 147–151. Available online: http://www.eeaoc.org.ar/upload/publicaciones/archivos/286/20121122085717000000.pdf (accessed on 25 April 2018).
- Murúa, M.G.; Cazado, L.E.; Casmuz, A.; Herrero, M.I.; Villagrán, M.E.; Vera, A.; Sosa Gómez, D.; Gastaminza, G. Species from the Heliothinae complex (Lepidoptera: Noctuidae) in Tucumán, Argentina, and update of geographical distribution of Helicoverpa armigera. J. Insect Sci. 2016, 16, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.I.; Fogliata, S.V.; Vera, A.; Casmuz, A.; Sosa Gómez, D.; Castagnaro, A.P.; Gastaminza, G.; Murúa, M.G. Biological characterization and mating compatibility of Helicoverpa gelotopoeon (D.) (Lepidoptera: Noctuidae) populations from different regions in Argentina. Bull. Entomol. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Pietrantonio, P.V.; Junek, T.A.; Parker, R.; Mott, D.; Siders, K.; Troxclair, N.; Vargas-Camplis, J.; Westbrook, J.K.; Vassiliou, V.A. Detection and evolution of resistance to the pyrethroid cypermethrin in Helicoverpa zea (Lepidoptera: Noctuidae) populations in Texas. Environ. Entomol. 2007, 36, 1174–1188. [Google Scholar] [CrossRef]
- Gao, Y.; Wu, K.; Gould, F.; Shen, Z. Cry2Ab tolerance response of Helicoverpa armigera (Lepidoptera: Noctuidae) populations from Cry1Ac cotton planting region. J. Econ. Entomol. 2009, 102, 1217–1223. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Xu, Z.; Zhu, Y.C.; Huang, F.; Wang, Y.; Li, H.; Gao, C.; Zhou, W.; Shen, J. Evidence of field-evolved resistance to Cry1Ac-expressing Bt cotton in Helicoverpa armigera (Lepidoptera: Noctuidae) in northern China. Pest Manag. Sci. 2010, 66, 155–161. [Google Scholar] [PubMed]
- Bird, L.J.; Downes, S.J. Toxicity and cross-resistance of insecticides to Cry2Ab-resistant and Cry2Ab-susceptible Helicoverpa armigera and Helicoverpa punctigera (Lepidoptera: Noctuidae). J. Econ. Entomol. 2014, 107, 1923–1930. [Google Scholar] [CrossRef] [PubMed]
- Brévault, T.; Tabashnik, B.E.; Carrière, Y. A seed mixture increases dominance of resistance to Bt cotton in Helicoverpa zea. Sci. Rep. 2015, 5, 9807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reisig, D.D.; Reay-Jones, F.P.F. Inhibition of Helicoverpa zea (Lepidoptera: Noctuidae) growth by transgenic corn expressing Bt toxins and development of resistance to Cry1Ab. Environ. Entomol. 2015, 44, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, O.; Sorgatto, R.J.; Barbosa, A.D.; Domingues, F.A.; Dourado, P.M.; Carvalho, R.A.; Martinelli, S.; Head, G.P.; Omoto, C. Low susceptibility of Spodoptera cosmiodes, Spodoptera eridania and Spodoptera frugiperda (Lepidoptera: Noctuidae) to genetically-modified soybean expressing Cry1Ac protein. Crop Prot. 2014, 58, 33–40. [Google Scholar] [CrossRef]
- Informativo Céleres. Available online: http://www.celeres.com.br/category/analise-de-safra-soja/ (accessed on 23 May 2018).
- Drees, B.M.; Rice, M.E. The vertical beat sheet: A new device for sampling soybeans insects. J. Econ. Entomol. 1985, 78, 1507–1510. [Google Scholar] [CrossRef]
- Velasco de Estacul, M.; Barral, J.M.; Orfila, R.N. Taxonomía, especificidad y caracteres biológicos diferenciados del complejo de especies denominadas “oruga del capullo” del algodón, “oruga de la espiga” del maíz, “oruga del brote” del tabaco y “bolillera” del lino. Rev. Invest. Agropec. Ser. Patol. Veg. 1969, 6, 19–68. [Google Scholar]
- Zenker, M.M.; Specht, A.; Corseuil, E. Estagios imaturos de Spodoptera cosmiodes (Walker), (Lepidoptera: Noctuidae). Rev. Bras. Zool. 2007, 24, 99–107. [Google Scholar] [CrossRef]
- Murúa, M.G.; Virla, E.G.; Defagó, V. Evaluación de cuatro dietas artificiales para la cría de Spodoptera frugiperda (Lep.: Noctuidae) destinada a mantener poblaciones experimentales de himenópteros parasitoides. Bol. San. Veg. Plagas 2003, 29, 43–51. [Google Scholar]
- Fehr, W.R.; Caviness, C.E.; Burmood, D.T.; Pennington, J.S. Stage of development descriptions for soybeans, Glycine max (L.) Merrill. Crop Sci. 1971, 11, 929–931. [Google Scholar] [CrossRef]
- Kogan, M.; Turnipseed, S.G. Soybean growth and assessment of damage by arthropods. In Sampling Methods in Soybean Entomology; Kogan, M., Herzog, D.C., Eds.; Springer Press: New York, NY, USA, 1980; p. 17. ISBN 978-1-4613-8069-6. [Google Scholar]
- Zar, J.H. Biostatistical Analysis; Prentice Hall: Upper Saddle River, NJ, USA, 1999; p. 750, ISBN-13 978-0321656865. [Google Scholar]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; González, L.; Tablada, M.; Robledo, C.W. InfoStat, Versión 2008; Grupo InfoStat, FCA, Universidad Nacional de Córdoba: Córdoba, Argentina, 2008. [Google Scholar]
- Justiniano, W.; Fernandes, M.G.; Tigre Pereira Viana, C.L. Diversity, composition and population dynamics of arthropods in the genetically modified soybeans Roundup Ready® RR1 (GT 40-3-2) and Intacta RR2 PRO® (MON87701 x MON89788). J. Agric. Sci. 2014, 6, 33–44. [Google Scholar] [CrossRef]
- Massoni, F.A.; Schlie, G.; Frana, J.E. Cultivo de soja Bt (RR2 PRO) y convencional (RR1) expuestos a poblaciones naturales de organismos plaga y depredadores. Información Técnica de Cultivos de Verano. Campaña 2013/2014, INTA-Estación Experimental Agropecuaria Rafaela, Argentina. Publ. Misc. 2015, 130, 98–103. Available online: http://www.aapresid.org.ar/wp-content/uploads/2015/01/Soja-Bt-vs-convencional-expuestos-a-organismos-plaga-y-depredadores_MASSONI_F-4.pdf (accessed on 22 May 2018).
- Perotti, E.; Russo, R.; López, R.; Zari, F.; Pradolini, E.; Sanmarti, N.; Maccari, G.; Boero, L.; Gamundi, J. Evaluación de estrategias de manejo de refugios asociados a cultivos de soja Bt sobre plagas blanco, no blanco y sus depredadores. Para Mejorar Producción Cultivos Estivales 2014, 52, 177–182. Available online: http://inta.gob.ar/sites/default/files/script-tmp-inta-evaluacin-estrategias-manejo-refugios-asociados-.pdf (accessed on 22 May 2018).
- Forti, L.A.; Sosa-Gómez, D.R. Suscetibilidade relativa de Spodoptera frugiperda, S. eridania, S. albula, S. cosmiodes e Anticarsia gemmatalis a os inseticidas chlorantraniliprole e flubendiamida. In Proceedings of the 9th Jornada Acadêmica da Embrapa Soja, Londrina, Brazil, 12 June 2014; pp. 126–129. Available online: https://ainfo.cnptia.embrapa.br/digital/bitstream/item/105824/1/JORN.2014.pdf (accessed on 22 May 2018).
- Pastrana, J.A. Los Lepidópteros Argentinos, Sus Plantas Hospedadoras y Otros Sustratos Alimenticios; Sociedad Entomológica Argentina: Buenos Aires, Argentina, 2004; p. 334. ISBN 987-21319-0-2. [Google Scholar]
- Sparks, A. A review of the biology of the fall armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Silva, M.D.; Bueno, F.A.; Stecca, S.C.; Andrade, K.; Neves, J.O.M.P.; Oliveira, N.M.C. Biology of Spodoptera eridania and Spodoptera cosmioides (Lepidoptera: Noctuidae) on different host plants. Fla. Entomol. 2017, 100, 752–760. [Google Scholar] [CrossRef]
- Angulo, A.O.; Olivares, T.S.; Weigert, G.T.H. Estados Inmaduros de Lepidópteros Noctuídos de Importancia Agrícola y Forestal en Chile y Claves Para su Identificación (Lepidoptera: Noctuidae); Universidad de Concepción: Concepción, Chile, 2008; p. 114. [Google Scholar]
Figure 1.
Percentage of defoliation produced by Helicoverpa gelotopoeon, Spodoptera albula, Spodoptera cosmiodes, Spodoptera eridania, and Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae in different soybean treatments (T1: Bt soybean, T2: non-Bt soybean with seed treatment, T3: non-Bt soybean without seed treatment). Means ± SE within species accompanied by different letters indicate significant differences (Tukey test, p < 0.05).
Figure 1.
Percentage of defoliation produced by Helicoverpa gelotopoeon, Spodoptera albula, Spodoptera cosmiodes, Spodoptera eridania, and Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae in different soybean treatments (T1: Bt soybean, T2: non-Bt soybean with seed treatment, T3: non-Bt soybean without seed treatment). Means ± SE within species accompanied by different letters indicate significant differences (Tukey test, p < 0.05).


{kind=link}
Table 1.
Summary of previous studies of the Spodoptera complex and Helicoverpa gelotopoeon (Lep.: Noctuidae), where the performance of each species was evaluated using different crops and/or artificial diet in field and laboratory studies.
Table 1.
Summary of previous studies of the Spodoptera complex and Helicoverpa gelotopoeon (Lep.: Noctuidae), where the performance of each species was evaluated using different crops and/or artificial diet in field and laboratory studies.
| Study | Species * | Identity of Species ** | Environmental Conditions (Controlled or Field Conditions) | Type of Infestation | Crop or Artificial Diet Used | Evaluations | Impacts |
|---|---|---|---|---|---|---|---|
| Present study | Sf, Se, Sa, Sc, Hg | individual | controlled (greenhouse) | artificial | soybean (Bt and non-Bt) | Performance and potencial risk | Bt soybean had the best control of Hg and a suppressive effect on Sf and Sa. Se and Sc were not susceptible to the Bt soybean. Se represents a potential risk in soybean crops |
| [60] | Sc, Se, Sf | individual | controlled | artificial | soybean (Bt and non-Bt) and diet | Susceptibility to Cry1Ac protein | Bt soybean showed poor control of Sc, Se, Sf |
| [16] | Sf, Se, Sc | individual | controlled | artificial | soybean genotypes | Larva consumption foliage | Sc defoliated nearly twice the leaf area of Sa and Sf |
| [5] | S. spp. | general | field conditions | natural | soybean | Different chemical alternatives for the management of the complex pest | Diamide + neonicotinoid had the best control |
| [6] | Sc | individual | field conditions | natural | soybean | Moment of application of different insecticides | Early application of diamide delayed the damage of larvae |
| [7] | H. spp., S. spp. | general | field conditions | natural | soybean (Bt and non-Bt) | Pest management | Early application of diamide delayed the damage of larvae |
| [8] | H. spp. | general | field conditions | natural | soybean (Bt and non-Bt) | Strategies for prevention of insect resistance | Low presence of H. spp. in Bt soybean and refuge |
| [19] | S. spp., H. spp. | general | field conditions | natural | soybean (Bt and non-Bt) | Behavior of Bt soybean on the pests and its predators | Bt soybean provided a good control of S. spp. and H. spp. and did not affect its predators |
| [18] | Se | individual | controlled | artificial | soybean cultivars (non-Bt) | Development, survival, and reproductive capacity | The development of Se was affected by the cultivar |
| [73] | Sf, Se, Sa, Sc | individual | controlled | artificial | diet | Susceptibility to chlorantraniliprole and flubendiamide | Chlorantraniliprole showed a higher mortality than flubendiamide for all Lepidoptera species tested |
| [70] | S. spp., Sf | general | field conditions | natural | soybean (Bt and non-Bt) | Diversity, composition, and population dynamics | Bt soybean reduced the target insect pests and favored populations of natural enemies |
| [71] | Sf, Sc, Hg | individual | field conditions | natural | soybean (Bt and non-Bt) | Insect abundance | Densities of the species were low in both treatment |
| [27] | Sa | controlled | artificial | diet | Developmental parameters and host plants | Complete detail of biological parameters of Sa and 55 host plant species of Sa are listed | |
| [26] | Se | individual | controlled | artificial | diet | Biotic potential and reproductive parameters | Complete detail of reproductive and population parameters of Se |
| [28] | Se | individual | controlled | artificial | diet | Developmental parameters and host plants | Complete detail of biological parameters of Se and 202 host plant species of Se are listed |
| [29] | Sa | controlled | artificial | diet | Biotic potential, life table parameters and fertility | Complete detail of reproductive and population parameters of Sa | |
| [72] | Sf, Sc, Hg | individual | field conditions | natural | soybean (Bt and non-Bt) | Strategies of refuge management | The management of refuges with selective insecticides and high persistence allowed to reduce the number of applications and to achieve greater survival of predators and adults of target pests |
| [15] | Se | individual | controlled | artificial | cotton, soybean, and Ipomoea grandifolia (L.) (Convolvulaceae) | Biology on different host plant | Soybean was the least suitable for the development of Se, and I. grandifolia was shown a suitable alternate host for Se |
| [20] | Sc | individual | controlled | artificial | corn (Bt and non-Bt), soybean (Bt and non-Bt), and diet | Development and reproduction | Bt and non-Bt corn adversely affect the development of Sc, and Bt soybean did not affect its biology, suggesting that Sc has major potential to become an important pest in Bt soybean crops |
| [76] | Se, Sc | individual | controlled | artificial | soybean, cotton, corn, Triticum aestivum (L.) (Poaceae), Avena sativa (L.), and diet | Biology on different host plants | Soybean and cotton were more suitable hosts for the development of Se and Sc |
* Species: (only the species involved in the present study are mentioned) S. spp.: Spodoptera species; Sf: S. frugiperda; Se: S. eridania; Sa: S. albula; Sc: S. cosmioides; H. spp.: Helicoverpa species; Hg: H. gelotopoeon. ** General: specific identity of each species was not considered in the study; Individual: specific identity of each species was considered in the study.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Murúa, M.G.; Vera, M.A.; Herrero, M.I.; Fogliata, S.V.; Michel, A. Defoliation of Soybean Expressing Cry1Ac by Lepidopteran Pests. Insects 2018, 9, 93. https://doi.org/10.3390/insects9030093
AMA Style
Murúa MG, Vera MA, Herrero MI, Fogliata SV, Michel A. Defoliation of Soybean Expressing Cry1Ac by Lepidopteran Pests. Insects. 2018; 9(3):93. https://doi.org/10.3390/insects9030093
Chicago/Turabian StyleMurúa, María G., Martín A. Vera, María I. Herrero, Sofia V. Fogliata, and Andrew Michel. 2018. "Defoliation of Soybean Expressing Cry1Ac by Lepidopteran Pests" Insects 9, no. 3: 93. https://doi.org/10.3390/insects9030093
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.