Detection of Volatile Constituents from Food Lures by Tephritid Fruit Flies

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Commercial Baits
- brewer’s yeast waste (St. George brewery, Addis Ababa, Ethiopia), is a byproduct from the brewing process that is normally tossed off. It is rich in yeast and has been exploited as an alternative source of protein for use in tephritid fruit fly management [28];
- baker’s yeast (Saccharomyces cervisiae) (Jästbolaget AB, Sollentuna, Sweden), activated with sugar, is a common diet for adult tephritid fruit flies. It is also an essential component of tephritid larval media (see Section 2.1);
- GF-120 success bait (Dow AgroSciences, Indianapolis, IN, USA), is a combination of spinosad, enzymatically hydrolyzed protein, sugars, adjuvants, and conditoners that attracts fruit flies [29];
- Anamed (ISCA Technologies, Riverside, CA, USA), is a protein-based attractant that targets a wide range of tephritid fruit flies;
2.3. Volatile Collection and GC-MS
2.4. Electrophysiology
2.5. Chemical Analysis
2.6. Synthetic Blends
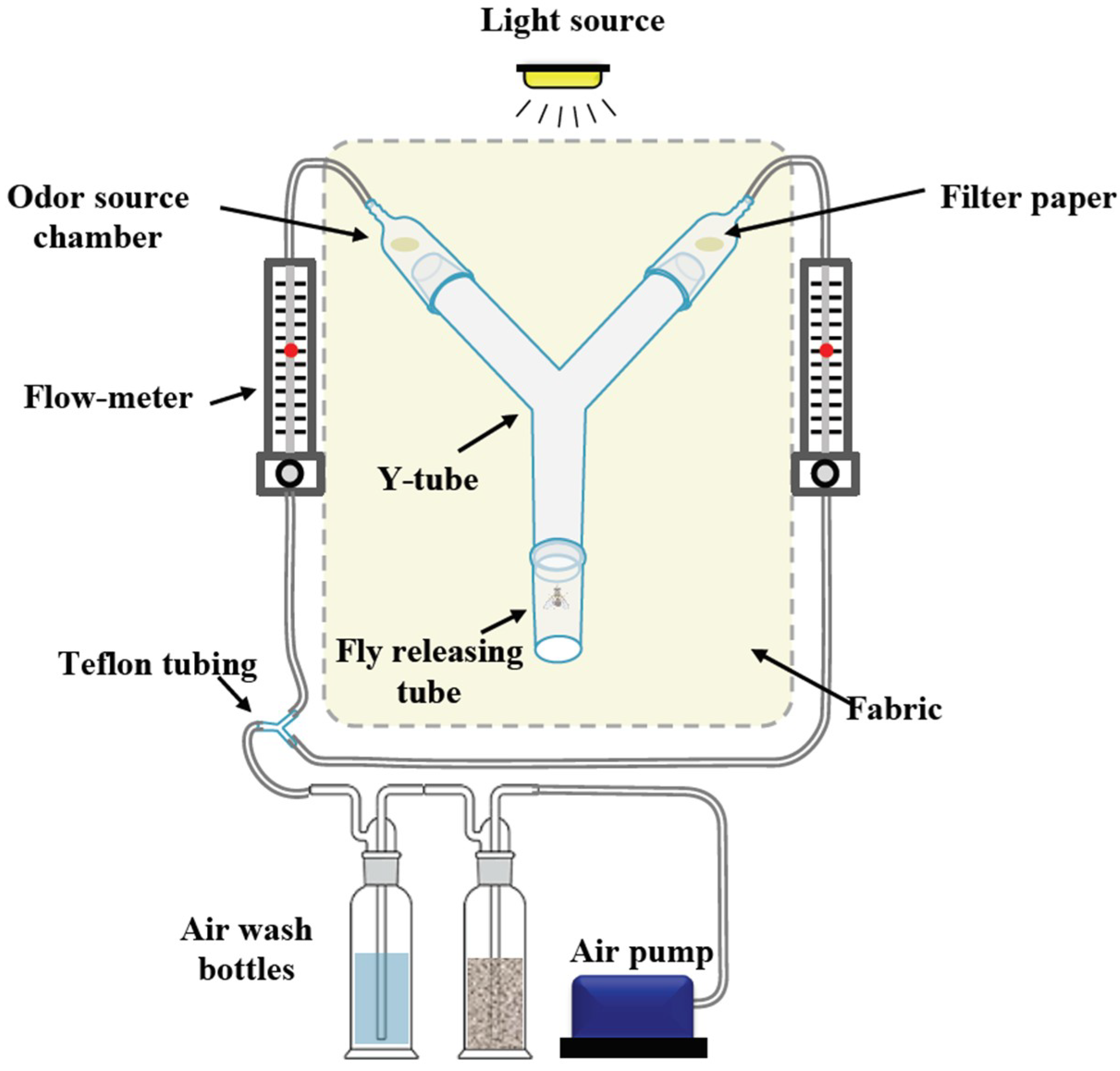
2.7. Olfactometer Experiments
2.8. Field Experiments
2.9. Statistical Analysis
3. Results
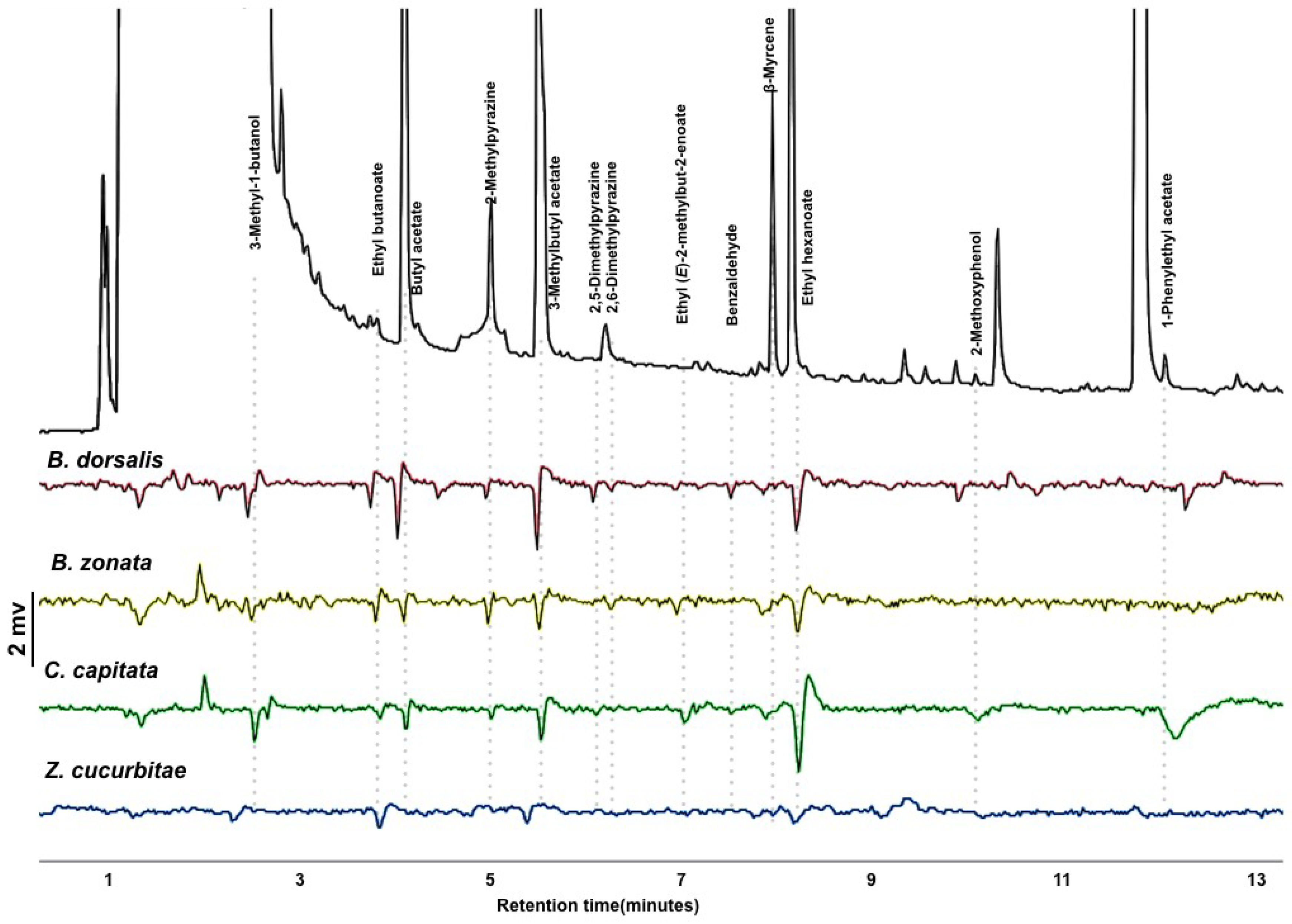
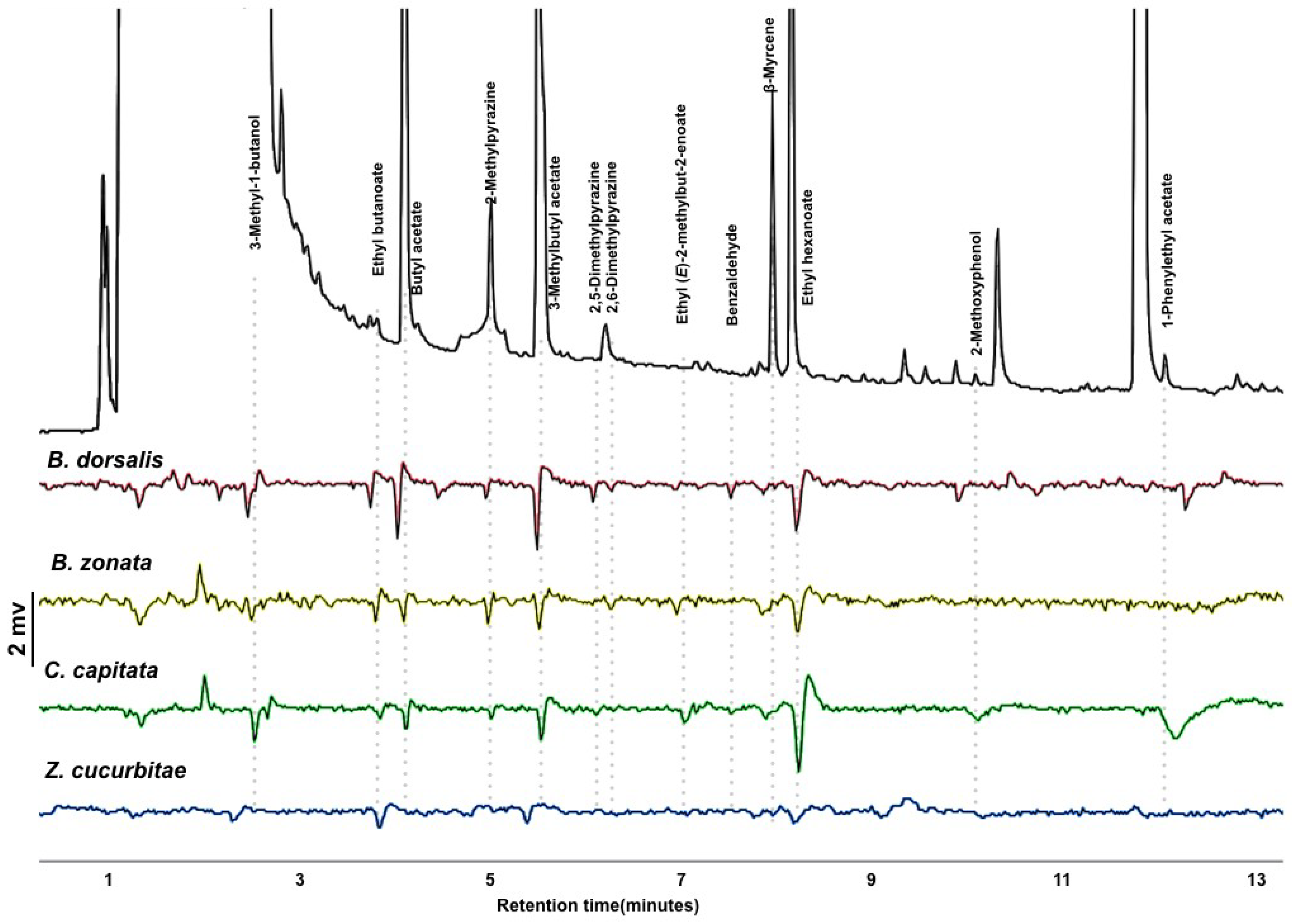
3.1. GC-EAD and Volatile Identification
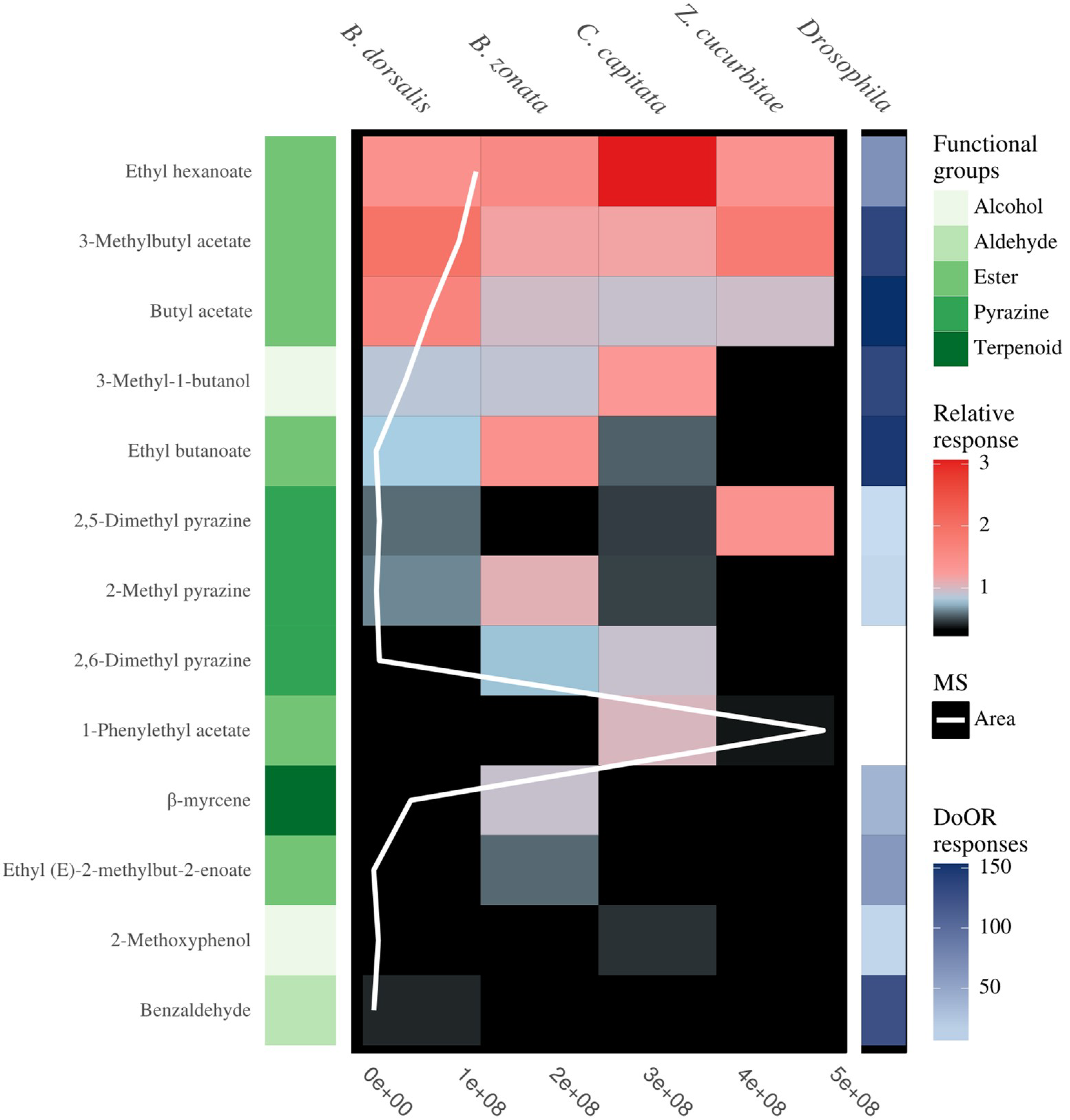
3.2. Antennal Sensitivity and DoOR Comparison
3.3. Olfactometer Experiments
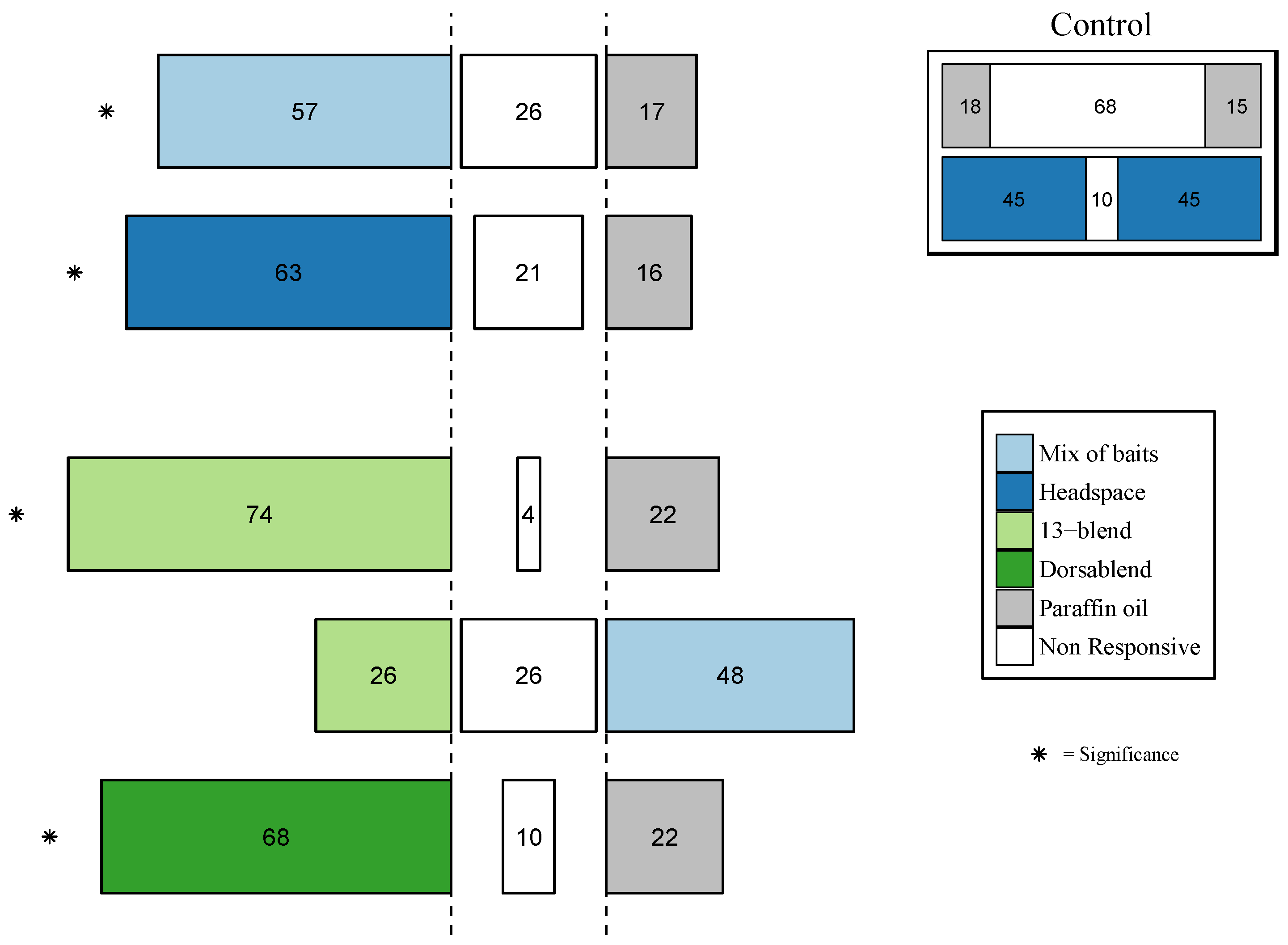
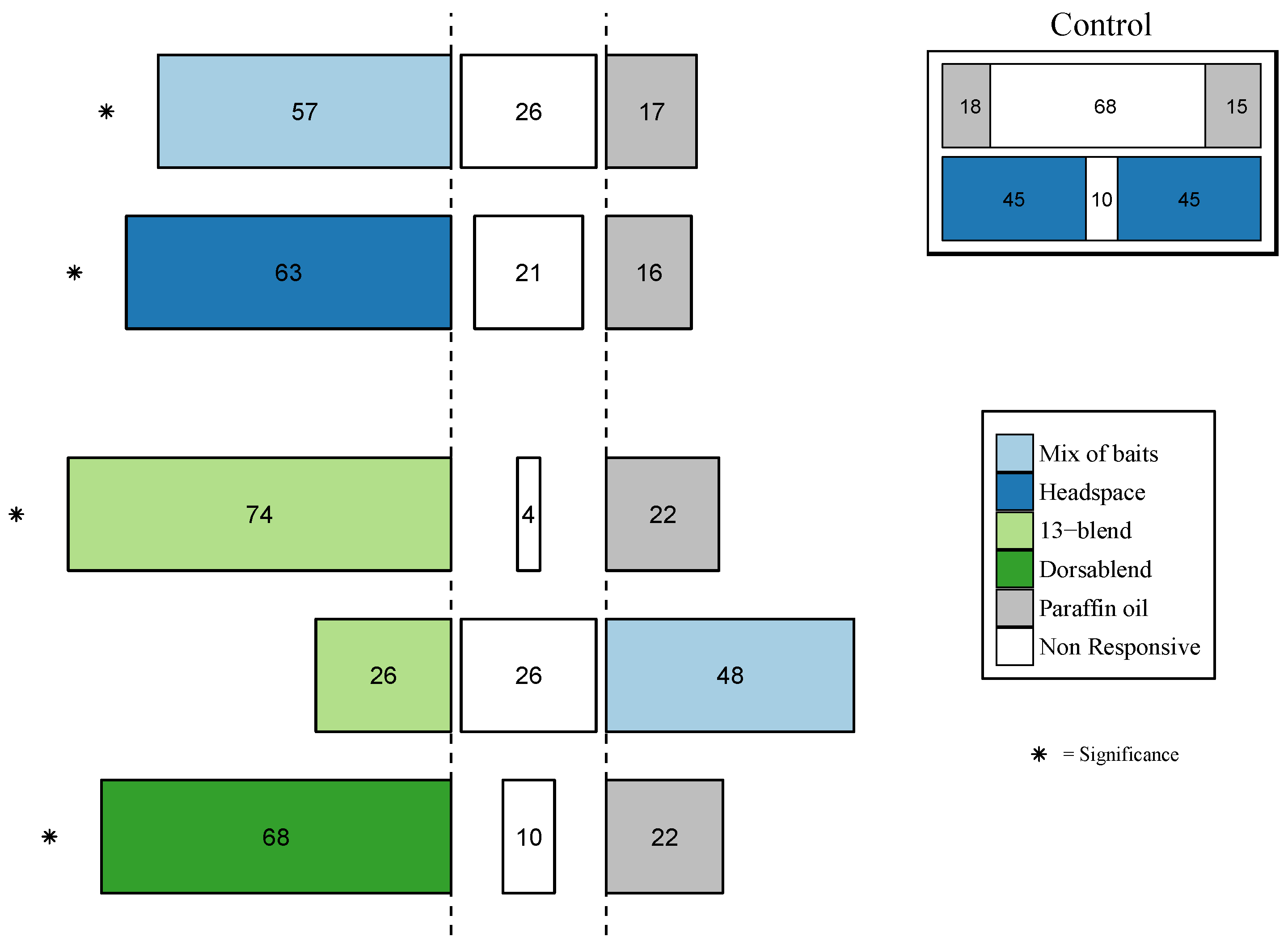
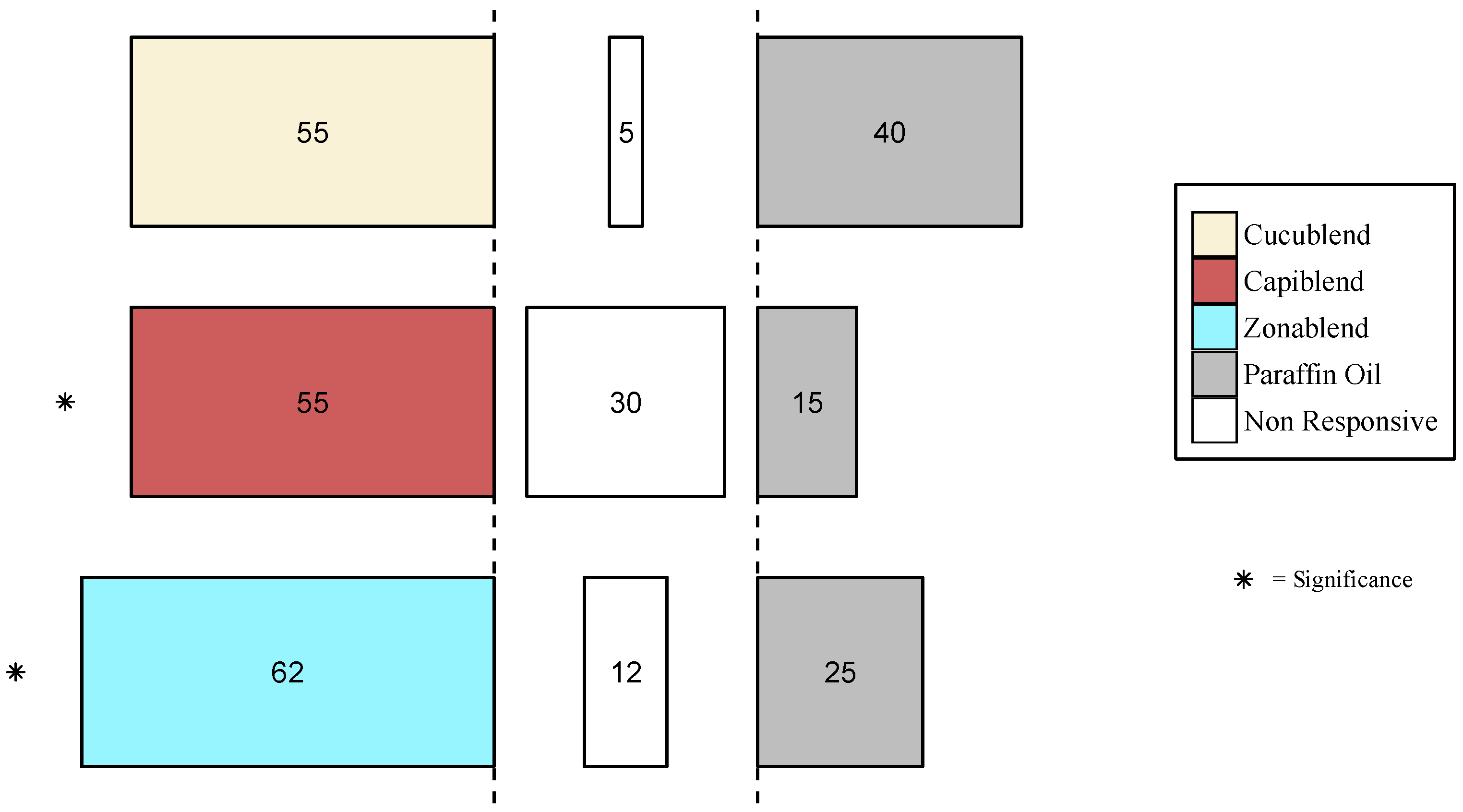
3.3.1. Behavioral Response of B. dorsalis
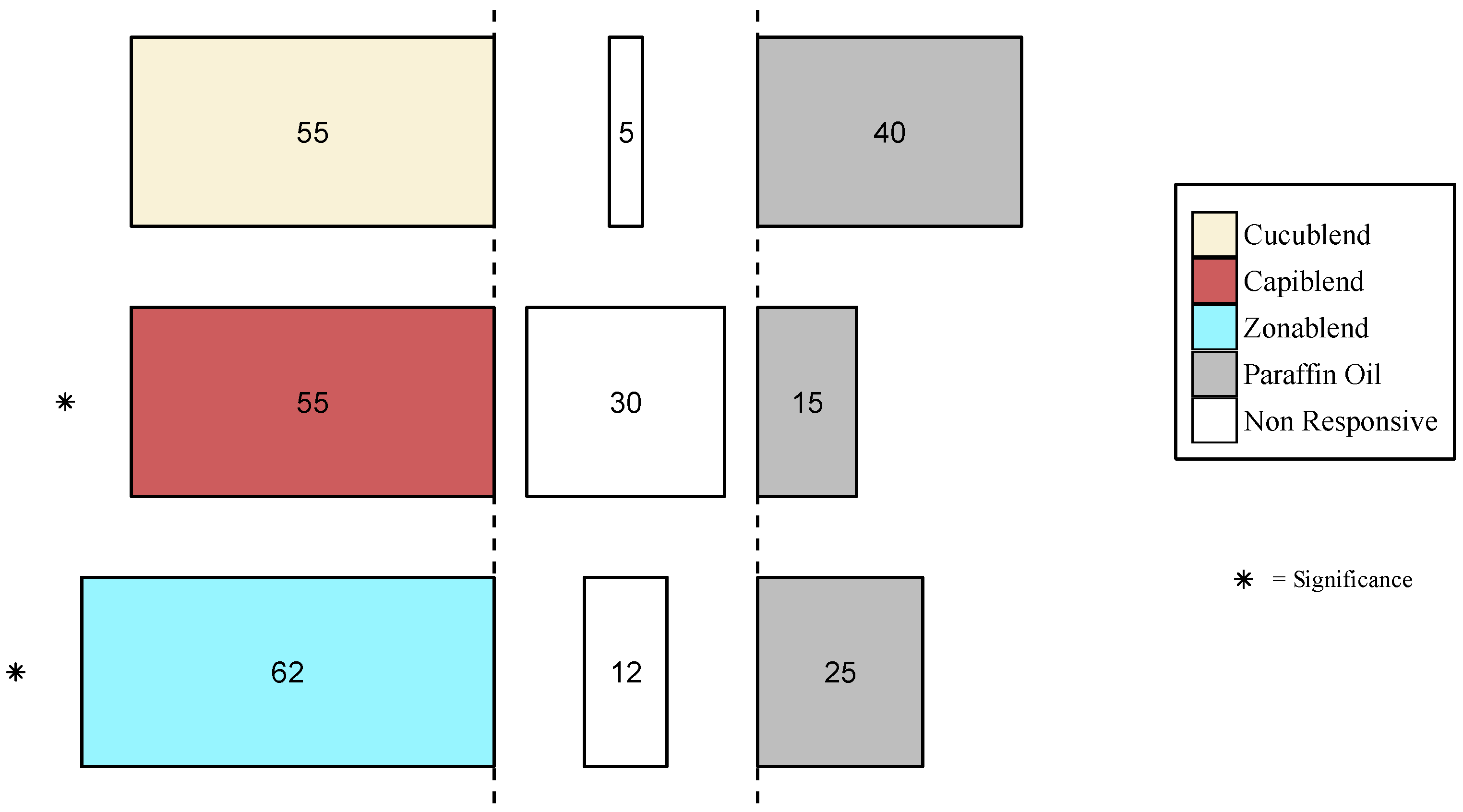
3.3.2. Behavioral Response of other Tephritid Species
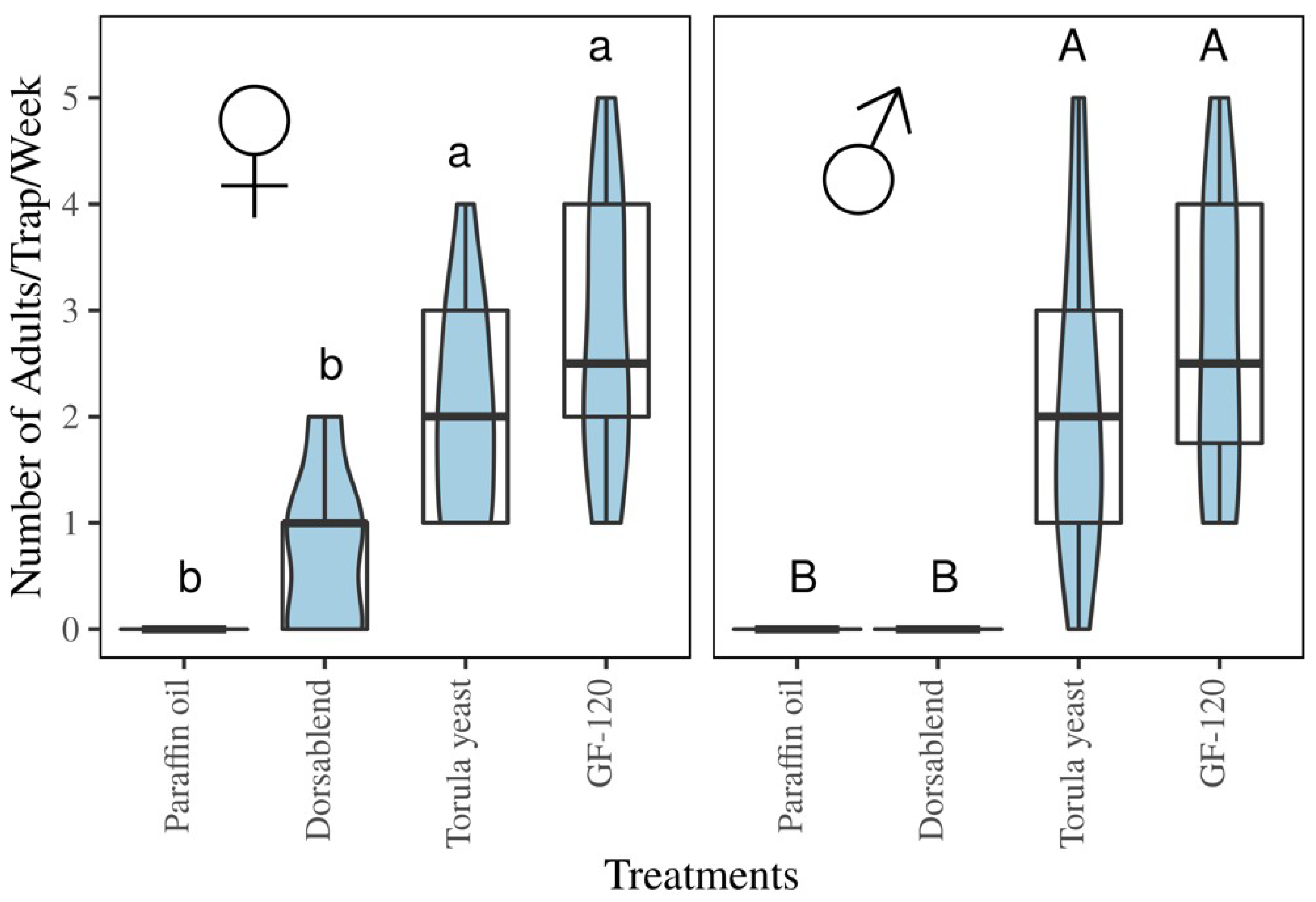
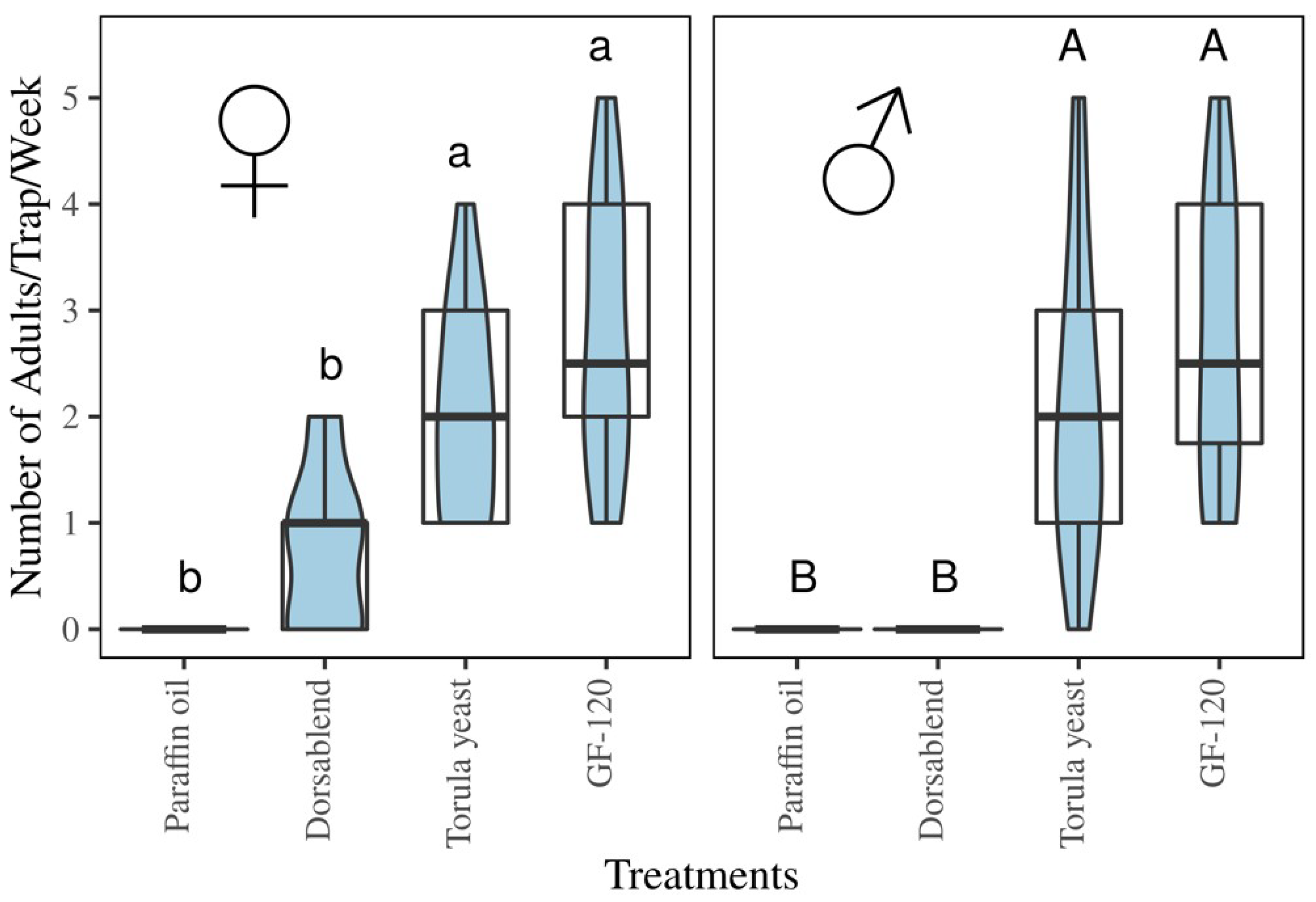
3.4. Field Experiment
Trap Catches of B. dorsalis with Protein-Based Baits
4. Discussion
5. Future Research
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Duyck, P.F.; David, P.; Quilici, S. A review of relationships between interspecific competition and invasions in fruit flies (Diptera: Tephritidae). Ecol. Entomol. 2004, 29, 511–520. [Google Scholar] [CrossRef] [Green Version]
- White, I.M.; Meyer, M.D.; Stonehouse, J. A Review of Native and Introduced Fruit Flies (Diptera, Tephritidae) in the Indian Ocean Islands of Mauritius, Réunion, Rodrigues and Seychelles. In Proceedings of the Indian Ocean Commission Regional Fruit Fly Symposium, Flic en Flac, Mauritius, 5–9 June 2000; Price, N.S., Seewooruthun, I., Eds.; Indian Ocean Commission: Quatre Bornes, Mauritius, 2000; pp. 15–21. [Google Scholar]
- Lux, S.A.; Copeland, R.S.; White, I.M.; Manrakhan, A.; Billah, M.K. A new invasive fruit fly species from the Bactrocera dorsalis (Hendel) group detected in East Africa. Int. J. Trop. Insect Sci. 2003, 23, 355–361. [Google Scholar] [CrossRef]
- De Meyer, M.; Delatte, H.; Mwatawala, M.; Quilici, S.; Vayssieres, J.F.; Virgilio, M.A. Review of the current knowledge on Zeugodacus cucurbitae (Coquillett) (Diptera, Tephritidae) in Africa, with a list of species included in Zeugodacus. ZooKeys 2015, 540, 539–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekesi, S.; Billah, M.K.; Nderitu, P.W.; Lux, S.A.; Rwomushana, I. Evidence for competitive displacement of Ceratitis cosyra by the invasive fruit fly Bactrocera invadens (Diptera: Tephritidae) on mango and mechanisms contributing to the displacement. J. Econ. Entomol. 2009, 102, 981–991. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, B. The biology of dacine fruit flies. Ann. Rev. Entomol. 1987, 32, 115–144. [Google Scholar] [CrossRef]
- Drew, R.A.I.; Yuval, B. The evolution of fruit fly feeding behavior. In Fruit Flies (Tephritidae); Martin, A., Allen, L.N., Eds.; CRC Press: Boca Raton, FL, USA, 1999; pp. 731–749. ISBN 0-8493-1275-2. [Google Scholar] [CrossRef]
- Manrakhan, A.; Lux, S.A. Contribution of natural food sources to reproductive behaviour, fecundity and longevity of Ceratitis cosyra, C. Fasciventris and C. capitata (Diptera: Tephritidae). Bull. Entomol. Res. 2006, 96, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Christenson, L.D.; Foote, R.H. Biology of fruit flies. Ann. Rev. Entomol. 1960, 5, 171–192. [Google Scholar] [CrossRef]
- Shelly, T.E.; Kurashima, R.S. Capture of Mediterranean fruit flies and melon flies (Diptera: Tephritidae) in food-baited traps in Hawaii. Proc. Hawaii Entomol. Soc. 2016, 48, 71–84. [Google Scholar]
- Epsky, N.D.; Kendra, P.E.; Schnell, E.Q. History and development of food-based attractants. In Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies; Shelly, T., Epsky, N., Jang, E., Reyes-Flores, J., Vargas, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 75–118. ISBN 978-94-017-9192-2. [Google Scholar] [CrossRef]
- Tan, K.H.; Nishida, R.; Jang, E.B.; Shelly, T.E. Pheromones, male lures, and trapping of tephritid fruit flies. In Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies; Shelly, T., Epsky, N., Jang, E., Reyes-Flores, J., Vargas, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 15–74. ISBN 978-94-017-9192-2. [Google Scholar] [CrossRef]
- Biasazin, T.D. The Chemical Ecology of the Oriental Fruit Fly Bactrocera dorsalis and the Potential for Novel Odor-Based Management Tools. Ph.D. Thesis, Swedish University of Agricultural Sciences, Alnarp, Sweden, September 2017. [Google Scholar]
- Davis, T.S.; Crippen, T.L.; Hofstetter, R.W.; Tomberlin, J.K. Microbial volatile emissions as insect semiochemicals. J. Chem. Ecol. 2013, 39, 840–859. [Google Scholar] [CrossRef] [PubMed]
- Drew, R.A.I.; Courtice, A.C.; Teakle, D.S. Bacteria as a natural source of food for adult fruit flies (Diptera: Tephritidae). Oecologia 1983, 60, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Ben-Yosef, M.; Jurkevitch, E.; Yuval, B. Effect of bacteria on nutritional status and reproductive success of the Mediterranean fruit fly Ceratitis capitata. Physiol. Entomol. 2008, 33, 145–154. [Google Scholar] [CrossRef]
- Yuval, B.; Ben-Ami, E.; Behar, A.; Ben-Yosef, M.; Jurkevitch, E. The Mediterranean fruit fly and its bacteria-potential for improving sterile insect technique operations. J. Appl. Entomol. 2010, 137, 39–42. [Google Scholar] [CrossRef]
- Van den Bosch, T.J.M.; Welte, C.U. Detoxifying symbionts in agriculturally important pest insects. Microb. Biotechnol. 2016, 10, 531–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd, A.C.; Drew, R.A.I.; Teakle, D.S.; Hayward, A.C. Bacteria associated with some Dacus species (Diptera: Tephritidae) and their host fruit in Queensland. Aust. J. Biol. Sci. 1986, 39, 361–368. [Google Scholar] [CrossRef]
- Bateman, M.; Morton, T. The importance of ammonia in proteinaceous attractants for fruit flies (Family: Tephritidae). Aust. J. Agric. Res. 1981, 32, 883. [Google Scholar] [CrossRef]
- Morton, T.; Bateman, M. Chemical studies on proteinaceous attractants for fruit flies, including the identification of volatile constituents. Aust. J. Agric. Res. 1981, 32, 905. [Google Scholar] [CrossRef]
- Heath, R.R.; Epsky, N.D.; Guzman, A.; Dueben, B.D.; Manukian, A.; Meyer, W.L. Development of a dry plastic insect trap with food-based synthetic attractant for the Mediterranean and Mexican fruit flies (Diptera: Tephritidae). J. Econ. Entomol. 1995, 88, 1307–1315. [Google Scholar] [CrossRef]
- Díaz-Fleischer, F.; Arredondo, J.; Flores, S.; Montoya, P.; Aluja, M. There is no magic fruit fly trap: Multiple biological factors influence the response of adult Anastrepha ludens and Anastrepha obliqua (Diptera: Tephritidae) individuals to multilure traps baited with BioLure or NuLure. J. Econ. Entomol. 2009, 102, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Leblanc, L.; Vargas, R.I.; Rubinoff, D. Attraction of Ceratitis capitata (Diptera: Tephritidae) and endemic and introduced non-target insects to BioLure bait and its individual components in Hawaii. Environ. Entomol. 2010, 39, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Leblanc, L.; Vargas, R.I.; Rubinoff, D. Captures of pest fruit flies (Diptera: Tephritidae) and non-target insects in BioLure and Torula yeast traps in Hawaii. Environ. Entomol. 2010, 39, 1626–1630. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.R.; Epsky, N.D.; Dueben, B.D.; Rizzo, J.; Jeronimo, F. Adding methyl-substituted ammonia derivatives to a food-based synthetic attractant on capture of the Mediterranean and Mexican fruit flies (Diptera: Tephritidae). J. Econ. Entomol. 1997, 90, 1584–1589. [Google Scholar] [CrossRef]
- IAEA. Development of improved attractants and their integration into fruit fly SIT management programmes. In Proceedings of the Research Coordination Meeting, Vienna, Austria, 5–7 May 2005; p. 238. [Google Scholar]
- Ekesi, S.; Tanga, C.M. Waste brewer’s yeast as an alternative source of protein for use as a bait in the management of tephritid fruit flies. In Fruit Fly Research and Development in Africa-Towards a Sustainable Management Strategy to Improve Horticulture; Ekesi, S., Mohamed, S., Eds.; Springer: Cham, Switzerland, 2016; pp. 293–306. ISBN 9783-3-319-43224-3. [Google Scholar] [CrossRef]
- Mangan, R.L.; Moreno, D.S.; Thompson, G.D. Bait dilution, spinosad concentration, and efficacy of GF-120 based fruit fly sprays. Crop Prot. 2006, 25, 125–133. [Google Scholar] [CrossRef]
- Ekesi, S.; Meyer, M.D.; Mohamed, S.A.; Virgilio, M.; Borgemeister, C. Taxonomy, ecology, and management of native and exotic fruit fly species in Africa. Annu. Rev. Entomol. 2016, 61, 219–238. [Google Scholar] [CrossRef] [PubMed]
- Piñero, J.C.; Souder, S.K.; Smith, T.R.; Vargas, R.I. Attraction of Bactrocera cucurbitae and Bactrocera dorsalis (Diptera: Tephritidae) to beer waste and other protein sources laced with ammonium acetate. Fla. Entomol. 2017, 100, 70–76. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2009. [Google Scholar]
- Münch, D.; Galizia, C.G. DoOR 2.0-comprehensive mapping of Drosophila melanogaster odorant responses. Sci. Rep. 2016, 6, 21841. [Google Scholar] [CrossRef] [PubMed]
- Grabe, V.; Baschwitz, A.; Dweck, H.K.; Lavista-Llanos, S.; Hansson, B.S.; Sachse, S. Elucidating the neuronal architecture of olfactory glomeruli in the Drosophila antennal lobe. Cell Rep. 2016, 16, 3401–3413. [Google Scholar] [CrossRef] [PubMed]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Becher, P.G.; Flick, G.; Rozpędowska, E.; Schmidt, A.; Hagman, A.; Lebreton, S.; Larsson, M.C.; Hansson, B.S.; Piškur, J.; Witzgall, P.; et al. Yeast, not fruit volatiles mediate Drosophila melanogaster attraction, oviposition and development. Funct. Ecol. 2012, 26, 822–828. [Google Scholar] [CrossRef]
- Lee, C.J.; Demilo, A.B.; Moreno, D.S.; Martinez, A.J. Analysis of the volatile components of a bacterial fermentation attractive to the Mexican fruit fly (Anastrepha ludens). J. Agric. Food Chem. 1995, 43, 1348–1351. [Google Scholar] [CrossRef]
- Robacker, D.C.; Martinez, A.J.; Garcia, J.A.; Bartelt, R.J. Volatiles attractive to the Mexican fruit fly (Diptera: Tephritidae) from eleven bacteria taxa. Fla. Entomol. 1998, 81, 497. [Google Scholar] [CrossRef]
- Scheidler, N.H.; Liu, C.; Hamby, K.A.; Zalom, F.G.; Syed, Z. Volatile codes: Correlation of olfactory signals and reception in Drosophila-yeast chemical communication. Sci. Rep. 2015, 5, 14059. [Google Scholar] [CrossRef] [PubMed]
- Mazzetto, F.; Elena, G.; Violetta, V.; Michail, S.; Marianna, P.; Sven, M.; Daniele, D.; Alberto, A. Olfactory attraction of Drosophila suzukii by symbiotic acetic acid bacteria. J. Pest Sci. 2016, 89, 783–792. [Google Scholar] [CrossRef]
- Bohman, B.; Phillips, R.D.; Menz, M.H.M.; Berntsson, B.W.; Flematti, G.R.; Barrow, R.A.; Dixon, K.W.; Peakall, R. Discovery of pyrazines as pollinator sex pheromones and orchid semiochemicals: Implications for the evolution of sexual deception. New Phytol. 2014, 203, 939–952. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.; Herbert, R.H.; Lomer, R.A. Chemical components of the rectal gland secretions of male Dacus cucurbitae, the melon fly. Experientia 1982, 38, 232–233. [Google Scholar] [CrossRef]
- Baker, P.S.; Howse, P.E.; Ondarza, R.N.; Reyes, J. Field trials of synthetic sex pheromone components of the male Mediterranean fruit fly (Diptera: Tephritidae) in Southern Mexico. J. Econ. Entomol. 1990, 83, 2235–2245. [Google Scholar] [CrossRef]
- Benelli, G.; Daane, K.M.; Canale, A.; Niu, C.-Y.; Messing, R.H.; Vargas, R.I. Sexual communication and related behaviours in Tephritidae: Current knowledge and potential applications for integrated pest management. J. Pest Sci. 2014, 87, 385–405. [Google Scholar] [CrossRef]
- Schulz, S.; Dickschat, J. Bacterial volatiles: The smell of small organisms. Nat. Prod. Rep. 2007, 24, 814–842. [Google Scholar] [CrossRef] [PubMed]
- Epsky, N.D.; Heath, R.R.; Dueben, B.D.; Lauzon, C.R.; Proveaux, A.T.; Maccollom, G.B. Attraction of 3-methyl-1-butanol and ammonia identified from Enterobacter agglomerans to Anastrepha suspensa. J. Chem. Ecol. 1998, 24, 1867–1880. [Google Scholar] [CrossRef]
- Buttery, R.G.; Ling, L.C.; Teranishi, R.; Mon, T.R. Insect attractants: Volatiles of hydrolyzed protein insect baits. J. Agric. Food Chem. 1983, 31, 689–692. [Google Scholar] [CrossRef]
- Robacker, D.C.; Garcia, J.A.; Hart, W.G. Attraction of a laboratory strain of Anastrepha ludens (Diptera: Tephritidae) to the odor of fermented chapote fruit and to pheromones in laboratory experiments. Environ. Entomol. 1990, 19, 403–408. [Google Scholar] [CrossRef]
- Abraham, J.; Zhang, A.; Angeli, S.; Abubeker, S.; Michel, C.; Feng, Y.; Rodriguez-Saona, C. Behavioral and antennal responses of Drosophila suzukii (Diptera: Drosophilidae) to volatiles from fruit extracts. Environ. Entomol. 2015, 44, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Uchida, G.K.; Mackey, B.E.; Vargas, R.I.; Beardsley, J.W.; Hardy, D.E.; Goff, M.L.; Stark, J.D. Response of non target insects to methyl eugenol, cue-lure, trimedlure, and protein bait bucket traps on Kauai Island, Hawaii, USA. Proc. Hawaii Entomol. Soc. 2006, 38, 61–71. [Google Scholar]
- Leblanc, L.; Vargas, R.I.; Rubinoff, D. A comparison of non-target captures in BioLure and Liquid Protein Food Lures in Hawaii. Proc. Hawaii Entomol. Soc. 2010, 42, 15–22. [Google Scholar]
- De Bruyne, M.; Smart, R.; Zammit, E.; Warr, C.G. Functional and molecular evolution of olfactory neurons and receptors for aliphatic esters across the Drosophila genus. J. Comp. Physiol. 2010, 196, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Nigg, H.N.; Mallory, L.L.; Simpson, S.E.; Callaham, S.B.; Toth, J.P.; Fraser, S.; Klim, M.; Nagy, S.; Nation, J.L.; Attaway, J.A. Caribbean fruit fly, Anastrepha suspensa (Loew), attraction to host fruit and host kairomones. J. Chem. Ecol. 1994, 20, 727–743. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Park, K.C.; Baker, T.C. Identification of odors from overripe mango that attract vinegar flies, Drosophila melanogaster. J. Chem. Ecol. 2003, 29, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Casana-Giner, V.; Gandia-Balaguer, A.; Hernández-Alamós, M.M.; Mengod-Puerta, C.; Garrido-Vivas, A.; Primo-Millo, J.; Primo-Yúfera, E. Attractiveness of 79 compounds and mixtures to wild Ceratitis capitata (Diptera: Tephritidae) in field trials. J. Econ. Entomol. 2001, 94, 898–904. [Google Scholar] [CrossRef] [PubMed]
- De Zeeuw, J.; Stricek, R.; Stidsen, J. An advanced base-deactivated capillary column for analysis of volatile amines, ammonia, and alcohols. Am. Lab. 2011, 43, 37–40. [Google Scholar]
- Hussain, A.; Zhang, M.; Üçpunar, H.K.; Svensson, T.; Quillery, E.; Gompel, N.; Ignell, R.; Kadow, I.C.G. Ionotropic chemosensory receptors mediate the taste and smell of polyamines. PLoS Biol. 2016, 14, e1002454. [Google Scholar] [CrossRef] [PubMed]
- Delventhal, R.; Menuz, K.; Joseph, R.; Park, J.; Sun, J.S.; Carlson, J.R. The taste response to ammonia in Drosophila. Sci. Rep. 2017, 7, 43754. [Google Scholar] [CrossRef] [PubMed]
- Dekker, T.; Ibba, I.; Siju, K.; Stensmyr, M.C.; Hansson, B.S. Olfactory shifts parallel superspecialism for toxic fruit in Drosophila melanogaster Sibling, D. sechellia. Curr. Biol. 2006, 16, 101–109. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blend Components (General Blend) | Ratio (ng µL−1) | CAS | Kovats Index | B. dorsalis dorsablend | B. zonata zonablend | C. capitate capiblend | Z. cucurbitae cucublend |
|---|---|---|---|---|---|---|---|
| 3-Methyl-1-butanol | 35 | 123-51-3 | 763 | * | * | * | |
| Ethyl butanoate | 3 | 105-54-4 | 810 | * | * | * | |
| Butyl acetate | 62 | 123-86-4 | 821 | * | * | * | * |
| 2-Methylpyrazine | 4 | 109-08-0 | 831 | * | * | * | |
| 3-Methylbutyl acetate | 94 | 123-92-2 | 877 | * | * | * | * |
| 2,5-Dimethylpyrazine | 7 | 123-32-0 | 911 | * | * | * | |
| Benzaldehyde | 1 | 100-52-7 | 963 | * | |||
| Ethyl hexanoate | 112 | 123-66-0 | 993 | * | * | * | * |
| 2,6-Dimethylpyrazine | 7 | 108-50-9 | 912 | * | * | ||
| Ethyl (E)-2-methylbut-2-enoate | 1 | 5837-78-5 | 937 | * | |||
| β-Myrcene | 41 | 123-35-3 | 987 | * | * | ||
| 2-Methoxyphenol | 6 | 90-05-1 | 1082 | * | |||
| 1-Phenylethyl acetate | 249 | 93-92-5 | 1189 | * | * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biasazin, T.D.; Chernet, H.T.; Herrera, S.L.; Bengtsson, M.; Karlsson, M.F.; Lemmen-Lechelt, J.K.; Dekker, T. Detection of Volatile Constituents from Food Lures by Tephritid Fruit Flies. Insects 2018, 9, 119. https://doi.org/10.3390/insects9030119
Biasazin TD, Chernet HT, Herrera SL, Bengtsson M, Karlsson MF, Lemmen-Lechelt JK, Dekker T. Detection of Volatile Constituents from Food Lures by Tephritid Fruit Flies. Insects. 2018; 9(3):119. https://doi.org/10.3390/insects9030119
Chicago/Turabian StyleBiasazin, Tibebe Dejene, Haimanot Teklemariam Chernet, Sebastian Larsson Herrera, Marie Bengtsson, Miriam Frida Karlsson, Joelle Kristin Lemmen-Lechelt, and Teun Dekker. 2018. "Detection of Volatile Constituents from Food Lures by Tephritid Fruit Flies" Insects 9, no. 3: 119. https://doi.org/10.3390/insects9030119