
Isolating Spermathecae and Determining Mating Status of Drosophila suzukii: A Protocol for Tissue Dissection and Its Applications


Abstract
:1. Introduction
2. Materials and Methods
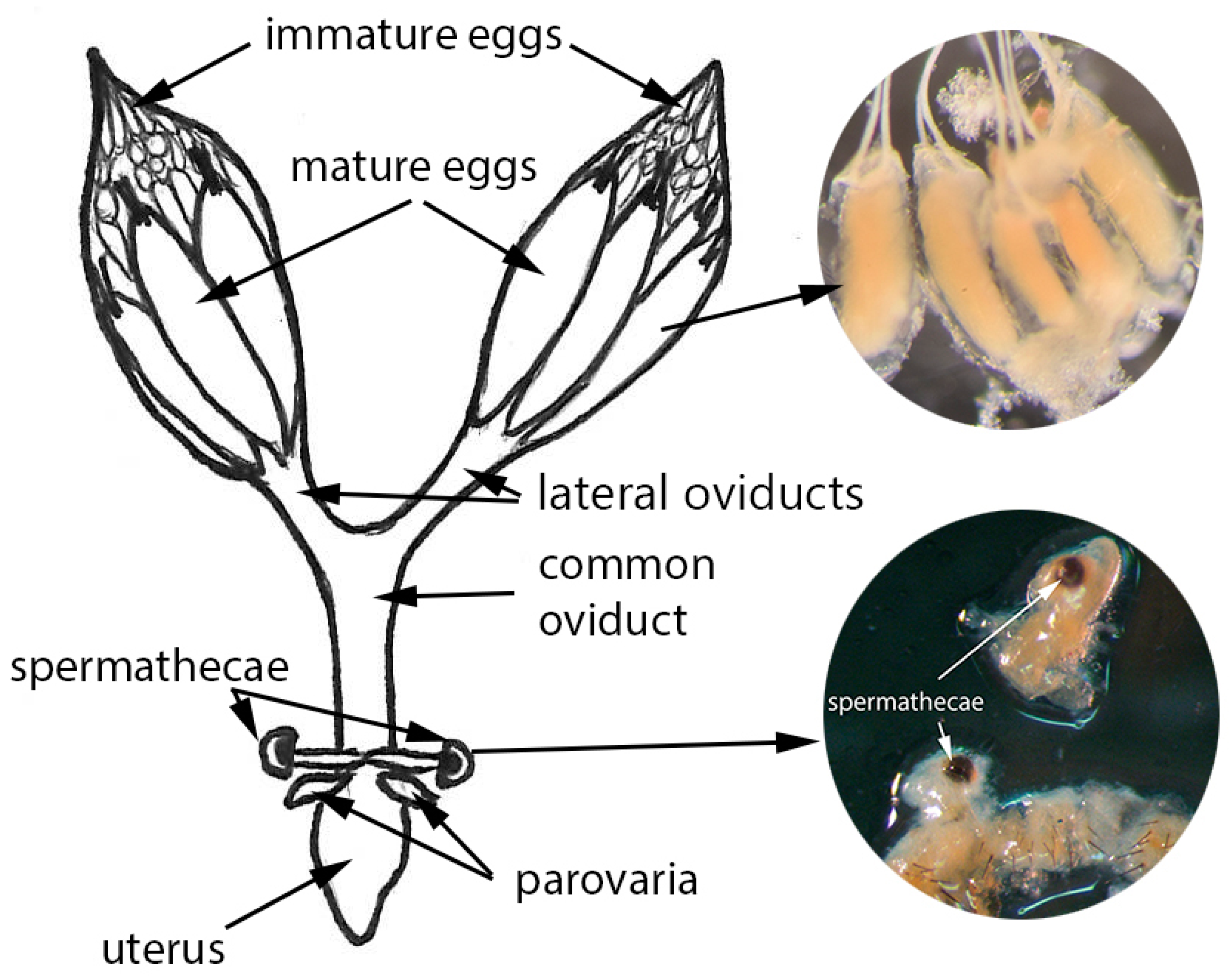
2.1. Study Species
2.2. Protocol Development
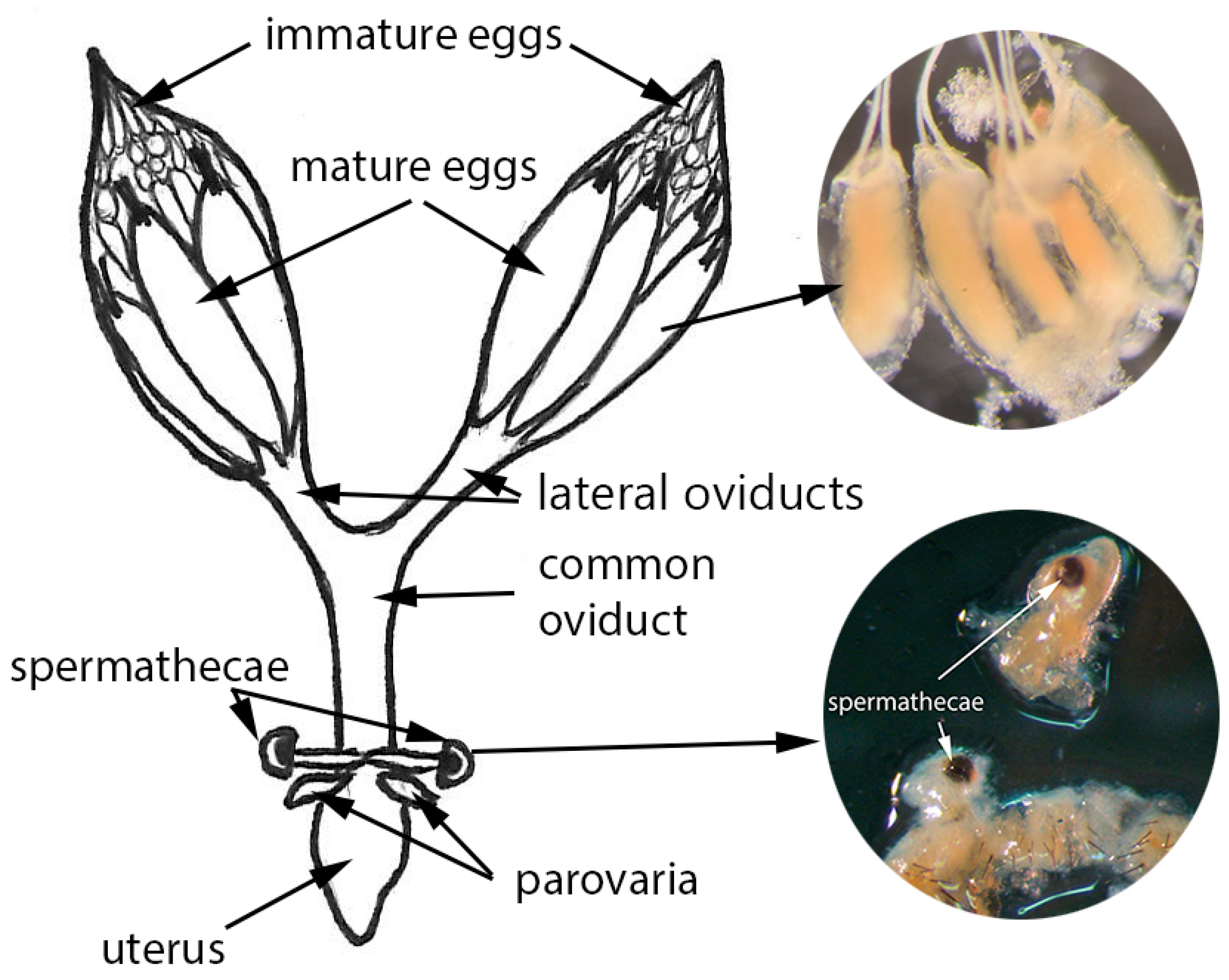
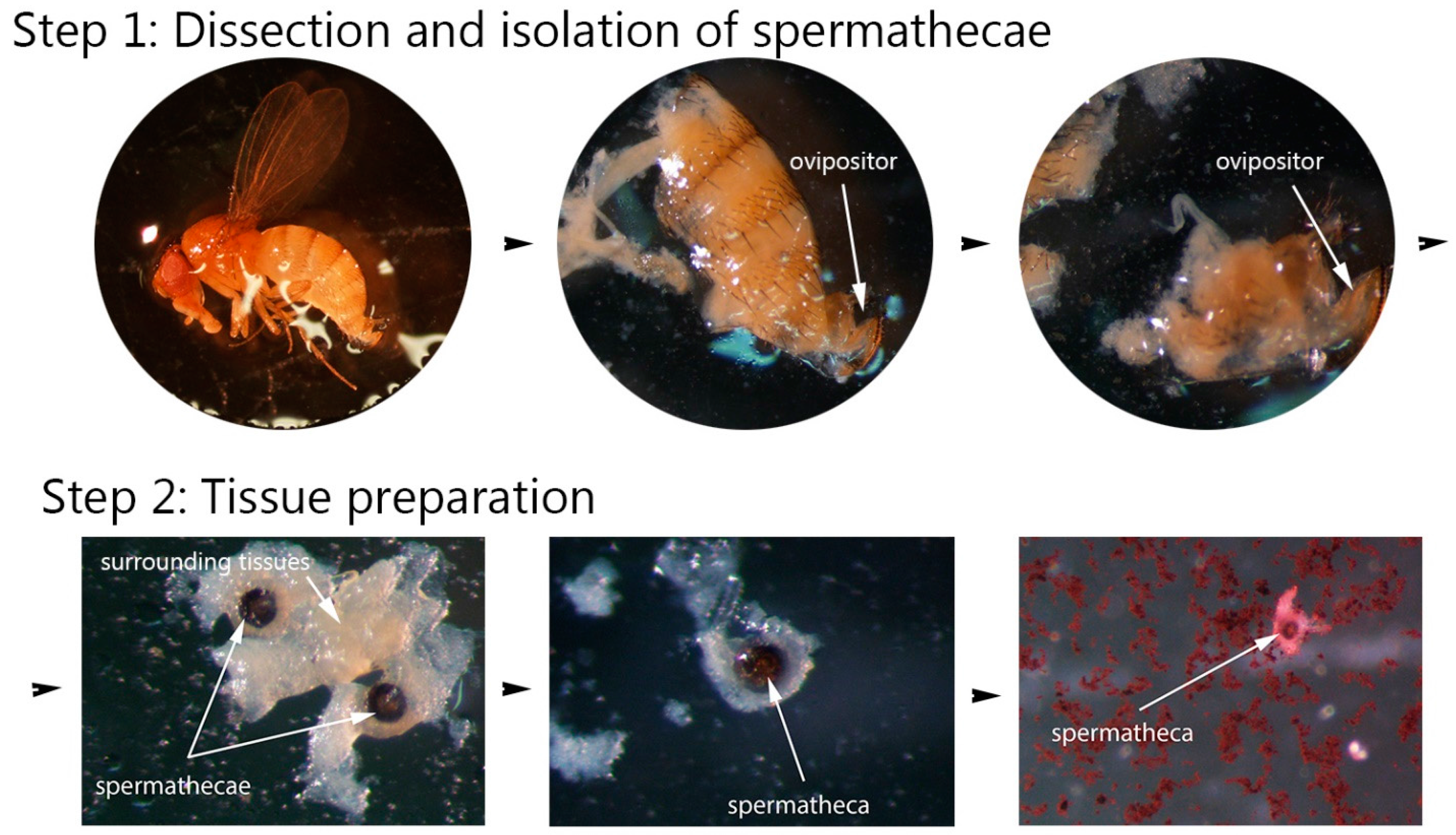
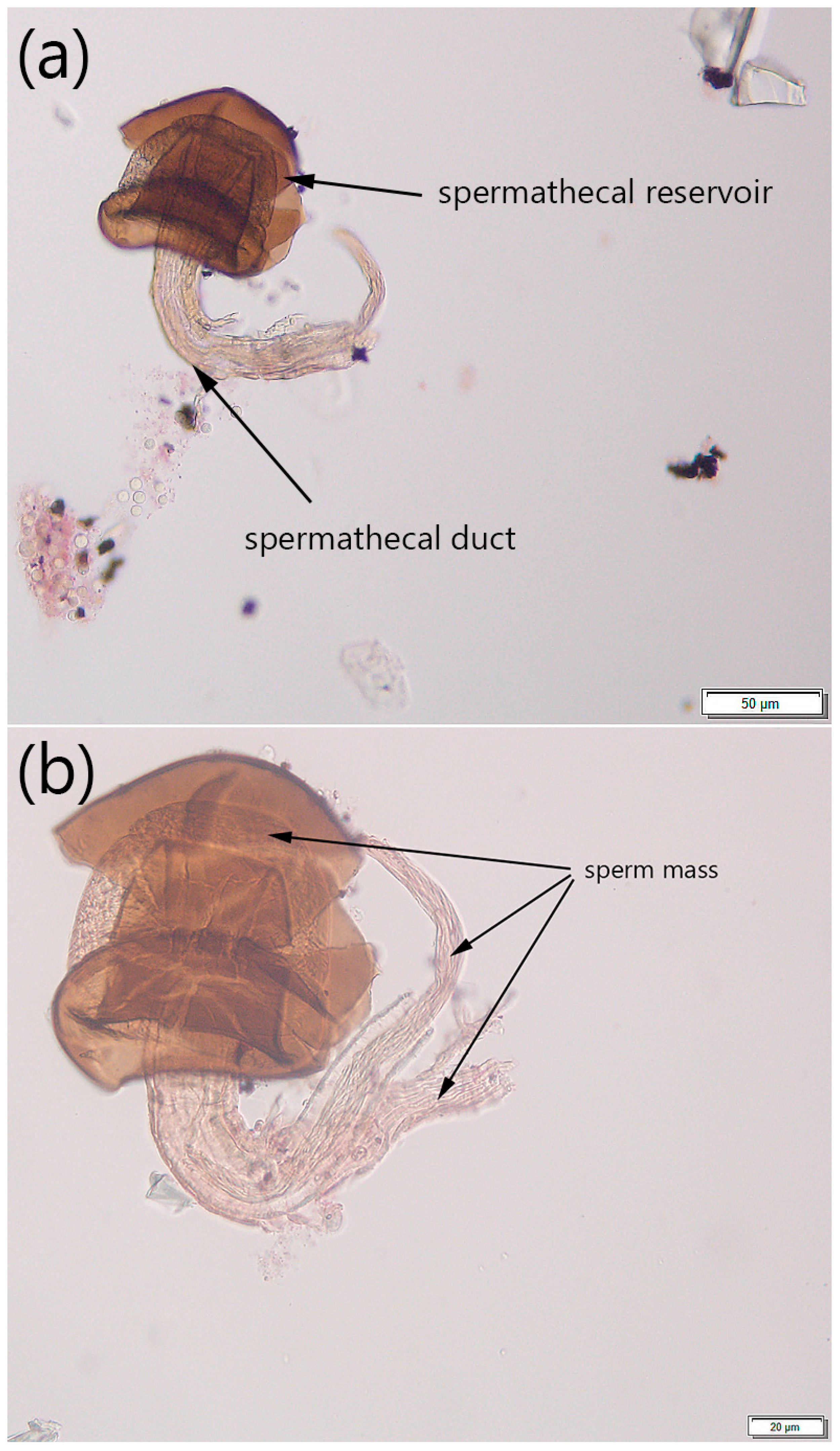
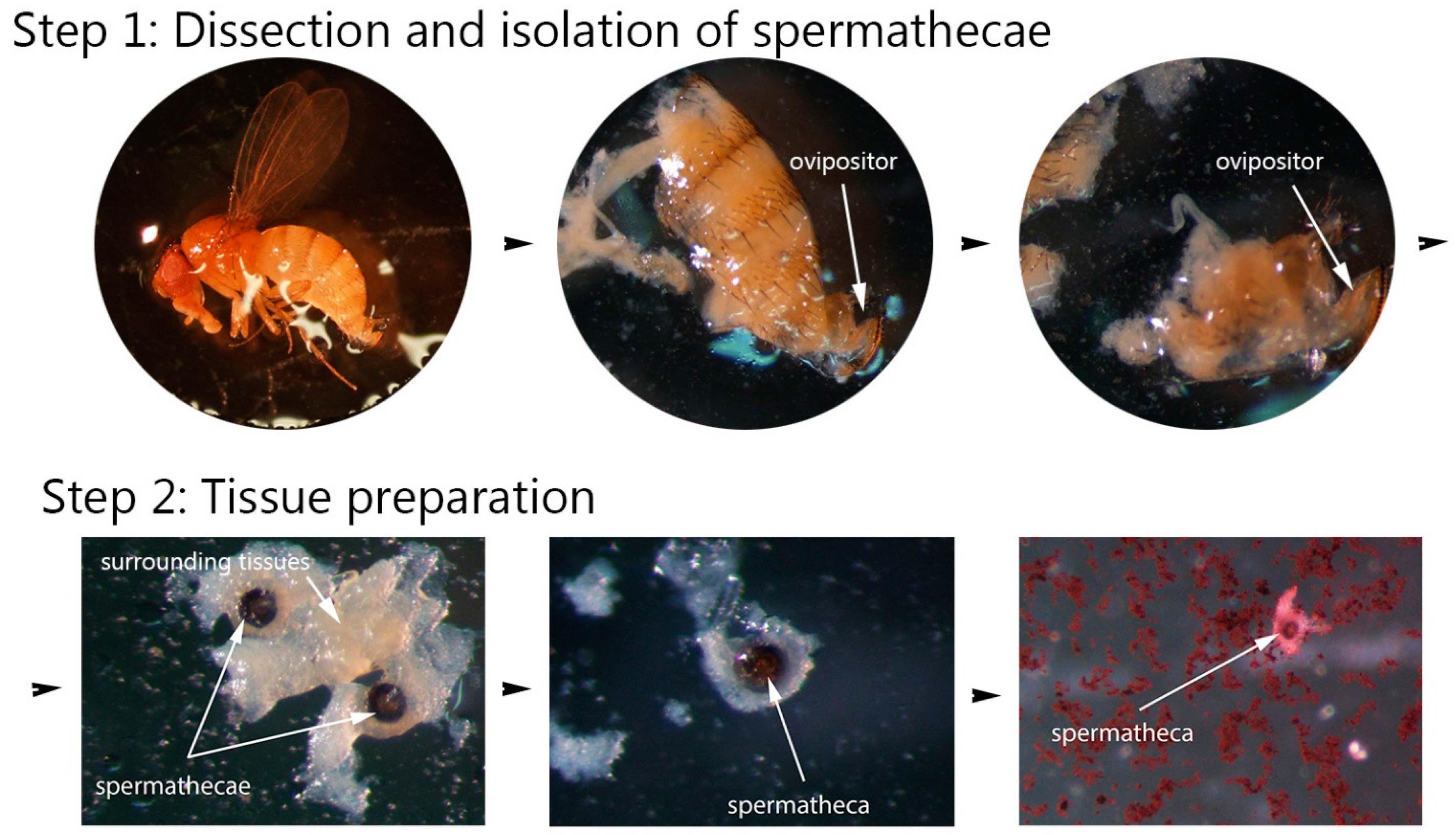
2.2.1. Step 1: Dissection and Isolation of Spermathecae
2.2.2. Step 2: Tissue Preparation
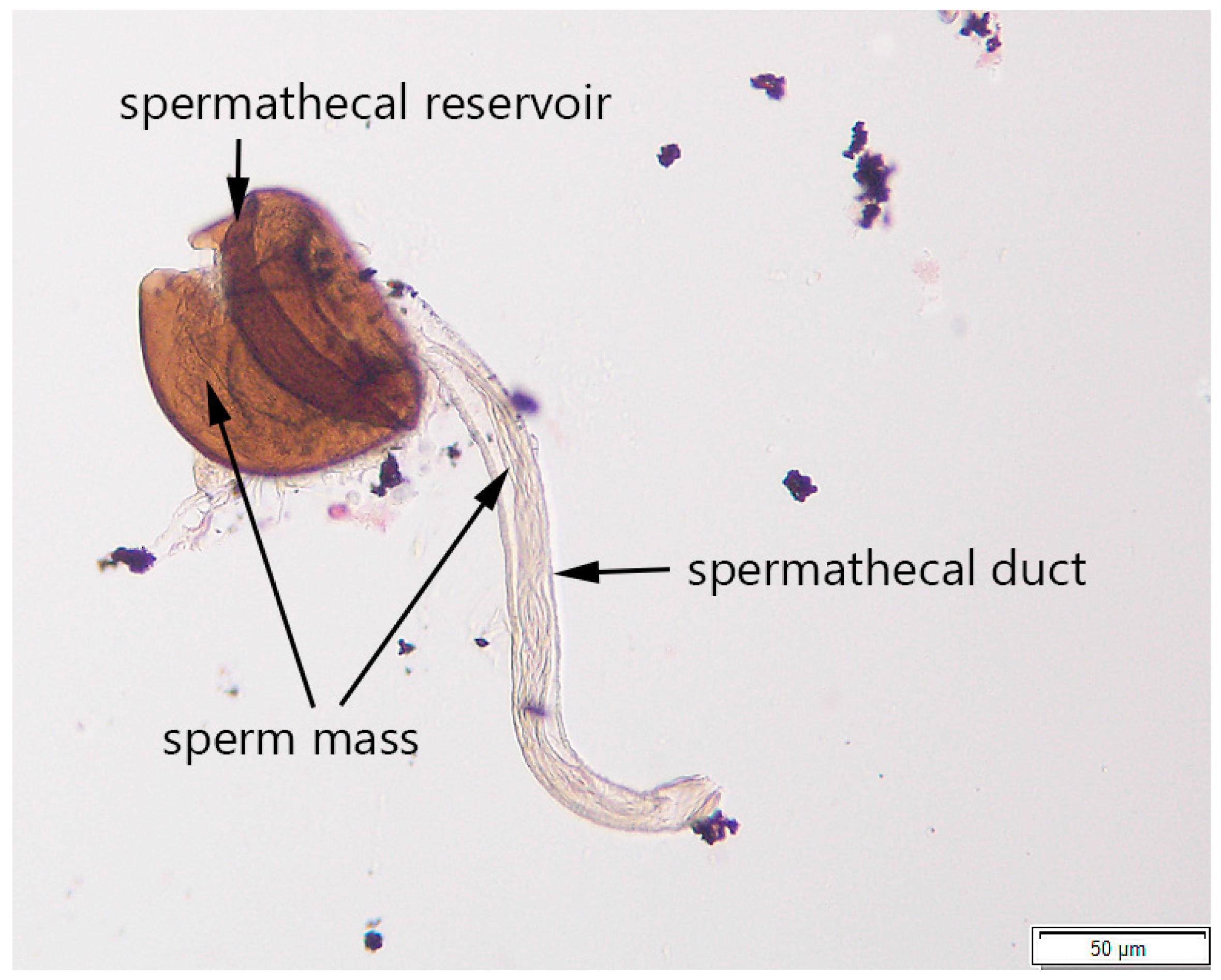
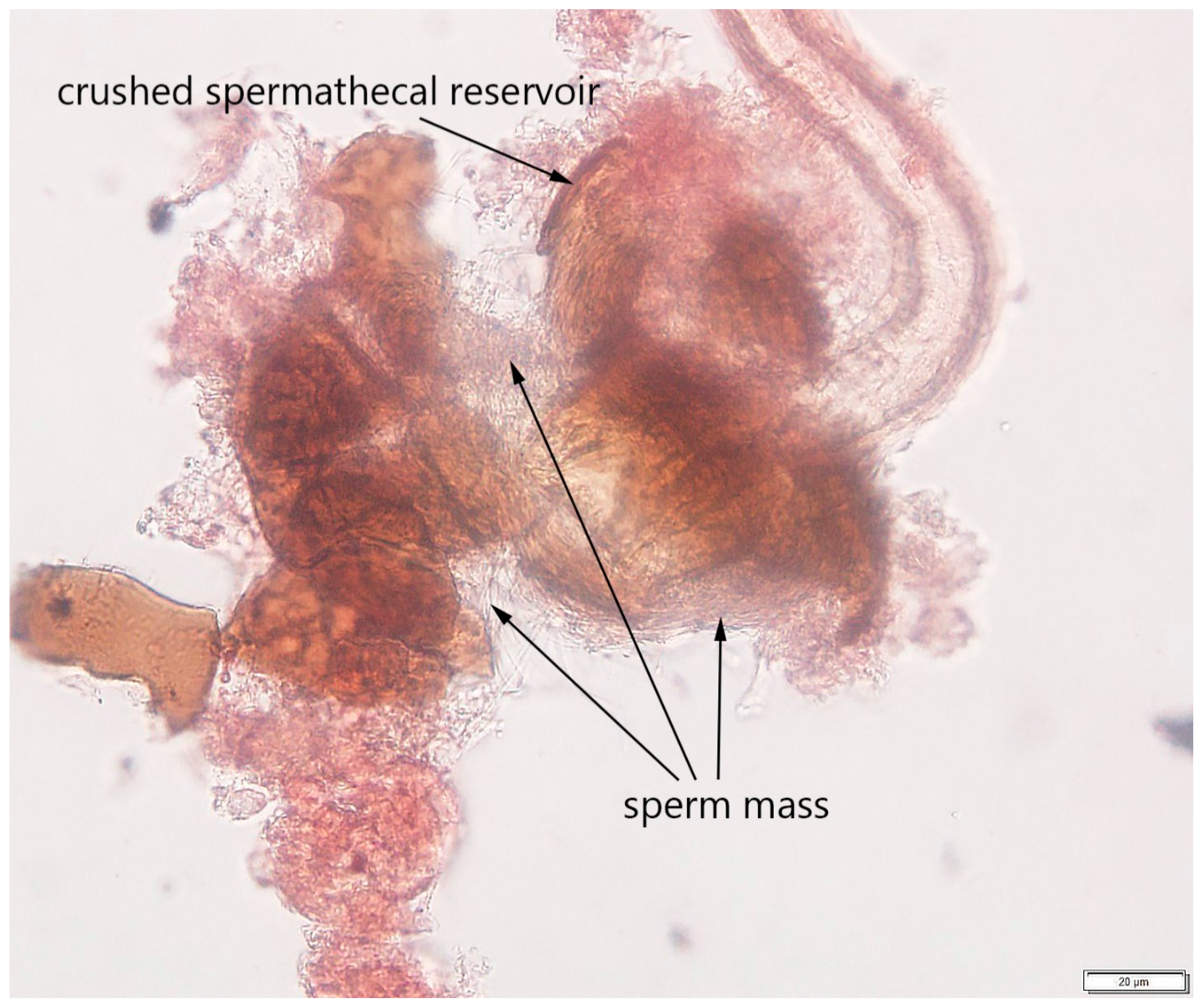
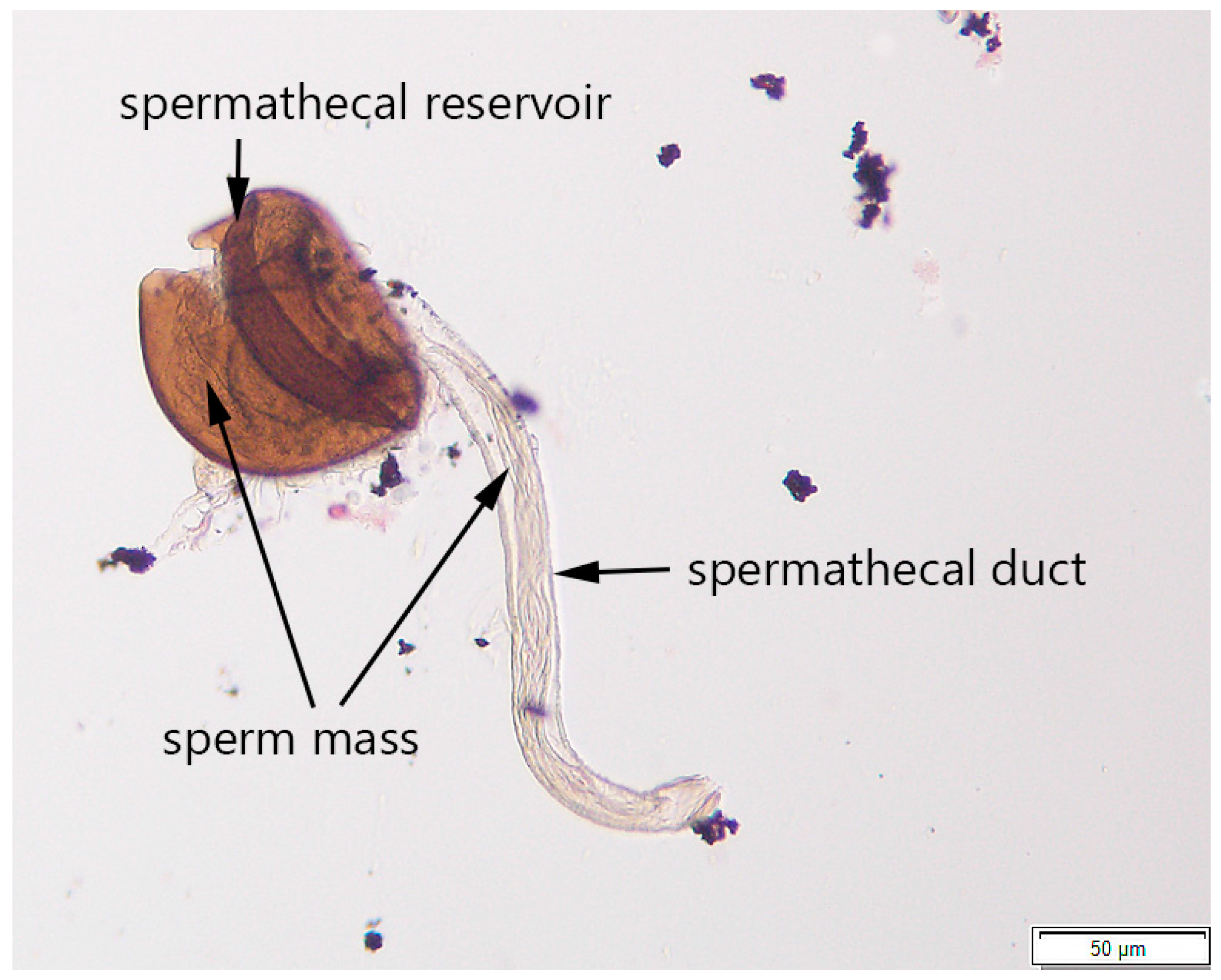
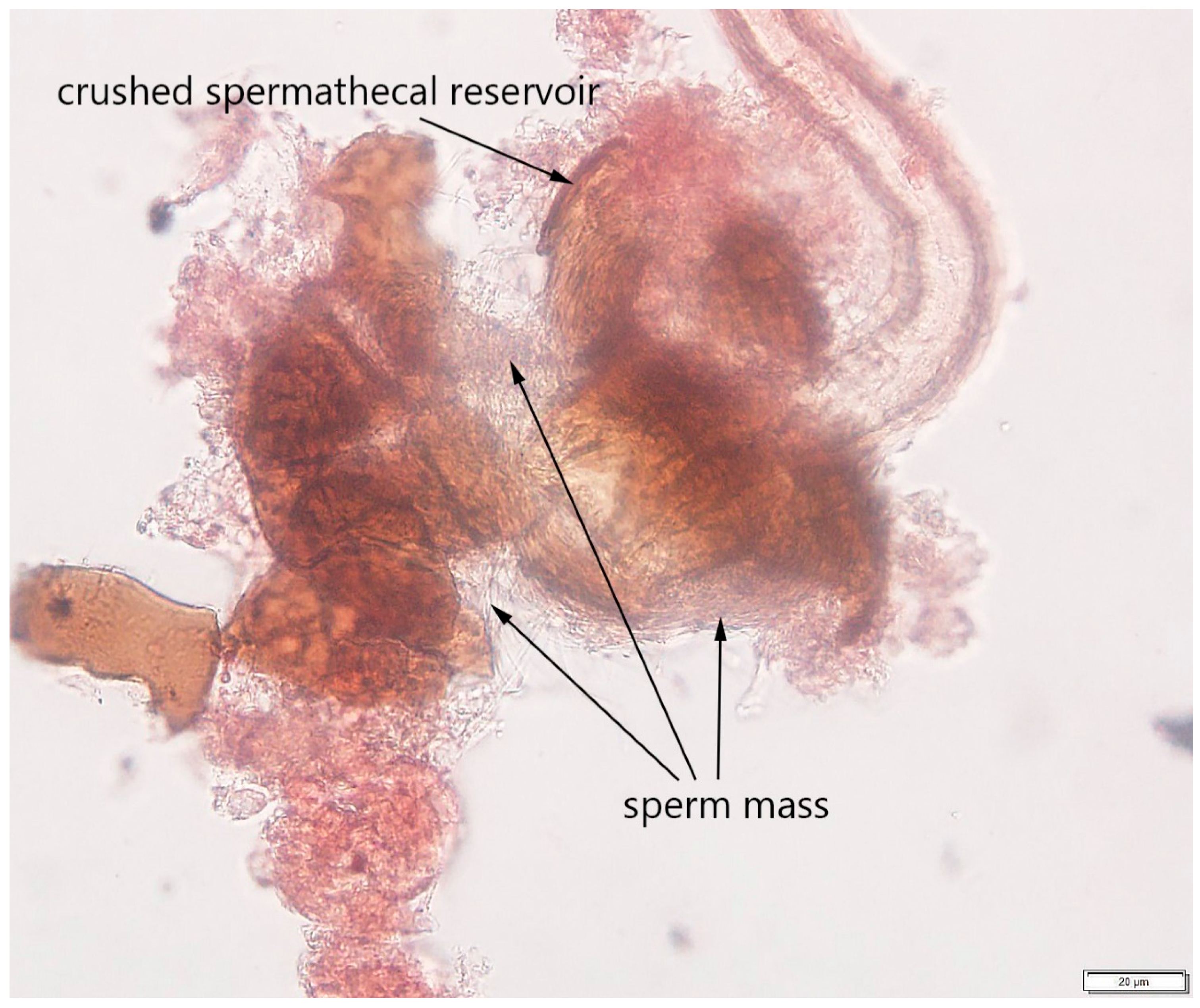
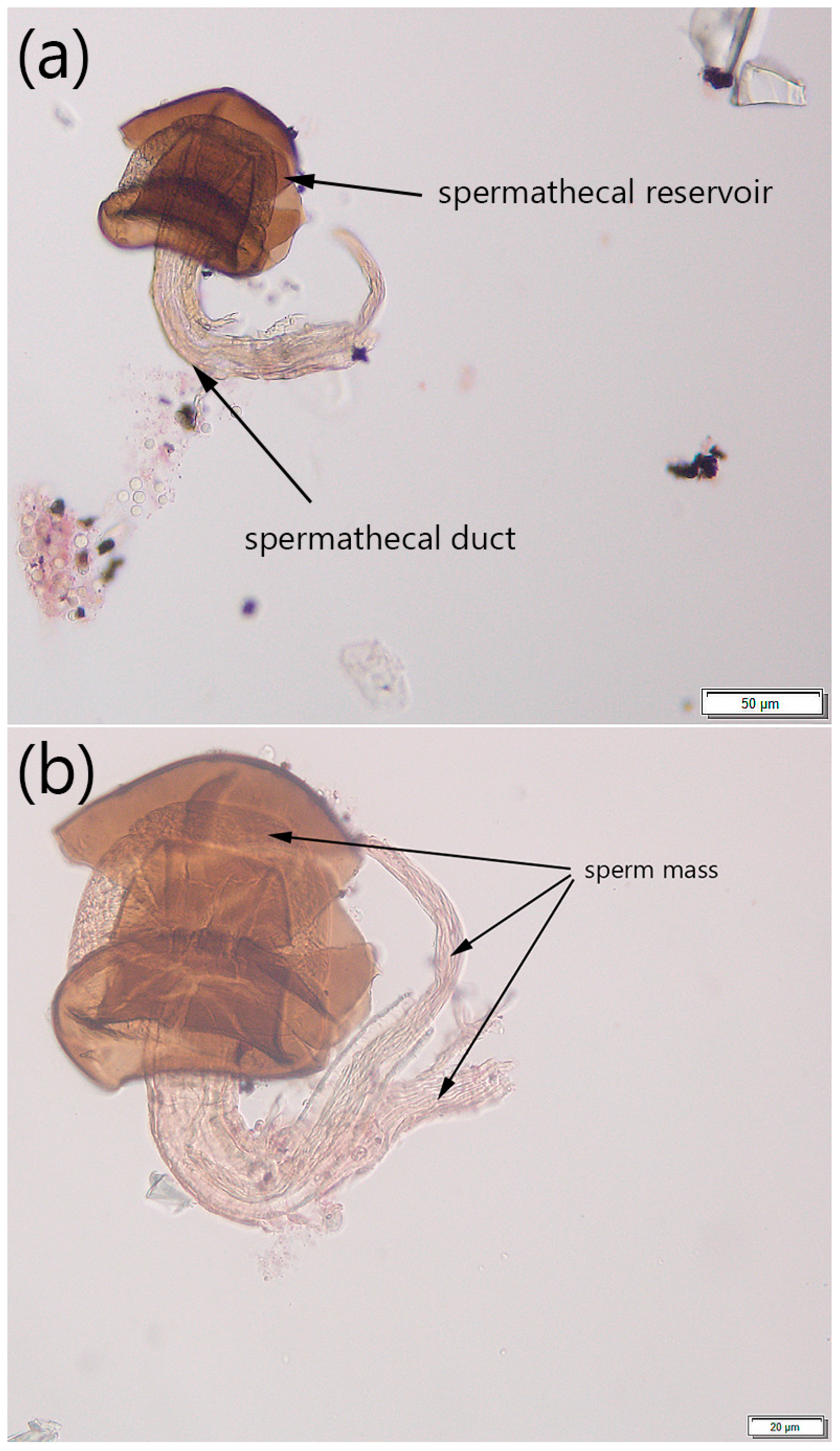
2.2.3. Step 3: Determining Mating Status
2.3. Testing the Protocol
3. Results
3.1. Protocol Development
3.2. Testing the Protocol
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Landolt, P.J.; Adams, T.; Rogg, H. Trapping spotted wing drosophila, Drosophila suzukii (Matsumura) (Diptera: Drosophilidae), with combinations of vinegar and wine, and acetic acid and ethanol. J. Appl. Entomol. 2012, 136, 148–154. [Google Scholar] [CrossRef]
- Lee, J.C.; Bruck, D.J.; Curry, H.; Edwards, D.; Haviland, D.R.; Van Steenwyk, R.A.; Yorgey, B.M. The susceptibility of small fruits and cherries to the spotted-wing drosophila, Drosophila suzukii. Pest Manag. Sci. 2011, 67, 1358–1367. [Google Scholar] [CrossRef] [PubMed]
- Burrack, H.J.; Fernandez, G.E.; Spivey, T.; Kraus, D.A. Variation in selection and utilization of host crops in the field and laboratory by Drosophila suzukii Matsumara (Diptera: Drosophilidae), an invasive frugivore. Pest Manag. Sci. 2013, 69, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Gilbert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Abraham, J.; Zhang, A.; Angeli, S.; Abubeker, S.; Michel, C.; Feng, Y.; Rodriguez-Saona, C. Behavioral and antennal responses of Drosophila suzukii (Diptera: Drosophilidae) to volatiles from fruit extracts. Environ. Entomol. 2015, 44, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Dalton, D.T.; Walton, V.M.; Shearer, P.W.; Walsh, D.B.; Caprile, J.; Isaacs, R. Laboratory survival of Drosophila suzukii under simulated winter conditions of the Pacific Northwest and seasonal field trapping in five primary regions of small and stone fruit production in the United States. Pest Manag. Sci. 2011, 67, 1368–1374. [Google Scholar] [CrossRef] [PubMed]
- Burrack, H.J.; Smith, J.P.; Pfeiffer, D.G.; Koeher, G.; Laforest, J. Using volunteer-based networks to track Drosophila suzukii (Diptera: Drosophilidae) an invasive pest of fruit crops. J. Integr. Pest Manag. 2012, 3, B1–B5. [Google Scholar] [CrossRef]
- Hampton, E.; Koski, C.; Barsoian, O.; Faubert, H.; Cowles, R.S.; Alm, S.R. Use of early ripening cultivars to avoid infestation and mass trapping to manage Drosophila suzukii (Diptera: Drosophilidae) in Vaccinium corymbosum (Ericales: Ericaceae). J. Econ. Entomol. 2014, 107, 1849–1857. [Google Scholar] [CrossRef] [PubMed]
- Joshi, N.K.; Butler, B.; Demchak, K.; Biddinger, D. Seasonal occurrence of spotted wing drosophila in various small fruits and berries in Pennsylvania and Maryland. J. Appl. Entomol. 2016. [Google Scholar] [CrossRef]
- Pelton, E.; Gratton, C.; Isaacs, R.; Van Timmeren, S.; Blanton, A.; Guédot, C. Earlier activity of Drosophila suzukii in high woodland landscapes but relative abundance is unaffected. J. Pest Sci. 2016, 89, 725–733. [Google Scholar] [CrossRef]
- Lin, Q.C.; Zhai, Y.F.; Zhou, C.G.; Li, L.L.; Zhuang, Q.Y.; Zhang, X.Y.; Zalom, F.G.; Yu, Y. Behavioral rhythms of Drosophila suzukii and Drosophila melanogaster. Fla. Entomol. 2014, 97, 1424–1433. [Google Scholar] [CrossRef]
- Dekker, T.; Revadi, S.; Mansourian, S.; Ramasamy, S.; Lebreton, S.; Becher, P.G.; Angeli, S.; Rota-Stabelli, O.; Anfora, G. Loss of Drosophila pheromone reverses its role in sexual communication in Drosophila suzukii. Proc. R. Soc. B 2015. [Google Scholar] [CrossRef] [PubMed]
- Revadi, S.; Lebreton, S.; Witzgall, P.; Anfora, G.; Dekker, T.; Becher, P.G. Sexual behavior of Drosophila suzukii. Insects 2015, 6, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Kapelnikov, A.; Rivlin, P.K.; Hoy, R.R.; Heifetz, Y. Tissue remodeling: A mating-induced differentiation program for the Drosophila oviduct. BMC Dev. Biol. 2008. [Google Scholar] [CrossRef] [PubMed]
- Long, T.A.F.; Pischedda, A.; Nichols, R.V.; Rice, W.R. The timing of mating influences reproductive success in Drosophila melanogaster: Implications for sexual conflict. J. Evol. Biol. 2010, 23, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Chapman, T.; Takahisa, M.; Smith, H.K.; Partridge, L. Interactions of mating, egg production and death rates in females of the Mediterranean fruitfly, Ceratitis capitata. Proc. R. Soc. B 1998, 265, 1879–1894. [Google Scholar] [CrossRef] [PubMed]
- Qazi, M.C.B.; Heifetz, Y.; Wolfner, M.F. The developments between gametogenesis and fertilization: Ovulation and female sperm storage in Drosophila melanogaster. Dev. Biol. 2003, 256, 195–211. [Google Scholar] [CrossRef]
- Allen, A.K.; Spradling, A.C. The Sf1-related nuclear hormone receptor Hr39 regulates Drosophila female reproductive tract development and function. Development 2008, 135, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Pitnick, S.; Markow, T.; Spicer, G.S. Evolution of multiple kinds of female sperm-storage organs in Drosophila. Evolution 1999, 53, 1804–1822. [Google Scholar] [CrossRef]
- Bangham, J.; Chapman, T.; Smith, H.K.; Partridge, L. Influence of female reproductive anatomy on the outcome of sperm competition in Drosophila melanogaster. Proc. R. Soc. B 2003, 270, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Mayhew, M.L.; Merritt, D.J. The morphogenesis of spermathecae and spermathecal glands in Drosophila melanogaster. Arthropod Struct. Dev. 2013, 42, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.L.; Smith, D.S. Fine structural organization of the spermatheca in the cockroach, Periplaneta americana. Tissue Cell 1969, 1, 295–324. [Google Scholar] [CrossRef]
- Lay, M.; Zissler, D.; Hartmann, R. Ultrastructural and functional aspects of the spermatheca of the African Migratory Locust Locusta migratoria migratorioides (Reiche and Fairmaire) (Orthoptera: Acrididae). Int. J. Insect Morphol. Embryol. 1999, 28, 349–361. [Google Scholar] [CrossRef]
- Happ, G.M.; Happ, C.M. Fine structure and histochemistry of the spermathecal gland in the mealworm beetle, Tenebrio molitor. Tissue Cell 1970, 2, 443–466. [Google Scholar] [CrossRef]
- Tombes, A.S.; Roppel, R.M. Scanning electron microscopy of the spermatheca in Sitophilus granarius (L.). Tissue Cell 1971, 3, 551–556. [Google Scholar] [CrossRef]
- Tombes, A.S.; Roppel, R.M. Ultrastructure of the spermatheca of the granary weevil, Sitophilus granarius (L.) (Coleoptera: Curculionidae). Int. J. Insect Morphol. Embryol. 1972, 1, 141–152. [Google Scholar] [CrossRef]
- Villavaso, E.J. The role of the spermathecal gland of the boll weevil, Anthonomus grandis. J. Insect Physiol. 1975, 21, 1457–1462. [Google Scholar] [CrossRef]
- Pendergrast, J.G. Studies on the reproductive organs of the Heteroptera with a consideration of their bearing on classification. Ecol. Entomol. 1957, 109, 1–63. [Google Scholar] [CrossRef]
- Huebner, E. Spermathecal ultrastructure of the insect Rhodnius prolixus Stal. J. Morphol. 1980, 166, 1–25. [Google Scholar] [CrossRef]
- Gschwentner, R.; Tadler, A. Functional anatomy of the spermatheca and its duct in the seed bug Lygaeus simulans (Heteroptera: Lygaeidae). Eur. J. Entomol. 2000, 97, 305–312. [Google Scholar] [CrossRef]
- Stacconi, M.V.R.; Romani, R. Ultrastructural and functional aspects of the spermatheca in the american harlequin bug, Murgantia histrionica (Hemiptera: Pentatomidae). Neotrop. Entomol. 2011, 40, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Pabalan, N.; Davey, K.G.; Packer, L. Comparative morphology of spermathecae in solitary and primitively eusocial bees (Hymenoptera; Apoidea). Can. J. Zool. 1996, 74, 802–808. [Google Scholar] [CrossRef]
- Gotoh, A.; Billen, J.; Hashim, R.; Ito, F. Comparison of spermatheca morphology between reproductive and non-reproductive females in social wasps. Arthropod Struct. Dev. 2008, 37, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Clements, A.N.; Potter, S.A. The fine structure of the spermathecae and their ducts in the mosquito Aedes aegypti. J. Insect Physiol. 1967, 13, 1825–1836. [Google Scholar] [CrossRef]
- Filosi, M.; Perotti, M.E. Fine structure of the spermatheca of Drosophila melanogaster Meig. J. Submicrosc. Cytol. Pathol. 1975, 7, 259–270. [Google Scholar]
- Kokwaro, E.D.; Okot-Kotber, B.M.; Odhiambo, T.R.; Murithi, J.K. Ultrastructural and histochemical study of the spermatheca of the tsetse Glossina morsitans morsitans Westwood. Insect Sci. Appl. 1981, 2, 135–143. [Google Scholar] [CrossRef]
- Fritz, A.H.; Turner, F.R. A light and electron microscopical study of the spermathecae and ventral receptacle of Anastrepha suspensa (Diptera: Tephritidae) and implications in female influence of sperm storage. Arthropod Struct. Dev. 2002, 30, 293–313. [Google Scholar] [CrossRef]
- Presgraves, D.C.; Baker, R.H.; Wilkinson, G.S. Coevolution of sperm and female reproductive tract morphology in stalk–eyed flies. Proc. R. Soc. B 1999, 266, 1041–1047. [Google Scholar] [CrossRef]
- Bartolucci, A.F.; Vera, M.T.; Yusef, V.; Oviedo, A. Morphological characterization of the reproductive system of irradiated Anastrepha fraterculus. In Proceedings of the 7th International Symposium on Fruit Flies of Economic Importance, Salvador, Brazil, 10–15 September 2006; pp. 45–52.
- Rossi-Stacconi, M.V.; Kaur, R.; Mazzoni, V.; Ometto, L.; Grassi, A.; Gottardello, A.; Rota-Stabelli, O.; Anfora, G. Multiple lines of evidence for reproductive winter diapause in the invasive pest Drosophila suzukii: Useful clues for control strategies. J. Pest Sci. 2016, 89, 689–700. [Google Scholar] [CrossRef]
- Burrack, H.J.; Asplen, M.; Bahder, L.; Collins, J.; Drummond, F.A.; Guédot, C.; Isaacs, R.; Johnson, D.; Blanton, A.; Lee, J.C.; et al. Multistate comparison of attractants for monitoring Drosophila suzukii (Diptera: Drosophilidae) in blueberries and caneberries. Environ. Entomol. 2015, 44, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Polak, M.; Wolf, L.L.; Starmer, W.T.; Barker, J.S.F. Function of the mating plug in Drosophila hibisci Bock. Behav. Ecol. Sociobiol. 2001, 49, 196–205. [Google Scholar] [CrossRef]
- O’Donnell, M.J.; Rheault, M.R.; Davies, S.A.; Rosay, P.; Harvey, B.J.; Maddrell, S.H.; Kaiser, K.; Dow, J.A. Hormonally controlled chloride movement across Drosophila tubules is via ion channels in stellate cells. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1998, 274, R1039–R1049. [Google Scholar]
- Budnik, V.; Gorczyca, M.; Prokop, A. Selected methods for the anatomical study of Drosophila embryonic and larval neuromuscular junctions. Int. Rev. Neurobiol. 2006, 75, 323–365. [Google Scholar] [PubMed]
- Shearer, P.W.; West, J.D.; Walton, V.M.; Brown, P.H.; Svetec, N.; Chiu, J.C. Seasonal cues induce phenotypic plasticity of Drosophila suzukii to enhance winter survival. BMC Ecol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Stephens, A.R.; Asplen, M.K.; Hutchison, W.D.; Venette, R.C. Cold hardiness of winter-acclimated Drosophila suzukii (Diptera: Drosophilidae) adults. Environ. Entomol. 2015, 44, 1619–1626. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Preservation Time (Days) | Mating Status 1 | |
|---|---|---|
| Virgin | Mated | |
| 565 | 0 | 5 |
| 572 | 0 | 4 |
| 575 | 0 | 5 |
| 579 | 2 | 2 |
| 579 | 0 | 1 |
| 582 | 0 | 3 |
| 589 | 0 | 3 |
| 590 | 1 | 0 |
| 593 | 0 | 1 |
| 603 | 0 | 5 |
| 603 | ? | ? |
| 645 | 1 | 0 |
| 667 | 0 | 5 |
| 691 | 0 | 5 |
| Parameters | Mating Status | Total Number of Flies | |
|---|---|---|---|
| Virgin | Mated | ||
| Number of flies per treatment | 10 | 10 | 20 |
| Number of correctly identified flies | 60% | 70% | 65% |
| Data comparisons (Correctly identified virgin vs. mated flies) | Kruskal-Wallis test: H = 0.2, df = 1, p > 0.05 | ||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avanesyan, A.; Jaffe, B.D.; Guédot, C. Isolating Spermathecae and Determining Mating Status of Drosophila suzukii: A Protocol for Tissue Dissection and Its Applications. Insects 2017, 8, 32. https://doi.org/10.3390/insects8010032
Avanesyan A, Jaffe BD, Guédot C. Isolating Spermathecae and Determining Mating Status of Drosophila suzukii: A Protocol for Tissue Dissection and Its Applications. Insects. 2017; 8(1):32. https://doi.org/10.3390/insects8010032
Chicago/Turabian StyleAvanesyan, Alina, Benjamin D. Jaffe, and Christelle Guédot. 2017. "Isolating Spermathecae and Determining Mating Status of Drosophila suzukii: A Protocol for Tissue Dissection and Its Applications" Insects 8, no. 1: 32. https://doi.org/10.3390/insects8010032