Contrasting Effects of Histone Deacetylase Inhibitors on Reward and Aversive Olfactory Memories in the Honey Bee


Abstract
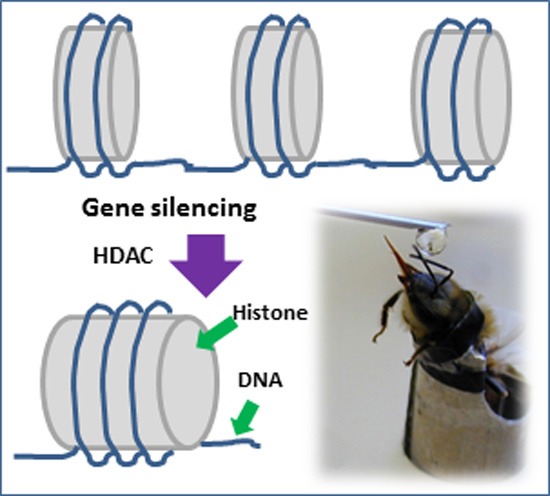
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Experimental Section
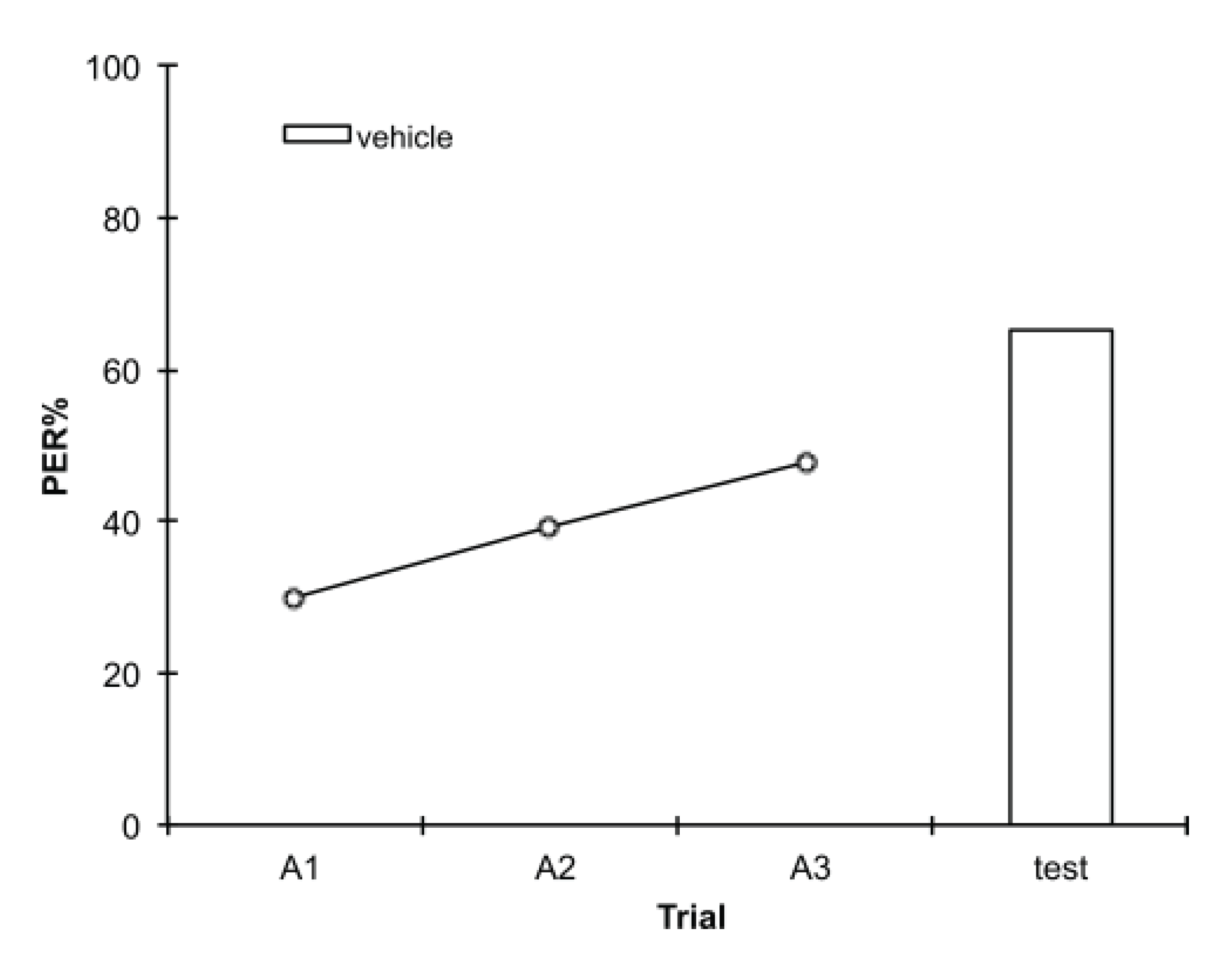
2.1. Olfactory Conditioning Assays
2.2. Sucrose Sensitivity Assays
2.3. Locomotor Activity Assays
2.4. Statistical Analyses
3. Results
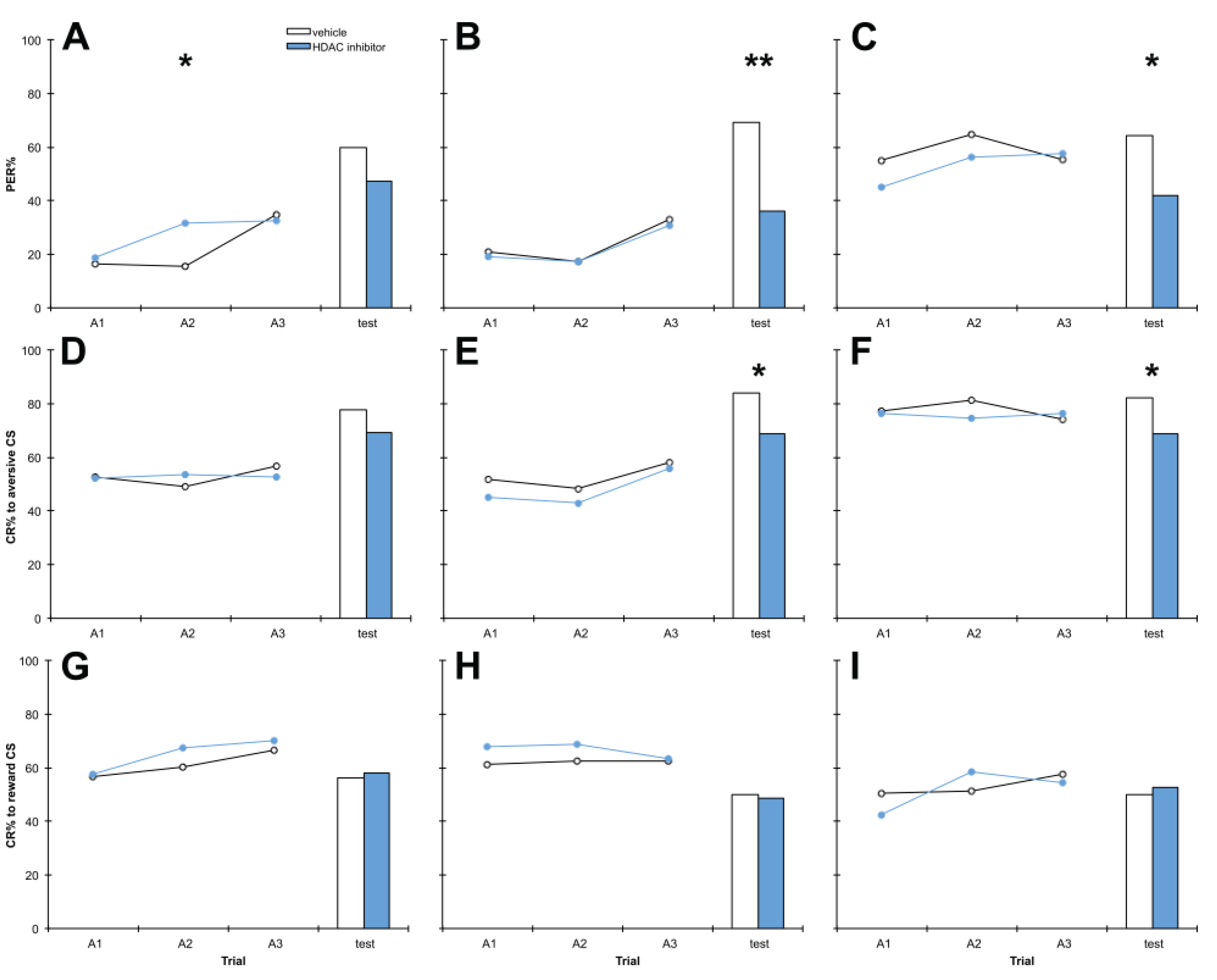
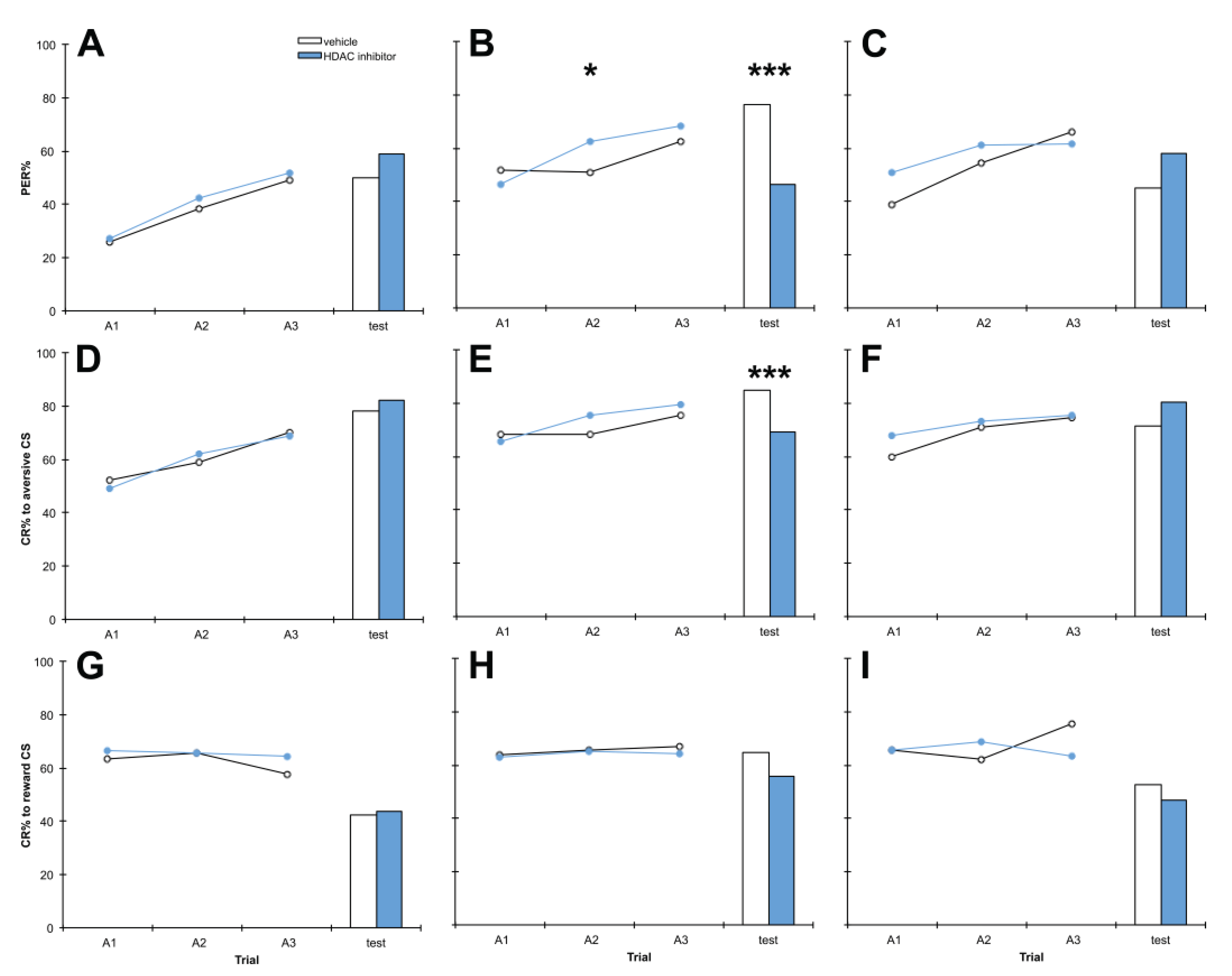
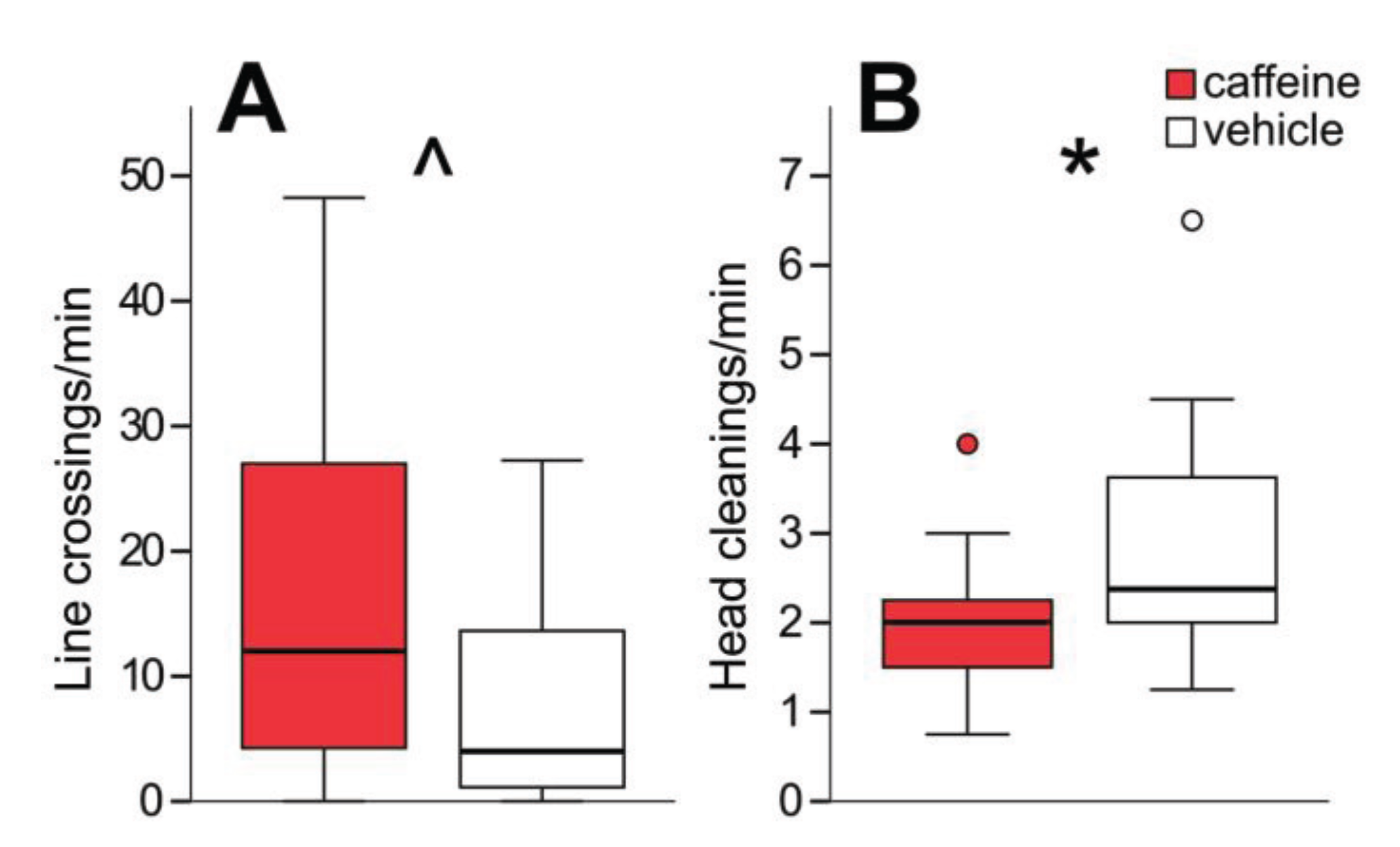
3.1. C8 and PB Impair Aversive Memory but not Reward Memory in a Dose-Dependent Manner

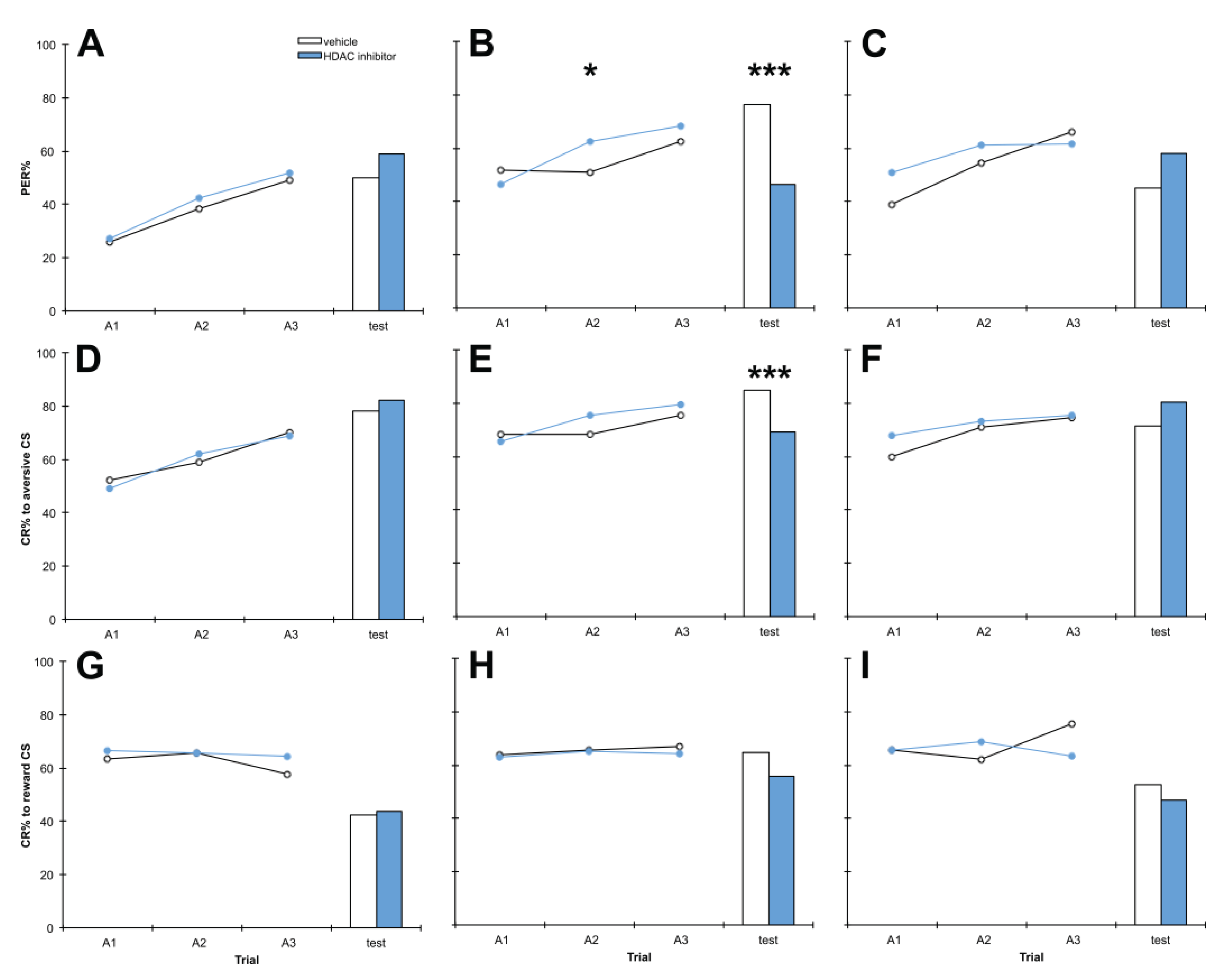
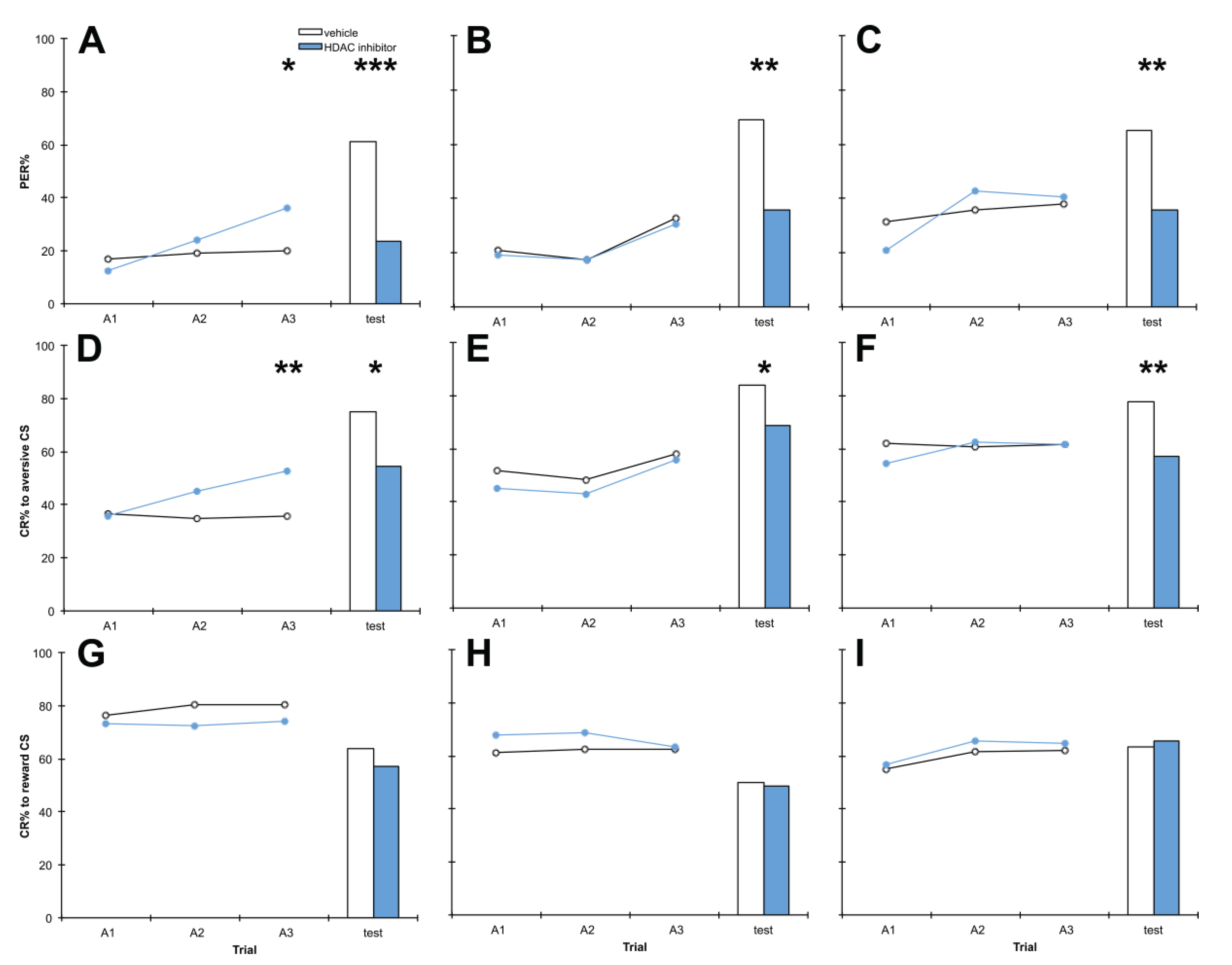
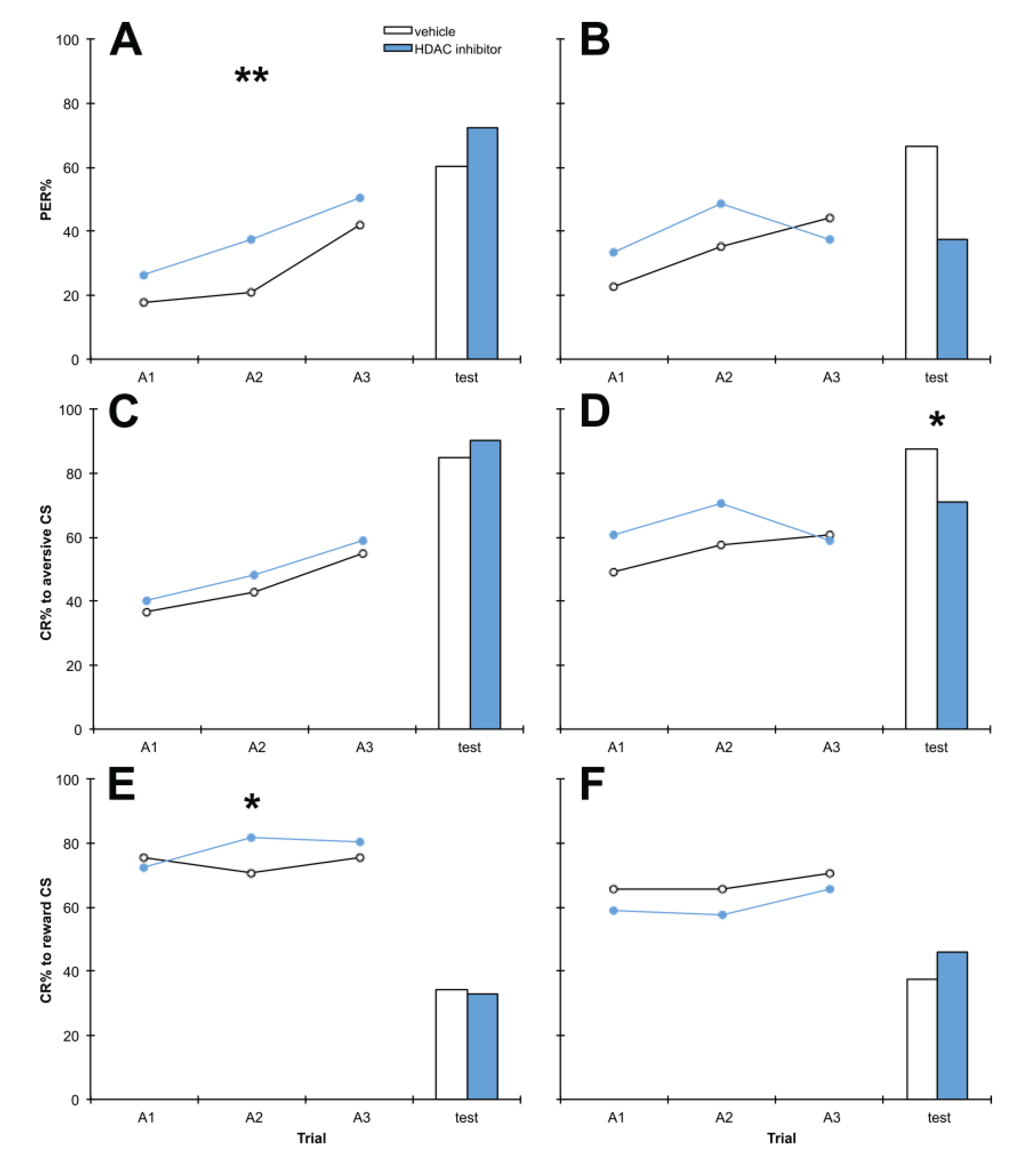
3.2. Treatment with C8 1 h before Training, 1 h after Training or 1 h before Testing Impairs Aversive Memory but Has no Effect on Reward Memory
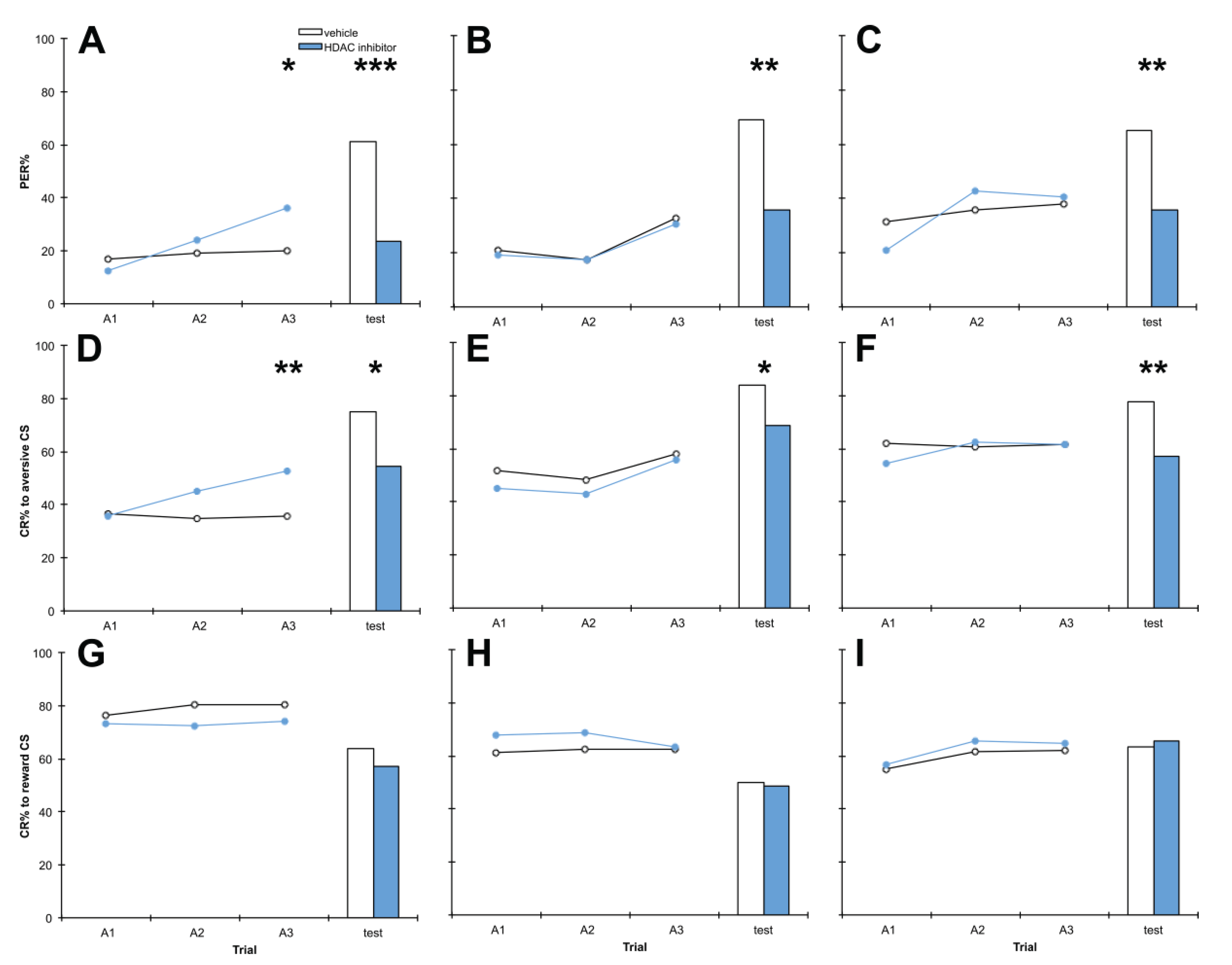
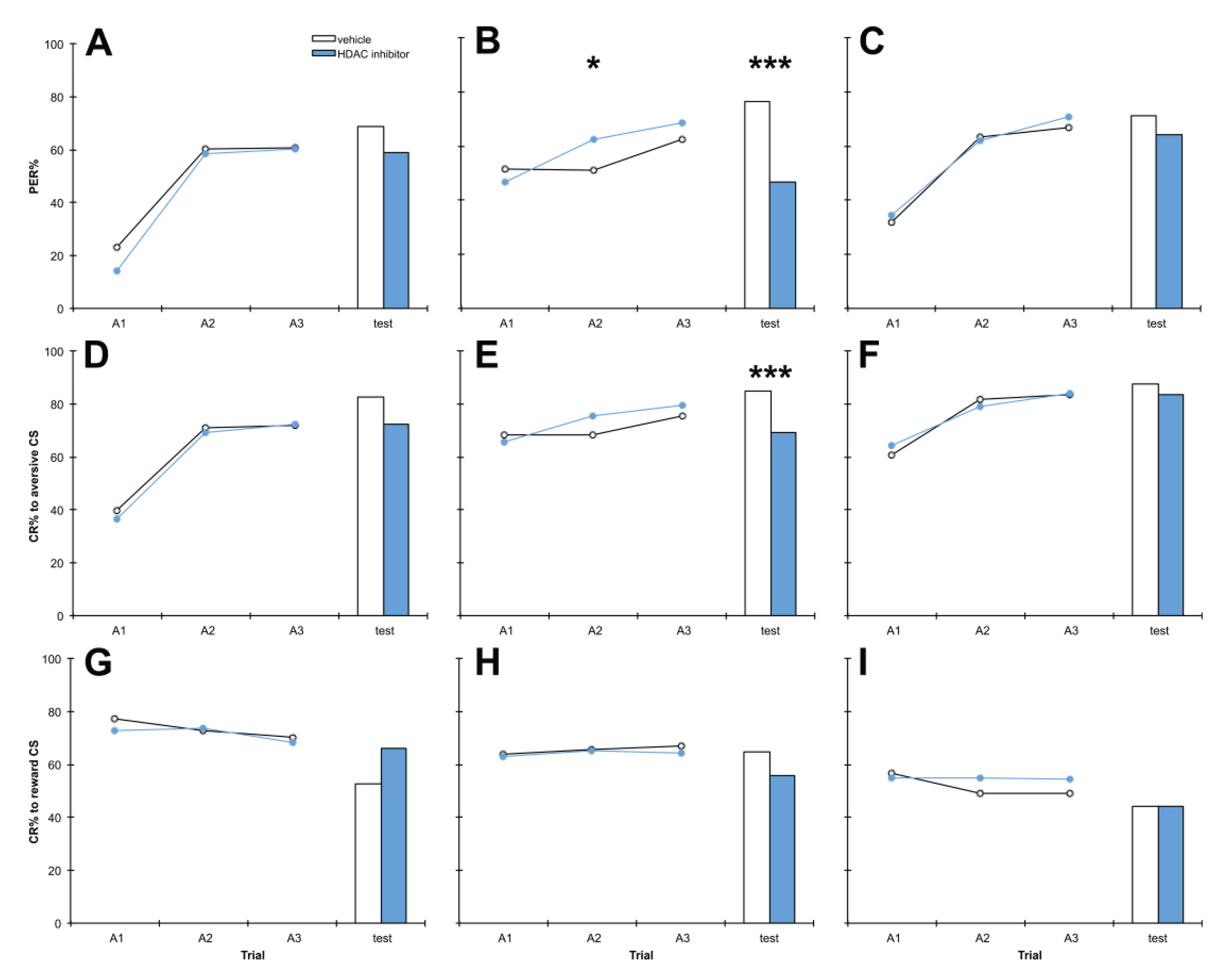
3.3. Treatment with PB 1 h after Training Impairs Aversive Memory but Has no Effect on Reward Memory
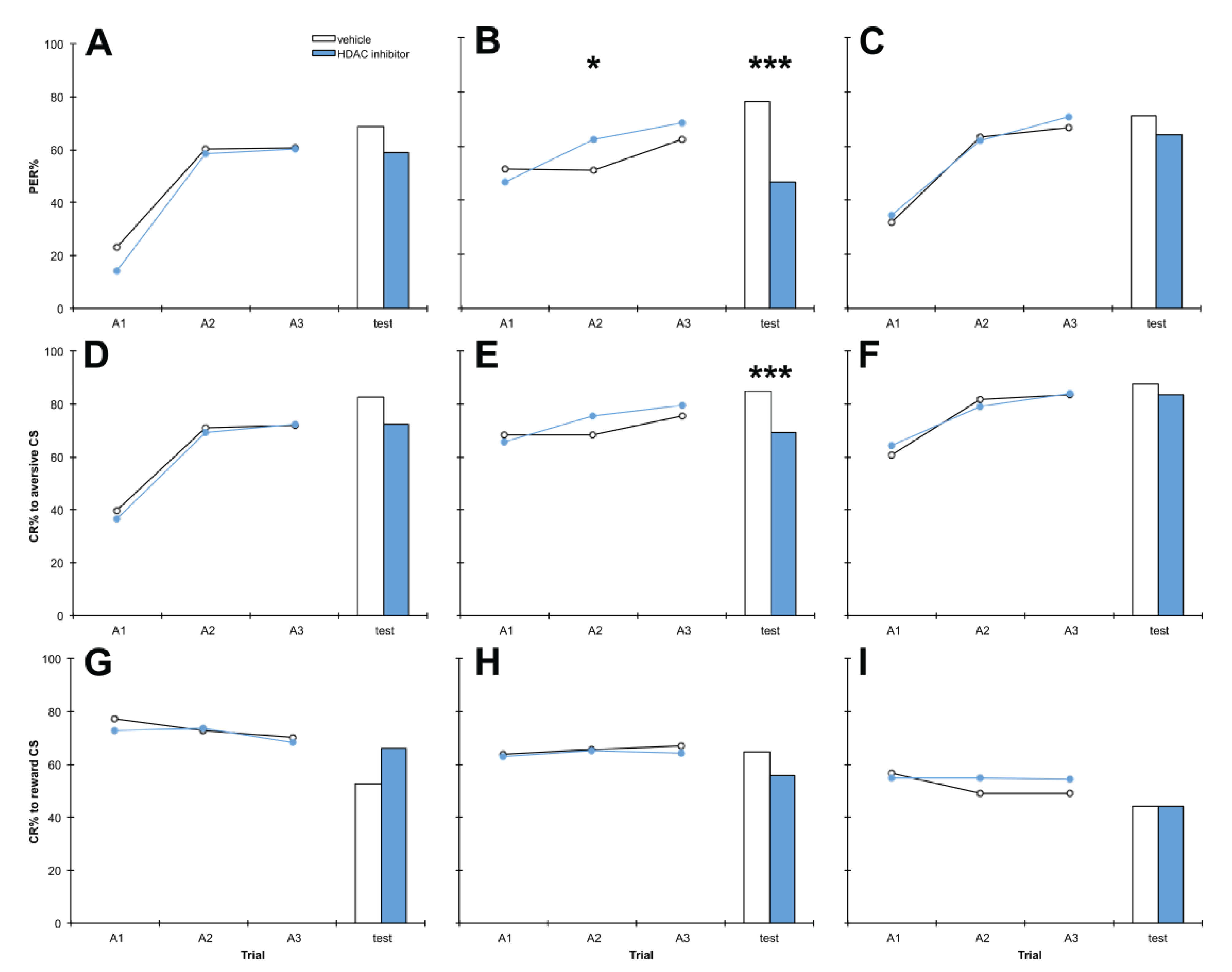
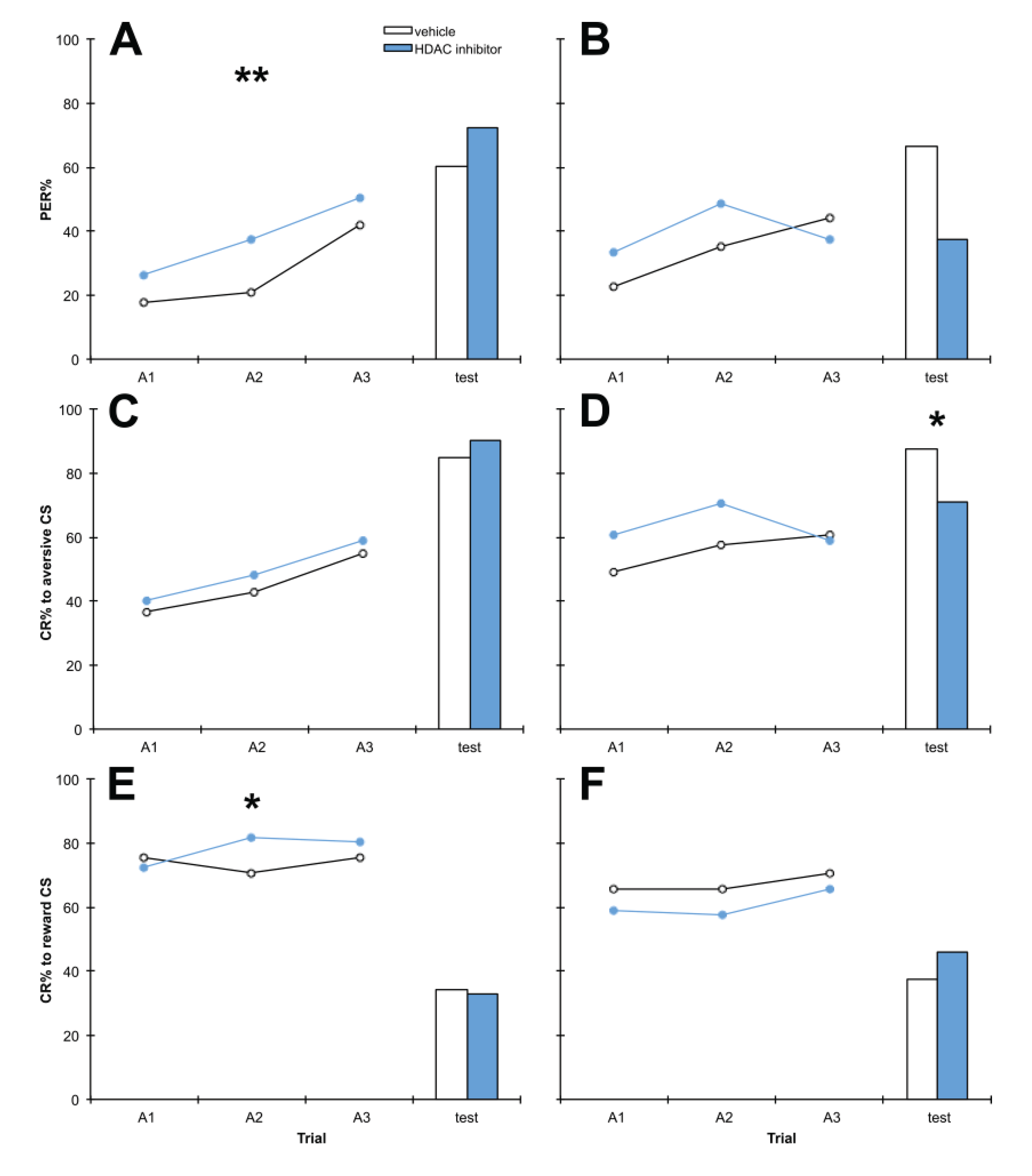
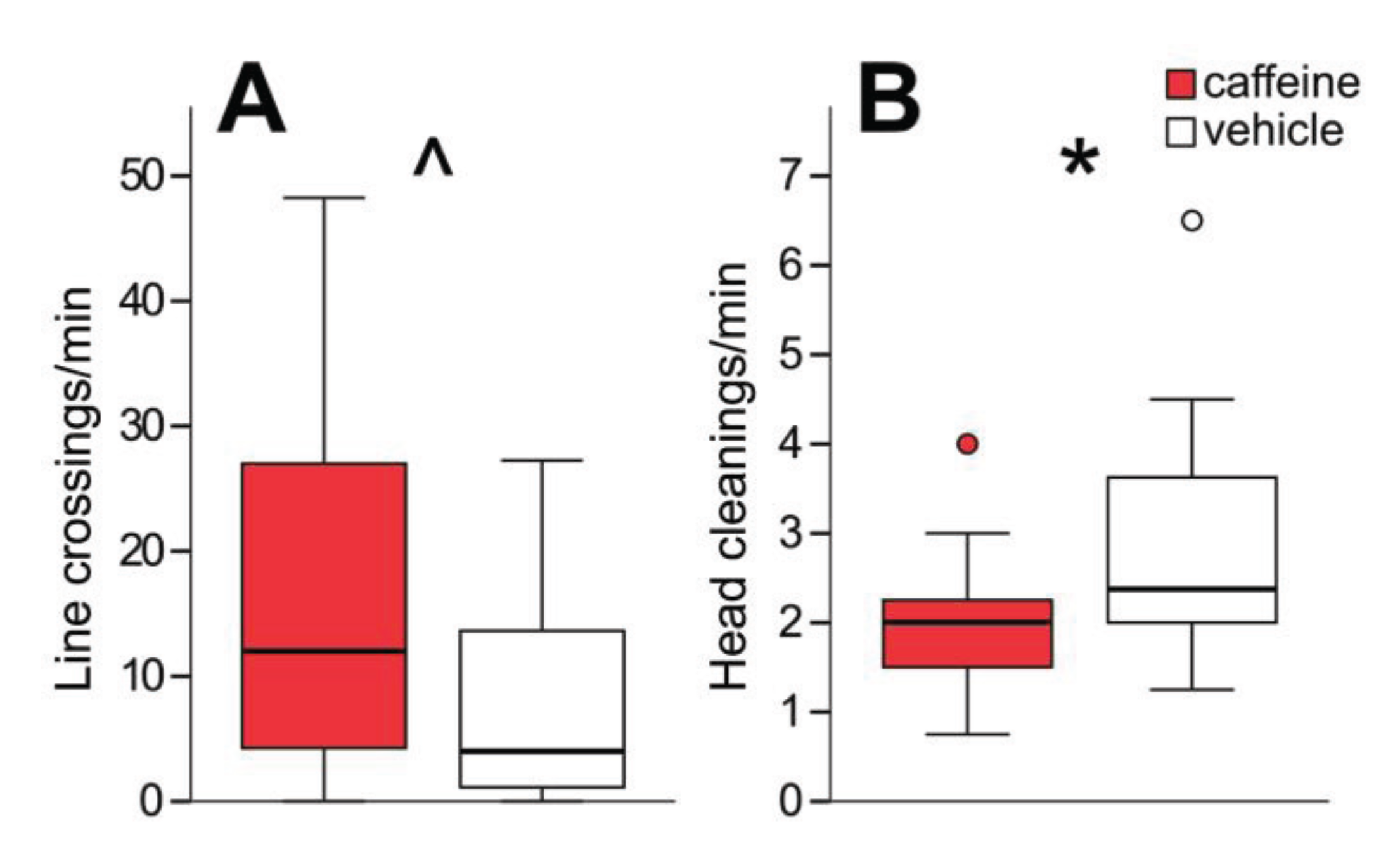
3.4. A Range of HDAC Inhibitors Selectively Impair Aversive Memory
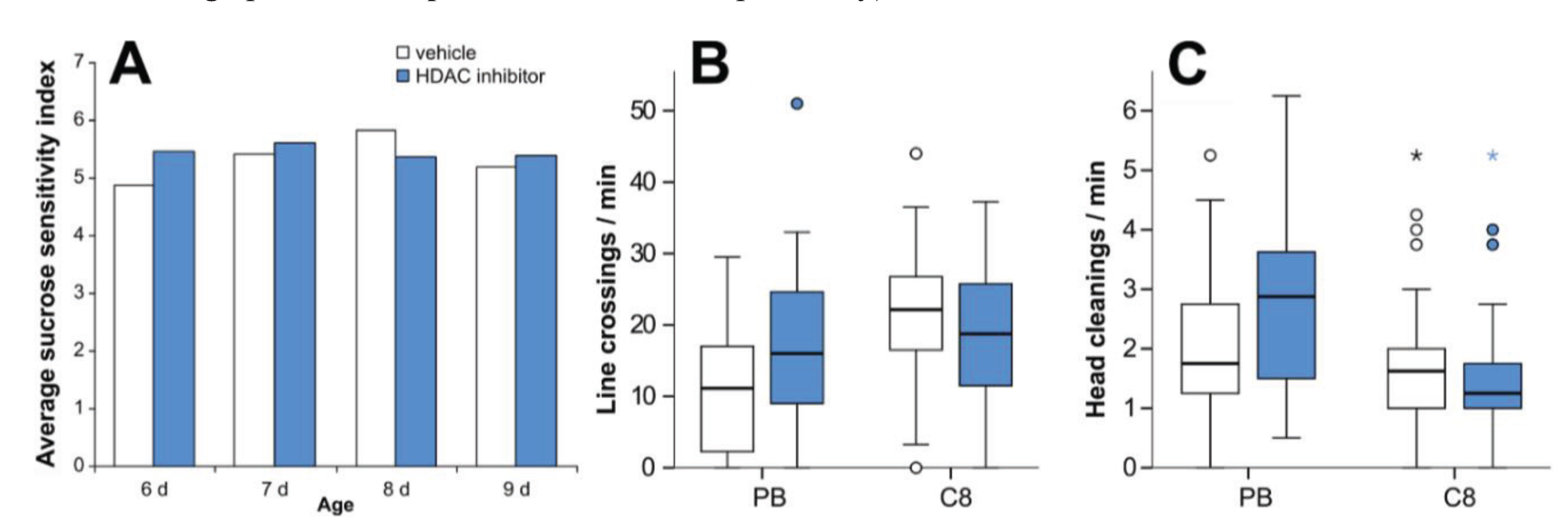
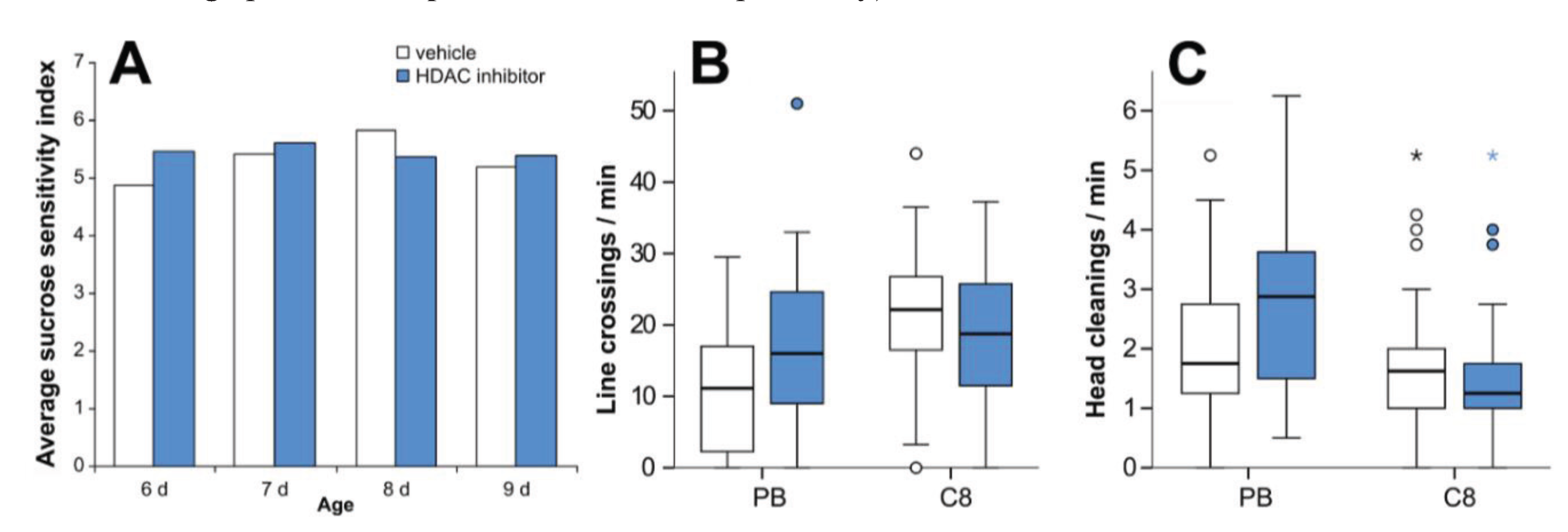
3.5. HDAC Inhibition Does not Affect Sucrose Sensitivity or Locomotor Activity
4. Discussion
4.1. C8 and PB Impair Aversive Memory but not Reward Memory in a Dose-Dependent Manner
4.2. Treatment with C8 1 h before Training, 1 h after Training or 1 h before Testing Impairs Aversive Memory but Has no Effect on Reward Memory
4.3. Treatment with PB 1 h after Training Impairs Aversive Memory but Has no Effect on Reward Memory
4.4. A Range of HDAC Inhibitors Selectively Impair Aversive Memory
4.5. HDAC Inhibition Does not Affect Locomotor Activity or Sucrose Sensitivity
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Roth, T.L.; Roth, E.D.; Sweatt, J.D. Epigenetic regulation of genes in learning and memory. Essays Biochem. 2010, 48, 263–274. [Google Scholar] [CrossRef]
- Lockett, G.A.; Wilkes, F.; Maleszka, R. Brain plasticity, memory and neurological disorders: An epigenetic perspective. NeuroReport 2010, 21, 909–913. [Google Scholar] [CrossRef]
- Lockett, G.A.; Patil, V.K.; Soto-Ramírez, N.; Ziyab, A.H.; Holloway, J.W.; Karmaus, W. Epigenomics and allergic disease. Epigenomics 2013, 5, 685–699. [Google Scholar] [CrossRef]
- Lockett, G.A.; Helliwell, P.; Maleszka, R. Involvement of DNA methylation in memory processing in the honey bee. NeuroReport 2010, 21, 812–816. [Google Scholar] [CrossRef]
- Day, J.J.; Sweatt, J.D. DNA methylation and memory formation. Nat. Neurosci. 2010, 13, 1319–1323. [Google Scholar] [CrossRef]
- Biergans, S.D.; Jones, J.C.; Treiber, N.; Galizia, C.G.; Szyszka, P. DNA Methylation mediates the discriminatory power of associative long-term memory in honeybees. PLoS One 2012, 7, e39349. [Google Scholar]
- Guan, Z.; Giustetto, M.; Lomvardas, S.; Kim, J.-H.; Miniaci, M.C.; Schwartz, J.H.; Thanos, D.; Kandel, E.R. Integration of long-term-memory-related synaptic plasticity involves bidirectional regulation of gene expression and chromatin structure. Cell 2002, 111, 483–493. [Google Scholar] [CrossRef]
- Federman, N.; Fustiñana, M.S.; Romano, A. Histone acetylation is recruited in consolidation as a molecular feature of stronger memories. Learn. Mem. 2009, 16, 600–606. [Google Scholar] [CrossRef]
- Grunstein, M. Histone acetylation in chromatin structure and transcription. Nature 1997, 389, 349–352. [Google Scholar] [CrossRef]
- Guan, J.S.; Haggarty, S.J.; Giacometti, E.; Dannenberg, J.H.; Joseph, N.; Gao, J.; Nieland, T.J.F.; Zhou, Y.; Wang, X.; Mazitschek, R. HDAC2 negatively regulates memory formation and synaptic plasticity. Nature 2009, 459, 55–60. [Google Scholar] [CrossRef]
- Levenson, J.M.; O’Riordan, K.J.; Brown, K.D.; Trinh, M.A.; Molfese, D.L.; Sweatt, J.D. Regulation of histone acetylation during memory formation in the hippocampus. J. Biol. Chem. 2004, 279, 40545–40559. [Google Scholar]
- Bredy, T.W.; Barad, M. The histone deacetylase inhibitor valproic acid enhances acquisition, extinction, and reconsolidation of conditioned fear. Learn. Mem. 2008, 15, 39–45. [Google Scholar] [CrossRef]
- Lattal, K.M.; Barrett, R.M.; Wood, M.A. Systemic or intrahippocampal delivery of histone deacetylase inhibitors facilitates fear extinction. Behav. Neurosci. 2007, 121, 1125–1131. [Google Scholar] [CrossRef]
- Yeh, S.-H.; Lin, C.-H.; Gean, P.-W. Acetylation of nuclear Factor-κB in rat amygdala improves long-term but not short-term retention of fear memory. Mol. Pharmacol. 2004, 65, 1286–1292. [Google Scholar] [CrossRef]
- Stefanko, D.P.; Barrett, R.M.; Ly, A.R.; Reolon, G.K.; Wood, M.A. Modulation of long-term memory for object recognition via HDAC inhibition. Proc. Natl. Acad. Sci. USA 2009, 106, 9447–9452. [Google Scholar] [CrossRef]
- Fontán-Lozano, Á.; Romero-Granados, R.; Troncoso, J.; Múnera, A.; Delgado-García, J.M.; Carrión, Á.M. Histone deacetylase inhibitors improve learning consolidation in young and in KA-induced-neurodegeneration and SAMP-8-mutant mice. Mol. Cell. Neurosci. 2008, 39, 193–201. [Google Scholar] [CrossRef]
- Swank, M.W.; Sweatt, J.D. Increased Histone acetyltransferase and lysine acetyltransferase activity and biphasic activation of the ERK/RSK cascade in insular cortex during novel taste learning. J. Neurosci. 2001, 21, 3383–3391. [Google Scholar]
- Fischer, A.; Sananbenesi, F.; Wang, X.; Dobbin, M.; Tsai, L. Recovery of learning and memory is associated with chromatin remodelling. Nature 2007, 447, 178–182. [Google Scholar] [CrossRef]
- Umka, J.; Mustafa, S.; ElBeltagy, M.; Thorpe, A.; Latif, L.; Bennett, G.; Wigmore, P.M. Valproic acid reduces spatial working memory and cell proliferation in the hippocampus. Neuroscience 2010, 166, 15–22. [Google Scholar] [CrossRef]
- Alarcón, J.M.; Malleret, G.; Touzani, K.; Vronskaya, S.; Ishii, S.; Kandel, E.R.; Barco, A. Chromatin acetylation, memory, and LTP are impaired in CBP+/− mice: A model for the cognitive deficit in Rubinstein-Taybi syndrome and its amelioration. Neuron 2004, 42, 947–959. [Google Scholar] [CrossRef]
- Merschbaecher, K.; Haettig, J.; Mueller, U. Acetylation-mediated suppression of transcription-independent memory: Bidirectional modulation of memory by acetylation. PLoS One 2012, 7, e45131. [Google Scholar] [CrossRef]
- Miller, C.A.; Campbell, S.L.; Sweatt, J.D. DNA methylation and histone acetylation work in concert to regulate memory formation and synaptic plasticity. Neurobiol. Learn. Mem. 2008, 89, 599–603. [Google Scholar] [CrossRef]
- Smith, B.H.; Abramson, C.I.; Tobin, T.R. Conditional withholding of proboscis extension in honeybees (Apis mellifera) during discriminative punishment. J. Comp. Psychol. 1991, 105, 345–356. [Google Scholar] [CrossRef]
- Vergoz, V.; Roussel, E.; Sandoz, J.-C.; Giurfa, M. Aversive learning in honeybees revealed by the olfactory conditioning of the sting extension reflex. PLoS One 2007, 2, e288. [Google Scholar] [CrossRef]
- Lockett, G.A.; Kucharski, R.; Maleszka, R. DNA methylation changes elicited by social stimuli in the brains of worker honey bees. Genes Brain Behav. 2012, 11, 235–242. [Google Scholar] [CrossRef]
- Foret, S.; Kucharski, R.; Pittelkow, Y.; Lockett, G.A.; Maleszka, R. Epigenetic regulation of the honey bee transcriptome: Unravelling the nature of methylated genes. BMC Genomics 2009, 10, 472. [Google Scholar] [CrossRef]
- Miklos, G.L.G.; Maleszka, R. Epigenomic communication systems in humans and honey bees: From molecules to behavior. Horm. Behav. 2011, 59, 399–406. [Google Scholar] [CrossRef]
- Kucharski, R.; Maleszka, J.; Foret, S.; Maleszka, R. Nutritional control of reproductive status in honeybees via DNA methylation. Science 2008, 319, 1827–1830. [Google Scholar] [CrossRef]
- Dickman, M.J.; Kucharski, R.; Maleszka, R.; Hurd, P.J. Extensive histone post-translational modification in honey bees. Insect Biochem. Mol. Biol. 2012, 43, 125–137. [Google Scholar]
- Menzel, R. The honeybee as a model for understanding the basis of cognition. Nat. Rev. Neurosci. 2012, 13, 758–768. [Google Scholar] [CrossRef]
- Bitterman, M.E.; Menzel, R.; Fietz, A.; Schäfer, S. Classical conditioning of proboscis extension in honeybees (Apis mellifera). J. Comp. Psychol. 1983, 97, 107–119. [Google Scholar] [CrossRef]
- Maleszka, R.; Helliwell, P.; Kucharski, R. Pharmacological interference with glutamate re-uptake impairs long-term memory in the honeybee, Apis mellifera. Behav. Brain Res. 2000, 115, 49–53. [Google Scholar] [CrossRef]
- Kuwabara, M. Bildung des bedingten Reflexes von Pavlovs Typus bei der Honigbiene, Apis mellifica. J. Fac. Sci. Hokkaido Univ. Ser. VI Zool. 1957, 13, 458–464. [Google Scholar]
- Massa, S.; Mai, A.; Sbardella, G.; Esposito, M.; Ragno, R.; Loidl, P.; Brosch, G. 3-(4-Aroyl-1H-pyrrol-2-yl)-N-hydroxy-2-propenamides, a new class of synthetic histone deacetylase inhibitors. J. Medicinal. Chem. 2001, 44, 2069–2072. [Google Scholar] [CrossRef]
- Mai, A.; Massa, S.; Ragno, R.; Esposito, M.; Sbardella, G.; Nocca, G.; Scatena, R.; Jesacher, F.; Loidl, P.; Brosch, G. Binding mode analysis of 3-(4-Benzoyl-1-methyl-1H-2-pyrrolyl)-N-hydroxy-2-propenamide: A new synthetic histone deacetylase inhibitor inducing histone hyperacetylation, growth inhibition, and terminal cell differentiation. J. Medicinal. Chem. 2002, 45, 1778–1784. [Google Scholar] [CrossRef]
- Mai, A.; Massa, S.; Ragno, R.; Cerbara, I.; Jesacher, F.; Loidl, P.; Brosch, G. 3-(4-Aroyl-1-methyl-1H-2-pyrrolyl)-N-hydroxy-2-alkylamides as a new class of synthetic histone deacetylase inhibitors. 1. Design, synthesis, biological evaluation, and binding mode studies performed through three different docking procedures. J. Medicinal. Chem. 2003, 46, 512–524. [Google Scholar] [CrossRef]
- Ricobaraza, A.; Cuadrado-Tejedor, M.; Pérez-Mediavilla, A.; Frechilla, D.; Del Río, J.; García-Osta, A. Phenylbutyrate Ameliorates cognitive deficit and reduces tau pathology in an alzheimer’s disease mouse model. Neuropsychopharmacology 2009, 34, 1721–1732. [Google Scholar] [CrossRef]
- Si, A.; Helliwell, P.; Maleszka, R. Effects of NMDA receptor antagonists on olfactory learning and memory in the honeybee (Apis mellifera). Pharmacol. Biochem. Behav. 2004, 77, 191–197. [Google Scholar] [CrossRef]
- Kucharski, R.; Mitri, C.; Grau, Y.; Maleszka, R. Characterization of a metabotropic glutamate receptor in the honeybee (Apis mellifera): Implications for memory formation. Invertebr. Neurosci. 2007, 7, 99–108. [Google Scholar] [CrossRef]
- Bicker, G. Transmitter-induced calcium signalling in cultured neurons of the insect brain. J. Neurosci. Methods 1996, 69, 33–41. [Google Scholar] [CrossRef]
- Barron, A.B.; Maleszka, J.; vander Meer, R.K.; Robinson, G.E.; Maleszka, R. Comparing injection, feeding and topical application methods for treatment of honeybees with octopamine. J. Insect Physiol. 2007, 53, 187–194. [Google Scholar] [CrossRef]
- Barron, A.B.; Maleszka, R.; Helliwell, P.G.; Robinson, G.E. Effects of cocaine on honey bee dance behaviour. J. Exp. Biol. 2009, 212, 163–168. [Google Scholar]
- Vergoz, V. Effects of Queen Mandibular Pheromone on Locomotor Behaviour and Learning in Worker Honey Bees Apis mellifera; Otago University: Dunedin, New Zealand, 2008. [Google Scholar]
- Beggs, K.T.; Glendining, K.A.; Marechal, N.M.; Vergoz, V.; Nakamura, I.; Slessor, K.N.; Mercer, A.R. Queen pheromone modulates brain dopamine function in worker honey bees. Proc. Natl. Acad. Sci. USA 2007, 104, 2460–2464. [Google Scholar]
- Si, A.; Zhang, S.-W.; Maleszka, R. Effects of caffeine on olfactory and visual learning in the honey bee (Apis mellifera). Pharmacol. Biochem. Behav. 2005, 82, 664–672. [Google Scholar] [CrossRef]
- Antoniou, K.; Kafetzopoulos, E.; Papadopoulou-Daifoti, Z.; Hyphantis, T.; Marselos, M. D-amphetamine, cocaine and caffeine: A comparative study of acute effects on locomotor activity and behavioural patterns in rats. Neurosci. Biobehav. Rev. 1998, 23, 189–196. [Google Scholar] [CrossRef]
- Malechuk, A. The Effects of Caffeine on Learning and Locomotor Behavior in the Honey Bee (Apis mellifera); Wake Forest University: Winston-Salem, NC, USA, 2009. [Google Scholar]
- Cysique, L.; Maruff, P.; Brew, B. Valproic acid is associated with cognitive decline in HIV-infected individuals: A clinical observational study. BMC Neurol. 2006, 6, 42. [Google Scholar] [CrossRef]
- Masmoudi, K.; Gras-Champel, V.; Masson, H.; Andréjak, M. Parkinsonism and/or cognitive impairment with valproic acid therapy: A report of ten cases. Pharmacopsychiatry 2006, 39, 9–12. [Google Scholar] [CrossRef]
- Ristić, A.J.; Vojvodić, N.; Janković, S.; Sindelić, A.; Sokić, D. The frequency of reversible parkinsonism and cognitive decline associated with valproate treatment: A study of 364 patients with different types of epilepsy. Epilepsia 2006, 47, 2183–2185. [Google Scholar] [CrossRef]
- Hommet, C.; Mondon, K.; de Toffol, B.; Constans, T. Reversible cognitive and neurological symptoms during valproic acid therapy. J. Am. Geriatr. Soc. 2007, 55, 628–628. [Google Scholar]
- Sleiman, S.F.; Basso, M.; Mahishi, L.; Kozikowski, A.P.; Donohoe, M.E.; Langley, B.; Ratan, R.R. Putting the ‘HAT’ back on survival signalling: The promises and challenges of HDAC inhibition in the treatment of neurological conditions. Expert Opin. Invest. Drugs 2009, 18, 573–584. [Google Scholar] [CrossRef]
- Schwaerzel, M.; Monastirioti, M.; Scholz, H.; Friggi-Grelin, F.; Birman, S.; Heisenberg, M. Dopamine and octopamine differentiate between aversive and appetitive olfactory memories in drosophila. J. Neurosci. 2003, 23, 10495–10502. [Google Scholar]
- Unoki, S.; Matsumoto, Y.; Mizunami, M. Participation of octopaminergic reward system and dopaminergic punishment system in insect olfactory learning revealed by pharmacological study. Eur. J. Neurosci. 2005, 22, 1409–1416. [Google Scholar] [CrossRef]
- Mercer, A.R.; Menzel, R. The effects of biogenic amines on conditioned and unconditioned responses to olfactory stimuli in the honeybee Apis mellifera. J. Comp. Physiol. A Neuroethol. Sens. Neural. Behav. Physiol. 1982, 145, 363–368. [Google Scholar] [CrossRef]
- Barron, A.B.; Maleszka, R.; vander Meer, R.K.; Robinson, G.E. Octopamine modulates honey bee dance behavior. Proc. Natl. Acad. Sci. USA 2007, 104, 1703–1707. [Google Scholar] [CrossRef]
- Karantzali, E.; Schulz, H.; Hummel, O.; Hubner, N.; Hatzopoulos, A.K.; Kretsovali, A. Histone deacetylase inhibition accelerates the early events of stem cell differentiation: Transcriptomic and epigenetic analysis. Genome Biol. 2008, 9, R65. [Google Scholar] [CrossRef]
- Vecsey, C.G.; Hawk, J.D.; Lattal, K.M.; Stein, J.M.; Fabian, S.A.; Attner, M.A.; Cabrera, S.M.; McDonough, C.B.; Brindle, P.K.; Abel, T. Histone deacetylase inhibitors enhance memory and synaptic plasticity via CREB:CBP-dependent transcriptional activation. J. Neurosci. 2007, 27, 6128–6140. [Google Scholar] [CrossRef]
- Levenson, J.M.; Sweatt, J.D. Epigenetic mechanisms in memory formation. Nat. Rev. Neurosci. 2005, 6, 108–118. [Google Scholar] [CrossRef]
- Lyko, F.; Foret, S.; Kucharski, R.; Wolf, S.; Falckenhayn, C.; Maleszka, R. The honey bee epigenomes: Differential methylation of brain DNA in queens and workers. PLoS Biol. 2010, 8, e1000506. [Google Scholar] [CrossRef]
- Kilgore, M.; Miller, C.A.; Fass, D.M.; Hennig, K.M.; Haggarty, S.J.; Sweatt, J.D.; Rumbaugh, G. Inhibitors of class 1 histone deacetylases reverse contextual memory deficits in a mouse model of alzheimer’s disease. Neuropsychopharmacology 2010, 35, 870–880. [Google Scholar] [CrossRef]
- Blackwell, L.; Norris, J.; Suto, C.M.; Janzen, W.P. The use of diversity profiling to characterize chemical modulators of the histone deacetylases. Life Sci. 2008, 82, 1050–1058. [Google Scholar] [CrossRef]
- Bolden, J.E.; Peart, M.J.; Johnstone, R.W. Anticancer activities of histone deacetylase inhibitors. Nat. Rev. Drug Discov. 2006, 5, 769–784. [Google Scholar] [CrossRef]
- Sugo, N.; Oshiro, H.; Takemura, M.; Kobayashi, T.; Kohno, Y.; Uesaka, N.; Song, W.-J.; Yamamoto, N. Nucleocytoplasmic translocation of HDAC9 regulates gene expression and dendritic growth in developing cortical neurons. Eur. J. Neurosci. 2010, 31, 1521–1532. [Google Scholar]
- Van Thuan, N.; Hong-Thuy, B.; Wakayama, S.; Kishigami, S.; Ohta, H.; Hikichi, T.; Mizutani, E.; Wakayama, T. 95 observation of chromosome decondensation, histone H3 modification, and HP1 protein in mouse cloned embryos following inhibition of histone deacetylations and cyclin-dependent kinase. Reprod. Fertil. Dev. 2007, 19, 164–165. [Google Scholar]
- Menzel, R. Massed and spaced learning in honeybees: The role of CS, US, the intertrial interval, and the test interval. Learn. Mem. 2001, 8, 198–208. [Google Scholar] [CrossRef]
- Mizunami, M.; Unoki, S.; Mori, Y.; Hirashima, D.; Hatano, A.; Matsumoto, Y. Roles of octopaminergic and dopaminergic neurons in appetitive and aversive memory recall in an insect. BMC Biol. 2009, 7, 46. [Google Scholar] [CrossRef]
- Lefer, D.; Perisse, E.; Hourcade, B.; Sandoz, J.; Devaud, J.-M. Two waves of transcription are required for long-term memory in the honeybee. Learn. Mem. 2013, 20, 29–33. [Google Scholar]
- Korzus, E.; Rosenfeld, M.G.; Mayford, M. CBP Histone acetyltransferase activity is a critical component of memory consolidation. Neuron 2004, 42, 961–972. [Google Scholar] [CrossRef]
- Menzel, R. Searching for the memory trace in a mini-brain, the honeybee. Learn. Mem. 2001, 8, 53–62. [Google Scholar] [CrossRef]
- Scheiner, R.; Barnert, M.; Erber, J. Variation in water and sucrose responsiveness during the foraging season affects proboscis extension learning in honey bees. Apidologie 2003, 34, 67–72. [Google Scholar] [CrossRef]
- Galizia, C.G.; Menzel, R. Odour perception in honeybees: Coding information in glomerular patterns. Curr. Opin. Neurobiol. 2000, 10, 504–510. [Google Scholar] [CrossRef]
- Guerrieri, F.; Schubert, M.; Sandoz, J.-C.; Giurfa, M. Perceptual and neural olfactory similarity in honeybees. PLoS Biol. 2005, 3, e60. [Google Scholar] [CrossRef]
- Maleszka, R.; Mason, P.H.; Barron, A.B. Epigenomics and the concept of degeneracy in biological systems. Brief. Funct. Genomics. 2014, 13, 191–202. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lockett, G.A.; Wilkes, F.; Helliwell, P.; Maleszka, R. Contrasting Effects of Histone Deacetylase Inhibitors on Reward and Aversive Olfactory Memories in the Honey Bee. Insects 2014, 5, 377-398. https://doi.org/10.3390/insects5020377
Lockett GA, Wilkes F, Helliwell P, Maleszka R. Contrasting Effects of Histone Deacetylase Inhibitors on Reward and Aversive Olfactory Memories in the Honey Bee. Insects. 2014; 5(2):377-398. https://doi.org/10.3390/insects5020377
Chicago/Turabian StyleLockett, Gabrielle A, Fiona Wilkes, Paul Helliwell, and Ryszard Maleszka. 2014. "Contrasting Effects of Histone Deacetylase Inhibitors on Reward and Aversive Olfactory Memories in the Honey Bee" Insects 5, no. 2: 377-398. https://doi.org/10.3390/insects5020377
APA StyleLockett, G. A., Wilkes, F., Helliwell, P., & Maleszka, R. (2014). Contrasting Effects of Histone Deacetylase Inhibitors on Reward and Aversive Olfactory Memories in the Honey Bee. Insects, 5(2), 377-398. https://doi.org/10.3390/insects5020377