Bacillus thuringiensis Is an Environmental Pathogen and Host-Specificity Has Developed as an Adaptation to Human-Generated Ecological Niches

Abstract
:1. Introduction
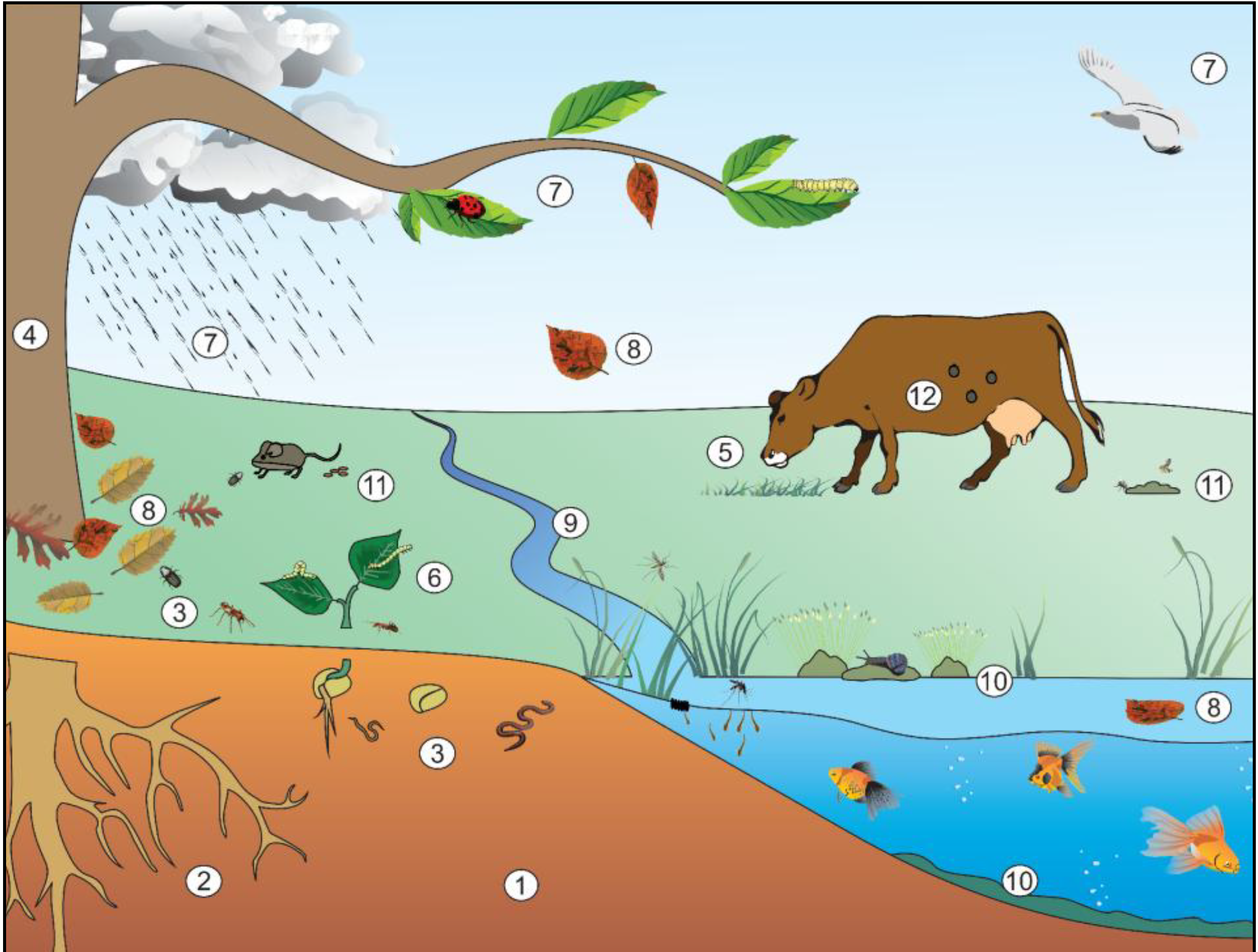
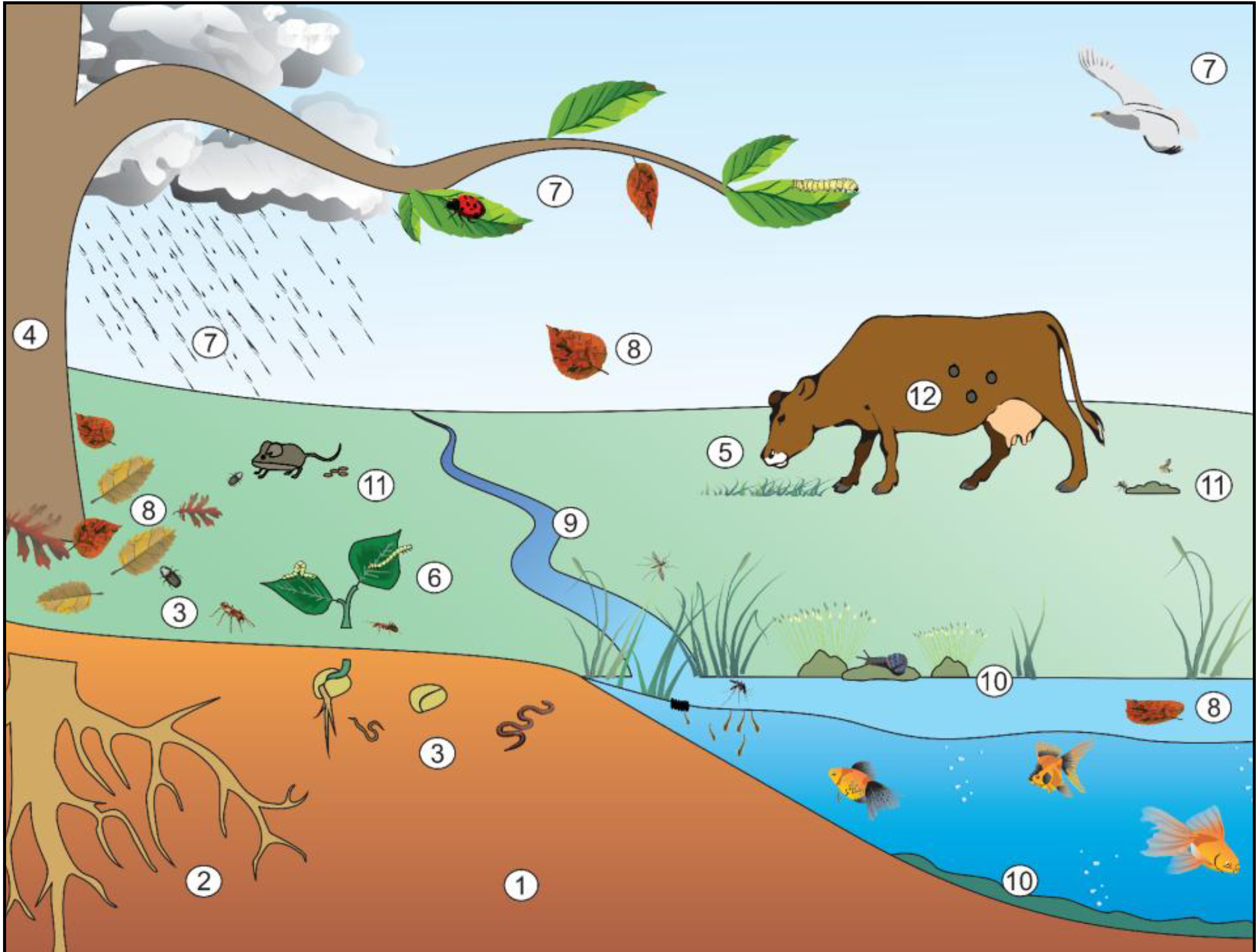
2. Niches Occupied by Bt
2.1. Soil Environment
2.2. Epiphytic Environment—Phylloplane
2.3. Epiphytic Environment—Rhizosphere
2.4. Endophytic Environment
2.5. Aquatic Environment
2.6. Paratenic Behavior
2.7. Pathogenic Behavior
2.7.1. Pathogenic Arsenal of Bt Cells towards Insects
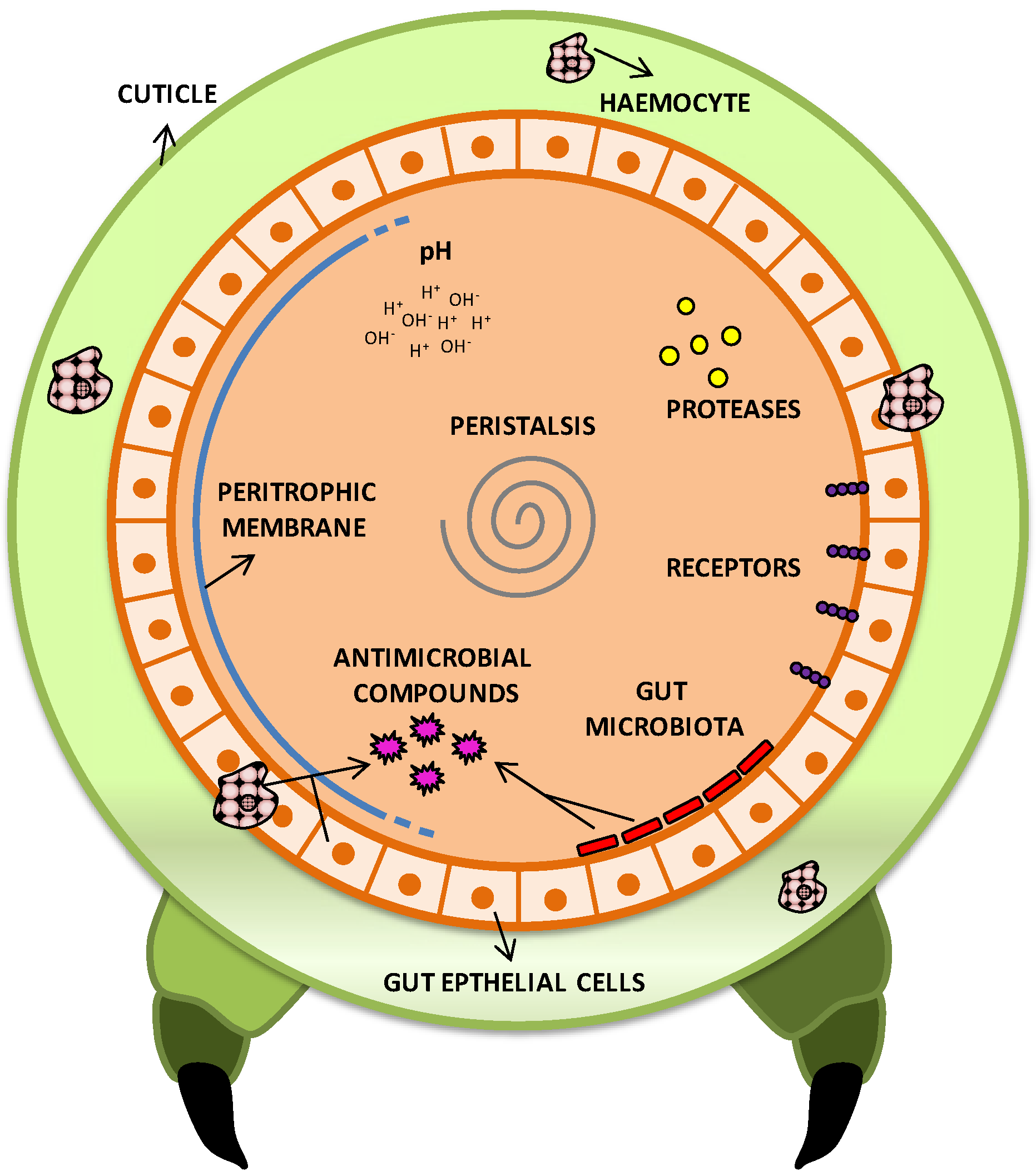

{kind=link}
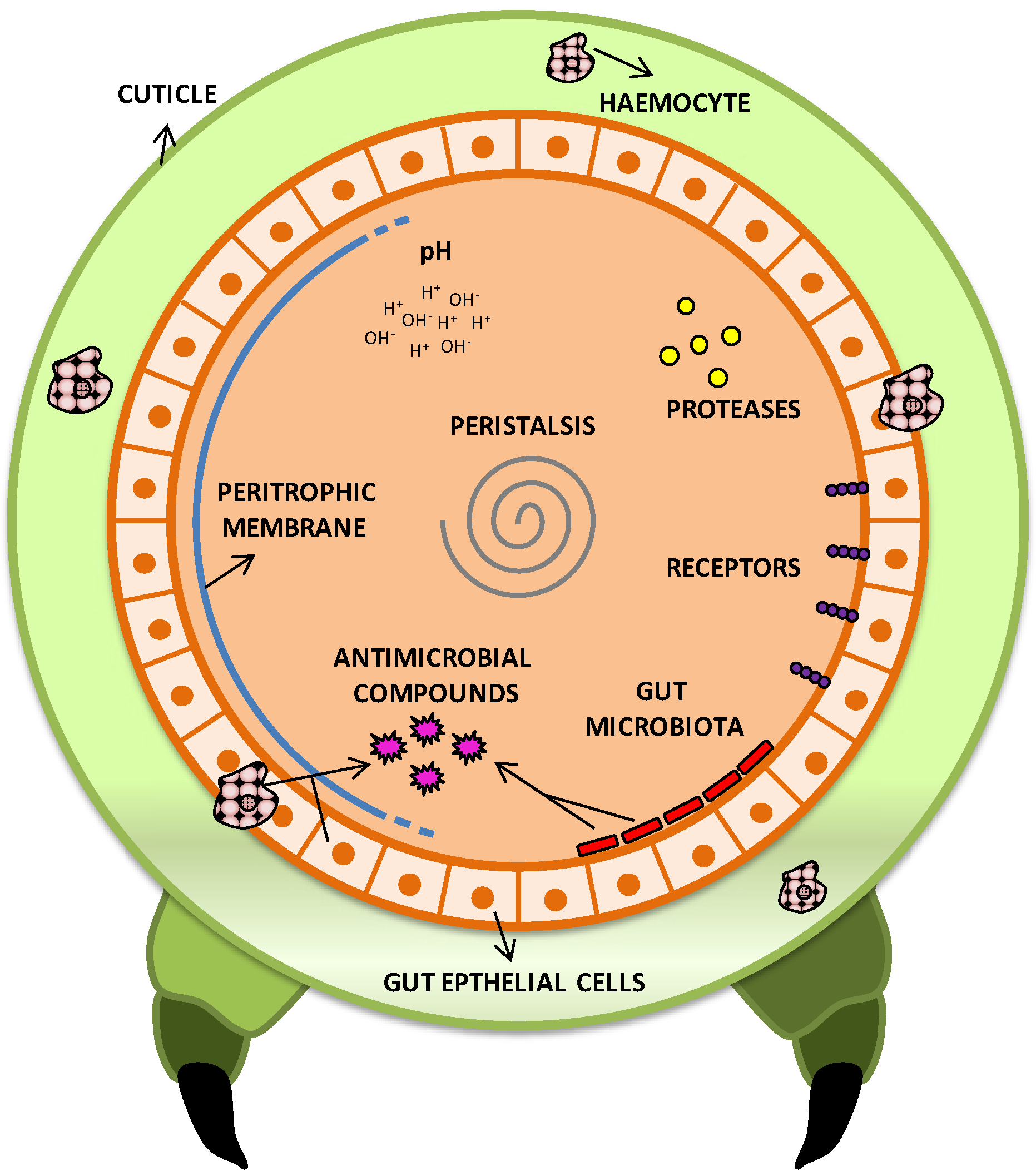{kind=link}
| Virulence strategies | Function | References |
|---|---|---|
| Toxins production | Degradation of intestinal membrane and release of nutrients, favoring spore germination and colonization. Gate opening to reach hemolymph and cause septicemia. | [1,2,6,66,93] |
| Antimicrobial resistance | Resistance to antimicrobials produced by the host and midgut resident microbiota, allowing colonization. | [94] |
| Antimicrobial production | Decrease of competition for space and nutrients, and reduction of antimicrobial compounds production by midgut microbiota. | [95,96] |
| Peristalsis and feeding reduction | Decrease of toxins and Bt cells elimination from intestine. | [6,97] |
| Production of degrading enzymes | Degradation of antimicrobial agents from host and facilitation of intestinal colonization. Direct toxicity in some cases. | [92,98,99,100,101,102,103] |
| Imune system resistance | Prevention of phagocytosis and resistance to antimicrobial compounds, digestive enzymes, pH, reactive oxygen species. | [92,104] |
2.7.1.1. Protein Toxins that Need Receptors in Host-Cells Membranes
2.7.1.2. Proteins and Toxins that Do Not Need Receptors in Host-Cell Membranes
2.7.1.3. Non-Proteinaceous Toxins
2.7.2. Pathogenicity to Other Taxa
3. Reconciling Alternative Ecological Views
4. Conclusions and Perspectives
Acknowledgments
Conflicts of Interest
References
- Aronson, A.I.; Beckman, W.; Dunn, P. Bacillus thuringiensis and related insect pathogens. Microbiol. Rev. 1986, 50, 1–24. [Google Scholar]
- Schnepf, E.; Crickmore, N.; van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D.R.; Dean, D.H. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. R. 1998, 62, 775–806. [Google Scholar]
- Bravo, A.; Sarabia, S.; Lopez, L.; Ontiveros, H.; Abarca, C.; Ortiz, A.; Ortiz, M.; Lina, L.; Villalobos, F.J.; Peña, G.; et al. Characterization of cry genes in a mexican Bacillus thuringiensis strain collection. Appl. Environ. Microb. 1998, 64, 4965–4972. [Google Scholar]
- Lecadet, M.M.; Frachon, E.; Dumanoir, V.C.; Ripouteau, H.; Hamon, S.; Laurent, P.; Thiéry, I. Updating the H-antigen classification of Bacillus thuringiensis. J. Appl. Microbiol. 1999, 86, 660–672. [Google Scholar] [CrossRef]
- Xu, D.; Côté, J.C. Sequence diversity of Bacillus thuringiensis flagellin (H antigen) protein at the intra-H serotype level. Appl. Environ. Microbiol. 2008, 74, 5524–5532. [Google Scholar] [CrossRef]
- Raymond, B.; Johnston, P.R.; Nielsen-LeRoux, C.; Lereclus, D.; Crickmore, N. Bacillus thuringiensis: An impotent pathogen? Trends Microbiol. 2010, 18, 189–194. [Google Scholar] [CrossRef]
- Crickmore, N. Beyond the spore: Past and future developments of Bacillus thuringiensis as a biopesticide. J. Appl. Microbiol. 2006, 101, 616–619. [Google Scholar] [CrossRef]
- Hendriksen, N.B.; Hansen, B.M. Long-term survival and germination of Bacillus thuringiensis var. kurstaki in a field trial. Can. J. Microbiol. 2002, 48, 256–261. [Google Scholar] [CrossRef]
- Marco, G.; Manuel, P. Ecological mysteries: Is Bacillus thuringiensis a real insect pathogen? Bt Res. 2012, 3, 1–2. [Google Scholar]
- Jensen, G.B.; Hansen, B.M.; Eilenberg, J.; Mahillon, J. The hidden lifestyles of Bacillus cereus and relatives. Appl. Environ. Microbiol. 2003, 5, 631–640. [Google Scholar]
- Cangelosi, G.A.; Freitag, N.E.; Buckley, M.R. From Outside to Inside: Environmental Microorganisms as Human Pathogens; American Society for Microbiology: Washington, DC, USA, 2004. [Google Scholar]
- Martin, P.A.W.; Travers, R.S. Worldwide abundance and distribution of Bacillus thuringiensis isolates. Appl. Environ. Microbiol. 1989, 55, 2437–2442. [Google Scholar]
- Fiuza, L.; Nielsen-Leroux, C.; Goze, E.; Frutos, R.; Charles, J. Binding of Bacillus thuringiensis Cry1 toxins to the midgut brush border membrane vesicles of Chilo. suppressalis (Lepidoptera: Pyralidae): Evidence of shared binding sites. Appl. Environ. Microbiol. 1996, 62, 1544–1549. [Google Scholar]
- Cappello, M.; Bungiro, R.D.; Harrison, L.M.; Bischof, L.J.; Griffitts, J.S.; Barrows, B.D.; Aroian, R.V. A purified Bacillus thuringiensis crystal protein with therapeutic activity against the hookworm parasite Ancylostoma. ceylanicum. Proc. Natl. Acad. Sci. USA 2006, 103, 15154–15159. [Google Scholar] [CrossRef]
- Fang, Y.; Li, Z.; Liu, J.; Shu, C.; Wang, X.; Zhang, X.; Yu, X.; Zhao, D.; Liu, G.; Hu, S.; et al. A pangenomic study of Bacillus thuringiensis. J. Genet. Genomics 2011, 38, 567–576. [Google Scholar] [CrossRef]
- Akiba, Y. Microbial ecology of Bacillus thuringiensis VI. Germination of Bacillus thuringiensis spore in the soil. Appl. Entomol. Zool. 1986, 21, 76–80. [Google Scholar]
- West, A.W.; Burges, H.D.; Dixon, T.J.; Wyborn, C.H. Effect of incubation in non-sterilised and autoclaved arable soil on survival of Bacillus thuringiensis and Bacillus cereus spore inocula. N. Z. J. Agric. Res. 1985, 28, 559–566. [Google Scholar] [CrossRef]
- West, A.W.; Burges, H.D.; Dixon, T.J. Survival of Bacillus thuringiensis and Bacillus cereus spore inocula in soil: Effects of ph, moisture, nutrient availability and indigenous microorganisms. Soil Biol. Biochem. 1985, 17, 657–665. [Google Scholar] [CrossRef]
- Saleh, S.M.; Harris, R.F.; Allen, O.N. Fate of Bacillus thuringiensis in soil: Effect of soil pH and organic amendment. Can. J. Microbiol. 1970, 16, 677–680. [Google Scholar] [CrossRef]
- Polanczyk, R.A.; Zanúncio, J.C.; Alves, S.B. Relationship between chemical properties of the soil and the occurrence of Bacillus thuringiensis. Ciênc. Rural 2009, 39, 1–5. [Google Scholar] [CrossRef]
- Akiba, Y.; Sekijima, Y.; Aizawa, K.; Fujiyoshi, N. Microbial ecological studies on Bacillus thuringiensis. II. Dynamics of Bacillus thuringiensis in sterilized soil. Jpn. J. Appl. Entomol. Zool. 1977, 21, 41–46. [Google Scholar] [CrossRef]
- West, A.W.; Burges, H.D.; White, R.J.; Wyborn, C.H. Persistence of Bacillus thuringiensis parasporal crystal insecticidal activity in soil. J. Invertebr. Pathol. 1984, 44, 128–133. [Google Scholar] [CrossRef]
- Akiba, Y. Microbial ecological studies on Bacillus thuringiensis. IV. The growth of Bacillus thuringiensis in soils of mulberry plantations. Jpn. J. Appl. Entomol. Zool. 1980, 24, 13–17. [Google Scholar] [CrossRef]
- Crecchio, C.S.G. Insecticidal activity and biodegradation of the toxin from Bacillus thuringiensis ssp. kurstaki bound to humic acids from soil. Soil Biol. Biochem. 1998, 30, 463–470. [Google Scholar] [CrossRef]
- Tapp, L.; Calamai, L.; Stotzky, G. Adsorption and binding of the insecticidal proteins from Bacillus thuringiensis subsp. kurstaki and subsp. tenebrionis on clay minerals. Soil Biol. Biochem. 1994, 26, 663–679. [Google Scholar] [CrossRef]
- Ohba, M.; Shisa, N.; Thaithanun, S.; Nakashima, K.; Lee, D.-H.; Ohgushi, A.; Wasano, N. A unique feature of Bacillus thuringiensis H-serotype flora in soils of a volcanic island of Japan. J. Gen. Appl. Microbiol. 2002, 48, 233–235. [Google Scholar] [CrossRef]
- Konecka, E.; Baranek, J.; Hrycak, A.; Kaznowski, A. Insecticidal activity of Bacillus thuringiensis strains isolated from soil and water. Sci. World J. 2012, 2012, 1–5. [Google Scholar]
- Monnerat, R.; Martins, E.; Queiroz, P.; Ordúz, S.; Jaramillo, G.; Benintende, G.; Cozzi, J.; Real, M.D.; Martinez-Ramirez, A.; Rausell, C.; et al. Genetic variability of Spodoptera. frugiperda Smith (Lepidoptera: Noctuidae) populations from Latin America is associated with variations in susceptibility to Bacillus thuringiensis cry toxins. Appl. Environ. Microbiol. 2006, 72, 7029–7035. [Google Scholar] [CrossRef]
- Guidi, V.; Patocchi, N.; Lüthy, P.; Tonolla, M. Distribution of Bacillus thuringiensis subsp. israelensis in soil of a Swiss Wetland reserve after 22 years of mosquito control. Appl. Environ. Microbiol. 2011, 77, 3663–3668. [Google Scholar] [CrossRef]
- Petras, S.F.; Casida, L.E., Jr. Survival of Bacillus thuringiensis spores in soil. Appl. Environ. Microbiol. 1985, 50, 1496–1501. [Google Scholar]
- Pedersen, J.C.; Damgaard, P.H.; Eilenberg, J.; Hansen, B.M. Dispersal of Bacillus thuringiensis var. kurstaki in an experimental cabbage field. Can. J. Microbiol. 1995, 41, 118–125. [Google Scholar] [CrossRef]
- Akiba, Y. Assessment of rainwater-mediated dispersion of field-sprayed Bacillus thuringiensis in the soil. Appl. Entomol. Zool. 1991, 26, 477–483. [Google Scholar]
- Wilcks, A.; Smidt, L.; Bahl, M.I.; Hansen, B.M.; Andrup, L.; Hendriksen, N.B.; Licht, T.R. Germination and conjugation of Bacillus thuringiensis subsp. israelensis in the intestine of gnotobiotic rats. J. Appl. Microbiol. 2008, 104, 1252–1259. [Google Scholar] [CrossRef]
- Bizzarri, M.F.; Bishop, A.H. The Ecology of Bacillus thuringiensis on the phylloplane: Colonization from soil, plasmid transfer, and interaction with larvae of Pieris brassica. Microb. Ecol. 2008, 56, 133–139. [Google Scholar] [CrossRef]
- Ammons, D.R.; Reyna, A.; Granados, J.C.; Samlal, M.S.; Rampersad, J.N. An investigation of Bacillus thuringiensis in rectal-collected fecal samples of cows. Curr. Microbiol. 2009, 59, 532–536. [Google Scholar] [CrossRef]
- Zhang, L.; Peng, Y.; Wu, S.; Sun, L.; Huang, E.; Huang, T.; Xu, L.; Wu, C.; Gelbic, I.; Guan, X. Microbial ecology and association of Bacillus thuringiensis in chicken feces originating from feed. Curr. Microbiol. 2012, 65, 784–791. [Google Scholar]
- Naryanan, M.S. Competitive Ability and Host Exploitation in Bacillus thuringiensis. MSc Thesis, University of Oxford, Oxford, UK, 2006. [Google Scholar]
- Dubois, T.; Faegri, K.; Perchat, S.; Lemy, C.; Buisson, C.; Nielsen-LeRoux, C.; Gohar, M.; Jacques, P.; Ramarao, N.; Kolstø, A.-B.; et al. Necrotrophism is a quorum-sensing-regulated lifestyle in Bacillus thuringiensis. PLoS Pathog. 2012, 8, e1002629. [Google Scholar] [CrossRef]
- Ichimatsu, T.; Mizuki, E.; Nishimura, K.; Akao, T.; Saitoh, H.; Higuchi, K.; Ohba, M. Occurrence of Bacillus thuringiensis in fresh waters of Japan. Curr. Microbiol. 2000, 40, 217–220. [Google Scholar] [CrossRef]
- Lachhab, K.; Tyagi, R.D.; Valéro, J.R. Production of Bacillus thuringiensis biopesticides using wastewater sludge as a raw material: Effect of inoculum and sludge solids concentration. Process Biochem. 2001, 37, 197–208. [Google Scholar] [CrossRef]
- Maheswaran, S.; Sreeramanan, S.; Josephine, C.M.R.; Marimuthu, K.; Xavier, R. Occurrence of Bacillus thuringiensis in faeces of herbivorous farm animals. Afr. J. Biotechnol. 2010, 9, 8013–8019. [Google Scholar]
- Maduell, P.; Armengol, G.; Llagostera, M.; Orduz, S.; Lindow, S. B. thuringiensis is a poor colonist of leaf surfaces. Microb. Ecol. 2008, 55, 212–219. [Google Scholar] [CrossRef]
- Prabhakar, A.; Bishop, A.H. Effect of Bacillus thuringiensis naturally colonising Brassica. campestris var. chinensis leaves on neonate larvae of Pieris. brassicae. J. Invertebr. Pathol. 2009, 100, 193–194. [Google Scholar] [CrossRef]
- Rodríguez-Sánchez, C.; Sittenfeld, A.; Janzen, D.H.; Espinoza, A.M. Bacillus thuringiensis in caterpillars and associated materials collected from protected tropical forests in northwestern Costa Rica. Rev. Biol. Trop. 2005, 54, 265–271. [Google Scholar]
- Bora, R.S.; Murty, M.G.; Shenbagarathai, R.; Sekar, V. Introduction of a Lepidopteran-specific insecticidal crystal protein gene of Bacillus thuringiensis subsp. kurstaki by conjugal transfer into a Bacillus megaterium strain that persists in the cotton phyllosphere. Appl. Environ. Microbiol. 1994, 60, 214–222. [Google Scholar]
- Devi, V.S.; Rao, P.A.; Sharma, S.P.; Sharma, H.C. Interaction of acid exudates in chickpea with biological activity of Bacillus thuringiensis towards Helicoverpa. armigera. J. Appl. Entomol. 2013. [Google Scholar] [CrossRef]
- Rabinovitch, L.; Fátima, C.; Cavados, G.; Chaves, J.Q.; Silva, K.R.A.; Seldin, L. A new strain of Bacillus thuringiensis serovar israelensis very active against Blackfly larvae. Mem. Inst. Oswaldo Cruz 1999, 94, 683–685. [Google Scholar] [CrossRef]
- Halverson, L.J.; Clayton, M.K.; Handelsman, J. Population biology of Bacillus cereus UW85 in the rhizosphere of field-grown soybeans. Soil Biol. Biochem. 1993, 25, 485–493. [Google Scholar] [CrossRef]
- Bisht, S.C.; Mishra, P.K. Ascending migration of endophytic Bacillus thuringiensis and assessment of benefits to different legumes of N.W. Himalayas. Eur. J. Soil Biol. 2013, 56, 56–64. [Google Scholar] [CrossRef]
- Subrahmanyan, P.; Reddy, M.N.; Rao, A.S. Exudation of certain organic compounds from seeds of groundnut. Seed Sci. Technol. 1983, 11, 267–272. [Google Scholar]
- Chanway, C.P. Bacterial endophytes: Ecological and practical implications. Sydowia 1998, 50, 149–170. [Google Scholar]
- Coombs, J.T.; Franco, C.M.M. Isolation and identification of Actinobacteria from surface-sterilized wheat roots. Appl. Environ. Microbiol. 2003, 69, 5603–5608. [Google Scholar] [CrossRef]
- Hallmann, J.; Quadt-Hallmann, A.; Mahaffee, W.F.; Kloepper, J.W. Bacterial endophytes in agricultural crops. Can. J. Microbiol. 1997, 43, 895–914. [Google Scholar] [CrossRef]
- Rosenblueth, M.; Martinez-Romero, E. Rhizobium. etli maize populations and their competitiveness for root colonization. Arch. Microbiol. 2004, 181, 337–344. [Google Scholar] [CrossRef]
- James, E.K.; Olivares, F.L.; de Oliveira, A.L.M.; dos Reis, F.B.; da Silva, L.G.; Reis, V.M. Further observations on the interaction between sugar cane and Gluconacetobacter diazotrophicus under laboratory and greenhouse conditions. J. Exp. Bot. 2001, 52, 747–760. [Google Scholar]
- Bacon, C. Endophytic and biological control potential of Bacillus mojavensis and related species. Biol. Contr. 2002, 23, 274–284. [Google Scholar] [CrossRef]
- Benhamou, N.; Kloepper, J.W.; Tuzun, S. Induction of resistance against Fusarium wilt of tomato by combination of chitosan with an endophytic bacterial strain: Ultrastructure and cytochemistry of the host response. Planta 1998, 204, 153–168. [Google Scholar] [CrossRef]
- Bent, E.; Chanway, C.P. The growth-promoting effects of a bacterial endophyte on lodgepole pine are partially inhibited by the presence of other rhizobacteria. Can. J. Microbiol. 1998, 44, 980–988. [Google Scholar] [CrossRef]
- Chanway, C.P. Inoculation of tree roots with plant growth promoting soil bacteria: An emerging technology for reforestation. For. Sci. 1997, 43, 99–112. [Google Scholar]
- Praça, L.B.; Gomes, A.C.M.M.; Cabral, G.; Martins, E.S.; Sujii, E.R.; Monnerat, R.G. Endophytic colonization by brazilian strains of Bacillus thuringiensis on cabbage seedlings grown in vitro. Bt Res. 2012, 3, 11–19. [Google Scholar]
- McInroy, J.A.; Kloepper, J.W. Population dynamics of endophytic bacteria in field-grown sweet corn and cotton. Can. J. Microbiol. 1995, 41, 895–901. [Google Scholar] [CrossRef]
- Suzuki, M.T.; Hernández-Rodríguez, C.S.; Araújo, W.L.; Ferré, J. Characterization of an endophytic Bacillus thuringiensis strain isolated from sugar cane. In Proceedings of 41st Annual Meeting of the Society for Invertebrate Pathology and 9th International Conference on Bacillus thuringiensis, University of Warwick, Coventry, UK, 3–7 August 2008.
- Mishra, P.K.; Mishra, S.; Selvakumar, G.; Bisht, J.K.; Kundu, S.; Gupta, H.S. Coinoculation of Bacillus thuringiensis-KR1 with Rhizobium. leguminosarum enhances plant growth and nodulation of pea (Pisum. sativum L.) and lentil (Lens culinaris L.). World J. Microbiol. Biotechnol. 2009, 25, 753–761. [Google Scholar] [CrossRef]
- Ornellas, R.M.S. Bioprospection of Rizobacteria for Beneficial Effects on ‘Yellow’ Passion Fruit (Passiflora Edulis) Seedlings. MSc Thesis, State University of Santa Cruz, Ilhéus-BA, Brazil, 2011. [Google Scholar]
- Da Silva, C.B. Holobionte Cacaueiro: Diversidade Genética da Porção Microbiana Associada a Frutos de Diferentes Clones de Theobroma cacao L. MSc Thesis, State University of Santa Cruz, Ilhéus-BA, Brazil, 2013. [Google Scholar]
- Estruch, J.J.; Warren, G.W.; Mullins, M.A.; Nye, G.J.; Craig, J.A.; Koziel, M.G. Vip3A, a novel Bacillus thuringiensis vegetative insecticidal protein with a wide spectrum of activities against lepidopteran insects. Proc. Natl. Acad. Sci. USA 1996, 93, 5389–5394. [Google Scholar] [CrossRef]
- McCully, E.M. Niches for bacterial endophytes in crop plants: A plant biologist’s view. Funct. Plant Biol. 2001, 28, 983–990. [Google Scholar] [CrossRef]
- Griego, V.M.; Spence, K.D. Inactivation of Bacillus thuringiensis spores by ultraviolet and visible light. Appl. Environ. Microbiol. 1978, 35, 906–910. [Google Scholar]
- Behle, R.; Mcguire, M.R.; Shasha, B.S. Effects of sunlight and simulated rain on residual activity of Bacillus thuringiensis formulations. J. Econ. Entomol. 1997, 90, 1560–1566. [Google Scholar]
- Ruan, L.; Yu, Z.; Fang, B.; He, W.; Wang, Y.; Shen, P. Melanin pigment formation and increased UV resistance in Bacillus thuringiensis following high temperature induction. Syst. Appl. Microbiol. 2004, 27, 286–289. [Google Scholar] [CrossRef]
- Stewart, C.N.; Adang, M.J.; All, J.N.; Boerma, H.R.; Cardineau, C.; Tucker, D.; Parrott, W.A. Genetic transformation, recovery, and characterization of fertile soybean transgenic for a synthetic Bacillus thuringiensis cry1Ac gene. Plant Physiol. 1996, 112, 121–129. [Google Scholar]
- Fischhoff, D.A.; Bowdish, K.S.; Perlak, F.J.; Marrone, P.G.; McCormick, S.M.; Niedermeyer, J.G.; Dean, D.A.; Kusano-Kretzmer, K.; Mayer, E.J.; Rochester, D.E.; et al. Insect tolerant transgenic tomato plants. Nat. Biotechnol. 1987, 5, 807–813. [Google Scholar] [CrossRef]
- Vaeck, M.; Reynaerts, A.; Hofte, H.; Jansens, S.; Debeuckeleer, M.; Dean, C.; Zabeau, M.; Vanmontagu, M.; Leemans, J. Transgenic plants protected from insect attack. Nature 1987, 328, 33–37. [Google Scholar] [CrossRef]
- Lampel, J.S.; Canter, G.L.; Dimock, M.B.; Kelly, J.L.; Anderson, J.J.; Uratani, B.B.; Foulke, J.S.; Turner, J.T. Integrative cloning, expression, and stability of the cryIA(c) gene from Bacillus thuringiensis subsp. kurstaki in a recombinant strain of Clavibacter xyli subsp. cynodontis. Appl. Environ. Microbiol. 1994, 60, 501–508. [Google Scholar]
- Skøt, L.; Timms, E.; Mytton, L. The effect of toxin-producing Rhizobium strains, on larvae of Sitona flavescens feeding on legume roots and nodules. Plant Soil 1994, 163, 141–150. [Google Scholar]
- Menon, A.S.; Mestral, J. Survival of Bacillus thuringiensis var. kurstaki in waters. Water Air Soil Pollut. 1985, 25, 265–274. [Google Scholar]
- De Amorim, G.V.; Whittome, B.; Shore, B.; Levin, D.B. Identification of Bacillus thuringiensis subsp. kurstaki strain HD1-Like bacteria from environmental and human samples after aerial spraying of Victoria, British Columbia, Canada, with foray 48B. Appl. Environ. Microbiol. 2001, 67, 1035–1043. [Google Scholar] [CrossRef]
- Boisvert, M.; Boisvert, J. Persistence of toxic activity and recycling of Bacillus thuringiensis var. israelensis in cold water: Field experiments using diffusion chambers in a pond. Biocontrol. Sci. Technol. 1999, 9, 507–522. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Su, T.; Mulla, M.S. Mosquito control and bacterial flora in water enriched with organic matter and treated with Bacillus thuringiensis subsp. israelensis and Bacillus sphaericus formulations. J. Vector Ecol. 1999, 24, 138–153. [Google Scholar]
- Manasherob, R.; Ben-Dov, E.; Zaritsky, A.; Barak, Z. Germination, growth, and sporulation of Bacillus thuringiensis subsp. israelensis in excreted food vacuoles of the protozoan Tetrahymena pyriformis. Appl. Environ. Microbiol. 1998, 64, 1750–1758. [Google Scholar]
- Kweon, C.; Choi, S.; Kwon, H.; Kim, E.; Kang, H.; Moon, J.; Jang, G.; Lee, H.; Kang, S.; Kim, J.; et al. Isolation, characterization, and evaluation of Bacillus thuringiensis isolated from cow milk. Korean J. Vet. Res. 2012, 52, 169–176. [Google Scholar]
- Swiecicka, I.; Fiedoruk, K.; Bednarz, G. The occurrence and properties of Bacillus thuringiensis isolated from free-living animals. Lett. Appl. Microbiol. 2002, 34, 194–198. [Google Scholar] [CrossRef]
- Peterson, J.W. Bacterial pathogenesis. In Medical Microbiology; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Rohmer, L.; Hocquet, D.; Miller, M.I. Are pathogenic bacteria just looking for food? Metabolism and microbial pathogenesis. Trends Ecol. Evol. 2012, 19, 341–348. [Google Scholar]
- Tanada, Y.; Kaya, H.K. Insect Pathology; Academic Press: San Diego, CA, USA, 1993. [Google Scholar]
- Johnston, P.R.; Crickmore, N. Gut bacteria not required for Bacillus thuringiensis insecticidal activity towards the tobacco hornworm, Manduca sexta. Appl. Environ. Microbiol. 2009, 75, 5094–5099. [Google Scholar] [CrossRef]
- Cerstiaens, A.; Verleyen, P.; Rie, J.V.A.N.; Kerkhove, E.V.A.N.; Schwartz, J.; Laprade, R.; Loof, A.D.E.; Schoofs, L. Effect of Bacillus thuringiensis Cry1 toxins in insect hemolymph and their neurotoxicity in brain cells of Lymantria dispar. Appl. Environ. Microbiol. 2001, 67, 3923–3927. [Google Scholar]
- Argôlo Filho, R.C.; Gomes, R.A.; Barreto, M.R.; Lana, U.G.P.; Valicente, F.H.; Loguercio, L.L. Growth variation among Bacillus thuringiensis strains can affect screening procedures for supernatant-secreted toxins against insect pests. Pest Manag. Sci. 2011, 67, 1184–1192. [Google Scholar]
- Loguercio, L.L.; Santos, C.G.; Barreto, M.R.; Guimaraes, C.T.; Paiva, E. Association of PCR and feeding bioassays as a large-scale method to screen tropical Bacillus thuringiensis isolates for a cry constitution with higher insecticidal effect against Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae. Lett. Appl. Microbiol. 2001, 32, 362–367. [Google Scholar] [CrossRef]
- Loguercio, L.L.; Barreto, M.L.; Rocha, T.L.; Santos, C.G.; Teixeira, F.F.; Paiva, E. Combined analysis of supernatant-based feeding bioassays and PCR as a first-tier screening strategy for Vip-derived activities in Bacillus thuringiensis strains effective against tropical fall armyworm. J. Appl. Microbiol. 2002, 93, 269–277. [Google Scholar] [CrossRef]
- Ohba, M.; Wasano, N.; Mizuki, E. Bacillus thuringiensis soil populations naturally occurring in the Ryukyus, a subtropic region of Japan. Microbiol. Res. 2000, 155, 17–22. [Google Scholar] [CrossRef]
- Vallet-gely, I.; Lemaitre, B.; Boccard, F. Bacterial strategies to overcome insect defences. Nat. Rev. Microbiol. 2008, 6, 302–313. [Google Scholar] [CrossRef]
- Fedhila, S.; Guillemet, E.; Nel, P.; Lereclus, D. Characterization of two Bacillus thuringiensis genes identified by in vivo screening of virulence factors. J. Bacteriol. 2004, 70, 4784–4791. [Google Scholar]
- Dalhammar, G.; Steiner, H. Characterization of inhibitor A, a protease from Bacillus thuringiensis which degrades attacins and cecropins, two classes of antibacterial proteins in insects. FEBS J. 1984, 139, 247–252. [Google Scholar]
- Raymond, B.; Lijek, R.S.; Griffiths, R.I.; Bonsall, M.B. Ecological consequences of ingestion of Bacillus cereus on Bacillus thuringiensis infections and on the gut flora of a lepidopteran host. J. Invertebr. Pathol. 2008, 99, 103–111. [Google Scholar] [CrossRef]
- Cherif, A.; Rezgui, W.; Raddadi, N.; Daffonchio, D.; Boudabous, A. Characterization and partial purification of entomocin 110, a newly identified bacteriocin from Bacillus thuringiensis subsp. entomocidus HD110. Microbiol. Res. 2008, 163, 684–692. [Google Scholar]
- Wabiko, H.; Yasuda, E. Bacillus thuringiensis protoxin: Location of toxic border and requirement of non-toxic domain for high-level in vivo production of active toxin. Microbiology 1995, 141, 629–639. [Google Scholar] [CrossRef]
- Volwerk, J.J.; Koke, J.A.; Wetherwax, P.B.; Griffith, O.H. Functional characteristics of phosphatidylinositol-specific phospholipases C from Bacillus cereus and Bacillus thuringiensis. FEMS Microbiol. Lett. 1989, 52, 237–241. [Google Scholar]
- Sampson, M.N.; Gooday, G.W. Involvement of chitinases of Bacillus thuringiensis during pathogenesis in insects. Microbiology 1998, 144, 2189–2194. [Google Scholar] [CrossRef]
- Fedhila, S.; Nel, P.; Lereclus, D. The InhA2 metalloprotease of Bacillus thuringiensis strain 407 is required for pathogenicity in insects infected via the oral route. J. Bacteriol. 2002, 184, 3296–3304. [Google Scholar] [CrossRef]
- Fortier, M.; Vachon, V.; Frutos, R.; Schwartz, J.-L.; Laprade, R. Effect of insect larval midgut proteases on the activity of Bacillus thuringiensis Cry toxins. Appl. Environ. Microbiol. 2007, 73, 6208–6213. [Google Scholar] [CrossRef]
- Tran, S.-L.; Guillemet, E.; Lereclus, D.; Ramarao, N. Iron regulates Bacillus thuringiensis haemolysin hlyII gene expression during insect infection. J. Invertebr. Pathol. 2013, 113, 205–208. [Google Scholar]
- Luo, X.; Chen, L.; Huang, Q.; Zheng, J.; Zhou, W.; Peng, D.; Ruan, L.; Sun, M. Bacillus thuringiensis metalloproteinase Bmp1 functions as a nematicidal virulence factor. Appl. Environ. Microbiol. 2013, 79, 460–468. [Google Scholar] [CrossRef]
- Ramarao, N.; Lereclus, D. The InhA1 metalloprotease allows spores of the B. cereus group to escape macrophages. Cell. Microbiol. 2005, 7, 1357–1364. [Google Scholar] [CrossRef]
- Dillon, R.J.; Dillon, V.M. The gut bacteria of insects: Nonpathogenic interactions. Annu. Rev. Entomol. 2004, 49, 71–92. [Google Scholar] [CrossRef]
- Wells, E.V.; Boulton, M.; Hall, W.; Bidol, S.A. Reptile-associated salmonellosis in preschool-aged children in Michigan, January 2001–June 2003. Clin. Infect. Dis. 2004, 39, 687–691. [Google Scholar] [CrossRef]
- Broderick, N.; Raffa, K.F.; Handelsman, J. Midgut bacteria required for Bacillus thuringiensis insecticidal activity. Proc. Natl. Acad. Sci. USA 2006, 103, 15196–15199. [Google Scholar] [CrossRef]
- Broderick, N.; Robinson, C.J.; McMahon, M.D.; Holt, J.; Handelsman, J.; Raffa, K.F. Contributions of gut bacteria to Bacillus thuringiensis-induced mortality vary across a range of Lepidoptera. BMC Biol. 2009, 7, 1–9. [Google Scholar] [CrossRef]
- Jarosz, J. Gut flora of Galleria mellonella suppressing ingested bacteria. J. Invertebr. Pathol. 1979, 34, 192–198. [Google Scholar] [CrossRef]
- Raymond, B.; Elliot, S.L.; Ellis, R.J. Quantifying the reproduction of Bacillus thuringiensis HD1 in cadavers and live larvae of Plutella xylostella. J. Invertebr. Pathol. 2008, 98, 307–313. [Google Scholar] [CrossRef]
- Guerchicoff, A.; Dele, A. The Bacillus thuringiensis cyt genes for hemolytic endotoxins constitute a gene family. Appl. Environ. Microbiol. 2001, 67, 1090–1096. [Google Scholar] [CrossRef]
- Bravo, A.; Gill, S.S.; Soberón, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar] [CrossRef]
- Kostichka, K.; Warren, G.W.; Mullins, M.; Mullins, A.D.; Palekar, N.V.; Craig, J.A.; Koziel, M.G.; Estruch, J.J. Cloning of a cryV-type insecticidal protein gene from Bacillus thuringiensis: The CryV-encoded protein is expressed early in stationary phase. J. Bacteriol. 1996, 178, 2141–2144. [Google Scholar]
- Song, F.; Zhang, J.; Gu, A.; Wu, Y.; Han, L.; He, K.; Chen, Z.; Yao, J.; Hu, Y.; Li, G.; et al. Identification of cry1I-type genes from Bacillus thuringiensis strains and characterization of a novel cry1I-type gene. Appl. Environ. Microbiol. 2003, 69, 5207–5211. [Google Scholar] [CrossRef]
- Li, J.D.; Carroll, J.; Ellar, D.J. Crystal structure of insecticidal delta-endotoxin from Bacillus thuringiensis at 2.5 A resolution. Nature 1991, 353, 815–821. [Google Scholar] [CrossRef]
- Vachon, V.; Laprade, R.; Schwartz, J. Current models of the mode of action of Bacillus thuringiensis insecticidal crystal proteins: A critical review. J. Invertebr. Pathol. 2012, 111, 1–12. [Google Scholar] [CrossRef]
- Koller, C.N.; Bauer, L.S.; Hollingworth, R.M. Characterization of the pH-mediated solubility of Bacillus thuringiensis var. san diego native delta-endotoxin crystals. Biochem. Biophys. Res. Commun. 1992, 184, 692–699. [Google Scholar] [CrossRef]
- Soberón, M.; Gill, S.S.; Bravo, A. Signaling versus punching hole: How do Bacillus thuringiensis toxins kill insect midgut cells? Cell. Mol. Life Sci. 2009, 66, 1337–1349. [Google Scholar] [CrossRef]
- Zhang, X.; Candas, M.; Griko, N.B. Cytotoxicity of Bacillus thuringiensis Cry1Ab toxin depends on specific binding of the toxin to the cadherin receptor BT-R1 expressed in insect cells. Cell Death Differ. 2005, 12, 1407–1416. [Google Scholar] [CrossRef]
- Zhang, X.; Candas, M.; Griko, N.B.; Taussig, R.; Bulla, L.A. A mechanism of cell death involving an adenylyl cyclase PKA signali ng pathway is induced by the Cry1Ab toxin of Bacillus thuringiensis. Proc. Natl. Acad. Sci. USA 2006, 103, 9897–9902. [Google Scholar]
- Glare, T.R.; O’Callaghan, M. Bacillus thuringiensis: Biology, Ecology and Safety; John Wiley: Chichester, UK, 2000. [Google Scholar]
- Butko, P. Cytolytic toxin Cyt1A and its mechanism of membrane damage: Data and hypotheses. Appl. Environ. Microbiol. 2003, 69, 2415–2422. [Google Scholar] [CrossRef]
- Parker, M.W.; Feil, S.C. Pore-forming protein toxins: From structure to function. Prog. Biophys. Mol. Biol. 2005, 88, 91–142. [Google Scholar] [CrossRef]
- Sayyed, A.L.I.H.; Crickmore, N.; Wright, D.J. Cyt1Aa from Bacillus thuringiensis subsp. israelensis is toxic to the diamondback moth, Plutella xylostella, and synergizes the activity of Cry1Ac towards a resistant strain. Appl. Environ. Microbiol. 2001, 67, 5859–5861. [Google Scholar] [CrossRef]
- Oestergaard, J.; Ehlers, R.; Mart, A.C.; Real, M.D. Binding of Cyt1Aa and Cry11Aa toxins of Bacillus thuringiensis serovar israelensis to brush border membrane vesicles of Tipula paludosa (Diptera: Nematocera) and subsequent pore formation. Appl. Environ. Microbiol. 2007, 73, 3623–3629. [Google Scholar] [CrossRef]
- Pérez, C.; Fernandez, L.E.; Sun, J.; Folch, J.L.; Gill, S.S.; Soberón, M.; Bravo, A. Bacillus thuringiensis subsp. israelensis Cyt1Aa synergizes Cry11Aa toxin by functioning as a membrane-bound receptor. Proc. Natl. Acad. Sci. USA 2005, 102, 18303–18308. [Google Scholar] [CrossRef]
- Rincón-Castro, M.C.; Barajas-Huerta, J.; Ibarra, J.E. Antagonism between Cry1Ac1 and Cyt1A1 toxins of Bacillus thuringiensis. Appl. Environ. Microbiol. 1999, 65, 2049–2053. [Google Scholar]
- Yu, C.G.; Mullins, M.A.; Warren, G.W.; Koziel, M.G.; Estruch, J.J. The Bacillus thuringiensis vegetative insecticidal protein Vip3A lyses midgut epithelium cells of susceptible insects. Appl. Environ. Microbiol. 1997, 63, 532–536. [Google Scholar]
- Rice, W.C. Specific primers for the detection of vip3A insecticidal gene within a Bacillus thuringiensis collection. Lett. Appl. Microbiol. 1999, 28, 378–382. [Google Scholar] [CrossRef]
- Lee, M.K.; Walters, F.S.; Hart, H.; Palekar, N.; Chen, J. The Mode of Action of the Bacillus thuringiensis Vegetative Insecticidal Protein Vip3A differs from that of Cry1Ab d-endotoxin. Appl. Environ. Microbiol. 2003, 69, 4648–4657. [Google Scholar] [CrossRef]
- Hernández-Rodríguez, C.S.; Boets, A.; van Rie, J.; Ferré, J. Screening and identification of vip genes in Bacillus thuringiensis strains. J. Appl. Microbiol. 2009, 107, 219–225. [Google Scholar] [CrossRef]
- Crickmore, N.; Baum, J.; Bravo, A.; Lereclus, D.; Narva, K.; Sampson, K.; Schnepf, E.; Sun, M.; Zeigler, D.R. Bacillus thuringiensis toxin nomenclature. Available online: http://www.btnomenclature.info/ (accessed on 20 October 2013).
- Warren, W. Vegetative insecticidal proteins: Novel proteins for control of corn pests. In Advances in Insect Control, the Role of Transgenic Plants; Carozzi, N.B., Koziel, M., Eds.; Taylors & Francis Ltd.: London, UK, 1997. [Google Scholar]
- Barth, H.; Aktories, K.; Popoff, M.R.; Stiles, B.G. Binary bacterial toxins: Biochemistry, biology, and applications of common Clostridium and Bacillus proteins. Microbiol. Mol. Biol. Rev. 2004, 68, 373–402. [Google Scholar] [CrossRef]
- Peng, D.; Chen, S.; Ruan, L.; Li, L.; Yu, Z.; Sun, M. Safety assessment of transgenic Bacillus thuringiensis with Vip insecticidal protein gene by feeding studies. Food Chem. Toxicol. 2007, 45, 1179–1185. [Google Scholar] [CrossRef]
- Callegan, M.C.; Cochran, D.C.; Kane, S.T.; Gilmore, M.S.; Gominet, M.; Lereclus, D. Contribution of membrane-damaging toxins to Bacillus endophthalmitis pathogenesis. Infect. Immun. 2002, 70, 5381–5389. [Google Scholar] [CrossRef]
- Ikezawa, H.; Nakabayashi, T.; Suzuki, K.; Nakajima, M.; Taguchi, T.; Taguchi, R. Complete purification of phosphatidylinositol-specific phospholipase C from a strain of Bacillus thuringiensis. J. Biochem. 1983, 93, 1717–1719. [Google Scholar]
- Hergenrother, P.J.; Martin, S.F. Determination of the kinetic parameters for phospholipase C (Bacillus cereus) on different phospholipid substrates using a chromogenic assay based on the quantitation of inorganic phosphate. Anal. Biochem. 1997, 251, 45–49. [Google Scholar] [CrossRef]
- Krieg, A. Concerning alpha-exotoxin produced by vegetative cells of Bacillus thuringiensis and Bacillus cereus. J. Invertebr. Pathol. 1971, 1, 134–135. [Google Scholar] [CrossRef]
- Faust, R.M.; Bulla, A.L., Jr. Bacterial and their toxins as insecticides. In Microbial and Viral Pesticides; Kurstaki, E., Ed.; Marcel Dekker Inc.: Nova York, NY, USA, 1982. [Google Scholar]
- Arora, N.; Ahmad, T.; Rajagopal, R.; Bhatnagar, R.K. A constitutively expressed 36 kDa exochitinase from Bacillus thuringiensis HD-1. Biochem. Biophys. Res. Commun. 2003, 307, 620–625. [Google Scholar] [CrossRef]
- Regev, A.; Keller, M.; Strizhov, N.; Sneh, B.; Prudovsky, E.; Chet, I.; Ginzberg, I.; Koncz-Kalman, Z.; Koncz, C.; Schell, J.; et al. Synergistic activity of a Bacillus thuringiensis delta-endotoxin and a bacterial endochitinase against Spodoptera littoralis larvae. Appl. Environ. Microbiol. 1996, 62, 3581–3586. [Google Scholar]
- Tantimavanich, S.; Pantuwatana, S.; Bhumiratana, A.; Panbangred, W. Cloning of a chitinase gene into Bacillus thuringiensis subsp. aizawai for enhanced insecticidal activity. J. Gen. Appl. Microbiol. 1997, 347, 341–347. [Google Scholar]
- Li, E.; Yousten, A.A. Metalloprotease from Bacillus thuringiensis. Appl. Microbiol. 1975, 30, 354–361. [Google Scholar]
- Andrews, R.E.; Bibilos, M.M.; Bulla, L.A. Protease activation of the entomocidal protoxin of Bacillus thuringiensis subsp. kurstaki. Appl. Environ. Microbiol. 1985, 50, 737–742. [Google Scholar]
- Guillemet, E.; Cadot, C.; Tran, S.-L.; Guinebretière, M.-H.; Lereclus, D.; Ramarao, N. The InhA Metalloproteases of Bacillus cereus contribute concomitantly to virulence. J. Bacteriol. 2010, 192, 286–294. [Google Scholar] [CrossRef]
- Donovan, W.P.; Engleman, J.T.; Donovan, J.C.; Baum, J.A.; Bunkers, G.J.; Chi, D.J.; Clinton, W.P.; English, L.; Heck, G.R.; Ilagan, O.M.; et al. Discovery and characterization of Sip1A: A novel secreted protein from Bacillus thuringiensis with activity against coleopteran larvae. Appl. Microbiol. Biot. 2006, 72, 713–719. [Google Scholar] [CrossRef]
- Farkas, J.; Sebesta, K.; Horská, K.; Samek, K.; Dolejs, L.; Sorm, F. The structure of exotoxin of Bacillus thuringiensis var. gelechiae. Collect. Czech. Chem. Commun. 1968, 34, 1118–1120. [Google Scholar]
- Beebee, T.; Korner, A.; Bond, R.P.M. Differential inhibition of mammalian ribonucleic acid polymerases by an exotoxin from Bacillus thuringiensis. The direct observation of nucleoplasmic ribonucleic acid polymerase activity in intact nuclei. Biochem. J. 1972, 127, 619–624. [Google Scholar]
- World Health Organization-WHO. Microbial Pest Control Agent: Bacillus thuringiensis; WHO: Geneva, Switzerland, 1999; p. 125. [Google Scholar]
- Calberg, G. Bacillus thuringiensis and microbial control of flies. MIRCEN J. Appl. Microb. Biotechnol. 1986, 2, 267–274. [Google Scholar] [CrossRef]
- Ohba, M.; Tantichodok, A.; Aizawa, K. Production of heat-stable exotoxin by Bacillus thuringiensis and related bacteria. J. Invertebr. Pathol. 1981, 38, 26–32. [Google Scholar] [CrossRef]
- Levinson, B.L.; Kasyan, K.J.; Chiu, S.U.E.S.; Currier, T.C.; González, J.M., Jr. Identification of b-exotoxin production, plasmid encoding, and a new exotoxin in Bacillus thuringiensis by using high-performance liquid chromatograpy. J. Bacteriol. 1990, 172, 3172–3179. [Google Scholar]
- Domingos, J.B.; Longhinotti, E.; Gageiro, V.; Nome, F. A química dos ésteres de fosfato. Quim. Nova 2003, 26, 745–753. [Google Scholar] [CrossRef]
- Espinasse, S.; Gohar, M.; Lereclus, D.; Sanchis, V. An extracytoplasmic-function sigma factor is involved in a pathway controlling b-exotoxin I production in Bacillus thuringiensis subsp. thuringiensis strain 407–1. J. Bacteriol. 2004, 186, 3108–3116. [Google Scholar] [CrossRef]
- Hassanain, M.A.; el Garhy, M.F.; Abdel-Ghaffar, F.A.; el-Sharaby, A.; Abdel Megeed, K.N. Biological control studies of soft and hard ticks in Egypt: The effect of Bacillus thuringiensis varieties on soft and hard ticks (ixodidae). Parasitol. Res. 1997, 83, 209–213. [Google Scholar] [CrossRef]
- Zhioua, E.; Heyer, K.; Browning, M.; Ginsberg, H.S.; LeBrun, R.A. Pathogenicity of Bacillus thuringiensis variety kurstaki to Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 1999, 36, 900–902. [Google Scholar]
- El-Kelesh, E.A.M.; El-Refaii, M.A.H. Insecticidal effect of Bacillus thuringiensis var. kurstaki against Hyalomma dromedarii on experimentally infested rabbits. Egypt. J. Agric. Res. 2006, 83, 993. [Google Scholar]
- Fernández-Ruvalcaba, M.; Peña-Chora, G.; Romo-Martínez, A.; Hernández-Velázquez, V.; de La Parra, A.B.; de La Rosa, D.P. Evaluation of Bacillus thuringiensis pathogenicity for a strain of the tick, Rhipicephalus microplus, resistant to chemical pesticides. J. Insect Sci. 2010, 10, 1–6. [Google Scholar]
- Habeeb, S.M.; El-hag, H.A.A. Ultrastructural changes in hemocyte cells of hard tick (Hyalomma dromedarii: Ixodidae): A model of Bacillus thuringiensis var. thuringiensis H14 d-endotoxin mode of action. Am. Euras. J. Agric. Environ. Sci. 2008, 3, 829–836. [Google Scholar]
- Erban, T.; Nesvorna, M.; Erbanova, M.; Hubert, J. Bacillus thuringiensis var. tenebrionis control of synanthropic mites (Acari: Acaridida) under laboratory conditions. Exp. Appl. Acarol. 2009, 49, 339–346. [Google Scholar] [CrossRef]
- Payne, J.; Cannon, R.J.C.; Ralph, A.L. Bacillus thuringiensis Isolates for Controlling Acarides. US Patent 5350576A, 27 September 1994. [Google Scholar]
- Wei, J.-Z.; Hale, K.; Carta, L.; Platzer, E.; Wong, C.; Fang, S.-C.; Aroian, R.V. Bacillus thuringiensis crystal proteins that target nematodes. Proc. Natl. Acad. Sci. USA 2003, 100, 2760–2765. [Google Scholar] [CrossRef]
- Mohammed, S.H.; El Saedy, M.A.E.; Enan, M.R.; Ibrahim, N.E.; Ghareeb, A.; Moustafa, A. Biocontrol efficiency of Bacillus thuringiensis toxins against root-knot nematode, Meloidogyne incognita. J. Cell Mol. Biol. 2008, 7, 57–66. [Google Scholar]
- Thompson, M.; Gaertner, F.H. Bacillus thuringiensis Isolate having Anti-Protozoan Activity. European Patent 461799A2, 28 December 1991. [Google Scholar]
- Kondo, S.; Mizuki, E.; Akao, T.; Ohba, M. Antitrichomonal strains of Bacillus thuringiensis. Parasitol. Res. 2002, 88, 1090–1092. [Google Scholar] [CrossRef]
- Mahillon, J.; Rezsöhazy, R.; Hallet, B.; Delcour, J. IS231 and other Bacillus thuringiensis transposable elements: A review. Genetica 1994, 93, 13–26. [Google Scholar] [CrossRef]
- Weisdorf, J.L. From foraging to farming: Explaining the neolithic revolution. J. Econ. Surv. 2005, 19, 561–586. [Google Scholar] [CrossRef]
- Loguercio, L.L.; Carneiro, N.P.; Carneiro, A.A. Milho Bt: Alternativa biotecnológica para controle biológico de insetos-praga. (in portuguese). Biotecnolog. Ciênc. Desenvolv. 2002, 24, 46–52. [Google Scholar]
- Bowden, S.E.; Drake, J.M. Ecology of multi-host pathogens of animals. Nat. Educ. Knowl. 2013, 4, 5. [Google Scholar]
- Pepin, K.M.; Lass, S.; Pulliam, J.R.C.; Read, A.F.; Lloyd-Smith, J.O. Identifying genetic markers of adaptation for surveillance of viral host jumps. Nat. Rev. Microbiol. 2010, 8, 802–813. [Google Scholar] [CrossRef]
- Garamszegi, L.Z. Patterns of co-speciation and host switching in primate malaria parasites. Malar. J. 2009, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Peeler, E.J.; Oidtmann, B.C.; Midtlyng, P.J.; Miossec, L.; Gozlan, R.E. Non-native aquatic animals introductions have driven disease emergence in Europe. Biol. Invasions 2011, 13, 1291–1303. [Google Scholar] [CrossRef]
- Mizuki, E.; Park, Y.S.; Saitoh, H.; Yamashita, S.; Akao, T.; Higuchi, K.; Ohba, M. Parasporin, a human leukemic cell-recognizing parasporal protein of Bacillus thuringiensis. Clin. Diagn. Lab. Immun. 2000, 7, 625–634. [Google Scholar]
- Mizuki, E.; Ohba, M.; Akao, T.; Yamashita, S.; Saitoh, H.; Park, Y.S. Unique activity associated with non-insecticidal Bacillus thuringiensis parasporal inclusions: In vitro cell-killing action on human cancer cells. J. Appl. Microbiol. 1999, 86, 477–486. [Google Scholar] [CrossRef]
- Ito, A.; Sasaguri, Y.; Kitada, S.; Kusaka, Y.; Kuwano, K.; Masutomi, K.; Mizuki, E.; Akao, T.; Ohba, M. A Bacillus thuringiensis crystal protein with selective cytocidal action to human cells. J. Biol. Chem. 2004, 279, 21282–21286. [Google Scholar] [CrossRef]
- Katayama, H.; Kusaka, Y.; Yokota, H.; Akao, T.; Kojima, M.; Nakamura, O.; Mekada, E.; Mizuki, E. Parasporin-1, a novel cytotoxic protein from Bacillus thuringiensis, induces Ca2+ influx and a sustained elevation of the cytoplasmic Ca2+ concentration in toxin-sensitive cells. J. Biol. Chem. 2007, 282, 7742–7752. [Google Scholar]
- Yamashita, S.; Katayama, H.; Saitoh, H.; Akao, T.; Park, Y.S.; Mizuki, E.; Ohba, M.; Ito, A. Typical three-domain cry proteins of Bacillus thuringiensis strain A1462 exhibit cytocidal activity on limited human cancer cells. J. Biochem. 2005, 138, 663–672. [Google Scholar] [CrossRef]
- Okumura, S.; Saitoh, H.; Ishikawa, T.; Wasano, N.; Yamashita, S.; Kusumoto, K.-I.; Akao, T.; Mizuki, E.; Ohba, M.; Inouye, K. Identification of a novel cytotoxic protein, Cry45Aa, from Bacillus thuringiensis A1470 and its selective cytotoxic activity against various mammalian cell lines. J. Agric. Food Chem. 2005, 53, 6313–6318. [Google Scholar] [CrossRef]
- Kitada, S.; Abe, Y.; Shimada, H.; Kusaka, Y.; Matsuo, Y.; Katayama, H.; Okumura, S.; Akao, T.; Mizuki, E.; Kuge, O.; et al. Cytocidal actions of parasporin-2, an anti-tumor crystal toxin from Bacillus thuringiensis. J. Biol. Chem. 2006, 281, 26350–26360. [Google Scholar] [CrossRef]
- Abe, Y.; Shimada, H.; Kitada, S. Raft-targeting and oligomerization of Parasporin-2, a Bacillus thuringiensis crystal protein with anti-tumour activity. J. Biochem. 2008, 143, 269–275. [Google Scholar]
- Fivaz, M.; Abrami, L.; Tsitrin, Y.; van der Goot, F.G. Aerolysin from Aeromonas. hydrophila and related toxins. Curr. Top. Microbiol. Immunol. 2001, 257, 35–52. [Google Scholar]
- Galinier, R.; Portela, J.; Moné, Y.; Allienne, J.F.; Henri, H.; Delbecq, S.; Mitta, G.; Gourbal, B.; Duval, D. Biomphalysin, a new β pore-forming toxin involved in Biomphalaria glabrata immune defense against Schistosoma mansoni. PLoS Pathog. 2013, 9, e1003216. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Argôlo-Filho, R.C.; Loguercio, L.L. Bacillus thuringiensis Is an Environmental Pathogen and Host-Specificity Has Developed as an Adaptation to Human-Generated Ecological Niches. Insects 2014, 5, 62-91. https://doi.org/10.3390/insects5010062
Argôlo-Filho RC, Loguercio LL. Bacillus thuringiensis Is an Environmental Pathogen and Host-Specificity Has Developed as an Adaptation to Human-Generated Ecological Niches. Insects. 2014; 5(1):62-91. https://doi.org/10.3390/insects5010062
Chicago/Turabian StyleArgôlo-Filho, Ronaldo Costa, and Leandro Lopes Loguercio. 2014. "Bacillus thuringiensis Is an Environmental Pathogen and Host-Specificity Has Developed as an Adaptation to Human-Generated Ecological Niches" Insects 5, no. 1: 62-91. https://doi.org/10.3390/insects5010062
APA StyleArgôlo-Filho, R. C., & Loguercio, L. L. (2014). Bacillus thuringiensis Is an Environmental Pathogen and Host-Specificity Has Developed as an Adaptation to Human-Generated Ecological Niches. Insects, 5(1), 62-91. https://doi.org/10.3390/insects5010062