Screening of Repellents against Vespid Wasps

Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Wasps, and Samples

{kind=link}
{kind=link}
{kind=link}
| Group | Date (d/m/y) a | Locality b | Remark | V.v. | V.g. | P.d. c | US d |
|---|---|---|---|---|---|---|---|
| 01 | 10/08/2011 | Vlezenbeek | In and around a garbage container | 16 | 29 | ||
| 02 | 16–17/08/2011 | Vlezenbeek | As Group 01 | 6 | 30 | ||
| 03 | 23/08/2011 | Vlezenbeek | As Group 01 | 2 | 30 | 1 (+1) | |
| 04 | 31/08/2011 | Ortho | From a nest at the base of a spruce trunk | 32 | 2 | ||
| 05 | 10–12/09/2011 | Vlezenbeek and Uccle | As Group 01, and from wasp traps placed in a garden | 6 | 3 | 10 (+3) | |
| 06 | 16–18/07/2012 | Vlezenbeek | From a nest at the base of a concrete wall | 49 | |||
| 07 | 24–26/07/2012 | Vlezenbeek | As Group 06 | 37 | |||
| 08 | 31/07–02/08/2012 | Vlezenbeek and Uccle | As Group 06, and from wasp traps placed in a garden | 26 | |||
| 09 | 04–12/08/2012 | Vlezenbeek and Uccle | As Group 08 | 21 | 1 | 2 | 1 |
| 10 | 14/08/2012 | Geraardsbergen | From trash cans in a park | 26 | 4 | ||
| 11 | 20/08/2012 | Geraardsbergen | As Group 10 | 31 | 7 | ||
| 12 | 27/08/2012 | Geraardsbergen | As Group 10 | 40 | 2 |
2.2. Experimental Setup
2.3. Bioassay

2.4. Screening of Volatiles
2.5. Dilutions and Mixtures of Selected Samples
2.6. Statistics and Calculations
3. Results
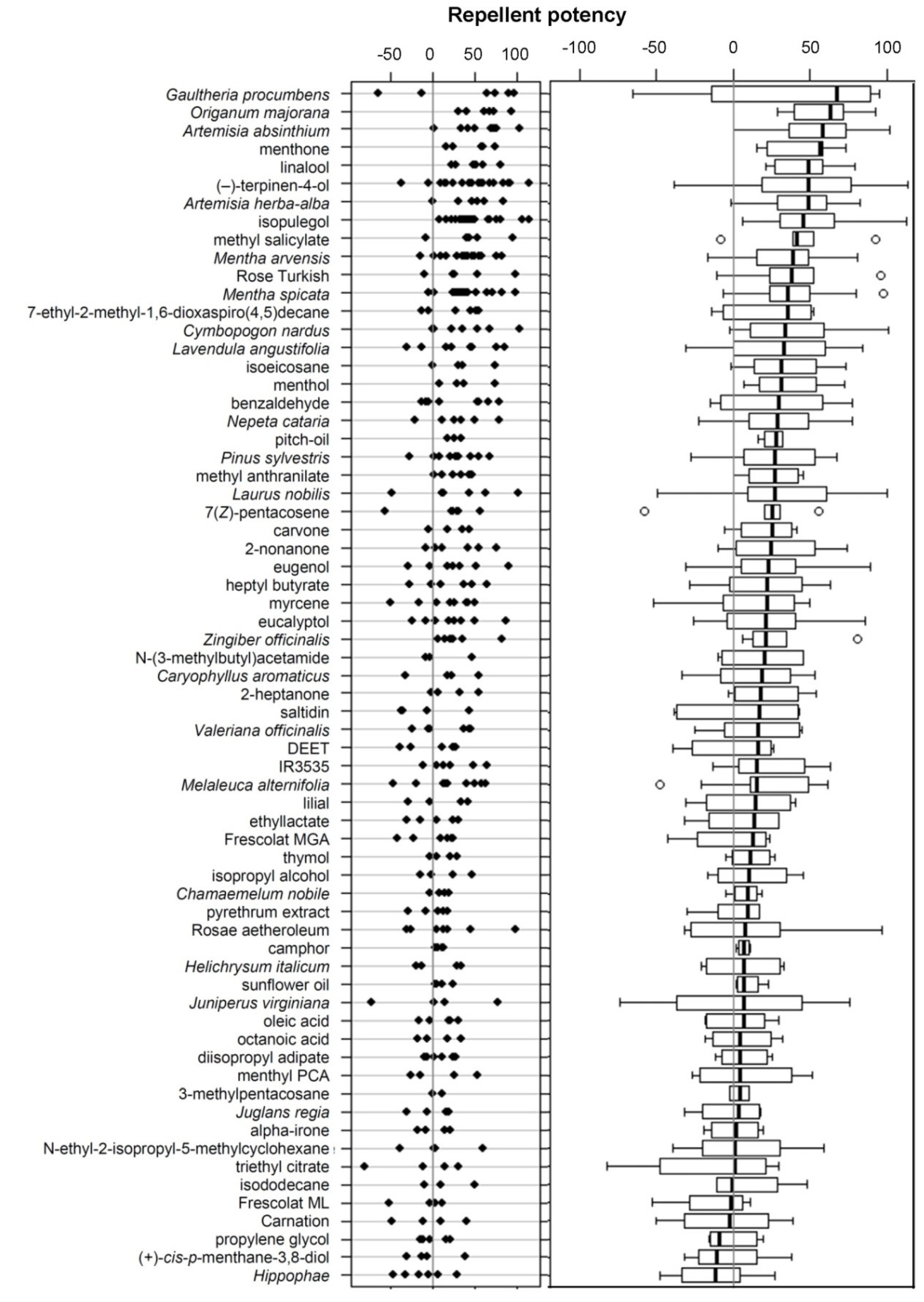
3.1. Repellency of Screened Samples
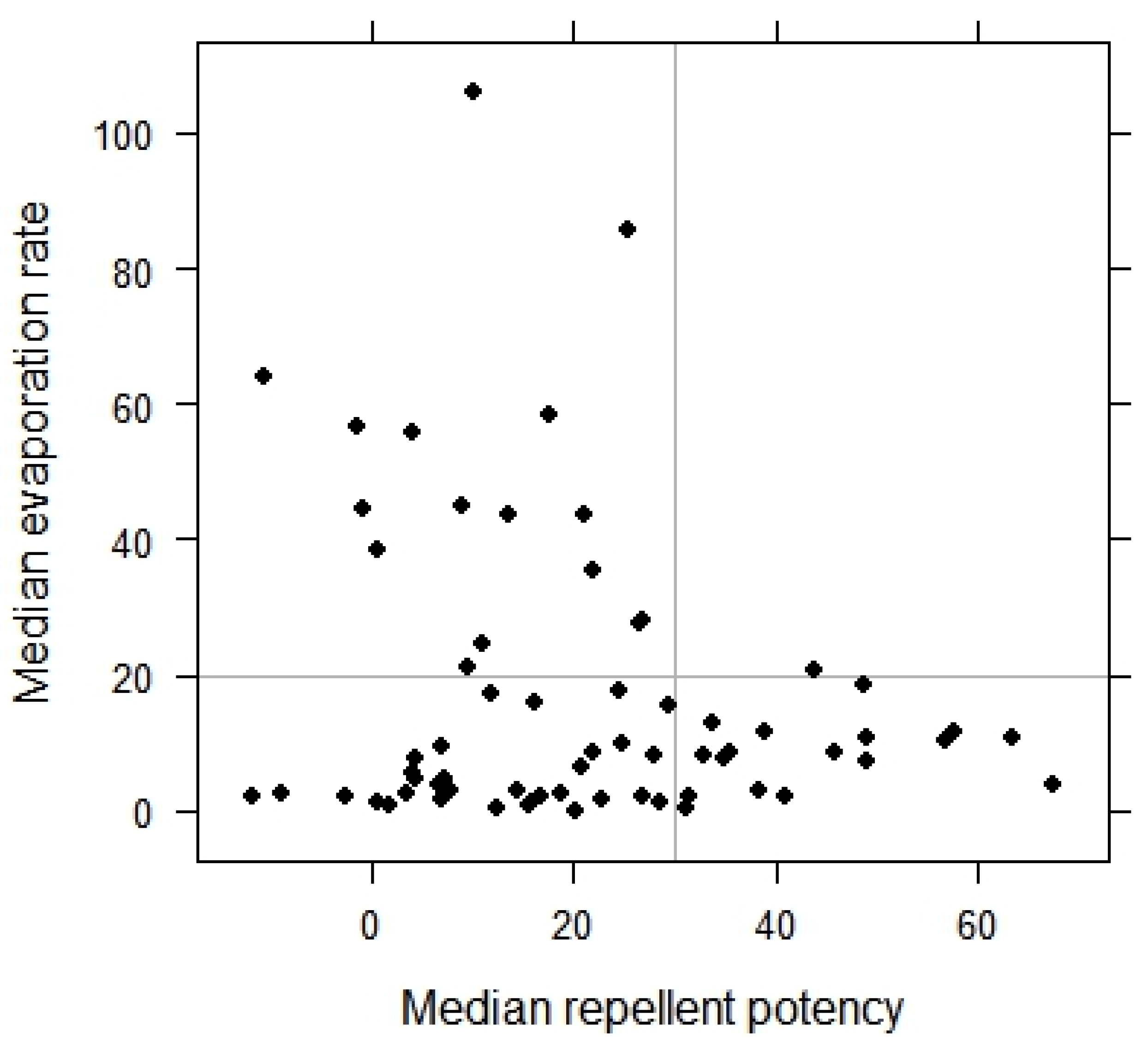
3.2. Repellency and Evaporation Rates
3.3. Repellency of Dilutions and Mixtures


| Essential oils and pure chemicals | Concentration b | ||||
|---|---|---|---|---|---|
| 0.00 | −0.25 | −0.50 | −0.75 | −1.00 | |
| Mentha spicata | 38.3 ± 27.3 | 28.3 ± 25.0 | 24.1 ± 17.7 | 21.8 ± 27.4 | −7.6 ± 19.1 |
| [18] | [6] | [6] | [6] | [6] | |
| Mentha arvensis | 36.3 ± 24.9 | 8.7 ± 14.1 | 0.3 ± 9.6 | 1.7 ± 13.1 | 2.3 ± 8.5 |
| [18] | [8] | [6] | [6] | [6] | |
| (−)-terpinen-4-ol | 47.4 ± 37.7 | 18.0 ± 19.3 | 28.4 ± 34.2 | 7.0 ± 13.2 | 8.5 ± 38.0 |
| [20] | [6] | [8] | [6] | [6] | |
| isopulegol | 49.9 ± 28.9 | 10.0 ± 36.1 | 15.5 ± 9.3 | 25.2 ± 14.4 | −5.3 ± 48.2 |
| [18] | [6] | [6] | [6] | [6] | |
| Mentha spicata | 20.0 ± 34.1 | 15.7 ± 17.8 | 16.4 ± 21.9 |
| [6] | Mentha arvensis | 8.2 ± 34.0 | 36.4 ± 23.4 |
| [6] | [6] | (−)-terpinen-4-ol | −2.3 ± 7.0 |
| [6] | [6] | [6] | isopulegol |
3.4. Bioassay Assessment
| Group | Runs (N) | Mean ± SD | Median | Min–Max | |||
|---|---|---|---|---|---|---|---|
| TTWW a | MNW b | TTWW a | MNW b | TTWW a | MNW b | ||
| 01 | 56 | 52.1 ± 56.0 | 4.2 ± 1.8 | 30 | 4 | 0–210 | 1–9 |
| 02 | 54 | 65.4 ± 70.6 | 4.5 ± 2.4 | 45 | 4 | 0–250 | 2–12 |
| 03 | 32 | 21.3 ± 38.8 | 6.0 ± 2.4 | 0 | 6 | 0–130 | 2–12 |
| 04 | 50 | 23.6 ± 36.4 | 5.5 ± 1.9 | 10 | 5 | 0–200 | 2–9 |
| 05 | 16 | 45.6 ± 45.2 | 3.6 ± 1.3 | 30 | 4 | 10–190 | 2–7 |
| 06 | 20 | 37.0 ± 49.2 | 5.1 ± 2.3 | 20 | 4 | 0–170 | 1–9 |
| 07 | 46 | 38.7 ± 43.0 | 4.4 ± 1.7 | 20 | 4 | 0–160 | 2–9 |
| 08 | 56 | 44.1 ± 70.0 | 4.8 ± 1.9 | 15 | 5 | 0–300 | 0–10 |
| 09 | 22 | 59.1 ± 58.1 | 3.4 ± 1.3 | 45 | 3 | 0–220 | 1–6 |
| 10 | 64 | 15.2 ± 17.8 | 6.4 ± 1.8 | 10 | 6 | 0–70 | 3–10 |
| 11 | 76 | 17.0 ± 22.2 | 6.3 ± 2.0 | 10 | 6 | 0–110 | 3–13 |
| 12 | 66 | 25.0 ± 34.4 | 6.1 ± 2.0 | 10 | 6 | 0–150 | 2–12 |
| Wasp species | Runs (N) | Mean ± SD | Median | Min–Max | |||
|---|---|---|---|---|---|---|---|
| TTWW a | MNW b | TTWW a | MNW b | TTWW a | MNW b | ||
| Vespula vulgaris | 400 | 28.5 ± 42.8 | 5.5 ± 2.1 | 10 | 5 | 0–300 | 0–13 |
| Vespula germanica | 142 | 50.2 ± 60.9 | 4.7 ± 2.3 | 30 | 4 | 0–250 | 1–12 |
| Polistes dominula | 16 | 45.6 ± 45.2 | 3.6 ± 1.3 | 30 | 3.5 | 10–190 | 2–7 |
4. Discussion
5. Conclusions
Acknowledgments
Author contributions
Conflicts of Interest
References
- Akre, R.D.; Davis, H.G. Biology and pest status of venomous wasps. Annu. Rev. Entomol. 1978, 23, 215–238. [Google Scholar] [CrossRef]
- Schmidt, J.O. Hymenopteran venoms: Striving toward the ultimate defense against vertebrates. In Insect Defenses: Adaptive Mechanisms and Strategies of Prey and Predators; State University of New York Press: Albany, NY, USA, 1990; pp. 387–419. [Google Scholar]
- Pasquet, A.; Cardot, J.; Leborgne, R. Wasp attacks and spider defence in the orb weaving species Zygiella x-notata. J. Insect Behav. 2007, 20, 553–564. [Google Scholar] [CrossRef]
- De Haro, L.; Labadie, M.; Chanseau, P.; Cabot, C.; Blanc-Brisset, I.; Penouil, F. Medical consequences of the Asian black hornet (Vespa velutina) invasion in Southwestern France. Toxicon 2010, 55, 650–652. [Google Scholar]
- Ono, M.; Terabe, H.; Hori, H.; Sasaki, M. Components of giant hornet alarm pheromone. Nature 2003, 424, 637–638. [Google Scholar] [CrossRef]
- Krishna, M.T.; Ewan, P.W.; Diwakar, L.; Durham, S.R.; Frew, A.J.; Leech, S.C.; Nasser, S.M. Diagnosis and management of hymenoptera venom allergy: British Society for Allergy and Clinical Immunology (BSACI) guidelines. Clin. Exp. Allergy 2011, 41, 1201–1220. [Google Scholar] [CrossRef]
- Spurr, E.B.; Drew, K.W.; Read, P.E.C.; Elliott, G. The effectiveness of a sulfluramid concentrate mixed with canned sardine cat-food for control of wasps. In Proceedings of the 49th New Zealand Plant Protection Conference, Nelson, New Zealand, 13–15 August 1996; pp. 132–136.
- Spurr, E.B. Freeze-dried bait for wasp control. In Proceedings of the 50th New Zealand Plant Protection Conference, Lincoln, New Zealand, 18–21 August 1997; pp. 401–404.
- Landolt, P.J.; Reed, H.C.; Aldrich, J.R.; Antonelli, A.L.; Dickey, C. Social wasps (Hymenoptera: Vespidae) trapped with acetic acid and isobutanol. Fla. Entomol. 1999, 82, 609–614. [Google Scholar] [CrossRef]
- Landolt, P.J.; Smithhisler, C.S.; Reed, H.C.; McDonough, L.M. Trapping social wasps (Hymenoptera: Vespidae) with acetic acid and saturated short chain alcohols. J. Econ. Entomol. 2000, 93, 1613–1618. [Google Scholar] [CrossRef]
- Landolt, P.J.; Tóth, M.; Jósvai, J. First European report of social wasps trapped in response to acetic acid, isobutanol, 2-methyl-2-propanol and heptyl butyrate in tests conducted in Hungary. Bull. Insectol. 2007, 60, 7–11. [Google Scholar]
- Makino, S.; Sayama, K. Species compositions of vespine wasps collected with bait traps in recreation forests in northern and central Japan (Insecta, Hymenoptera, Vespidae). Bull. FFPRI 2005, 4, 283–289. [Google Scholar]
- Bacandritsos, N.; Papanastasiou, I.; Saitanis, C.; Roinioti, E. Three non-toxic insect traps useful in trapping wasps enemies of honey bees. Bull. Insectol. 2006, 59, 135–145. [Google Scholar]
- Reierson, D.A.; Rust, M.K.; Vetter, R.S. Traps and protein bait to suppress populations of yellowjackets (Hymenoptera: Vespidae). In Proceedings of the Sixth International Conference on Urban Pests, Veszprém, Hungary, 13–16 July 2008; Robinson, W.H., Bajomi, D., Eds.; pp. 267–274.
- Rust, M.K.; Reierson, D.A.; Vetter, R. Developing Baits for the Control of Yellowjackets in California; Department of Entomology, University of California: Riverside, CA, USA, 2010; pp. 1–33. [Google Scholar]
- Merino, L.; France, A.; Gerding, M. Selection of native fungi strains pathogenic to Vespula germanica (Hymenoptera: Vespidae). Agric. Técnica 2007, 67, 335–342. [Google Scholar]
- Toft, R.J.; Harris, R.J. Can trapping control Asian paper wasp (Polistes chinensis antennalis) populations? N. Z. J. Ecol. 2004, 28, 279–282. [Google Scholar]
- Spurr, E.B. Carbohydrate bait preferences of wasps (Vespula vulgaris and V. germanica) (Hymenoptera: Vespidae) in New Zealand. N. Z. J. Zool. 1996, 23, 315–324. [Google Scholar] [CrossRef]
- Crowder, D.W.; Snyder, W.E. Eating their way to the top? Mechanisms underlying the success of invasive insect generalist predators. Biol. Invasions 2010, 12, 2857–2876. [Google Scholar] [CrossRef]
- Harris, R.J.; Oliver, E.H. Prey diets and polulation densities of the wasps Vespula vulgaris and V. germanica in scrubland-pasture. N. Z. J. Ecol. 1993, 17, 5–12. [Google Scholar]
- Barr, K.; Moller, H.; Christmas, E.; Lyver, P.; Beggs, J. Impacts of introduced common wasps (Vespula vulgaris) on experimentally placed mealworms in a New Zealand beech forest. Oecologia 1996, 105, 266–270. [Google Scholar]
- Beggs, J.R.; Rees, J.S. Restructuring of Lepidoptera communities by introduced Vespula wasps in a New Zealand beech forest. Oecologia 1999, 119, 565–571. [Google Scholar] [CrossRef]
- Richter, M.R. Social wasp (Hymenoptera: Vespidae) foraging behavior. Annu. Rev. Entomol. 2000, 45, 121–150. [Google Scholar] [CrossRef]
- Wilson, E.E.; Mullen, L.M.; Holway, D.A. Life history plasticity magnifies the ecological effects of a social wasp invasion. Proc. Natl. Acad. Sci. USA 2009, 106, 12809–12813. [Google Scholar] [CrossRef]
- Markwell, T.J.; Kelly, D.; Duncan, K.W. Competition netween honey bees (Apis mellifera) and wasps (Vespula spp.) in honeydew beech (Nothofagus solandri var. solandri) forest. N. Z. J. Ecol. 1993, 17, 85–93. [Google Scholar]
- Beggs, J.R.; Wardle, D.A. Keystone species: Competition for honeydew among exotic and indigenous species. In Ecological Studies, Vol. 186; Allen, R.B., Lee, W.G., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 2006; Volume 186, pp. 281–294. [Google Scholar]
- Kenis, M.; Auger-Rozenberg, M.-A.; Roques, A.; Timms, L.; Péré, C.; Cock, M.J.W.; Settele, J.; Augustin, S.; Lopez-Vaamonde, C. Ecological effects of invasive alien insects. Biol. Invasions 2009, 11, 21–45. [Google Scholar] [CrossRef]
- Wardle, D.A.; Karl, B.J.; Beggs, J.R.; Yeates, G.W.; Williamson, W.M.; Bonner, K.I. Determining the impact of scale insect honeydew, and invasive wasps and rodents, on the decomposer subsystem in a New Zealand beech forest. Biol. Invasions 2009, 12, 2619–2638. [Google Scholar]
- Dvořák, L.; Roberts, S.P.M. Key to the paper and social wasps of Central Europe (Hymenoptera: Vespidae). Acta Entomol. Musei Natl. Pragae 2006, 46, 221–244. [Google Scholar]
- Rome, Q.; Perrard, A.; Muller, F.; Villemant, C. Monitoring and control modalities of a honeybee predator, the yellow-legged hornet Vespa velutina nigrithorax (Hymenoptera: Vespidae). Aliens Invasive Species Bull. 2011, 31, 7–15. [Google Scholar]
- Zhang, Q.-H.; Schneidmiller, R.G.; Hoover, D.R. Essential oils and their compositions as spatial repellents for pestiferous social wasps. Pest Manag. Sci. 2012, 69, 542–552. [Google Scholar]
- Veith, H.J.; Koeniger, N.; Maschwitz, U. 2-Methyl-3-butene-2-ol, a major component of the alarm pheromone of the hornet Vespa crabro. Naturwissenschaften 1984, 71, 328–329. [Google Scholar] [CrossRef]
- Heath, R.R.; Landolt, P.J. The isolation , identification and synthesis of the alarm pheromone of Vespula squamosa (Drury) (Hymenoptera: Vespidae) and associated behavior. Experientia 1988, 44, 82–83. [Google Scholar] [CrossRef]
- Fortunato, A.; Dani, F.R.; Sledge, M.F.; Fondelli, L.; Turillazzi, S. Alarm communication in Ropalidia social wasps. Insectes Soc. 2004, 51, 299–305. [Google Scholar]
- Bruschini, C.; Dani, F.R.; Pieraccini, G.; Guarna, F.; Turillazzi, S. Volatiles from the venom of five species of paper wasps (Polistes dominulus, P. gallicus, P. nimphus, P. sulcifer and P. olivaceus). Toxicon 2006, 47, 812–825. [Google Scholar] [CrossRef]
- Ono, M.; Igarashi, T.; Ohno, E.; Sasaki, M. Unusual thermal defence by a honeybee against mass attack by hornets. Nature 1995, 377, 334–336. [Google Scholar] [CrossRef]
- Ikan, R.; Gottlieb, R.; Bergmann, E.D.; Ishay, J. The pheromone of the queen of the oriental hornet, Vespa orientalis. J. Insect Physiol. 1969, 15, 1709–1712. [Google Scholar] [CrossRef]
- Ruther, J.; Sieben, S.; Schricker, B. Role of cuticular lipids in nestmate recognition of the European hornet Vespa crabro L. (Hymenoptera: Vespidae). Insectes Soc. 1998, 45, 169–179. [Google Scholar] [CrossRef]
- Ruther, J.; Sieben, S.; Schricker, B. Nestmate recognition in social wasps: Manipulation of hydrocarbon profiles induces aggression in the European hornet. Naturwissenschaften 2002, 89, 111–114. [Google Scholar] [CrossRef]
- Steinmetz, I.; Sieben, S.; Schmolz, E. Chemical trails used for orientation in nest cavities by two vespine wasps, Vespa crabro and Vespula vulgaris. Insectes Soc. 2002, 49, 354–356. [Google Scholar]
- Steinmetz, I.; Schmolz, E.; Ruther, J. Cuticular lipids as trail pheromone in a social wasp. Proc. R. Soc. Biol. Sci. 2003, 270, 385–391. [Google Scholar] [CrossRef]
- Ono, M.; Sasaki, M. Sex pheromones and their cross-activities in six Japanese sympatric species of the genus Vespa. Insectes Soc. 1987, 34, 252–260. [Google Scholar] [CrossRef]
- Spiewok, S.; Schmolz, E.; Ruther, J. Mating system of the European hornet Vespa crabro: Male seeking strategies and evidence for the involvement of a sex pheromone. J. Chem. Ecol. 2006, 32, 2777–2788. [Google Scholar] [CrossRef]
- Taylor, B.J.; Schalk, D.R.; Jeanne, R.L. Yellowjackets use nest-based cues to differentially exploit higher-quality resources. Naturwissenschaften 2010, 97, 1041–1046. [Google Scholar] [CrossRef]
- MacKenzie, J.K.; Landolt, P.J.; Zack, R.S. Attraction to ornamental peony (Paeonia lactiflora, Paeoniaceae) by Polistes dominulus Christ (Hymenoptera: Vespidae) demonstrated using olfactometers. J. Kansas Entomol. Soc. 2006, 79, 231–238. [Google Scholar] [CrossRef]
- Brodmann, J.; Twele, R.; Francke, W.; Hölzler, G.; Zhang, Q.-H.; Ayasse, M. Orchids mimic green-leaf volatiles to attract prey-hunting wasps for pollination. Curr. Biol. 2008, 18, 740–744. [Google Scholar] [CrossRef]
- Jander, R. Olfactory learning of fruit odors in the Eastern yellow jacket, Vespula maculifrons (Hymenoptera: Vespidae). J. Insect Behav. 1998, 11, 879–888. [Google Scholar] [CrossRef]
- Aldrich, J.R.; Kochansky, J.P.; Sexton, J.D. Chemical attraction of the eastern yellowjacket, Vespula maculifrons (Hymenoptera: Vespidae). Experientia 1985, 41, 420–422. [Google Scholar] [CrossRef]
- Aldrich, J.R.; Lusby, W.R.; Kochansky, J.P. Identification of a new predaceous stink bug pheromone and its attractiveness to the eastern yellowjacket. Experientia 1986, 42, 583–585. [Google Scholar] [CrossRef]
- Landolt, P.J.; Heath, R.R.; Reed, H.C.; Manning, K. Pheromonal mediation of alarm in the Eastern yellowjacket (Hymenoptera: Vespidae). Fla. Entomol. 1995, 78, 101–108. [Google Scholar]
- Weston, R.J.; Woolhouse, A.D.; Spurr, E.B.; Harris, R.J.; Suckling, D.M. Spiroacetals and other venom constituents as potential wasp attractants. J. Chem. Ecol. 1997, 23, 553–568. [Google Scholar] [CrossRef]
- Siegel, S. Nonparametric Statistics for the Behavioral Sciences; McGraw-Hill: New York, NY, USA, 1956; pp. 1–312. [Google Scholar]
- Baser, K.H.C.; Buchbauer, G. Handbook of Essential Oils: Science, Technology, and Applications; CRC Press/Taylor and Francis: Boca Raton, FL, USA, 2010; p. 975. [Google Scholar]
- Cook, S.M.; Khan, Z.R.; Picket, J.A. The use of push-pull strategies in Integrated Pest Management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Boevé, J.-L.; Honraet, K.; Rossel, B. Screening of Repellents against Vespid Wasps. Insects 2014, 5, 272-286. https://doi.org/10.3390/insects5010272
Boevé J-L, Honraet K, Rossel B. Screening of Repellents against Vespid Wasps. Insects. 2014; 5(1):272-286. https://doi.org/10.3390/insects5010272
Chicago/Turabian StyleBoevé, Jean-Luc, Kris Honraet, and Bart Rossel. 2014. "Screening of Repellents against Vespid Wasps" Insects 5, no. 1: 272-286. https://doi.org/10.3390/insects5010272
APA StyleBoevé, J.-L., Honraet, K., & Rossel, B. (2014). Screening of Repellents against Vespid Wasps. Insects, 5(1), 272-286. https://doi.org/10.3390/insects5010272