Horizontal Transmission of Metarhizium anisopliae in Fruit Flies and Effect of Fungal Infection on Egg Laying and Fertility

Abstract
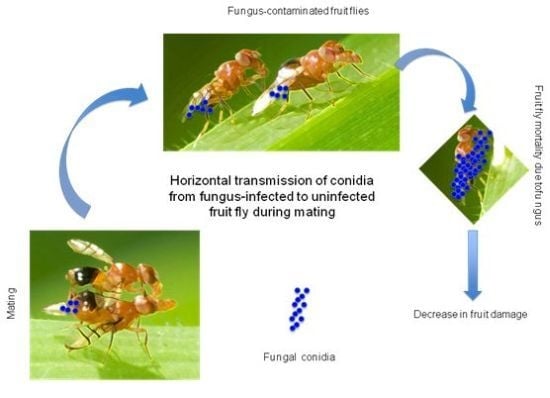
:
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Fungal Isolate
2.3. Quantification of Conidial Uptake by Fruit Flies from Treated Substrate
2.4. Transfer of Inoculum
2.5. Transfer of Inoculum through a Chain of Individuals
2.6. Effect of Fungal Infection by M. anisopliae on Reproduction Potential
2.7. Data Analysis
3. Results and Discussion
3.1. Retention of Conidia

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time after treatment (h) | Mean number of conidia per fly |
|---|---|
| 0 | 1.1 × 106 |
| 2 | 7.2 × 105 |
| 4 | 6.1 × 105 |
| 6 | 4.3 × 105 |
| 8 | 4.1 × 105 |
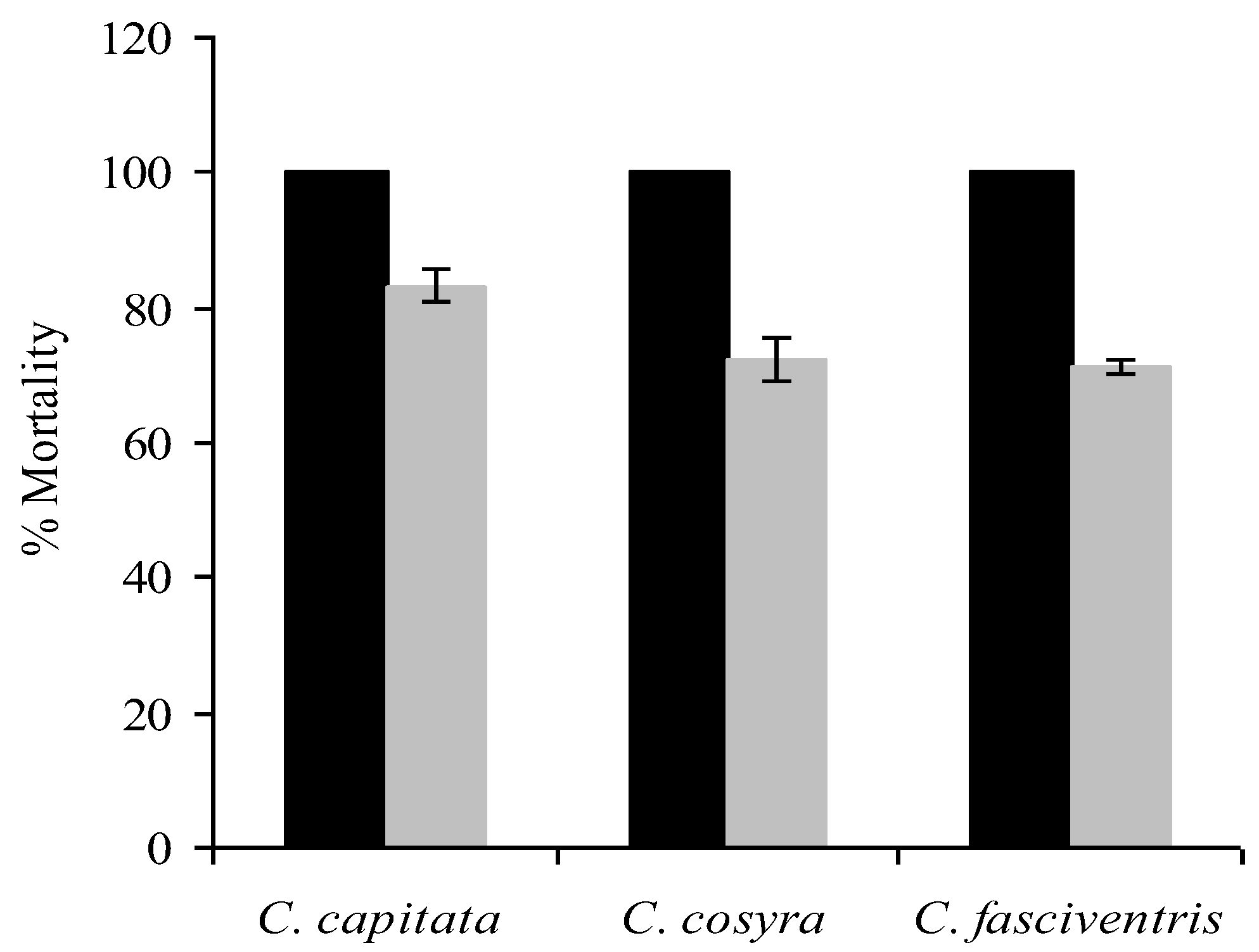
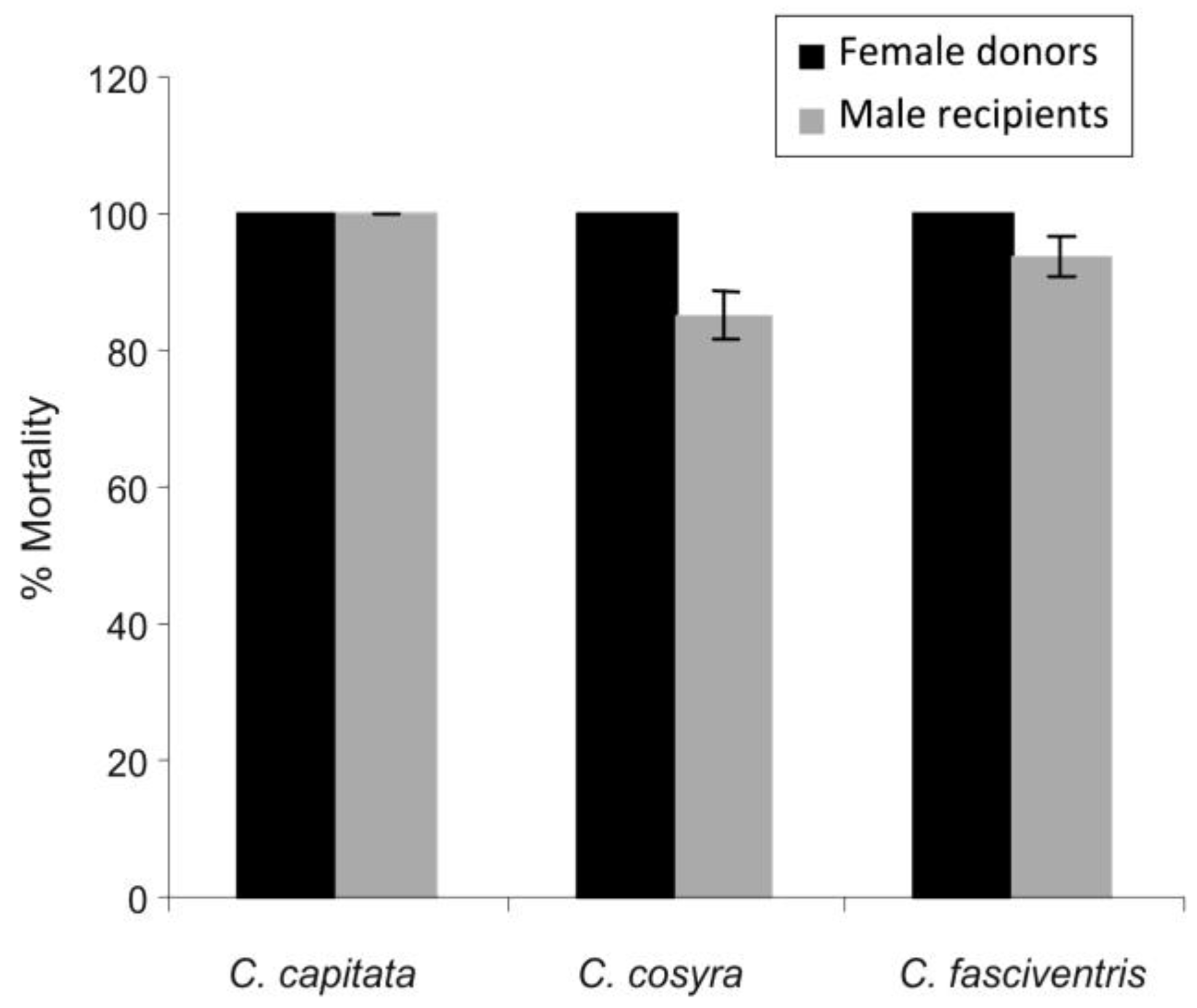
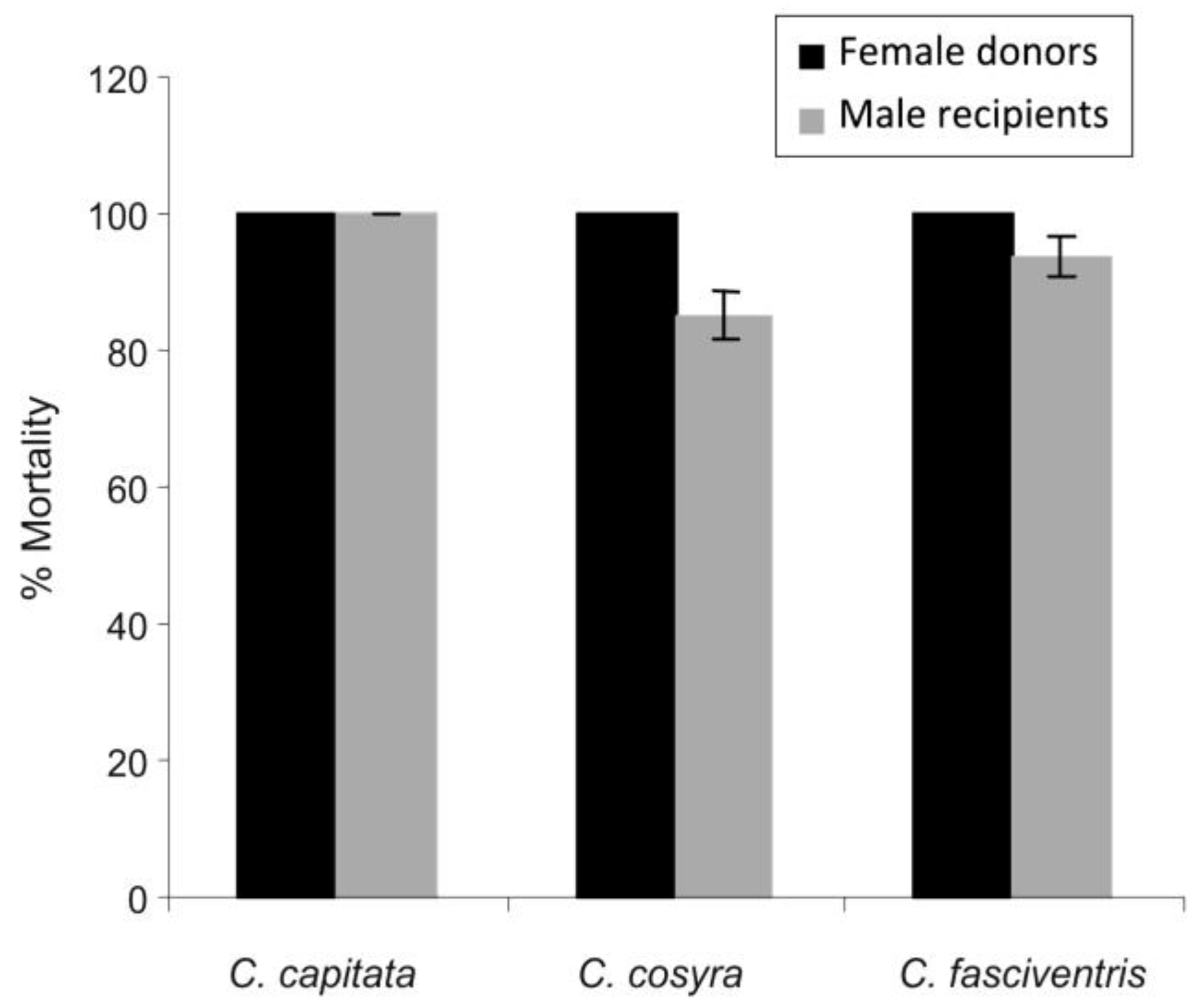
3.2. Transfer of Inoculum

| LT50 day (X ± SE) | |||
|---|---|---|---|
| C. capitata | C. cosyra | C. fasciventris | |
| Male “donors” | 2.6 ± 0.1 | 2.9 ± 0.2 | 2.9 ± 0.1 |
| Female “donor” | 2.8 ± 0.1 | 3.4 ± 0.3 | 3.1 ± 0.1 |
| Female “recipient” | 8.1 ± 0.4 | 10.4 ± 0.4 | 10.5 ± 0.4 |
| Male “recipient” | 5.6 ± 0.5 | 7.9 ± 0.3 | 7.7 ± 0.2 |
3.3. Transfer of Inoculum through a Chain of Individuals
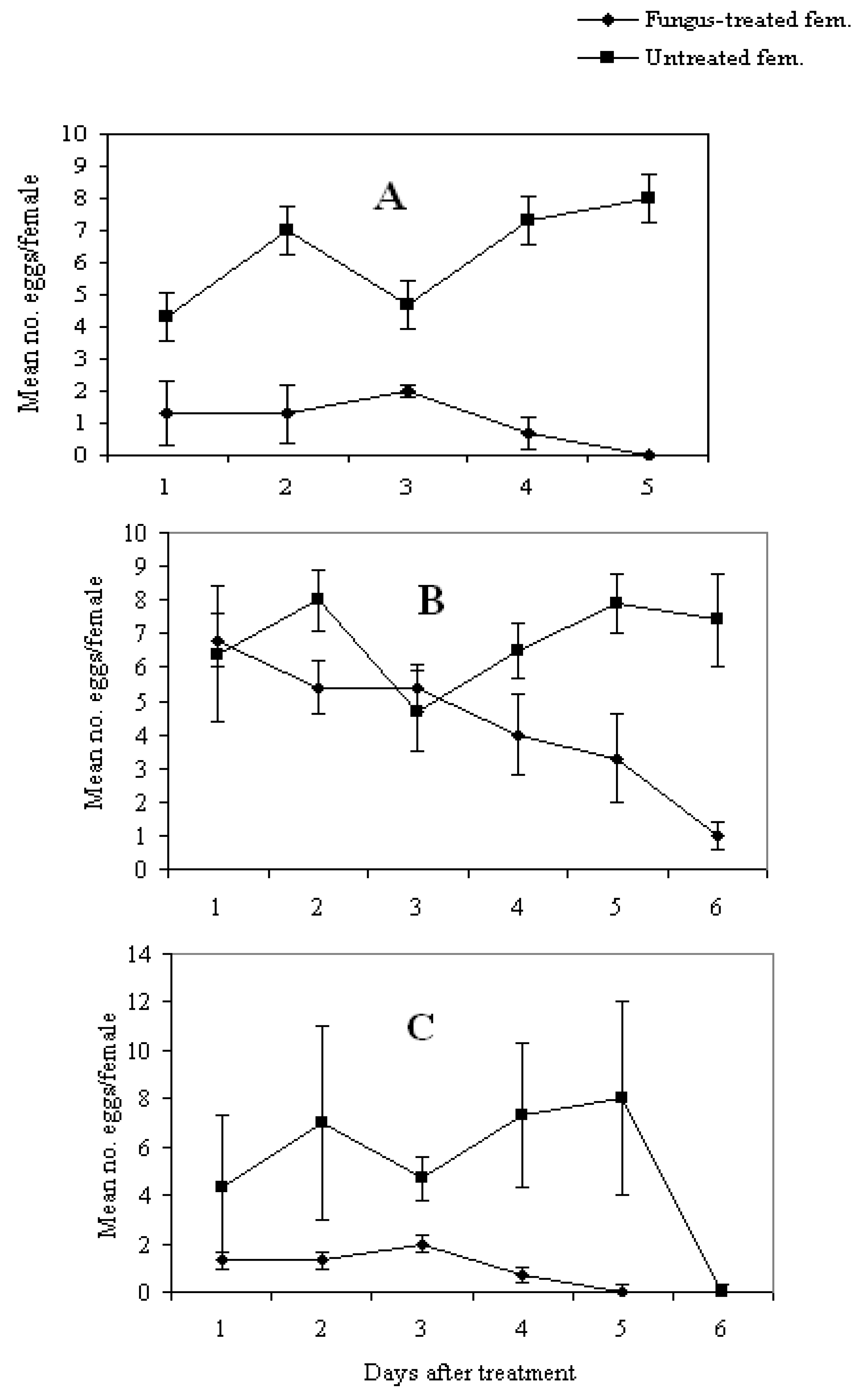
3.4. Effect of Fungal Infection on Reproduction Potential
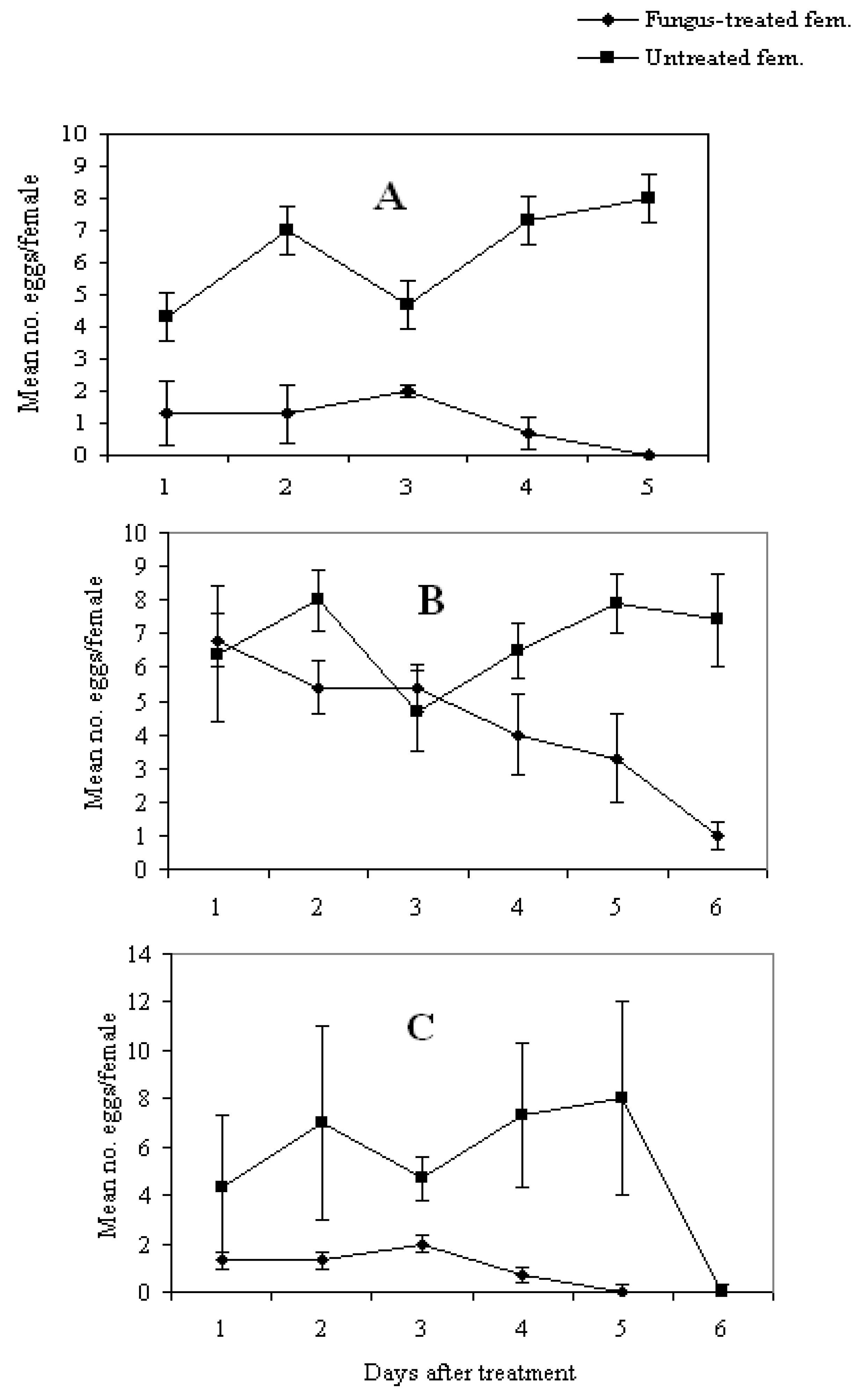
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Lux, S.A.; Ekesi, S.; Dimbi, S.; Mohamed, S.; Billah, M. Mango infesting fruit flies in Africa—Perspectives and limitations of biological approaches to their management. In Biological Control in Integrated Pest Management Systems in Africa; Neuenschwander, P., Borgemeister, C., Langewald, J., Eds.; CABI: Wallingford, CT, USA, 2003; pp. 277–293. [Google Scholar]
- Ekesi, S.; Nderitu, P.W.; Rwomushana, I. Field infestation and demographic parameters of the fruit fly Bactrocera invadens (Diptera: Tephritidae) in Africa. Bull. Entomol. Res. 2006, 96, 379–386. [Google Scholar]
- Ekesi, S.; Maniania, N.K.; Lux, S.A. Mortality in three African tephritid fruit fly puparia and adults caused by the entomopathogenic fungi, Metarhizium anisopliae and Beauveria bassiana. Biocontrol Sci. Technol. 2002, 12, 7–17. [Google Scholar] [CrossRef]
- Saul, S.H.; Tsuda, D.; Wong, T.T.Y. Laboratory and field trials of soil applications of methoprene and other insecticides for control of the Mediterranean fruit fly (Diptera: Tephritidae). J. Econ. Entomol. 1983, 76, 174–177. [Google Scholar]
- Roessler, Y. Insecticidal bait and cover sprays. In Fruit flies: Their Biology, Natural Enemies and Control. World Crop Pests; Robinson, A.S., Hooper, G., Eds.; Elsevier: Amsterdam, the Netherlands, 1989; pp. 329–336. [Google Scholar]
- Ekesi, S.; Dimbi, S.; Maniania, N.K. The role of entomopathogenic fungi in the integrated management of fruit flies (Diptera: Tephritidae) with emphasis on species occurring in Africa. In Use of Entomopathogenic Fungi in Biological Pest Management; Ekesi, S., Maniania, N.K., Eds.; Research SignPost: Kerala, India, 2007; pp. 239–274. [Google Scholar]
- Ekesi, S.; Maniania, N.K.; Mohamed, S.A. Efficacy of soil application of Metarhizium anisopliae and the use of GF-120 spinosad bait spray for suppression of Bactrocera invadens (Diptera: Tephritidae) in mango orchards. Biocontrol Sci. Technol. 2011, 21, 299–316. [Google Scholar] [CrossRef]
- Dimbi, S.; Maniania, N.K.; Lux, S.A.; Ekesi, S.; Mueke, J.K. Pathogenicity of Metarhizium anisopliae (Metsch.) Sorokin and Beauveria bassiana (Balsamo) Vuillemin, to three adult fruit fly species: Ceratitis capitata (Weidemann), C. rosa var. fasciventris Karsch and C. cosyra (Walker) (Diptera: Tephritidae). Mycopathologia 2003, 156, 375–382. [Google Scholar] [CrossRef]
- Ekesi, S.; Maniania, N.K.; Mohamed, S.A.; Lux, S.A. Effect of soil application of different formulations of Metarhizium anisopliae on African tephritid fruit flies and their associated endoparasitoids. Biol. Control. 2005, 35, 83–91. [Google Scholar] [CrossRef]
- Toledo, J.; Liedo, P.; Flores, S.; Campos, S.E.; Villasenor, A.; Montoya, P. Use of Beauveria bassiana and Metarhizium anisopliae for fruit fly control: A novel approach. In Proceedsing of 7th International Symposium on Fruit Flies of Economic Importance, Salvador, Brazil, 10–15 September 2006; pp. 127–132.
- Kaaya, G.P.; Okech, M.A. Horizontal transmission of mycotic infection in adult tsetse, Glossina morsitans morsitnans. Entomophaga. 1990, 35, 589–600. [Google Scholar] [CrossRef]
- Maniania, N.K. A laboratory technique for infecting tsetse flies with a fungal pathogen. Insect Sci. Appl. 1994, 15, 421–426. [Google Scholar]
- Meadow, R.; Vandenberg, J.D.; Shelton, M. Exchange of inoculum of Beauveria bassiana (Bals.) Vuill. (Hyphomycetes) between adult flies of the cabbage maggot Delia radicum L. (Diptera: Anthomyiidae). Biocontrol Sci. Technol. 2000, 10, 479–485. [Google Scholar] [CrossRef]
- Scholte, E.J.; Knols, B.G.J.; Samson, R.A.; Takken, W. Entomopathogenic fungi for mosquito control: A review. J. Insect Sci. 2004, 4, 24. [Google Scholar]
- Toledo, J.; Campos, S.E.; Flores, S.; Liedo, P.F.; Barrera, J.; Villaseñor, A.; Montoya, P. Horizontal transmission of Beauveria bassiana in Anastrepha ludens (Diptera: Tephritidae) under laboratory and field cage conditions. J. Econ. Entomol. 2007, 100, 291–297. [Google Scholar]
- Quesada-Moraga, E.; Martin-Carballo, I.; Garrido-Jurado, I.; Santiago-Álvarez, C. Horizontal transmission of Metarhizium anisopliae among laboratory populations of Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). Biol. Control. 2008, 47, 115–124. [Google Scholar] [CrossRef]
- Maniania, N.K.; Okech, M.A.; Adino, J.O.; Opere, J.O.; Ekesi, S. Transfer of inoculum of Metarhizium anisopliae between adult Glossina morsitans morsitans and effects of fungal infection on blood feeding and mating behaviors. J. Pest Sci. 2012. [Google Scholar] [CrossRef]
- Eilenberg, J. Abnormal egg laying behavior of female carrot flies (Psila roase) induced by fungus Entomophthora muscae. Entomol. Exp. Appl. 1987, 43, 61–65. [Google Scholar]
- Watson, D.W.; Petersen, J.J. Sexual activity of male Musca domestica (Diptera: Muscidae) infected with Entomophthora muscae (Entomophthoraceae: Entomophthorales). Biol. Control. 1993, 3, 22–26. [Google Scholar] [CrossRef]
- Castillo, M.A.; Moya, P.; Primo-Yứfera, E. Susceptibility of Ceratitis capitata Wiedemann (Diptera: Tephritidae) to entomopathogenic fungi and their extracts. Biol. Control. 2000, 19, 274–282. [Google Scholar] [CrossRef]
- Mouatcho, J.C.; Koekemoer, L.L.; Coetzee, M.; Brooke, B.D. The effect of entomopathogenic fungus infection on female fecundity of the major malaria vector, Anopheles funestus. Afr. Entomol. 2011, 19, 725–729. [Google Scholar] [CrossRef]
- Falcon, L.A. Development and use of microbial insecticides. In Biological Control in Agricultural IPM Systems; Hoy, M.A., Herzog, D.C., Eds.; Academic Press: Orlando, FL, USA, 1985; pp. 229–242. [Google Scholar]
- Hooper, G.H.S. Application of quality control procedures for large scale rearing of the Mediterranean fruit fly. Entomol. Exp. Appl. 1987, 44, 161–167. [Google Scholar] [CrossRef]
- Goettel, M.S.; Inglis, D.G. Fungi Hyphomycetes. In Manual of Techniques in Insect Pathology; Lacey, L.A., Ed.; Academics Press: New York, NY, USA, 1997; pp. 213–249. [Google Scholar]
- Abbott, W.S. A method for computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar]
- SAS Institute Inc. SAS/STAT User’s Guide: Version 6, 4th ed.; SAS Institute Inc.: Cary, NC, USA, 1990; Volume 2. [Google Scholar]
- SAS, SAS/STAT. User’s Guide Release 8.E; SAS Institute Inc.: Cary, NC, USA, 1999. [Google Scholar]
- Prokopy, R.J.; Hendrichs, J. Mating behavior of Ceratitis capitata on a field-caged host tree. Ann. Entomol. Soc. Am. 1979, 72, 642–648. [Google Scholar]
- Hedström, I.; Monge-Nájera, J. Is sexually transmitted fungal infection evidence for size-related mating success in Neotropical guava fruit flies? J. Trop. Biol. 1998, 46, 1129–1132. [Google Scholar]
- Shelly, T.E.; Villalobos, E.M. Cue lure and the mating behavior of male melonies (Diptera: Tephritidae). Fla. Entomol. 1995, 78, 473–482. [Google Scholar] [CrossRef]
- Lance, D.R.; Mcinnis, D.O.; Rendon, P.; Jackson, C.G. Courtship among sterile and wild Ceratitis capitata (Diptera: Tephritidae) in field cages in Hawaii and Guatemala. Ann. Entomol. Soc. Am. 2000, 93, 1179–1185. [Google Scholar] [CrossRef]
- Scholte, E.-J.; Knols, B.G.J.; Takken, W. Infection of Anopheles gambiae with the entomopathogenic fungus Metarhizium anisopliae reduces blood feeding and fecundity. J. Invertebr. Pathol. 2006, 91, 43–49. [Google Scholar] [CrossRef]
- Blanford, S.; Shi, W.; Riann, C.; Marden, J.H.; Koekemoer, L.L.; Brooke, B.D.; Coetzee, M.; Read, A.F.; Thomas, M.B. Lethal and Pre-lethal effects of a fungal biopesticide contribute to substantial and rapid vector control. PLoS One 2011, 6, e23591. [Google Scholar]
- Maniania, N.K.; Ekesi, S.; Odulaja, A.; Okech, M.A.; Nadel, D.J. Prospects of a fungus-contamination device for the control of tsetse fly Glossina fuscipes fuscipes. Biocontrol Sci. Technol. 2006, 16, 129–139. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Dimbi, S.; Maniania, N.K.; Ekesi, S. Horizontal Transmission of Metarhizium anisopliae in Fruit Flies and Effect of Fungal Infection on Egg Laying and Fertility. Insects 2013, 4, 206-216. https://doi.org/10.3390/insects4020206
Dimbi S, Maniania NK, Ekesi S. Horizontal Transmission of Metarhizium anisopliae in Fruit Flies and Effect of Fungal Infection on Egg Laying and Fertility. Insects. 2013; 4(2):206-216. https://doi.org/10.3390/insects4020206
Chicago/Turabian StyleDimbi, Susan, Nguya K. Maniania, and Sunday Ekesi. 2013. "Horizontal Transmission of Metarhizium anisopliae in Fruit Flies and Effect of Fungal Infection on Egg Laying and Fertility" Insects 4, no. 2: 206-216. https://doi.org/10.3390/insects4020206