Neotenic Phenotype and Sex Ratios Provide Insight into Developmental Pathways in Reticulitermes flavipes (Isoptera: Rhinotermitidae)

Abstract
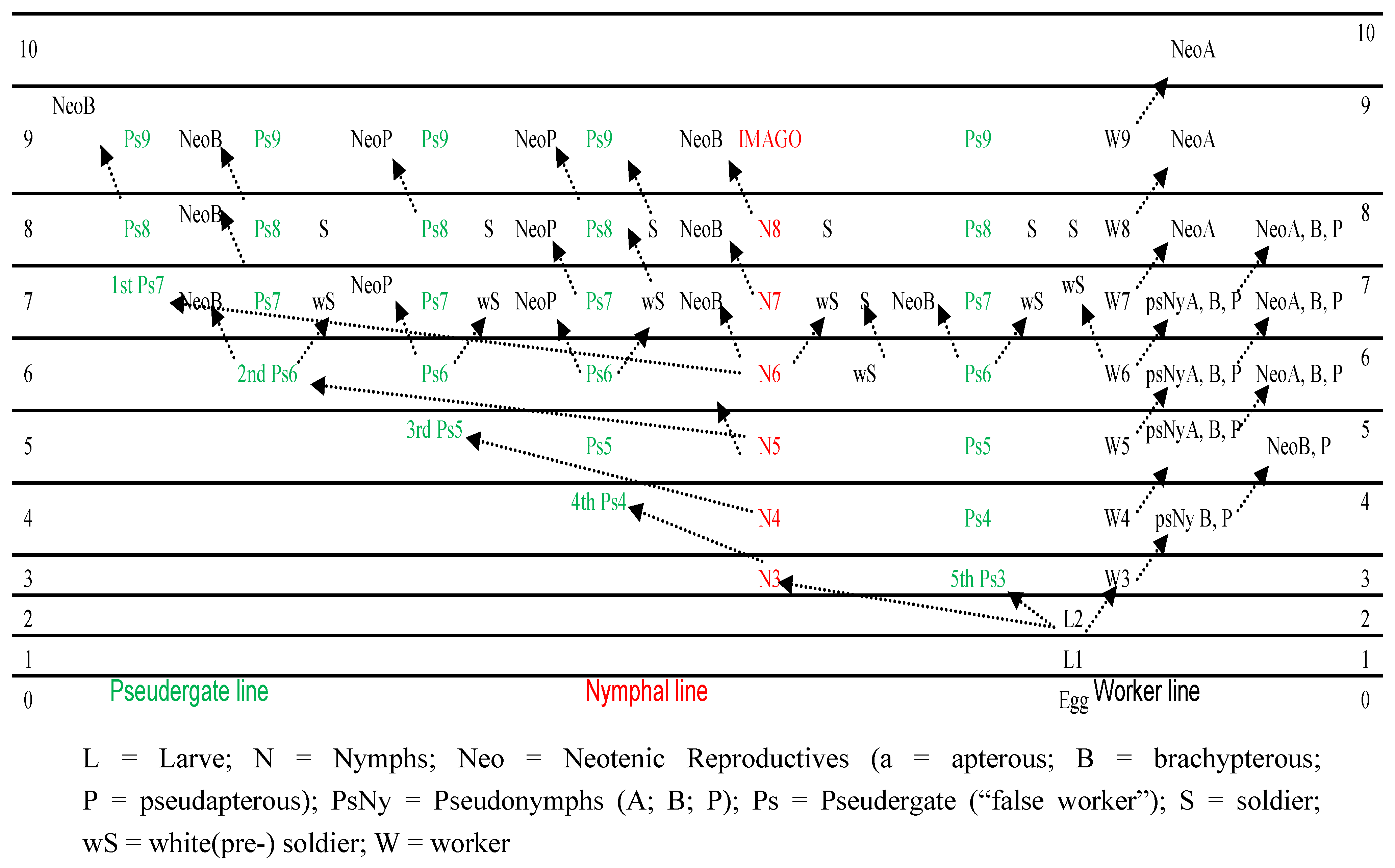
:1. Introduction
2. Material and Methods
2.1. Specimen Collection

2.2. Sex determination

{kind=link}
{kind=link}
| Collection | Caste | ♂ (male) | ♀ (female) | Sex ratio |
|---|---|---|---|---|
| Log A | Regular neotenic | 306 | 335 | 0.91 |
| Black-headed neotenic | 373 | 5 | 73.50 | |
| Black neotenic | 51 | 0 | exclusively male | |
| Soldier | 157 | 309 | 0.51 | |
| Worker | 226 | 174 | 1.30 | |
| Nymph | 155 | 200 | 0.78 | |
| Alates | 351 | 271 | 1.30 | |
| Neotenic +Soldier | 887 | 649 | 1.37 | |
| Log B | Regular neotenic | 243 | 774 | 0.31 |
| Black-headed neotenic | 94 | 5 | 15.40 | |
| Soldier | 133 | 125 | 1.06 | |
| Worker | 34 | 20 | 1.70 | |
| Neotenic +Soldier | 470 | 904 | 0.52 | |
| Log C | Regular neotenic | 4 | 14 | 0.29 |
| Black-headed neotenic | 1 | 3 | 0.33 | |
| Black neotenic | 5 | 0 | exclusively male | |
| Soldier | 11 | 21 | 0.52 | |
| Worker | 137 | 68 | 2.01 | |
| Nymph | 5 | 13 | 0.38 | |
| Neotenic +Soldier | 21 | 38 | 0.55 |
2.3. Results
2.4. Discussion
| Collection Paired date | Caste | Date of census | ♂ (male) | ♀ (female) | Sex ratio |
|---|---|---|---|---|---|
| Colony I 2/3/2003 | Regular neotenic | 10/10/2008 | 9 | 41 | 0.22 |
| Black-headed neotenic | 6 | 5 | 1.20 | ||
| Nymph | 8 | 9 | 0.89 | ||
| Soldier | 11 | 6 | 1.83 | ||
| Worker | 113 | 65 | 1.74 | ||
| Colony II 3/11/2003 | Regular neotenic | 10/10/2008 | 0 | 2 | exclusively female |
| Nymph | 81 | 132 | 0.61 | ||
| Soldier | 17 | 12 | 1.42 | ||
| Worker | 155 | 48 | 3.23 | ||
| Colony III 2/21/2000 | Reproductive (K, K, Q) | 12/4/2008 | 2 | 1 | 2.00 |
| Nymph | 61 | 10 | 0.87 | ||
| Soldier | 68 | 43 | 1.58 | ||
| Worker | 97 | 60 | 1.62 | ||
| Colony IV 4/21/2003 | Reproductive (K, Q) | 7/31/2008 | 1 | 1 | 1.00 |
| Nymph | 1 | 1 | 1.00 | ||
| Soldier | 33 | 46 | 0.72 | ||
| Worker | 84 | 43 | 1.95 | ||
| Colony V 4/21/2003 | Reproductive (K, Q) | 10/23/2008 | 1 | 1 | 1.00 |
| Soldier | 32 | 26 | 1.23 | ||
| Worker | 135 | 91 | 1.48 | ||
| Colony VI 4/21/2003 | Reproductive (K, Q) | 12/5/2008 | 1 | 1 | 1.00 |
| Nymph | 1 | 1 | 1.00 | ||
| Soldier | 31 | 7 | 4.43 | ||
| Worker | 82 | 33 | 2.48 | ||
| Colony VII 10/6/2004 | Reproductive (K, Q) | 6/1/2009 | 1 | 1 | 1.00 |
| Nymph | 0 | 2 | exclusively female | ||
| Soldier | 9 | 3 | 3.00 | ||
| Worker | 84 | 35 | 2.40 | ||
| Colony VIII 7/25/1995 | Reproductive (K, Q) | 6/1/2009 | 1 | 1 | 1.00 |
| Nymph | 0 | 4 | exclusively female | ||
| Soldier | 19 | 26 | 0.73 | ||
| Worker | 127 | 47 | 2.70 |
3. Neotenic Differentiation and Development
4. Sex Ratios Provide Hints to Developmental Pathways

5. Sex Ratios and Castes of Laboratory Colonies
6. Summary
Acknowledgments
References
- Roisin, Y. Diversity and Evolution of Caste Patterns. In Termites: Evolution, Sociality, Symbioses, Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 95–120. [Google Scholar]
- Korb, J.; Hartfelder, K. Life history and development—A framework for understanding developmental plasticity in lower termites. Biol. Rev. 2008, 83, 295–313. [Google Scholar]
- Miller, E.M. Caste Differentiation in the Lower Termites. In Biology of Termites; Krishna, K., Weesner, F.M., Eds.; Academic Press: New York, NY, USA, 1969; pp. 283–310. [Google Scholar]
- Hrdy, I.; Krecek, J. Development of superfluous soldiers induced by juvenile-hormone analogs in termite, Reticulitermes Lucifugus Santonensis. Insectes Soc. 1972, 19, 105–109. [Google Scholar] [CrossRef]
- Noirot, C. Sexual Castes and Reproductive Strategies in Termites. In An Evolutionary Approach to Castes and Reproduction; Engels, W., Ed.; Springer-Verlag, Berlin, Germany, 1999; pp. 5–35. [Google Scholar]
- Laine, L.V.; Wright, D.J. The life cycle of Reticulitermes spp. (Isopter: Rhinotermitidae): What do we know? Bull. Entomol. Res. 2003, 93, 267–278. [Google Scholar]
- Mao, L.X.; Henderson, G.; Liu, Y.X.; Laine, R.A. Formosan subterranean termite (Isoptera: Rhinotermitidae) soldiers regulate juvenile hormone levels and caste differentiation in workers. Ann. Entomol. Soc. Am. 2005, 98, 340–345. [Google Scholar] [CrossRef]
- Cornette, R.; Gotoh, H.; Koshikawa, S.; Miura, T. Juvenile hormone titers and caste differentiation in the damp-wood termite Hodotermopsis sjostedri (Isoptera: Termopsidae). J. Insect Physiol. 2008, 54, 922–930. [Google Scholar] [CrossRef]
- Elliott, K.L.; Stay, B. Changes in juvenile hormone synthesis in the termite Reticulitermes flavipes during development of soldiers and neotenic reproductives from groups of isolated workers. J. Insect Physiol. 2008, 54, 492–500. [Google Scholar] [CrossRef]
- Muller, H.; Korb, J. Male or female soldiers? An evaluation of several factors which may influence soldier sex ratio in lower termites. Insectes Soc. 2008, 55, 213–219. [Google Scholar] [CrossRef]
- Raina, A.; Park, Y.; Gelman, D. Molting in workers of the Formosan subterranean termite Coptotermes formosanus. J. Insect Physiol. 2008, 54, 155–161. [Google Scholar] [CrossRef]
- Snyder, T.E. Insects Injurious to Forests and Forest Products. Biology of the Termites of the Eastern United States, with Preventative and Remedial Measures; U.S. Government Printing Office: Washinton, DC, USA, 1915; pp. 13–95. [Google Scholar]
- Hare, L. Caste determination and differentiation with special reference to the genus Reticulitermes (Isoptera). J. Morph. 1934, 56, 267–293. [Google Scholar] [CrossRef]
- Grassé, P.-P. Ordre des Isoptère ou Termites. In Traité de Zoologie IX; Grassé, P.P., Ed.; Masson et cie: Paris, France, 1949; pp. 408–544. [Google Scholar]
- Buchli, H.R. L’origine des castes et les potentialités ontogéniques des termites européens du genre Reticulitermes Holmgren. Ann. Nat. Zool. 1958, 11, 263–429. [Google Scholar]
- Noirot, C.; Pasteels, J.M. The worker caste is polyphyletic in termites. Sociobiology 1988, 14, 15–20. [Google Scholar]
- Vieau, F. Les stades nymphaux 7 et 8 chez Reticulitermes santonensis Feytaud (Isoptera, Rhinotermitidae). Actes Colloq Insectes Soc. 1994, 9, 61–66. [Google Scholar]
- Thorne, B.L.; Traniello, J.F.A.; Adams, E.S.; Bulmer, M. Reproductive dynamics and colony structure of subterranean termites of the genus Reticulitermes (Isoptera Rhinotermitidae): A review of the evidence from behavioral, ecological, and genetic studies. Ethol. Ecol. Evol. 1999, 11, 149–169. [Google Scholar] [CrossRef]
- Snyder, T.E. Record of the finding of a true queen of termes flavipes KOL. Proc. Entomol. Soc. Wash. 1912, 14, 107–109. [Google Scholar]
- Myles, T.G. Review of secondary reproduction in termites (Insecta: Isoptera) with comments on its role in termite ecology and social evolution. Sociobiology 1999, 33, 1–91. [Google Scholar]
- Forschler, B.T.; Townsend, M.L. Mark-release-recapture estimates of Reticulitermes spp. (Isoptera: Rhinotermitidae) colony foraging populations from Georgia, USA. Environ. Entomol. 1996, 25, 952–962. [Google Scholar]
- Grube, S.; Forschler, B.T. Census of monogyne and polygyne laboratory colonies illuminates dynamics of population growth in Reticulitermes flavipes (Isoptera: Rhinotermitidae). Ann. Entomol. Soc. Am. 2004, 97, 466–475. [Google Scholar] [CrossRef]
- Thorne, B.L. Termite terminology. Sociobiology 1996, 28, 253–263. [Google Scholar]
- Zimet, M.; Stuart, A.M. Sexual dimorphism in the immature stages of the termite, Reticulitermes flavipes (Isoptera: Rhinotermitidae). Sociobiology 1982, 7, 1–7. [Google Scholar]
- Hu, J.; Forschler, B.T. Sample size requirement in the study of subterranean termite (Isoptera: Rhinotermitidae) sex ratio. J. Entomol. Sci. 2011, 46, 23–29. [Google Scholar]
- Leponce, M.; Rosin, Y.; Pasteels, J.M. Reproductive mechanisms and dynamics of habitat colonization in Mircocerotermes biroi (Ispotera: Termitidae). Ecol. Entomol. 1996, 21, 178–184. [Google Scholar] [CrossRef]
- Whitman, D.W.; Ananthakrishnan, T.N. What Is Phenotypic Plasticityand Why Is It Important? In Phenotypic Plasticity of Insects; Whitman, D.W., Ananthakrishnan, T.N., Eds.; Science Publishers: Enfield, NH, USA, 2009; pp. 1–64. [Google Scholar]
- Snyder, T.E. The origin of the castes in termites. Proc. Biol. Soc. Wash. 1925, 38, 57–68. [Google Scholar]
- Howard, R.W.; Haverty, M.I. Reproductives in mature colonies of Reticulitermes flavipes: Sex-ratio and association with soldiers. Environ. Entomol. 1980, 9, 458–460. [Google Scholar]
- Lenz, M.; Kard, B.; Evans, T.A.; Mauldin, J.K.; Etheridge, J.L.; Abbey, H.M. Differential use of identical food resources by Reticulitermes flavipes (Isoptera: Rhinotermitidae) in two types of habitats. Environ. Entomol. 2009, 38, 35–42. [Google Scholar] [CrossRef]
- Buchli, H. Die Neotenie bei Reticulitermes. Insectes Soc. 1956, 3, 131–143. [Google Scholar] [CrossRef]
- Long, C.E.; Thorne, B.L.; Breisch, N.L. Termite colony ontogeny: A long-term assessment of reproductive lifespan, caste ratios and colony size in Reticulitermes flavipes (Isoptera: Rhinotermitidae). Bull. Entomol. Res. 2003, 93, 439–445. [Google Scholar]
- Long, C.E.; Thorne, B.L.; Breisch, N.L. Termite colony ontogeny: Supplemental data in the long-term assessment of reproductive lifespan, female neotenic production and colony size in Reticulitermes flavipes (Isoptera: Rhinotermitidae). Bull. Entomol. Res. 2007, 97, 321–325. [Google Scholar] [CrossRef]
- Noirot, C. Formation of Castes in the Higher Termites. In Biology of Termites; Krishna, K., Weesner, F.M., Eds.; Academic Press: New York, NY, USA, 1969; pp. 311–350. [Google Scholar]
- Korb, J.; Katrantzis, S. Influence of environmental conditions on the expression of the sexual dispersal phenotype in a lower termite: Implications for the evolution of workers in termites. Evol. Dev. 2004, 6, 342–352. [Google Scholar] [CrossRef]
- Matsuura, K.; Fujimoto, M.; Goka, K. Sexual and asexual colony foundation and the mechanism of facultative parthenogenesis in the termite Reticulitermes speratus (Isoptera: Rhinotermitidae). Insectes Soc. 2004, 51, 325–332. [Google Scholar] [CrossRef]
- Hayashi, Y.; Lo, N.; Miyata, H.; Kitade, O. Sex-linked genetic influence on caste determination in a termite. Science 2007, 318, 985–987. [Google Scholar] [CrossRef]
- Matsuura, K.; Vargo, E.L.; Kawatsu, K.; Labadie, P.E.; Nakano, H.; Yashiro, T.; Tsuji, K. Queen succession through asexual reproduction in termites. Science 2009, 323, 1687–1687. [Google Scholar]
- Lo, N.; Hayashi, Y.; Kitade, O. Should environmental caste determination be assumed for termites? Am. Nat. 2009, 173, 848–853. [Google Scholar] [CrossRef]
- Vargo, E.L.; Husseneder, C. Biology of subterranean termites: Insights from molecular studies of Reticulitermes and Coptotermes. Annu. Rev. Entomol. 2009, 54, 379–403. [Google Scholar] [CrossRef]
- Lenz, M. Laboratory bioassay with subterranean termites (Isoptera)—The importance of termite biology. Sociobiology 2009, 53, 573–595. [Google Scholar]
- Husseneder, C.; Vargo, E.L.; Grace, J.K. Multilocus DNA fingerprinting and microsatellite genotyping: Complementary molecular approaches to investigating colony and population genetic structure in subterranean termites. Sociobiology 2002, 40, 217–226. [Google Scholar]
- Scharf, M.E.; Ratliff, C.R.; Wu-Scharf, D.; Zhou, X.; Pittendrigh, B.R.; Bennett, G.W. Effects of juvenile hormone III on Reticulitermes flavipes: Changes in hemolymph protein composition and gene expression. Insect Biochem. Mol. Biol. 2005, 35, 207–215. [Google Scholar] [CrossRef]
- Zhou, X.; Tarver, M.R.; Bennett, G.W.; Oi, F.M.; Scharf, M.E. Hexamerin genes from the termite Reticulitermes flavipes: Sequence, expression, and proposed functions in caste régulation. Gene 2006, 376, 47–58. [Google Scholar] [CrossRef]
- Esenther, G.R. Termites in Wisconsin. Ann. Entomol. Soc. Am. 1969, 62, 1274–1284. [Google Scholar]
- Greenberg, S.L.W.; Stuart, A.M. Precocious reproductive development (Neoteny) by larvae of a primitive termite Zootermopsis angusticollis (Hagen). Insectes Soc. 1982, 29, 535–547. [Google Scholar] [CrossRef]
- Lenz, M.; Runko, S. Long-term impact of orphaning on field colonies of Coptotermes lacteus (Froggatt) (Isoptera: Rhinotermitidae). Insectes Soc. 1993, 40, 439–456. [Google Scholar] [CrossRef]
- Dean, S.R.; Gold, R.E. Sex ratios and development of the reproductive system in castes of Reticulitermes flavipes (Kollar) (Isoptera: Rhinotermitidae). Ann. Entomol. Soc. Am. 2004, 97, 147–152. [Google Scholar] [CrossRef]
- Costa-Leonardo, A.M.; Arab, A.; Casarin, F.E. Neotenic formation in laboratory colonies of the termite Coptotermes gestroi after orphaning. J. Insect Sci. 2004, 4, 1–6. [Google Scholar]
- Miyata, H.; Furuichi, H.; Kitade, O. Patterns of neotenic differentiation in a subterranean termite, Reticulitermes speratus (Isoptera: Rhinotermitidae). Entomol. Sci. 2004, 7, 309–314. [Google Scholar] [CrossRef]
- Luykx, P. A cytogenetic survey of 25 species of lower termites from Australia. Genome 1990, 33, 80–88. [Google Scholar] [CrossRef]
- Bergamaschi, S.; Dawes-Gromadzki, T.Z.; Scali, V.; Marini, M.; Mantovani, B. Karyology, mitochondrial DNA and the phylogeny of Australian termites. Chromosome Res. 2007, 15, 735–753. [Google Scholar] [CrossRef]
- Martins, V. Karyotype evolution in the Termitidae (Isoptera). Sociobiology 1999, 34, 395–405. [Google Scholar]
- Roisin, Y.; Lenz, M. Origin of male-biased sex allocation in orphaned colonies of the termite, Coptotermes lacteus. Behav. Ecol. Sociobiol. 2002, 51, 472–479. [Google Scholar] [CrossRef]
- Jenkins, T.M.; Dean, R.E.; Verkerk, R.; Forschler, B.T. Phylogenetic analyses of two mitochondrial genes and one nuclear intron region illuminate European subterranean termite (Isoptera: Rhinotermitidae) gene flow, taxonomy, and introduction dynamic. Mol. Phylogenetics Evol. 2001, 20, 286–293. [Google Scholar] [CrossRef]
- Austin, J.W.; Szalanski, A.L.; Scheffrahn, R.H.; Messenger, M.T.; Dronnet, S.; Bagneres, A.-G. Genetic evidence for the synonymy of two Reticulitermes species: Reticulitermes flavipes and Reticulitermes santonensis. Ann. Entomol. Soc. Am. 2005, 98, 395–401. [Google Scholar] [CrossRef]
- Evans, T.A. Invasive Termites. In Biology of Termites: A Modern Synthesis; Bignell, D.E., Roisin, Y., Lo, N., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 519–562. [Google Scholar]
- Noirot, C. Pathways of Caste Development in the Lower Termites. In Caste Differentiation in Social Insects; Watson, J.A.L., Okot-Kotber, B.M., Noirot, C., Eds.; Pergamon Press: Oxford, UK, 1985; pp. 41–57. [Google Scholar]
- Vieau, F. Le termite de Saintonge: Un danger pour l‘Ouest de la France. Penn Bed 1991, 140, 19–32. [Google Scholar]
- Scharf, M.E.; Ratliff, C.R.; Hoteling, J.T.; Pittendrigh, B.R.; Bennett, G.W. Caste differentiation responses of two sympatric Reticulitermes termite species to juvenile hormone homologs. Insectes Soc. 2003, 50, 346–354. [Google Scholar] [CrossRef]
- Matsuura, K.; Himuro, C.; Yokoi, T.; Yamamoto, Y.; Vargo, E.L.; Keller, L. Identification of a pheromone regulating caste differentiation in a termite. Proc. Natl. Acad. Sci. USA 2010, 107, 12963–12968. [Google Scholar]
- Korb, J.; Lenz, M. Reproductive decision-making in the termite, Cryptotermes secundus (Kalotermitidae), under variable food conditions. Behav. Ecol. 2004, 15, 390–395. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hu, J.; Forschler, B.T. Neotenic Phenotype and Sex Ratios Provide Insight into Developmental Pathways in Reticulitermes flavipes (Isoptera: Rhinotermitidae). Insects 2012, 3, 538-552. https://doi.org/10.3390/insects3020538
Hu J, Forschler BT. Neotenic Phenotype and Sex Ratios Provide Insight into Developmental Pathways in Reticulitermes flavipes (Isoptera: Rhinotermitidae). Insects. 2012; 3(2):538-552. https://doi.org/10.3390/insects3020538
Chicago/Turabian StyleHu, Jian, and Brian T. Forschler. 2012. "Neotenic Phenotype and Sex Ratios Provide Insight into Developmental Pathways in Reticulitermes flavipes (Isoptera: Rhinotermitidae)" Insects 3, no. 2: 538-552. https://doi.org/10.3390/insects3020538