Polydnaviruses of Parasitic Wasps: Domestication of Viruses To Act as Gene Delivery Vectors


Abstract
:1. Introduction
2. PDV Distribution, Origins and Function
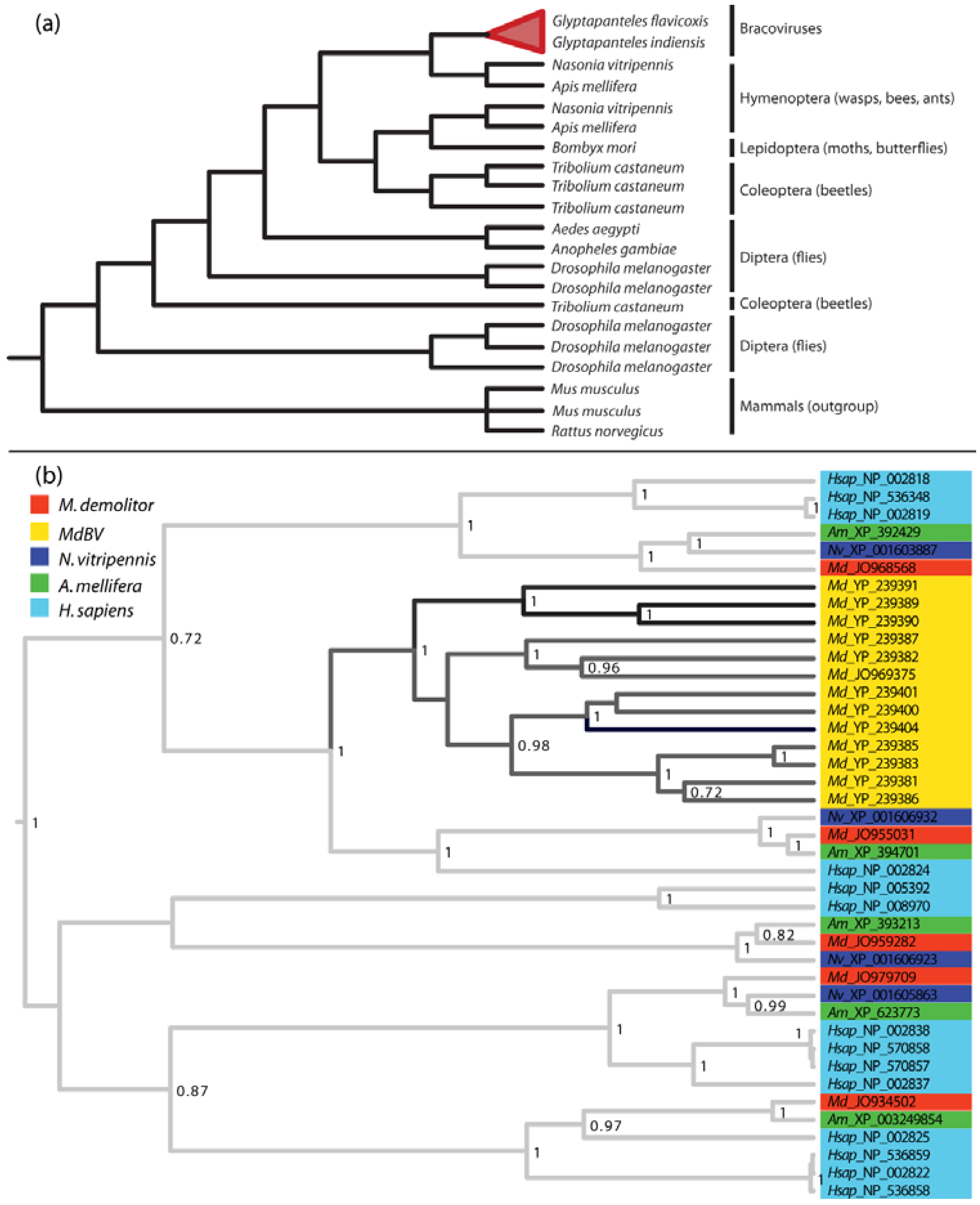
2.1. PDVs Are Associated with Wasps in Two Families
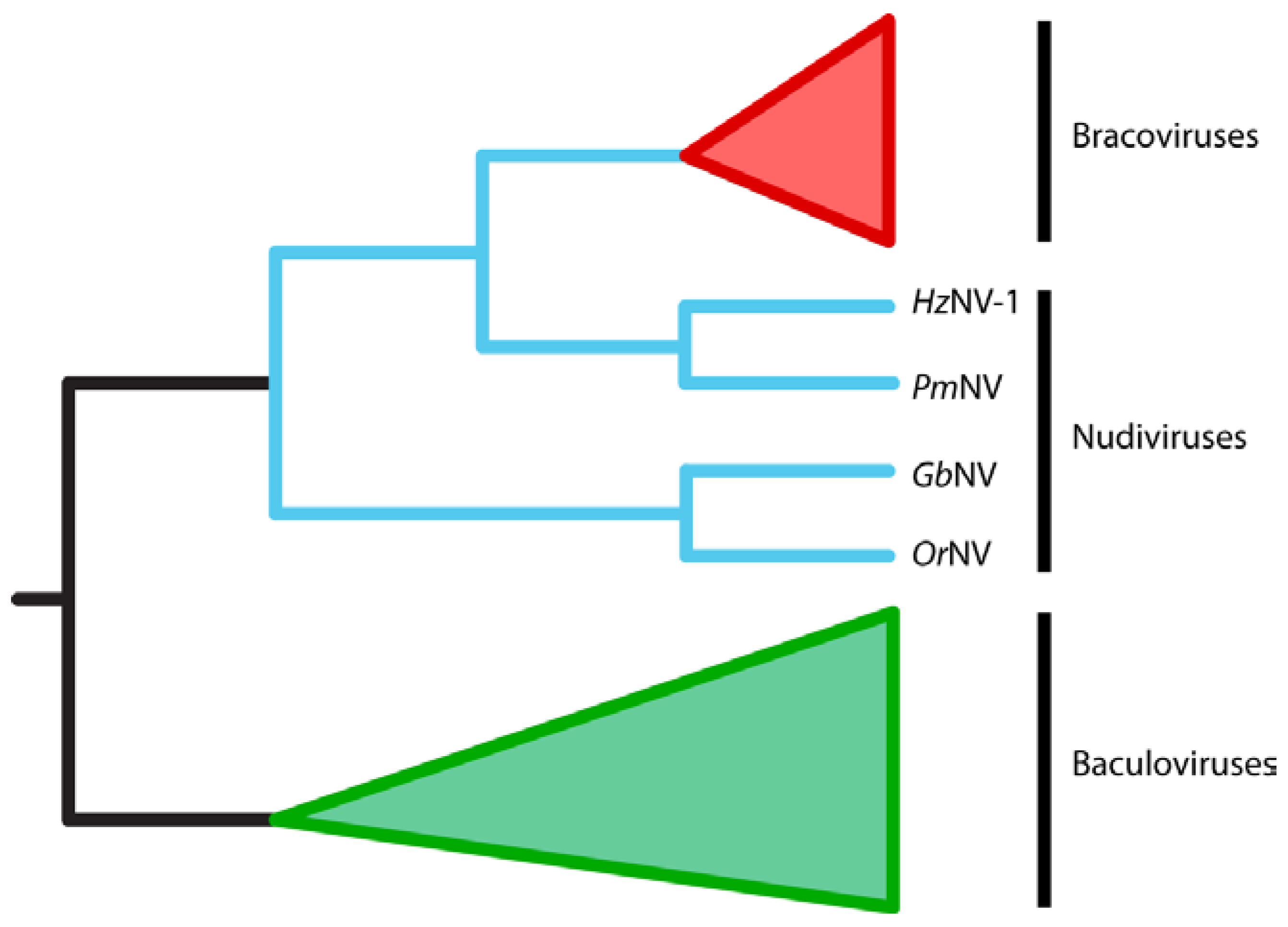
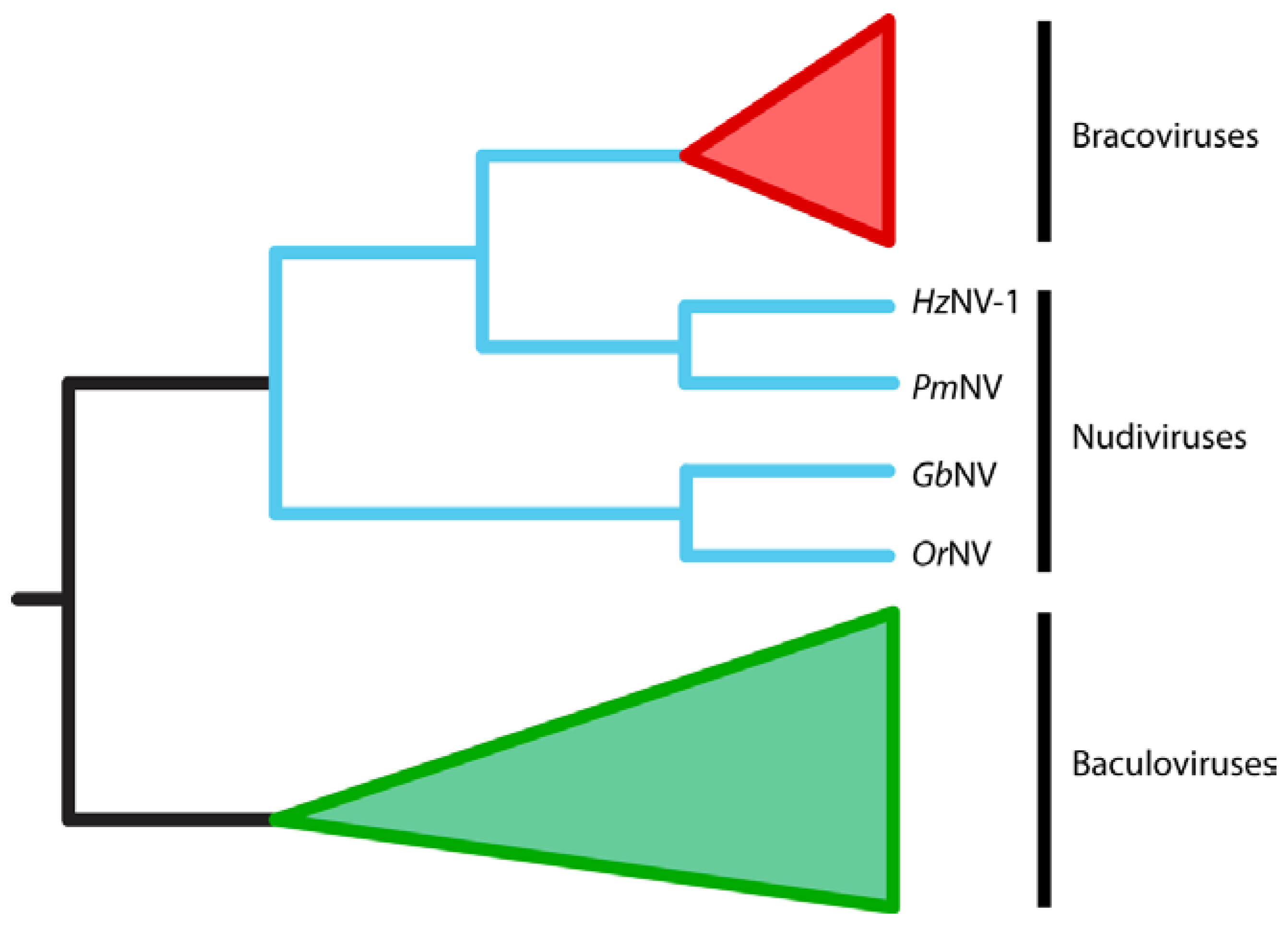
2.3. BVs Evolved from a Nudivirus Ancestor
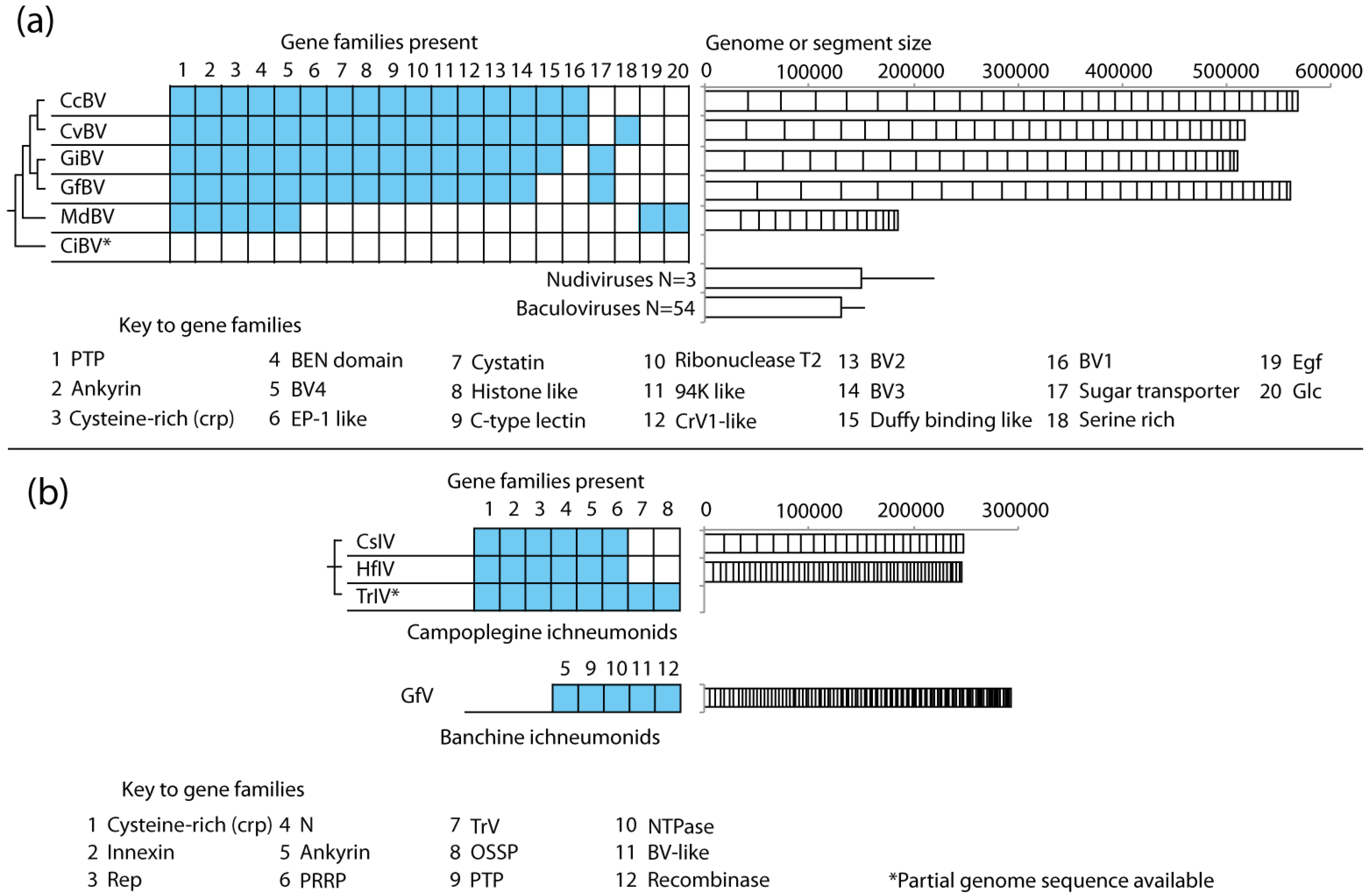
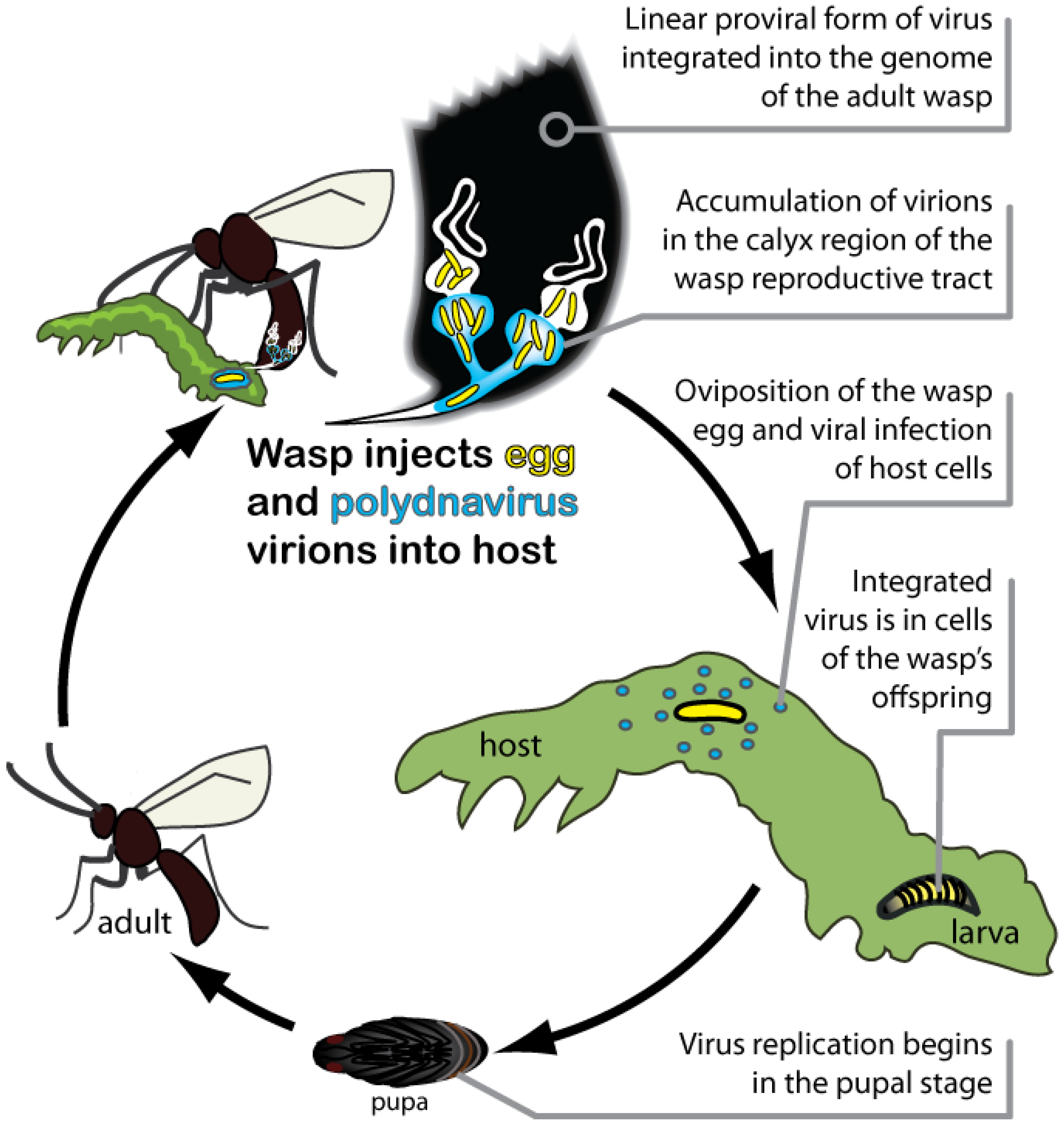
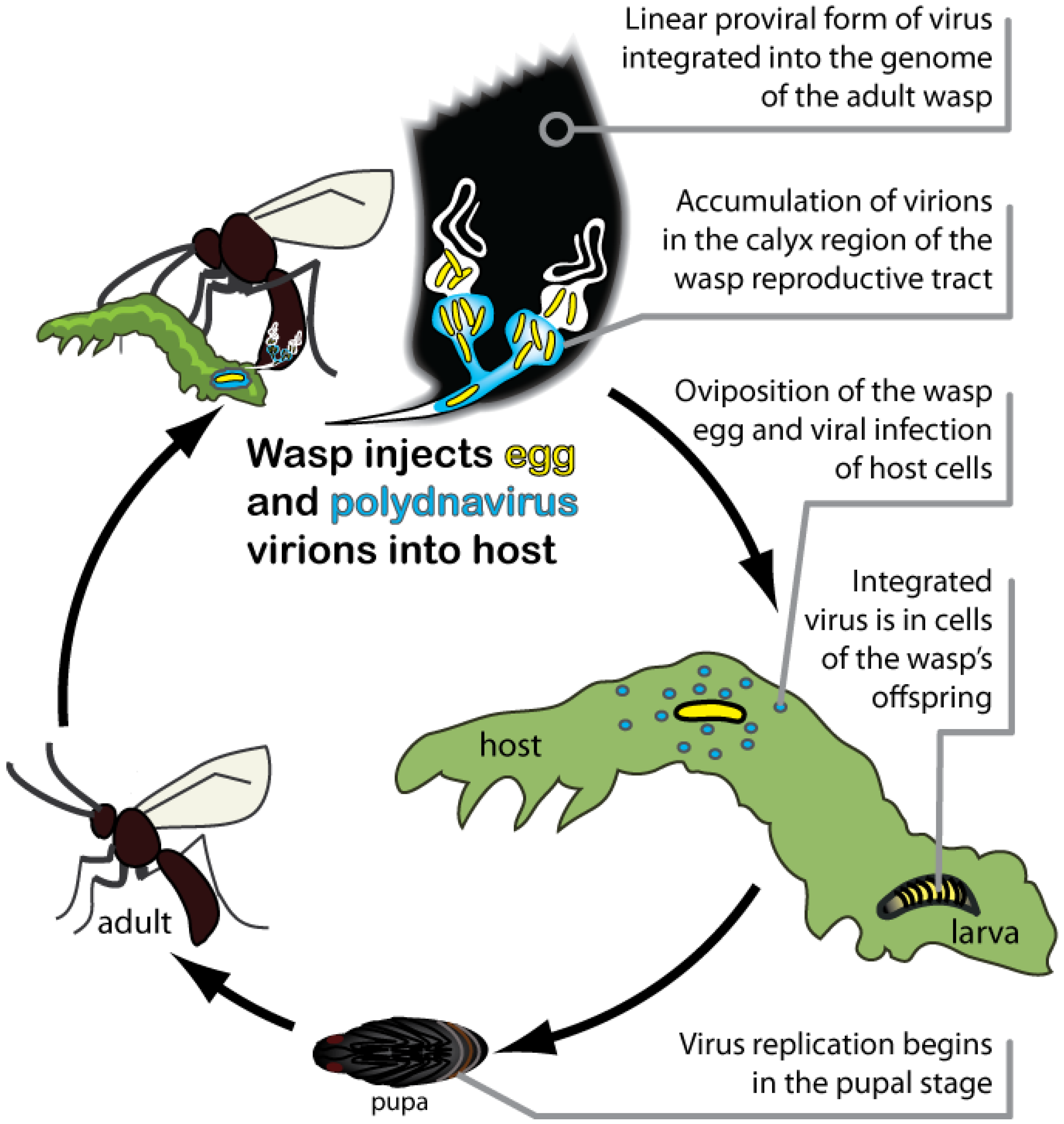
2.4. PDVs Are Essential for Successful Parasitism of Hosts by Wasps
3. PDVs as Domesticated Extensions of Parasitic Wasps
3.1. BVs Likely Evolved from a Pathogenic Nudivirus
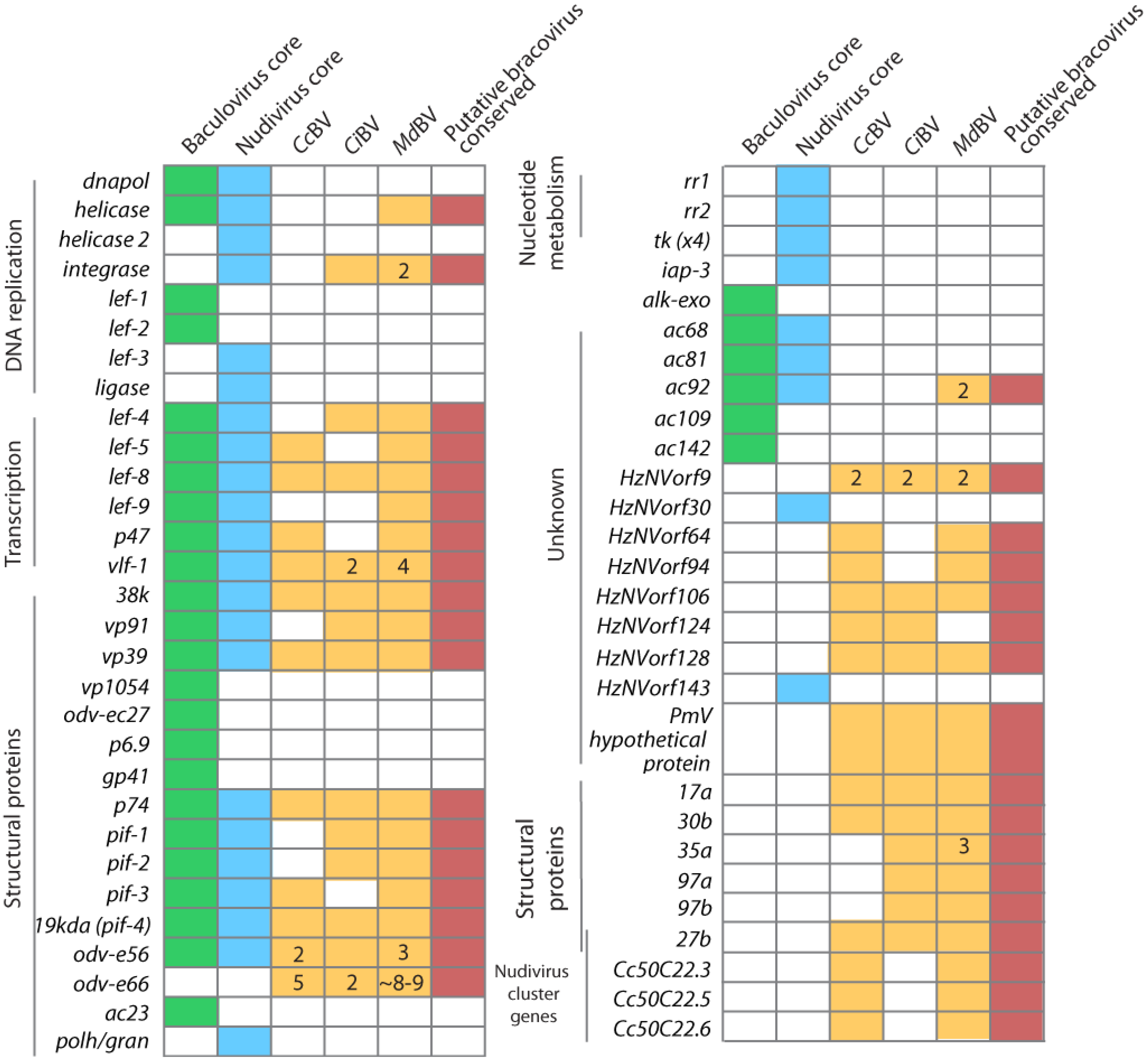
3.2. BVs Were Domesticated by Immobilizing Virus Replication Genes in the Wasp Genome

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism and gene set | G+C% | N * |
| M. demolitor transcripts | 34.0 | 66,298 transcripts (32674 loci) |
| M. demolitor nudivirus-like | 31.5 | 133 transcripts (41 loci) |
| C. congregata nudivirus-like | 34.0 | 20 |
| C. inanitus nudivirus-like | 31.4 | 18 |
| HzNV -1 all coding sequences | 45.6 | 154 |
| HzNV -1 genes conserved in BVs | 47.2 | 22 |
3.3. Consequences of Domestication upon Mechanisms of Virus Replication
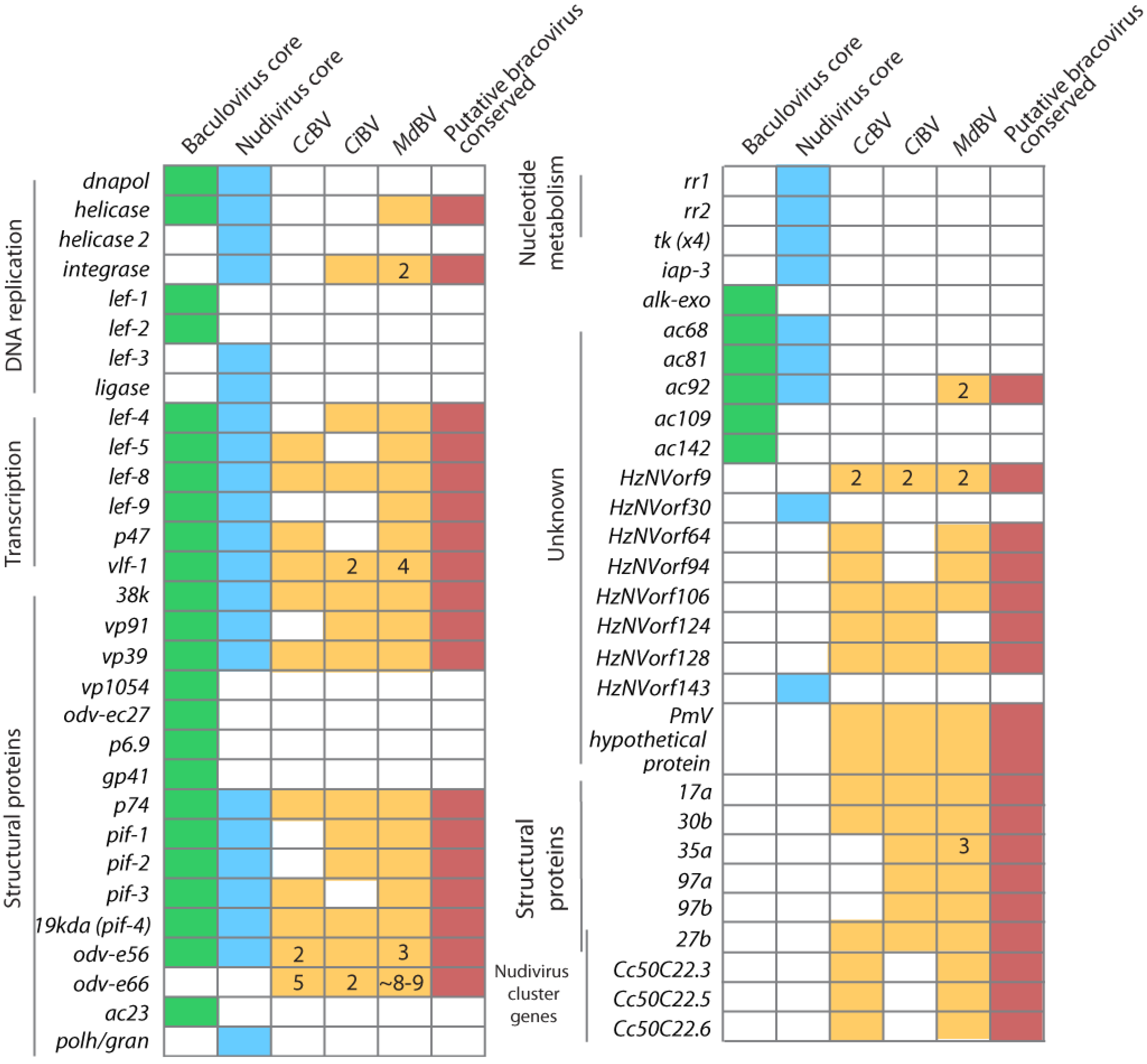
3.4. Wasps Control PDV Replication
3.5. Wasp Genes with Potential Roles in Regulating Replication of Viral DNA
3.6. Regulation of PDV Virulence Gene Expression and Segment Proliferation in Wasps
3.7. Consequences of Virus Domestication on the Rate of Genome Evolution
4. PDVs as Wasp Gene Delivery Systems
4.1. BVs Likely Use Cell Entry Mechanisms Similar to Those of Baculoviruses
4.2. The Encapsidated Genomes of PDVs Have Insect-Like Architectural Features
| Group | Genome | AT content | Introns (% of genes) | Coding density |
|---|---|---|---|---|
| BV genomes | CcBV | 66 | 69 | 27 |
| CpBV | 65 | 41 | 32 | |
| CvBV | 66 | 57 | 27 | |
| GiBV | 64 | 58 | 33 | |
| GfBV | 64 | 63 | 33 | |
| MdBV | 66 | 14 | 17 | |
| CiBV | NA | NA | NA | |
| BV relatives | HzNV-1 | 58 | 0 | 67 |
| AcMNPV | 59 | 0 | 94 | |
| IV genomes | ||||
| Campoplegine IVs | CsIV | 59 | 10 | 29 |
| HfIV | 57 | NA | 30 | |
| TrIV | 58 | NA | 22 | |
| Banchine IVs | GfV | 63 | Near 0 | 20 |
| Hymenopteran genomes | N. vitripennis | 59 | 5 introns/gene | 14 |
| A. mellifera | 67 | 5.8 introns/gene | 12 |
4.3. Most Genes in the Encapsidated Genome of PDVs form Multimember Families
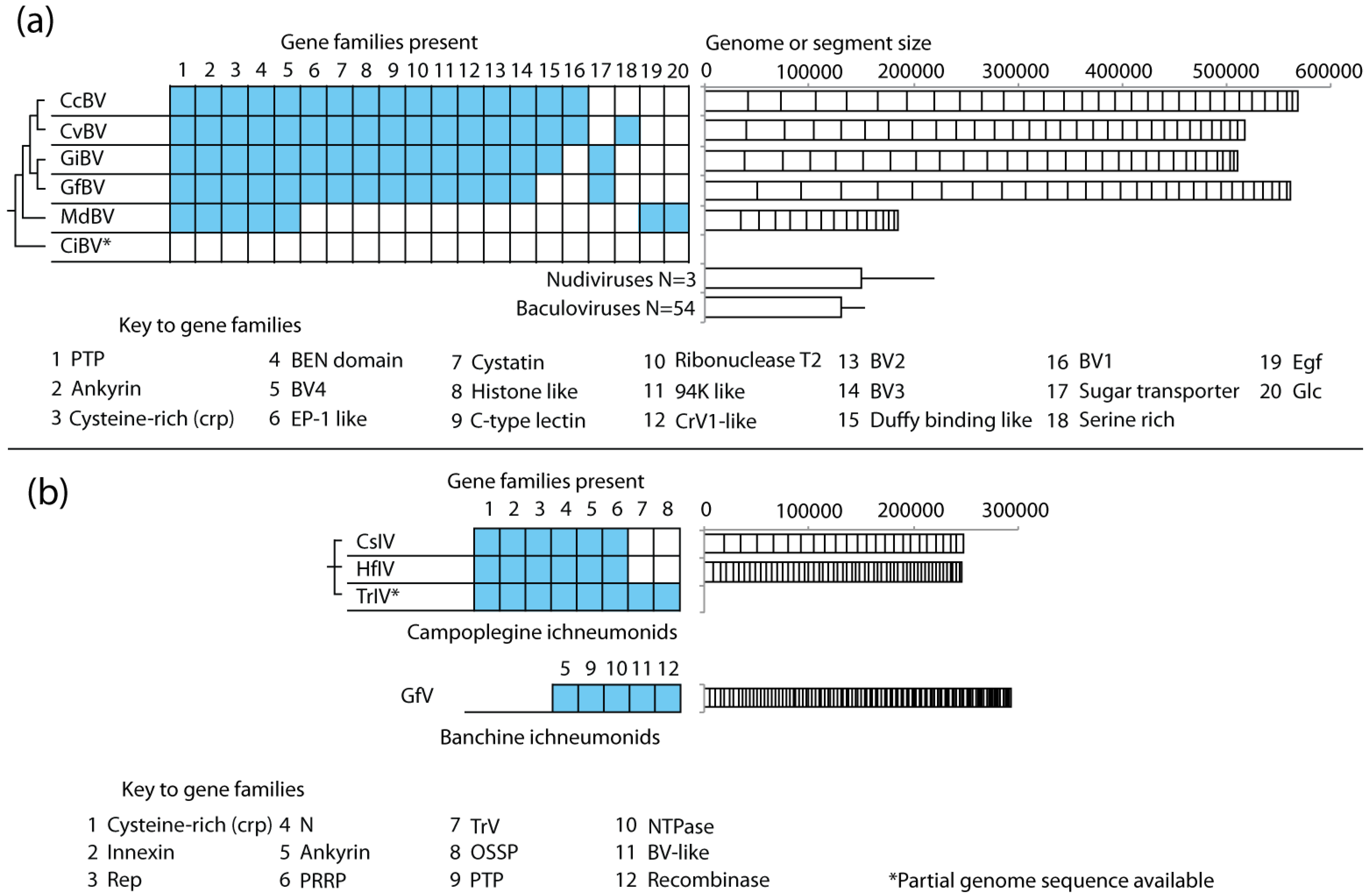
4.4. PDV Gene Families Have Diverse Origins
4.5. Genome Size and Gene Content Are Highly Dynamic in PDVs
4.6. DNA Segments of the Encapsidated Genomes Integrate into Host Cells
5. Concluding Remarks
Acknowledgements
References
- Moran, N.A. Symbiosis. Curr. Biol. 2006, 16, R866–R871. [Google Scholar] [CrossRef]
- Newton, I.L.; Woyke, T.; Auchtung, T.A.; Dilly, G.F.; Dutton, R.J.; Fisher, M.C.; Fontanez, K.M.; Lau, E.; Stewart, F.J.; Richardson, P.M.; et al. The Calyptogena magnifica chemoautotrophic symbiont genome. Science 2007, 315, 998–1000. [Google Scholar]
- Baumann, P. Biology bacteriocyte-associated endosymbionts of plant sap-sucking insects. Annu. Rev. Microbiol. 2005, 59, 155–189. [Google Scholar] [CrossRef]
- Aylward, F.O.; Currie, C.R.; Suen, G. The evolutionary innovation of nutritional symbiosis in leaf-cutter ants. Insects 2012, 3, 41–61. [Google Scholar] [CrossRef]
- Nowack, E.C.; Melkonian, M. Endosymbiotic associations within protists. Philos. Trans. R. Soc. B 2010, 365, 699–712. [Google Scholar] [CrossRef]
- Strand, M.R. Polydnaviruses. In Insect Virology; Asgari, S., Johnson, K.N., Eds.; Caister Academic Press: Norwich, UK, 2010; pp. 171–197. [Google Scholar]
- Webb, B.A.; Strand, M.R. The Biology and Genomics of Polydnaviruses. In Comprehensive Molecular Insect Science; Iatrou, K., Gill, S., Eds.; Elsevier Pergamon: Amsterdam, the Netherlands, 2005; Volume 6, pp. 323–360. [Google Scholar]
- Moreira, D.; Lopez-Garcia, P. Ten reasons to exclude viruses from the tree of life. Nat. Rev. Microbiol. 2009, 7, 306–311. [Google Scholar]
- Whitfield, J.B. Phylogeny of the non-aculeate Apocrita and the evolution of parasitism in the Hymenoptera. J. Hymenopt. Res. 1992, 1, 3–14. [Google Scholar]
- Grimaldi, D.A.; Engel, M.S. Evolution of the Insects; Cambridge University Press: New York, NY, USA, 2005. [Google Scholar]
- Whitfield, J.B. Phylogeny and evolution of host-parasitoid interactions in hymenoptera. Annu. Rev. Entomol. 1998, 43, 129–151. [Google Scholar] [CrossRef]
- Whitfield, J.B. Estimating the age of the polydnavirus/braconid wasp symbiosis. Proc. Natl. Acad. Sci. USA 2002, 99, 7508–7513. [Google Scholar] [CrossRef]
- Jones, O.W.; Purvis, A.; Baumgart, E.; Quicke, D.L.J. Using taxonomic revision data to estimate the geographic and taxonomic distribution of undescribed species richness in the Braconidae (Hymenoptera: Ichneumonoidea). Insect Conserv. Diver. 2009, 2, 204–212. [Google Scholar] [CrossRef]
- Murphy, N.; Banks, J.C.; Whitfield, J.B.; Austin, A.D. Phylogeny of the parasitic microgastroid subfamilies (Hymenoptera: Braconidae) based on sequence data from seven genes, with an improved time estimate of the origin of the lineage. Mol. Phylogenet. Evol. 2008, 47, 378–395. [Google Scholar] [CrossRef]
- Stoltz, D.B.; Vinson, S.B. Viruses and parasitism in insects. Adv. Virus Res. 1979, 24, 125–171. [Google Scholar] [CrossRef]
- Albrecht, U.; Wyler, T.; Pfister-Wilhelm, R.; Gruber, A.; Stettler, P.; Heiniger, P.; Kurt, E.; Schumperli, D.; Lanzrein, B. Polydnavirus of the parasitic wasp Chelonus inanitus (Braconidae): Characterization, genome organization and time point of replication. J. Gen. Virol. 1994, 75, 3353–3363. [Google Scholar] [CrossRef]
- Wyder, S.; Tschannen, A.; Hochuli, A.; Gruber, A.; Saladin, V.; Zumbach, S.; Lanzrein, B. Characterization of Chelonus inanitus polydnavirus segments: Sequences and analysis, excision site and demonstration of clustering. J. Gen. Virol. 2002, 83, 247–256. [Google Scholar]
- Wyler, T.; Lanzrein, B. Ovary development and polydnavirus morphogenesis in the parasitic wasp Chelonus inanitus. II. Ultrastructural analysis of calyx cell development, virion formation and release. J. Gen. Virol. 2003, 84, 1151–1163. [Google Scholar] [CrossRef]
- Norton, W.N.; Vinson, S.B. Correlating the initiation of virus replication with a specific pupal developmental phase of an ichneumonid parasitoid. Cell Tissue Res. 1983, 231, 387–398. [Google Scholar]
- Kroemer, J.A.; Webb, B.A. Polydnavirus genes and genomes: Emerging gene families and new insights into polydnavirus replication. Annu. Rev. Entomol. 2004, 49, 431–456. [Google Scholar] [CrossRef]
- Beck, M.H.; Inman, R.B.; Strand, M.R. Microplitis demolitor bracovirus genome segments vary in abundance and are individually packaged in virions. Virology 2007, 359, 179–189. [Google Scholar] [CrossRef]
- Webb, B.A. Polydnavirus Biology, Genome Structure, and Evolution. In The Insect Viruses; Fraenkel-Conrat, H., Wagner, R.R., Eds.; Plenum Press: New York, NY, USA, 1998; pp. 105–139. [Google Scholar]
- Choi, J.H.; Kwon, S.J.; Roh, J.Y.; Yang, T.J.; Yoon, S.H.; Kim, H.; Li, M.S.; Park, B.S.; Woo, S.D.; Jin, B.R.; et al. Sequence and gene organization of 24 circles from the Cotesia plutellae bracovirus genome. Arch. Virol. 2009, 154, 1313–1327. [Google Scholar] [CrossRef]
- Desjardins, C.A.; Gundersen-Rindal, D.E.; Hostetler, J.B.; Tallon, L.J.; Fadrosh, D.W.; Fuester, R.W.; Pedroni, M.J.; Haas, B.J.; Schatz, M.C.; Jones, K.M.; et al. Comparative genomics of mutualistic viruses of Glyptapanteles parasitic wasps. Genome Biol. 2008, 9, R183. [Google Scholar] [CrossRef]
- Espagne, E.; Dupuy, C.; Huguet, E.; Cattolico, L.; Provost, B.; Martins, N.; Poirié, M.; Periquet, G.; Drezen, J.M. Genome sequence of a polydnavirus: insights into symbiotic virus evolution. Science 2004, 306, 286–289. [Google Scholar]
- Webb, B.A.; Strand, M.R.; Dickey, S.E.; Beck, M.H.; Hilgarth, R.S.; Barney, W.E.; Kadash, K.; Kroemer, J.A.; Lindstrom, K.G.; Rattanadechakul, W.; et al. Polydnavirus genomes reflect their dual roles as mutualists and pathogens. Virology 2006, 347, 160–174. [Google Scholar] [CrossRef]
- Weber, B.; Annaheim, M.; Lanzrein, B. Transcriptional analysis of polydnaviral genes in the course of parasitization reveals segment-specific patterns. Arch. Insect Biochem. 2007, 66, 9–22. [Google Scholar] [CrossRef]
- Chen, Y.F.; Gao, F.; Ye, X.Q.; Wei, S.J.; Shi, M.; Zheng, H.J.; Chen, X.X. Deep sequencing of Cotesia vestalis bracovirus reveals the complexity of a polydnavirus genome. Virology 2011, 414, 42–50. [Google Scholar] [CrossRef]
- Lapointe, R.; Tanaka, K.; Barney, W.E.; Whitfield, J.B.; Banks, J.C.; Beliveau, C.; Stoltz, D.; Webb, B.A.; Cusson, M. Genomic and morphological features of a banchine polydnavirus: comparison with bracoviruses and ichnoviruses. J. Virol. 2007, 81, 6491–6501. [Google Scholar]
- Tanaka, K.; Lapointe, R.; Barney, W.E.; Makkay, A.M.; Stoltz, D.; Cusson, M.; Webb, B.A. Shared and species-specific features among ichnovirus genomes. Virology 2007, 363, 26–35. [Google Scholar] [CrossRef]
- Dupuy, C.; Gundersen-Rindal, D.; Cusson, M. Genomics and Replication of Polydnaviruses. In Parasitoid Viruses; Beckage, N.E., Drezen, J.M., Eds.; Academic Press: London, UK, 2011; pp. 47–62. [Google Scholar]
- Stoltz, D.B.; Vinson, S.B. Baculovirus-like particles in the reproductive tracts of female parasitoid wasps. II. The genus Apanteles. Can. J. Microbiol. 1977, 23, 28–37. [Google Scholar] [CrossRef]
- Stoltz, D.B.; Vinson, S.B.; MacKinnon, E.A. Baculovirus-like particles in the reproductive tracts of female parasitoid wasps. Can. J. Microbiol. 1976, 22, 1013–1023. [Google Scholar] [CrossRef]
- Federici, B.A.; Bigot, Y. Origin and evolution of polydnaviruses by symbiogenesis of insect DNA viruses in endoparasitic wasps. J. Insect Physiol. 2003, 49, 419–432. [Google Scholar] [CrossRef]
- Bigot, Y.; Samain, S.; Auge-Gouillou, C.; Federici, B.A. Molecular evidence for the evolution of ichnoviruses from ascoviruses by symbiogenesis. BMC Evol. Biol. 2008, 8, 253:1–253:12. [Google Scholar]
- Bèzier, A.; Annaheim, M.; Herbinière, J.; Wetterwald, C.; Gyapay, G.; Bernard-Samain, S.; Wincker, P.; Roditi, I.; Heller, M.; Belghazi, M.; et al. Polydnaviruses of braconid wasps derive from an ancestral nudivirus. Science 2009, 323, 926–930. [Google Scholar]
- Jehle, J.A. Nudiviruses. In Insect Virology; Asgari, S., Johnson, K.N., Eds.; Caister Academic Press: Norwich, UK, 2010; pp. 153–170. [Google Scholar]
- Theze, J.; Bèzier, A.; Periquet, G.; Drezen, J.M.; Herniou, E.A. Paleozoic origin of insect large dsDNA viruses. Proc. Natl. Acad. Sci. USA 2011, 108, 15931–15935. [Google Scholar]
- Volkoff, A.N.; Jouan, V.; Urbach, S.; Samain, S.; Bergoin, M.; Wincker, P.; Demettre, E.; Cousserans, F.; Provost, B.; Coulibaly, F.; et al. Analysis of virion structural components reveals vestiges of the ancestral ichnovirus genome. PLoS Pathog. 2010, 6, e1000923. [Google Scholar]
- Cerenius, L.; Soderhall, K. The prophenoloxidase-activating system in invertebrates. Immunol. Rev. 2004, 198, 116–126. [Google Scholar] [CrossRef]
- Theopold, U.; Schmidt, O.; Soderhall, K.; Dushay, M.S. Coagulation in arthropods: defence, wound closure and healing. Trends Immunol. 2004, 25, 289–294. [Google Scholar] [CrossRef]
- Imler, J.L.; Bulet, P. Antimicrobial peptides in Drosophila: Structures, activities and gene regulation. Chem. Immunol. Allergy 2005, 86, 1–21. [Google Scholar] [CrossRef]
- Kanost, M.R.; Gorman, M.J. Phenoloxidases in Insect Immunity. In Insect Immunity; Beckage, N.E., Ed.; Academic Press: San Diego, CA, USA, 2008; pp. 69–96. [Google Scholar]
- Strand, M.R. Insect Hemocytes and Their Role in Immunity. In Insect Immunity; Beckage, N.E., Ed.; Academic Press: San Diego, CA, USA, 2008; pp. 25–47. [Google Scholar]
- Strand, M.R.; Pech, L.L. Immunological basis for compatibility in parasitoid-host relationships. Annu. Rev. Entomol. 1995, 40, 31–56. [Google Scholar] [CrossRef]
- Strand, M.R. Bracoviruses (Polydnaviridae). In The Springer Index of Viruses; Tidona, C.A., Darai, G., Lange, R., Eds.; Springer: Berlin, Germany, 2001; pp. 820–825. [Google Scholar]
- Moreau, S.J.M.; Huguet, E.; Drezen, J.M. Polydnaviruses as Tools to Deliver Wasp Virulence Factors to Impair Lepidopteran Host Immunity. In Insect Infection and Immunity: Evolution, Ecology and Mechanisms; Reynolds, S., Ed.; Oxford University Press: Oxford, UK, 2009; pp. 137–158. [Google Scholar]
- Pech, L.L.; Trudeau, D.; Strand, M.R. Effects of basement membranes on the behavior of hemocytes from Pseudoplusia includens: Development of an in vitro encapsulation assay. J. Insect Physiol. 1995, 41, 801–807. [Google Scholar] [CrossRef]
- Pennacchio, F.; Strand, M.R. Evolution of developmental strategies in parasitic hymenoptera. Annu. Rev. Entomol. 2006, 51, 233–258. [Google Scholar] [CrossRef]
- Strand, M.R. Microplitis demolitor polydnavirus infects and expresses in specific morphotypes of Pseudoplusia includens haemocytes. J. Gen. Virol. 1994, 75, 3007–3020. [Google Scholar] [CrossRef]
- Beck, M.H.; Strand, M.R. A novel polydnavirus protein inhibits the insect prophenoloxidase activation pathway. Proc. Natl. Acad. Sci. USA 2007, 104, 19267–19272. [Google Scholar]
- Beliveau, C.; Laforge, M.; Cusson, M.; Bellemare, G. Expression of a Tranosema rostrale polydnavirus gene in the spruce budworm, Choristoneura fumiferana. J. Gen. Virol. 2000, 81, 1871–1880. [Google Scholar]
- Galibert, L.; Rocher, J.; Ravallec, M.; Duonor-Cerutti, M.; Webb, B.A.; Volkoff, A.N. Two Hyposoter didmator ichnovirus genes expressed in the lepidopteran host encode secreted or membrane-associated serine and threonine rich proteins in segments that may be nested. J. Insect Physiol. 2003, 49, 441–451. [Google Scholar] [CrossRef]
- Yin, L.; Zhang, C.; Qin, J.; Wang, C. Polydnavirus of Campoletis chlorideae: Characterization and temporal effect on host Helicoverpa armigera cellular immune response. Arch. Insect Biochem. 2003, 52, 104–113. [Google Scholar] [CrossRef]
- Lanzrein, B.; Pfister-Wilhelm, R.; Kaeslin, M.; Wespi, G.; Roth, T. The orchestrated manipulation of the host by Chelonus inanitus and its polydnavirus. In Parasitoid viruses; Beckage, N.E., Drezen, J.M., Eds.; Academic Press: London, UK, 2012; pp. 169–178. [Google Scholar]
- Beckage, N.E. Polydnaviruses as Endocrine Regulators. In Parasitoid Viruses; Beckage, N.E., Drezen, J.M., Eds.; Academic Press: London, UK, 2012; pp. 163–168. [Google Scholar]
- Rohrmann, G.F. Baculovirus Molecular Biology, 2nd ed.; National Library of Medicine, National Center for Biotechnology Information: Bethesda, MD, USA, 2011. [Google Scholar]
- Chao, Y.C.; Lee, S.T.; Chang, M.C.; Chen, H.H.; Chen, S.S.; Wu, T.Y.; Liu, F.H.; Hsu, E.L.; Hou, R.F. A 2.9-kilobase noncoding nuclear RNA functions in the establishment of persistent Hz-1 viral infection. J. Virol. 1998, 72, 2233–2245. [Google Scholar]
- Lin, C.L.; Lee, J.C.; Chen, S.S.; Wood, H.A.; Li, M.L.; Li, C.F.; Chao, Y.C. Persistent Hz-1 virus infection in insect cells: Evidence for insertion of viral DNA into host chromosomes and viral infection in a latent status. J. Virol. 1999, 73, 128–139. [Google Scholar]
- Burand, J.P. The sexually transmitted insect virus, Hz-2V. Virol. Sin. 2009, 24, 428–435. [Google Scholar] [CrossRef]
- Annaheim, M.; Lanzrein, B. Genome organization of the Chelonus inanitus polydnavirus: Excision sites, spacers and abundance of proviral and excised segments. J. Gen. Virol. 2007, 88, 450–457. [Google Scholar] [CrossRef]
- Beck, M.H.; Zhang, S.; Bitra, K.; Burke, G.R.; Strand, M.R. The encapsidated genome of Microplitis demolitor bracovirus integrates into the host Pseudoplusia includens. J. Virol. 2011, 85, 11685–11696. [Google Scholar]
- Cheng, C.H.; Liu, S.M.; Chow, T.Y.; Hsiao, Y.Y.; Wang, D.P.; Huang, J.J.; Chen, H.H. Analysis of the complete genome sequence of the Hz-1 virus suggests that it is related to members of the Baculoviridae. J. Virol. 2002, 76, 9024–9034. [Google Scholar]
- Burke, G.R.; Strand, M.R. Deep sequencing identifies viral and wasp genes with potential roles in replication of Microplitis demolitor bracovirus. J. Virol. 2012. accepted. [Google Scholar]
- Wu, Y.L.; Wu, C.P.; Lee, S.T.; Tang, H.; Chang, C.H.; Chen, H.H.; Chao, Y.C. The early gene hhi1 reactivates Heliothis zea nudivirus 1 in latently infected cells. J. Virol. 2010, 84, 1057–1065. [Google Scholar] [CrossRef]
- Herniou, E.A.; Olszewski, J.A.; Cory, J.S.; O’Reilly, D.R. The genome sequence and evolution of baculoviruses. Annu. Rev. Entomol. 2003, 48, 211–234. [Google Scholar] [CrossRef]
- Wang, Y.; Jehle, J.A. Nudiviruses and other large, double-stranded circular DNA viruses of invertebrates: new insights on an old topic. J. Invertebr. Pathol. 2009, 101, 187–193. [Google Scholar] [CrossRef]
- Wetterwald, C.; Roth, T.; Kaeslin, M.; Annaheim, M.; Wespi, G.; Heller, M.; Mäser, P.; Roditi, I.; Pfister-Wilhelm, R.; Bèzier, A.; et al. Identification of bracovirus particle proteins and analysis of their transcript levels at the stage of virion formation. J. Gen. Virol. 2010, 91, 2610–2619. [Google Scholar]
- Huang, N.; Clem, R.J.; Rohrmann, G.F. Characterization of cDNAs encoding p53 of Bombyx mori and Spodoptera frugiperda. Insect Biochem. Mol. 2011, 41, 613–619. [Google Scholar] [CrossRef]
- Marti, D.; Grossniklaus-Burgin, C.; Wyder, S.; Wyler, T.; Lanzrein, B. Ovary development and polydnavirus morphogenesis in the parasitic wasp Chelonus inanitus. I. Ovary morphogenesis, amplification of viral DNA and ecdysteroid titres. J. Gen. Virol. 2003, 84, 1141–1150. [Google Scholar] [CrossRef]
- Tower, J. Developmental gene amplification and origin regulation. Annu. Rev. Genet. 2004, 38, 273–304. [Google Scholar] [CrossRef]
- Pasquier-Barre, F.; Dupuy, C.; Huguet, E.; Monerio, F.; Moreau, A.; Poirié, M.; Drezen, J.M. Polydnavirus replication: the EP1 segment of the parasitoid wasp Cotesia congregata is amplified within a larger precursor molecule. J. Gen. Virol. 2002, 83, 2035–2045. [Google Scholar]
- Savary, S.; Beckage, N.; Tan, F.; Periquet, G.; Drezen, J.M. Excision of the polydnavirus chromosomal integrated EP1 sequence of the parasitoid wasp Cotesia congregata (Braconidae, Microgastinae) at potential recombinase binding sites. J. Gen. Virol. 1997, 78, 3125–3134. [Google Scholar]
- Savary, S.; Drezen, J.M.; Tan, F.; Beckage, N.E.; Periquet, G. The excision of polydnavirus sequences from the genome of the wasp Cotesia congregata (Braconidae, microgastrinae) is developmentally regulated but not strictly restricted to the ovaries in the adult. Insect Mol. Biol. 1999, 8, 319–327. [Google Scholar]
- Bitra, K.; Zhang, S.; Strand, M.R. Transcriptomic profiling of Microplitis demolitor bracovirus reveals host, tissue and stage-specific patterns of activity. J. Gen. Virol. 2011, 92, 2060–2071. [Google Scholar] [CrossRef]
- Provost, B.; Varricchio, P.; Arana, E.; Espagne, E.; Falabella, P.; Huguet, E.; La Scaleia, R.; Cattolico, L.; Poirié, M.; Malva, C.; et al. Bracoviruses contain a large multigene family coding for protein tyrosine phosphatases. J. Virol. 2004, 78, 13090–13103. [Google Scholar]
- Deng, L.; Webb, B.A. Cloning and expression of a gene encoding a Campoletis sonorensis polydnavirus structural protein. Arch. Insect Biochem. 1999, 40, 30–40. [Google Scholar] [CrossRef]
- Theilmann, D.A.; Summers, M.D. Molecular analysis of Campoletis sonorensis virus DNA in the lepidopteran host Heliothis virescens. J. Gen. Virol. 1986, 67, 1961–1969. [Google Scholar] [CrossRef]
- Hilgarth, R.S.; Webb, B.A. Characterization of Campoletis sonorensis ichnovirus segment I genes as members of the repeat element gene family. J. Gen. Virol. 2002, 83, 2393–2402. [Google Scholar]
- Turnbull, M.; Webb, B. Perspectives on polydnavirus origins and evolution. Adv. Virus Res. 2002, 58, 203–254. [Google Scholar] [CrossRef]
- Rasoolizadeh, A.; Beliveau, C.; Stewart, D.; Cloutier, C.; Cusson, M. Tranosema rostrale ichnovirus repeat element genes display distinct transcriptional patterns in caterpillar and wasp hosts. J. Gen. Virol. 2009, 90, 1505–1514. [Google Scholar] [CrossRef]
- Brookfield, J.F. Host-parasite relationships in the genome. BMC Biol. 2011, 9, 67:1–67:3. [Google Scholar]
- Stoltz, D.B. Evidence for chromosomal transmission of polydnavirus DNA. J. Gen. Virol. 1990, 71, 1051–1056. [Google Scholar] [CrossRef]
- Stoltz, D.B.; Xu, D. Polymorphism in polydnavirus genomes. Can. J. Microbiol. 1990, 36, 538–543. [Google Scholar] [CrossRef]
- Keightley, P.D.; Trivedi, U.; Thomson, M.; Oliver, F.; Kumar, S.; Blaxter, M.L. Analysis of the genome sequences of three Drosophila melanogaster spontaneous mutation accumulation lines. Genome Res. 2009, 19, 1195–1201. [Google Scholar] [CrossRef]
- Lynch, M. Evolution of the mutation rate. Trends Genet. 2010, 26, 345–352. [Google Scholar] [CrossRef]
- Gopalapillai, R.; Kadono-Okuda, K.; Okuda, T. Molecular cloning and analysis of a novel teratocyte-specific carboxylesterase from the parasitic wasp, Dinocampus coccinellae. Insect Biochem. Mol. Biol. 2005, 35, 1171–1180. [Google Scholar] [CrossRef]
- Consoli, F.L.; Lewis, D.; Keeley, L.; Vinson, S.B. Characterization of a cDNA encoding a putative chitinase from teratocytes of the endoparasitoid Toxoneuron nigriceps. Entomol. Exp. Appl. 2007, 122, 271–278. [Google Scholar] [CrossRef]
- Falabella, P.; Riviello, L.; De Stradis, M.L.; Stigliano, C.; Varricchio, P.; Grimaldi, A.; de Eguileor, M.; Graziani, F.; Gigliotti, S.; Pennacchio, F. Aphidius ervi teratocytes release an extracellular enolase. Insect Biochem. Mol. 2009, 39, 801–813. [Google Scholar] [CrossRef]
- Vincent, B.; Kaeslin, M.; Roth, T.; Heller, M.; Poulain, J.; Cousserans, F.; Schaller, J.; Poirié, M.; Lanzrein, B.; Drezen, J.M.; et al. The venom composition of the parasitic wasp Chelonus inanitus resolved by combined expressed sequence tags analysis and proteomic approach. BMC Genomics 2010, 11, 693:1–693:15. [Google Scholar]
- Wang, R.; Deng, F.; Hou, D.; Zhao, Y.; Guo, L.; Wang, H.; Hu, Z. Proteomics of the Autographa californica nucleopolyhedrovirus budded virions. J. Virol. 2010, 84, 7233–7242. [Google Scholar] [CrossRef]
- Means, J.C.; Passarelli, A.L. Viral fibroblast growth factor, matrix metalloproteases, and caspases are associated with enhancing systemic infection by baculoviruses. Proc. Natl. Acad. Sci. USA 2010, 107, 9825–9830. [Google Scholar]
- Ohkawa, T.; Volkman, L.E.; Welch, M.D. Actin-based motility drives baculovirus transit to the nucleus and cell surface. J. Cell Biol. 2010, 190, 187–195. [Google Scholar] [CrossRef]
- Andersen, J.N.; Mortensen, O.H.; Peters, G.H.; Drake, P.G.; Iversen, L.F.; Olsen, O.H.; Jansen, P.G.; Andersen, H.S.; Tonks, N.K.; Moller, N.P. Structural and evolutionary relationships among protein tyrosine phosphatase domains. Mol. Cell Biol. 2001, 21, 7117–7136. [Google Scholar]
- Pruijssers, A.J.; Strand, M.R. PTP-H2 and PTP-H3 from Microplitis demolitor Bracovirus localize to focal adhesions and are antiphagocytic in insect immune cells. J. Virol. 2007, 81, 1209–1219. [Google Scholar] [CrossRef]
- Suderman, R.J.; Pruijssers, A.J.; Strand, M.R. Protein tyrosine phosphatase-H2 from a polydnavirus induces apoptosis of insect cells. J. Gen. Virol. 2008, 89, 1411–1420. [Google Scholar] [CrossRef]
- Chen, Y.P.; Higgins, J.A.; Gundersen-Rindal, D.E. Quantitation of a Glyptapanteles indiensis polydnavirus gene expressed in parasitized host, Lymantria dispar, by real-time quantitative RT-PCR. J. Virol. Methods 2003, 114, 125–133. [Google Scholar] [CrossRef]
- Chen, Y.P.; Taylor, P.B.; Shapiro, M.; Gundersen-Rindal, D.E. Quantitative expression analysis of a Glyptapanteles indiensis polydnavirus protein tyrosine phosphatase gene in its natural lepidopteran host, Lymantria dispar. Insect Mol. Biol. 2003, 12, 271–280. [Google Scholar] [CrossRef]
- Falabella, P.; Caccialupi, P.; Varricchio, P.; Malva, C.; Pennacchio, F. Protein tyrosine phosphatases of Toxoneuron nigriceps bracovirus as potential disrupters of host prothoracic gland function. Arch. Insect Biochem. 2006, 61, 157–169. [Google Scholar] [CrossRef]
- Whitfield, J. Phylogeny of Microgastroid Braconid Wasps, and What it Tells Us About Polydnavirus Evolution. In The Hymenoptera: Evolution, Biodiversity and Biological Control; Austin, A.D., Dowton, M., Eds.; CSIRO Publishing: Melbourne, Australia, 2000; pp. 97–105. [Google Scholar]
- Iyer, L.M.; Aravind, L.; Koonin, E.V. Common origin of four diverse families of large eukaryotic DNA viruses. J. Virol. 2001, 75, 11720–11734. [Google Scholar] [CrossRef]
- Drezen, J.M.; Bèzier, A.; Lesobre, J.; Huguet, E.; Cattolico, L.; Periquet, G.; Dupuy, C. The few virus-like genes of Cotesia congregata bracovirus. Arch. Insect Biochem. 2006, 61, 110–122. [Google Scholar] [CrossRef]
- Bigot, Y.; Rabouille, A.; Doury, G.; Sizaret, P.Y.; Delbost, F.; Hamelin, M.H.; Periquet, G. Biological and molecular features of the relationships between Diadromus pulchellus ascovirus, a parasitoid hymenopteran wasp (Diadromus pulchellus) and its lepidopteran host, Acrolepiopsis assectella. J. Gen. Virol. 1997, 78, 1149–1163. [Google Scholar]
- DPI&F Entomology. IPM: Microplitis demolitor and ascovirus: Important natural enemies of Helicoverpa. Available online: http://www.dpi.qld.gov.au (accessed on 6 January 2012).
- Bèzier, A.; Herbinière, J.; Serbielle, C.; Lesobre, J.; Wincker, P.; Huguet, E.; Drezen, J.M. Bracovirus gene products are highly divergent from insect proteins. Arch. Insect Biochem. 2008, 67, 172–187. [Google Scholar] [CrossRef]
- Serbielle, C.; Chowdhury, S.; Pichon, S.; Dupas, S.; Lesobre, J.; Purisima, E.O.; Drezen, J.M.; Huguet, E. Viral cystatin evolution and three-dimensional structure modelling: A case of directional selection acting on a viral protein involved in a host-parasitoid interaction. BMC Biol. 2008, 6, 38:1–38:16. [Google Scholar]
- Lu, Z.; Beck, M.H.; Strand, M.R. Egf1. 5 is a second phenoloxidase cascade inhibitor encoded by Microplitis demolitor bracovirus. Insect Biochem. Mol. 2010, 40, 497–505. [Google Scholar] [CrossRef]
- Friedman, R.; Hughes, A.L. Pattern of gene duplication in the Cotesia congregata Bracovirus. Infect. Genet. Evol. 2006, 6, 315–322. [Google Scholar] [CrossRef]
- Leister, D. Tandem and segmental gene duplication and recombination in the evolution of plant disease resistance gene. Trends Genet. 2004, 20, 116–122. [Google Scholar] [CrossRef]
- Dupuy, C.; Periquet, G.; Serbielle, C.; Bèzier, A.; Louis, F.; Drezen, J.M. Transfer of a chromosomal Maverick to endogenous bracovirus in a parasitoid wasp. Genetica 2011, 139, 489–496. [Google Scholar] [CrossRef]
- Esnault, C.; Maestre, J.; Heidmann, T. Human LINE retrotransposons generate processed pseudogenes. Nat. Genet. 2000, 24, 363–367. [Google Scholar] [CrossRef]
- Falabella, P.; Varricchio, P.; Gigliotti, S.; Tranfaglia, A.; Pennacchio, F.; Malva, C. Toxoneuron nigriceps polydnavirus encodes a putative aspartyl protease highly expressed in parasitized host larvae. Insect Mol. Biol. 2003, 12, 9–17. [Google Scholar] [CrossRef]
- Gundersen-Rindal, D. Integration of Polydnavirus DNA into Host Cellular Genomic DNA. In Parasitoid Viruses; Beckage, N.E., Drezen, J.M., Eds.; Academic Press: London, UK, 2012; pp. 99–114. [Google Scholar]
- Moran, N.A.; Degnan, P.H.; Santos, S.R.; Dunbar, H.E.; Ochman, H. The players in a mutualistic symbiosis: Insects, bacteria, viruses, and virulence genes. Proc. Natl. Acad. Sci. USA 2005, 102, 16919–16926. [Google Scholar]
- Oliver, K.M.; Degnan, P.H.; Hunter, M.S.; Moran, N.A. Bacteriophages encode factors required for protection in a symbiotic mutualism. Science 2009, 325, 992–994. [Google Scholar] [CrossRef]
- Oliver, K.M.; Moran, N.A.; Hunter, M.S. Variation in resistance to parasitism in aphids is due to symbionts not host genotype. Proc. Natl. Acad. Sci. USA 2005, 102, 12795–12800. [Google Scholar]
- Bordenstein, S.R.; Marshall, M.L.; Fry, A.J.; Kim, U.; Wernegreen, J.J. The tripartite associations between bacteriophage, Wolbachia, and arthropods. PLoS Pathog. 2006, 2, e43. [Google Scholar] [CrossRef]
- Kukan, B. Vertical transmission of nucleopolyhedrovirus in insects. J. Invertebr Pathol. 1999, 74, 103–111. [Google Scholar] [CrossRef]
- Burden, J.P.; Possee, R.D.; Sait, S.M.; King, L.A.; Hails, R.S. Phenotypic and genotypic characterisation of persistent baculovirus infections in populations of the cabbage moth (Mamestra brassicae) within the British Isles. Arch. Virol. 2006, 151, 635–649. [Google Scholar] [CrossRef]
- Burden, J.P.; Griffiths, C.M.; Cory, J.S.; Smith, P.; Sait, S.M. Vertical transmission of sublethal granulovirus infection in the Indian meal moth, Plodia interpunctella. Mol. Ecol. 2002, 11, 547–555. [Google Scholar]
- Villarreal, L.P. Virus-host symbiosis mediated by persistence. Symbiosis 2007, 44, 1–9. [Google Scholar]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar]
- Ng, T.F.; Willner, D.L.; Lim, Y.W.; Schmieder, R.; Chau, B.; Nilsson, C.; Anthony, S.; Ruan, Y.; Rohwer, F.; Breitbart, M. Broad surveys of DNA viral diversity obtained through viral metagenomics of mosquitoes. PLoS One 2011, 6, e20579. [Google Scholar]
- Rosario, K.; Marinov, M.; Stainton, D.; Kraberger, S.; Wiltshire, E.J.; Collings, D.A.; Walters, M.; Martin, D.P.; Breitbart, M.; Varsani, A. Dragonfly cyclovirus, a novel single-stranded DNA virus discovered in dragonflies (Odonata: Anisoptera). J. Gen. Virol. 2011, 92, 1302–1308. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Burke, G.R.; Strand, M.R. Polydnaviruses of Parasitic Wasps: Domestication of Viruses To Act as Gene Delivery Vectors. Insects 2012, 3, 91-119. https://doi.org/10.3390/insects3010091
Burke GR, Strand MR. Polydnaviruses of Parasitic Wasps: Domestication of Viruses To Act as Gene Delivery Vectors. Insects. 2012; 3(1):91-119. https://doi.org/10.3390/insects3010091
Chicago/Turabian StyleBurke, Gaelen R., and Michael R. Strand. 2012. "Polydnaviruses of Parasitic Wasps: Domestication of Viruses To Act as Gene Delivery Vectors" Insects 3, no. 1: 91-119. https://doi.org/10.3390/insects3010091