
Impacts of Non-Lethal High-Temperature Stress on the Development and Reproductive Organs of Bradysia odoriphaga

1
Department of Plant Protection, College of Agriculture, Yangtze University, Jingzhou 434025, China
2
Forewarning and Management of Agricultural and Forestry Pests, Hubei Engineering Technology Center, Yangtze University, Jingzhou 434025, China
3
Henan Engineering Research Center of Chinese Chives, Pindingshan Academy of Agricultural Sciences, Pindingshan 467000, China
*
Author to whom correspondence should be addressed.
Insects 2022, 13(1), 74; https://doi.org/10.3390/insects13010074
Submission received: 19 November 2021
/
Revised: 29 December 2021
/
Accepted: 7 January 2022
/
Published: 10 January 2022
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Bradysia odoriphaga is a soil-dwelling insect native to China, and its preferred host is Chinese chives. In this study, non-lethal high-temperature as an important limiting factor to affect the population and development of B. odoriphaga was assessed. Meanwhile, the physiological mechanism on non-lethal high-temperature to reduce the population was also identified. These would lay a key theoretical foundation for the future development of high-temperature products for environment friendly pest control strategies.
Abstract
Bradysia odoriphaga is an agricultural pest in China’s vegetable industry. In this study, pupae and adults were exposed to various non-lethal high-temperatures. The results demonstrated a decreased rate of eclosion once the pupae were exposed to temperatures exceeding 37 °C for 1 h. No effect on the lifespan of unmated female adults was observed after exposure to temperature stress, while unmated male adult lifespan decreased (>37 °C for 2 h). The size of the testis and ovaries for unmated male and female adults decreased, as did the fecundity and egg hatching rate for mated females. Compared with the control group (25 °C), the testis size of unmated male adults decreased after high-temperature stress followed by recovery at 25 °C for 1 h, though the size of the ovaries of female adults did not change. Additionally, the size of the testis and ovaries for unmated male and female adults decreased following high-temperature stress and 24 h of recovery at 25 °C. High temperatures affected males more than females; 37 °C is the critical temperature to control the population of B. odoriphaga. These results lay the foundation for the future development of environmentally friendly high-temperature prevention and pest-control strategies.
1. Introduction
Bradysia odoriphaga Yang et Zhang (Diptera: Sciaridae) is commonly known as the chive maggot and consumes roots [1]. It has a wide range of hosts and can damage over thirty kinds of vegetables and fruits from seven families, as well as edible fungi [2,3]. These hosts include, but are not limited to, Welsh onion, garlic, lettuce, celery, Chinese cabbage, and Chinese chives (Allium tuberosum Rottler ex Spreng). If control measures are not used against B. odoriphaga, yields of A. tuberosum can be reduced by 50%. In severe cases, the seedlings can be destroyed, leading to near-total crop failure [4].
The methods currently used to prevent and control B. odoriphaga include releasing entomopathogenic nematodes [5,6], hanging black sticky boards [7], and adjusting cultivation patterns [8]. However, these methods are limited by high costs and poor efficacy. Vegetable farmers use root irrigation of overdose chemical pesticides [9], which has conferred resistance on B. odoriphaga and resulted in environmental pollution and food safety concerns [10]. Therefore, research on green control technology for B. odoriphaga would be critical for ensuring healthy and sustainable agricultural productivity.
While many environmental factors influence insect growth, reproduction, and population structure [11,12], temperature is one of the most important limiting factors [13]. High temperatures can impact insect survival, growth and development, lifespan, reproductive rate, sex ratio, gene expression, and heat shock protein synthesis [14]. For instance, extreme high-temperatures hinder the feeding of Leucoptera coffeella Guérin-Méneville (Lepidoptera: Lyonetiidae) [15], affect the egg laying of Spodoptera exigua Hübner (Lepidoptera: Noctuidae) [16]. Bactrocera cucurbitae Coquillet (Diptera: Tephritidae) adults will die when the temperatures exceed 51 °C [17]. In short, if the temperatures are higher than the optimum temperature range for development, the adaptability of insects will instantly fall with the temperature increase [18]. Recently, Shi et al. [19] also identified a negative relationship between the population of B. odoriphaga and non-lethal high-temperatures in nature. A similar study demonstrated that non-lethal high-temperature stress on B. odoriphaga larvae negatively impacts subsequent growth and development, which can decrease their population [20]. Despite this understanding of temperature effects on larvae, it is unclear how high temperatures impact other development stages of B. odoriphaga and the ecological mechanism by which high temperatures decrease the population growth. If the above assertions were made certain, we could improve the environment temperature to the suitable non-lethal temperature to limit B. odoriphaga development, or control their population under the level of economic damage threshold, which would decrease the damage of Chinese chives and protect the ecological balance of the species. Therefore, we designed our study to expose the pupae and unmated female and male adults of B. odoriphaga to different degrees of non-lethal high-temperature stress. In particular, we assessed the impact of heat stress on B. odoriphaga reproduction by qualitatively examining the reproductive organs of adults. Additionally, we observed the effects of non-lethal heat stress on pupae and adult growth and development, and on reproductive success (measured as the female reproductive capacity and egg hatching rate). This provides a key theoretical foundation for the development of high-temperature products for environmentally friendly pest control.
2. Materials and Methods
2.1. Bradysia odoriphaga Rearing and Maintenance
The B. odoriphaga population was supplied by the Institute of Vegetables and Flowers, at the Chinese Academy of Agricultural Sciences, Beijing, China. They were cultured under a constant temperature (25 ± 1 °C) in an incubator (MLR-352H-PC) (70 ± 5% RH) at a 14:10 (L:D) h photoperiod in a laboratory located at the Agricultural College of Yangtze University. Three generations of this population were raised with A. tuberosum rhizomes for subsequent use.
2.2. Impacts of High-Temperature Stress on Bradysia odoriphaga Pupae
According to our research experience, female and male pupae can be distinguished by their size: females are significantly larger than males at the same age (unpublished). Male and female pupae were separated by sex within 8 h of pupation into two clearly labeled Petri dishes. The dish bottom was covered with a 3 mm thick layer of 2% solidified agar (A109143; Aladdin, Shanghai, China), while filter paper was placed on the surface of the agar to maintain the moisture content.
2.2.1. High-Temperature Impacts Growth and Development of Bradysia odoriphaga Pupae
A single pupa was placed on each Petri dish (Φ = 3 cm); 30 male and 30 female pupae were placed into the control or experimental group at a time. The experimental treatments exposed each group to 34, 37, and 40 °C temperatures for 1, 2, or 4 h. The control was exposed to 25 °C, with 30 females and 30 males for each treatment. Following the treatment, the Petri dishes were placed in an incubator at a constant temperature of 25 °C. Each treatment, as well as the control, was performed five times. The emergence of pupae was observed once a day, while the pupae were considered dead if eclosion did not begin within 10 days. Once eclosion occurred, the pupal sex was observed, while the pupae were maintained in their original Petri dishes. Adult survival was observed each day until all had died.
2.2.2. Impacts of High-Temperature Stress on Egg Laying and Egg Hatching in Bradysia odoriphaga Pupae
Forty male and forty female pupae were placed into the control or experimental group at a time; each was placed in an individual Petri dish (Φ = 6 cm). The experimental treatments exposed each group to 34, 37, and 40 °C temperatures for 1, 2, or 4 h. Following the treatment, the Petri dishes were placed in an incubator at a constant temperature of 25 °C. Once emergence occurred, female and male adults were placed, one by one, into a new Petri dish within 8 h. Every six pairs were considered a replicate that were placed in an incubator at 25 °C for typical feeding. Each treatment, as well as the control, was performed five times. Adult lifespan, the number of eggs each female laid, and rate of egg hatching were all recorded.
2.2.3. Impacts of High-Temperature Stress on Unmated Adult Reproductive Organs Size in Bradysia odoriphaga Pupae
Forty female and forty male pupae were individually placed into two Petri dishes (Φ = 6 cm) as a treatment or the control. Treatments were as follows: 34 °C and 37 °C for either 2 h or 4 h. The control was exposed to a constant temperature of 25 °C. After treatment, the Petri dishes were put into a constant temperature incubator at 25 °C. After pupae eclosion, the male and female adults (within 8 h of emergence) per treatment or the control were dissected, and the length of testis and ovaries was measured. Six pupae of the same sex were sampled as one replicate. Each treatment or control was repeated five times.
2.3. Impacts of High-Temperature Stress on the Morphology and Structure of Testis and Ovaries in Unmated Bradysia odoriphaga Adults
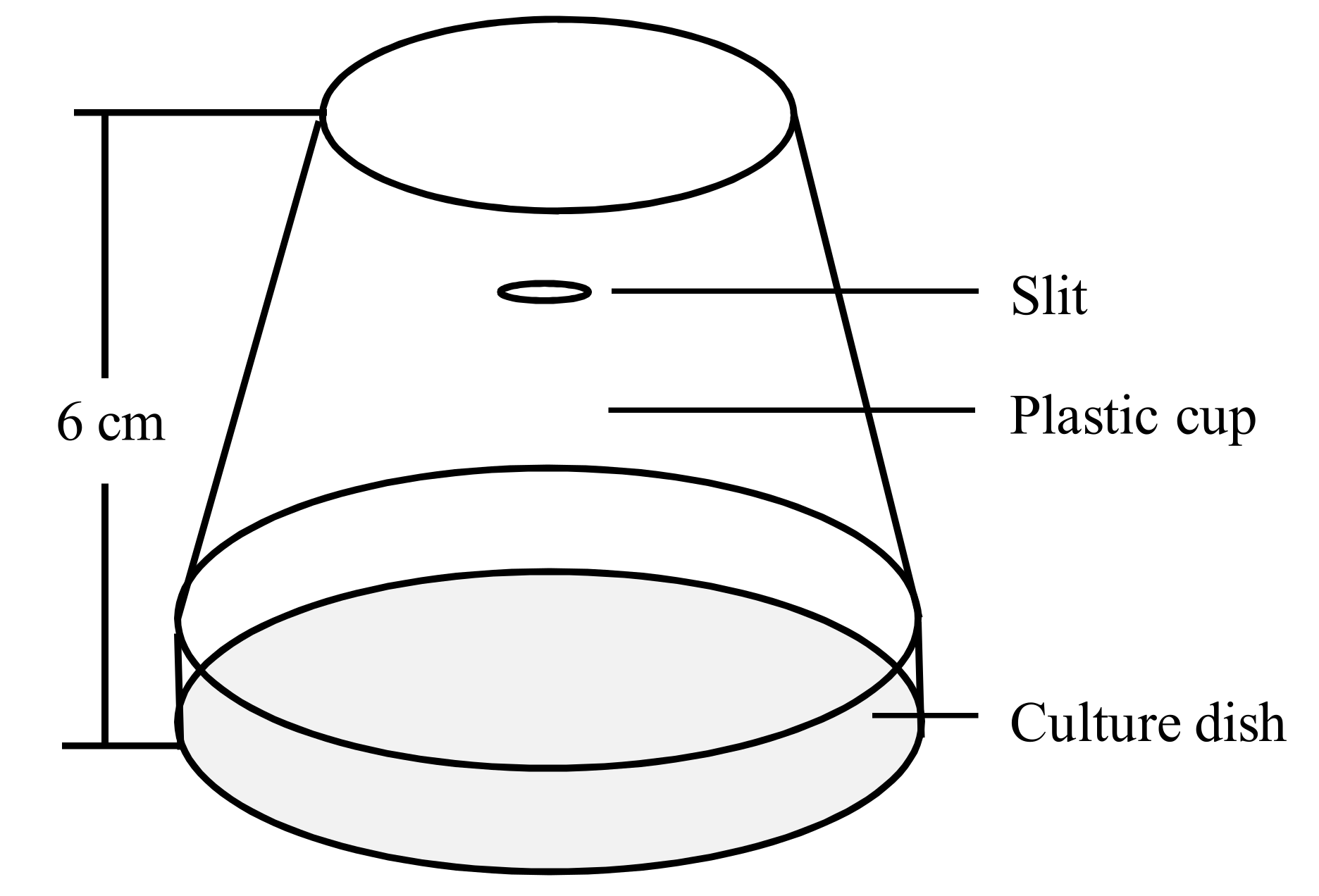
Forty female and forty male adults (within 8 h of emergence) were individually placed into two insect cages (Figure 1) as a treatment or the control. The treatments were as follows: exposure to 34 °C and 37 °C for either 2 h or 4 h. The control was exposed to a constant temperature of 25 °C. After treatment, the insect cages were placed into a constant temperature incubator at 25 °C to recover for 1 h or 24 h, individually. Afterward, the male and female adults were dissected and photographed, while the size of testis and ovaries was measured. Six adults of the same sex were sampled as one replicate. Each treatment or the control was repeated five times.
2.4. Data Analysis
Data entry was performed using Excel version 2007, while one-way analyses of variance (ANOVA) were performed using SPSS version 17.0 (SPSS Inc., Chicago, IL, USA). Data were presented as mean ± SD. Except lifetime in female and male was compared by Student’s t-test, other treatments were assessed by Tukey’s test. The results were considered significant when p < 0.05.
3. Results
3.1. Impacts of High-Temperature Stress on Bradysia odoriphaga Pupae
3.1.1. Impacts of High-Temperature Stress on Bradysia odoriphaga Pupae Emergence Rate
The rate of emergence of the female pupae decreased as the duration and temperature of exposure increased (F8,36 = 1919.523, p < 0.01). The rate of emergence of female pupae when exposed to 34 °C for 1, 2, and 4 h did not significantly differ from the rate observed in the 25 °C control group. No significant differences were observed in treatments at 37 °C for 1, 2, and 4 h, and 40 °C for 1 h. The lowest rate of emergence, 9.3%, was observed in female pupae exposed to 40 °C for 2 h (Figure 2A).
The rate of emergence of the male pupae decreased as the duration and temperature of exposure increased (F8,36 = 3679.482, p < 0.01). The rate of emergence of male pupae exposed to 34 °C for 4 h significantly differed from the 25 °C control group. Once temperatures exceeded 37 °C, the rate of male eclosion decreased significantly below that of the 25 °C control group. No significant difference was observed among treatments at 37 °C for 1, 2, and 4 h, and 40 °C for 1 h. The lowest rate of emergence, 6.0%, was observed in male pupae exposed to 40 °C for 2 h (Figure 2B).
3.1.2. Impacts of High-Temperature Stress on Adults Longevity in Bradysia odoriphaga Pupae
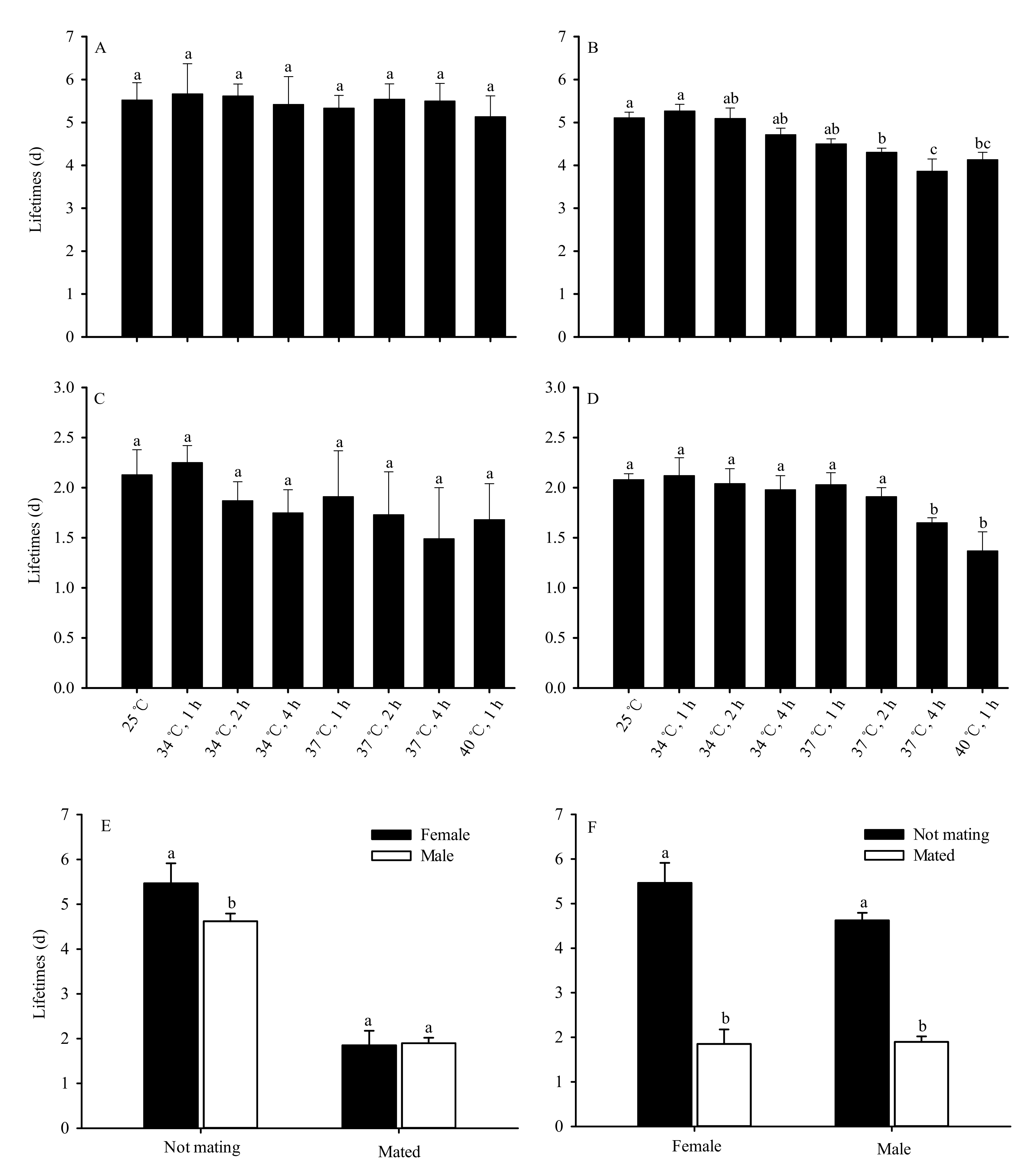
No significant differences were observed in the lifespan of unmated female adults after the pupae were subjected to various levels of short-term high-temperature stress (F7,32 = 0.451, p = 0.857) (Figure 3A). However, the lifespan of unmated male adults significantly decreased as the duration and temperature of exposure increased (F7,32 = 94.865, p < 0.01). Compared with the 25 °C control group, no significant differences were observed in the lifespan of male adults following exposure of the pupae to 34 °C for 1, 2 and 4 h, and 37 °C for 1 h. However, the lifespan of the 37 °C groups treated for 2 h and 4 h and the group treated at 40 °C for 1 h were significantly shorter than the 25 °C control group. There was no difference in male lifespan between the 37 °C group (2 h of exposure) and the 40 °C group (1 h of exposure). The shortest male lifespan, 3.86 days, occurred in the 37 °C group (4 h of exposure) (Figure 3B).
No significant differences were also observed in the lifespan of mated female adults after the pupae were subjected to various levels of short-term high-temperature stress (F7,32 = 1.174, p = 0.345) (Figure 3C). The lifespan of mated male adults also significantly decreased as duration and temperature of exposure increased (F7,32 43.554, p < 0.01) (Figure 3D). However, the average lifespan of unmated female adults (5.47 d) significantly exceeded that of unmated male adults (4.62 d) (t = 11.380, df = 78, p < 0.01). The average lifespan of female adults was lower than that of male adults (1.85 days and 1.89 days respectively) after mating, though the difference was not significant (t = 0.272, df = 63.322, p = 0.787) (Figure 3E). The average lifespan of female (t = 40.474, df = 78, p < 0.01) and male (t = 39.138, df = 78, p < 0.01) adults significantly decreased following mating (Figure 3F).
3.1.3. Impacts of High-Temperature Stress on Reproductive Capacity and Egg Hatchability in Bradysia odoriphaga
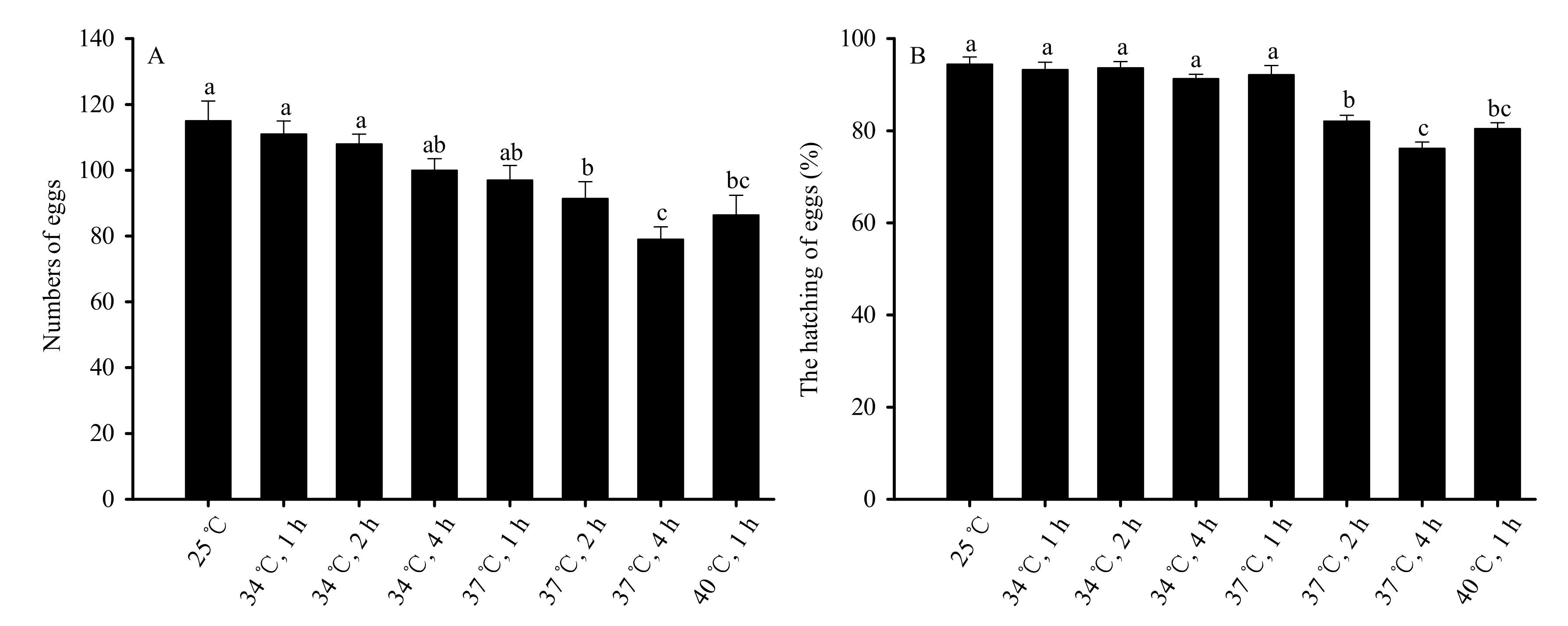
Female reproductive capacity was measured by the number of eggs that each female laid, and decreased significantly when the pupal duration and temperature of stress exposure increased (F7,32 = 44.586, p < 0.01). No significant differences were observed in the ability of mated female adults to reproduce at 34 °C of stress for 1, 2, and 4 h, or under 37 °C for 1 h when compared to the control group (25 °C). The reproductive capacity decreased significantly compared to the control group under 37 °C of treatment for 2 h and 4 h and 40 °C of treatment for 1 h. No significant difference was observed in groups exposed to 40 °C for 1 h and groups exposed to 37 °C for 2 h. The capacity of females to reproduce was the lowest following 4 h of treatment at 37 °C (Figure 4A).
High-temperature stress also significantly affected the rate of hatching (F7,32 = 114.882, p < 0.01). No significant difference was observed in the rate of hatching of eggs laid by mated females in the groups stressed at 34 °C for 1, 2, or 4 h, or 37 °C for 1 h compared to the control. Significant decreases were observed in groups exposed to 37 °C for 2 h and 4 h and 40 °C for 1 h compared to the control group. However, the difference between the 40 °C group (1 h of exposure) and the 37 °C group (2 h of exposure) was not significantly different. The rates of egg hatching were lowest following 4 h of stress at 37 °C (Figure 4B).
3.1.4. Impacts of High-Temperature Stress on the Reproductive Organs of Unmated Adults in Bradysia odoriphaga Pupae
Treatment of pupae with short-term high-temperature stress significantly impacted the size of the reproductive organs of the adults (Figure 5). As temperature and duration increased, ovaries length decreased (F4,20 = 40.783, p < 0.01). The length of ovaries of the 25 °C control group was the longest and was significantly longer than that of 34 °C for 4 h, 37 °C for 2 h and 37 °C for 4 h treatment groups. The ovaries of the treatment groups at 37 °C for 2 h and 37 °C for 4 h were the shortest (Figure 5A). Similarly, as the temperature and duration increased, the testis length decreased (F4,20 = 115.544, p < 0.01). The testis length was longest in the 25 °C control group and was significantly longer than that of all other treatment groups. The shortest testis was found in the group treated at 37 °C for 4 h (Figure 5B).
3.2. Impacts of High-Temperature Stress of Bradysia odoriphaga Unmated Adults on the Size of Reproductive Organs
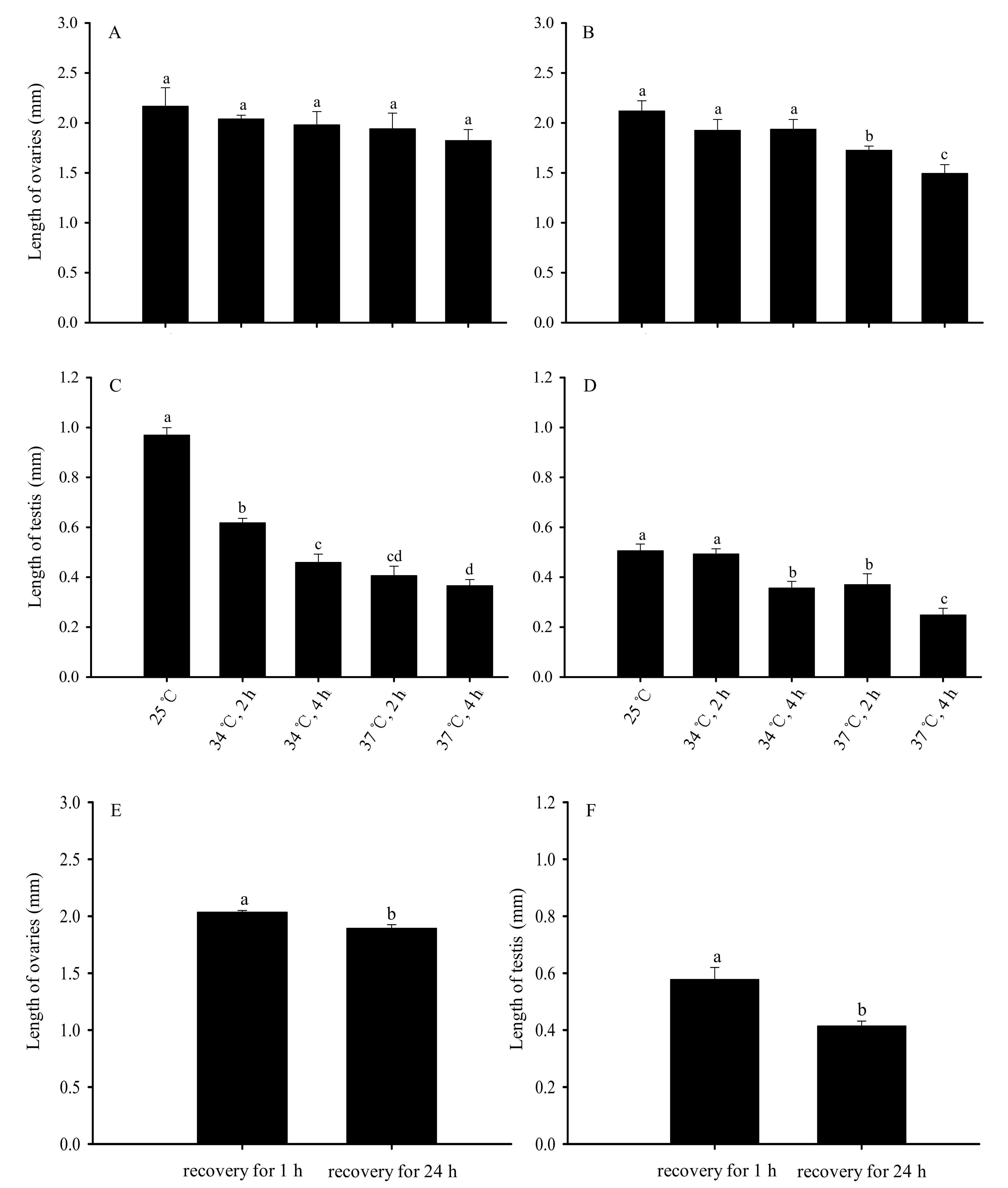
When adults were treated with short-term high-temperature stress, the size of the reproductive organs of subsequent unmated adults significantly changed (Figure 6). Compared with the ovaries of unmated female adults normally reared at 25 °C, there was no significant difference in the length of ovaries on unmated female adults after short-term high-temperature stress and 1 h recovery at 25 °C (F4,20 = 2.684, p = 0.062) (Figure 6A). Differing from the ovaries of unmated female adults normally reared at 25 °C, the length of the ovaries in the group that recovered at 25 °C for 24 h after short-term high-temperature treatment decreased as the temperature and duration of the stress increased (F4,20 = 43.036, p < 0.01). The shortest ovaries were surveyed in the group exposed to 37 °C for 4 h (Figure 6B).
Compared with the testis of unmated male adults normally reared at 25 °C, there was a significant difference in the length of testis on unmated male adults after short-term high-temperature stress and 1 h recovery at 25 °C (F4,20 = 293.858, p < 0.01) (Figure 6C). Additionally, under the same treatment conditions, the length of the testis of male adults after 24 h recovery was also significant difference (F4,20 = 222.723, p < 0.01). The shortest testis were surveyed in the group exposed to 37 °C for 4 h (Figure 6D).
Additionally, under the same treatment conditions, the length of the ovaries of female adults after 24 h recovery was significantly smaller than those of female adults after 1 h of recovery (t = 3.943, df = 35.302, p < 0.01) (Figure 6E). The length of the testis of male adults after 24 h recovery was also significantly smaller than those of male adults after 1 h of recovery (t = 3.560, df = 31.926, p < 0.01) (Figure 6F).
4. Discussion
The temperature of the surrounding environment is the primary factor limiting the activity, development, distribution, and reproduction of insects. High temperatures can disrupt the growth, development, and reproduction of insects [21,22]. The sensitivity of various insect species to high temperatures varies, and different insects have different physiological responses to this stress. For instance, closely related species can have significantly different diapause (Heliothis virescent Fabricius (Lepidoptera: Noctuidae), 32 °C [16] and Helicoverpa armigera Hübner (Lepidoptera: Noctuidae), 37 °C [23]), while pupae and adults of Plutella xylostella Linnaeus (Lepidoptera: Plutellidae) larvae exposed to 35 °C decrease in size and cannot normally reproduce [24]. Adults of Tribolium castaneum Herbst (Coleoptera: Tenebrionidae) cannot reproduce after being exposed to 50 °C for 39 min [25]. Insects are poikilotherms, meaning their body temperatures change rapidly as the temperature of their environment is altered, which can result in death [26]. The growth, development, and reproductive capacity can still be disrupted even if the insect is not killed. High-temperature exposure increases the preoviposition period in Liriomyza sativae Blanchard (Diptera: Agromyzidae), decreases the lifespan of Laodelphax striatellus Fallen (Hemiptera: Delphacidae) [27], decreases the ability of Myzus persicae Sulzer (Hemiptera: Aphididae) to reproduce [28], alters the ratio of males and females in the offspring of Frankliniella occidentalis Pergande (Thysanoptera: Thripidae), and alters reproduction in Harmonia axyridis Pallas (Coleoptera: Coccinellidae) [29]. Our results demonstrate that exposing B. odoriphaga pupae to heat stress (at or exceeding 37 °C) significantly lowers the rate of eclosion as the temperature and duration of exposure increase. This indicates that 37 °C is a threshold temperature for the eclosion of pupae. Shi et al. [26] determined that B. odoriphaga pupae were more resistant to high-temperature stresses than the other life-history stages. As high temperatures decrease the eclosion rate, other insect life stages could be affected. This study confirms that the 37 °C threshold does affect the development of pupae and the ability of adults to reproduce. This aligns with previous findings that B. odoriphaga populations decrease in the summertime because temperatures in certain regions of China can exceed 37 °C [19].
We determined that if pupae successfully develop into adults under heat stress, there is little effect on the lifespan of unmated female adults. However, the male lifespan decreases significantly. Liang et al. [30] demonstrated that B. odoriphaga males can mate several times throughout their lives. If the male lifespan decreases significantly, it will decrease the times of mating, thus reducing the population of the insect. Therefore, our study further indicated that high-temperature stress can affect the population of B. odoriphaga by decreasing the lifespan of males and lowering their reproductive capacity.
The lifespan of an insect is associated with its intake of energy: insects that live a long time require a lot of energy [31]. However, B. odoriphaga adults do not eat and have a fairly short lifespan. Male adults are significantly smaller than females, though the ability of females to fly is lower than that of males [32]. The association between energy use, energy consumption, and development would suggest that females eat more and live longer than males. This aligns with our findings. The lifespan of unmated male adults is much shorter than that of unmated female adults, though we found no significant difference in the lifespan of female and male adults following mating. This indicates that females spend more energy on reproduction and oviposition than on lifespan. The lifespan of treated females significantly decreased after they mated. Mating requires significant amounts of energy; B. odoriphaga males can mate several times in their lives [30]. The stress of high temperatures significantly affected male pupae and the lifespan of unmated adult males. This suggests that the restoration of reproductive and developmental functioning following stress significantly affects male lifespan. Male lifespan in B. odoriphaga is already shorter than female lifespan, and heat stress further widens this gap.
High-temperature stress impacts development and propagation in insects [33,34]. Our research demonstrated that female reproductive capacity and the egg hatchability of mated adult females exposed to heat stress as pupae decreased significantly as the temperature and duration increased. The reproductive capacity and rate of egg hatching were the lowest after 4 h of exposure to 37 °C. Additionally, female ovaries became shorter as the temperature and duration of heat stress increased, while the shortest ovary was found in the treatment group exposed to 37 °C for 4 h. This relationship can explain why the adult female reproductive capacity and egg hatchability both significantly decrease after the pupae are exposed to high-temperature stress. After male pupae were exposed to heat stress, the testis size in adult B. odoriphaga became smaller with the increase in stress temperature and duration, which may also affect the survival quality of male sperm and reduce egg hatchability [35]. The influence of high-temperature stress on the sperm survival rate of B. odoriphaga requires further study.
The reproductive organs of male and female B. odoriphaga gradually began to develop and mature during the pupal stage [28]. We found that applying non-lethal high-temperature stress to the pupae of B. odoriphaga impacted adult reproductive anatomy more than applying heat stress to adults. Our research also demonstrated that applying high-temperature stress to unmated female adults and providing recovery at the control temperature for 1 h had no significant effect on ovary length, while, when the recovery time increased to 24 h, the size of the ovary decreased significantly. This indicates that short-term high-temperature stress has a continuous effect on the ovaries. Research has demonstrated that female adults (B. odoriphaga) will discharge some of their eggs as they recover bodily functions following exposure to high-temperature stress [36]. It indicated that eggs in the ovaries could be discharged following exposure to high-temperature stress or with recovery time increased, thus decreasing the size of the ovaries. Meanwhile, unfertilized eggs discharged would indirectly decrease the population number of B. odoriphaga.
High temperatures adversely affect both male and female propagation [29]. Our research demonstrates that high-temperature stress on unmated B. odoriphaga male adults was more significant than that on unmated female adults. Male testis shrank significantly as the temperature and duration increased, and B. odoriphaga testis were the smallest after 4 h of exposure to 37 °C. Moreover, under the same treatment temperature and duration, the testis of male adults that were allowed to recover for 24 h were significantly smaller than the testis of male adults that were allowed to recover for 1 h. Previous research has demonstrated that high temperatures can cause Grapholitha molesta Busck (Lepidoptera: Tortricidae) to expel semen from the testis, thereby affecting the size of the testis [37]. Consequently, we suspect that high temperatures impact the formation of male testis, including the potential loss of sperm count or motility, thus indirectly decreasing the population number of B. odoriphaga. Certainly, the specific effects of high-temperature on the formation and activity of B. odoriphaga testis requires further study.
5. Conclusions
We found that B. odoriphaga pupae exposed to heat stress exhibit greater constraints on growth and development, female reproductive capacity, hatchability, and testis and ovary size than adults. We also found that high-temperature stress impacts males more than females. Moreover, our results demonstrate that 37 °C is a crucial temperature for controlling populations of B. odoriphaga. Altogether, these results demonstrate that heat stress can be used to negatively affect B. odoriphaga, providing an environmentally friendly control strategy for an economically devastating pest.
Author Contributions
Conceptualization, J.H. and C.S.; methodology, C.S.; software, J.H.; validation, J.H., R.G.M. and S.Z.; formal analysis, P.M.; investigation, J.H. and S.Z.; resources, J.H.; data curation, J.H.; writing—original draft preparation, J.H.; writing—review and editing, R.G.M. and C.S.; visualization, J.H.; supervision, J.H.; project administration, J.H.; funding acquisition, C.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the National Natural Science Foundation of China (32172400), the Project of Education Department in Hubei Province (B2020038).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article.
Acknowledgments
The authors would like to thank Guanglin Xie (Institute of Insects Research, College of Agriculture, Yangtze University) for providing Olypums microscope used in this study.
Conflicts of Interest
The authors declare that they have no conflict of interest in this study.
References
- Yang, J.K.; Zhang, X.M. Notes on the fragrant onion gnats with descriptions of two new species of Bradysia (Diptera: Sciaridae). Acta Agric. Univ. Pekin. 1985, 11, 153–156. [Google Scholar]
- Yang, Y.T.; Li, W.X.; Xie, W.; Wu, Q.J.; Xu, B.Y.; Wang, S.L.; Zhang, Y.J. Development of Bradysia odoriphaga (Diptera: Sciaridae) as affected by humidity: An age-stage, two-sex, life-table study. Appl. Entomol. Zool. 2015, 50, 3–10. [Google Scholar] [CrossRef]
- Tao, Y.L.; Guo, Y.N.; Wang, J.; Li, L.L.; Yu, Y.; Chu, D. Detection and identification of Wolbachia in Bradysia odoriphaga (Diptera: Sciaridae) populations from Shangdong Province, China. Acta Entomol. Sin. 2015, 58, 454–459. [Google Scholar]
- Shan, T.S.; Chen, C.Y.; Ding, Q.; Chen, X.W.; Zhang, H.H.; Chen, A.Q.; Shi, X.Y.; Gao, X.W. Molecular characterization and expression profiles of nicotinic acetylcholine receptors in Bradysia odoriphaga. Pestic. Biochem. Physiol. 2020, 165, 104563. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.H.; Li, A.H.; Han, R.C.; Cao, L.; Liu, X.L. Factors affecting the control of Bradysia odoriphaga with entomopathogenic nematode Heterorhabditis indica LN2. Nat. Enemies Insects 2004, 26, 150–155. [Google Scholar]
- Ma, J.; Chen, S.L.; Moens, M.; Han, R.C.; Clercq, P.D. Efficacy of entomopathogenic nematodes (Rhabditida: Steinernematidae and Heterorhabditidae) against the chive gnat, Bradysia odoriphaga. J. Pest Sci. 2013, 86, 551–561. [Google Scholar] [CrossRef]
- Wang, H.T.; Song, C.F.; Wang, Y.Z. The tendency for Bradysia odoriphaga among different color and the luring effect on yellow plate. J. Agr. Sci. Jiangsu 2015, 43, 133–134. [Google Scholar]
- Dang, Z.H.; Dong, J.H.; Gao, Z.L. Biology and injury of Bradysia odoriphaga on leek in different types of cultivation. J. Agric. Univ. Hebei 2002, 24, 65–68. [Google Scholar]
- Ullah, F.; Gul, H.; Desneux, N.; Sai, F.; Gao, X.W.; Song, D.L. Fitness costs in chlorfenapyr-resistant populations of the chive maggot, Bradysia odoriphage. Ecotoxicology 2020, 29, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Liu, F.; Mu, W.; Li, Q.H.; Wang, H.; Chen, C.Y. Life table study of the effects of sublethal concentrations of thiamethoxam on Bradysia odoriphage Yang and Zhang. Pestic. Biochem Phys. 2014, 111, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Boggs, C.L. The fingerprints of global climate change on insect populations. Curr. Opin. Insect Sci. 2016, 17, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Hamby, K.A.; Bellamy, D.E.; Chiu, J.C.; Lee, J.C.; Walton, V.M.; Wiman, N.G.; Biondi, A. Biotic and abiotic factors impacting development, behavior, phenology, and reproductive biology of Drosophila suzukii. J. Pest Sci. 2016, 89, 605–619. [Google Scholar] [CrossRef]
- Cui, X.H.; Wan, F.H.; Xie, M.; Liu, T.X. Effects of heat shock on survival and reproduction of two whitefly species, Trialeurodes vaporariorum and Bemisia tabaci biotype B. J. Insect Sci. 2018, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kobori, Y.; Hanboonsong, Y. Effect of temperature on the development and reproduction of the sugarcane white leaf insect vector, Matsumuratettix hiroglyphicus (Matsumura) (Hemiptera: Cicadellidae). J. Asia-Pac. Entomol. 2017, 20, 281–284. [Google Scholar] [CrossRef]
- Lomeli-Flores, J.R.; Barrera, J.F.; Bernal, J.S. Impacts of weather, shade cover and elevation on coffee leafminer Leucoptera coffeella (Lepidoptera: Lyonetiidae) population dynamics and natural enemies. Crop Prot. 2010, 29, 1039–1048. [Google Scholar] [CrossRef]
- Wang, H.S.; Xu, H.F.; Cui, F. Effect of high temperature on fecundity and ovary development of beet armyworm Spodoptera exigua (Hǜbner). Southwest China J. Agr. Sci. 2006, 19, 916–919. [Google Scholar]
- Zeng, B.; Zhu, W.J.; Fu, Y.G.; Zhou, S.H. Influence of high-temperature exposure on the mating, oviposition and thermotaxis of Bactrocera cucurbitae (Coquillet) (Diptera: Tephritidae). PLoS ONE 2018, 13, e0204065. [Google Scholar] [CrossRef] [PubMed]
- Fields, P.G. The control of stored-product insects and mites with extreme temperatures. J. Stored Prod. Res. 1992, 28, 89–118. [Google Scholar] [CrossRef]
- Shi, C.H.; Hu, J.R.; Zhang, Y.J. The effects of temperature and humidity on a field population of Bradysia odoriphaga (Diptera: Sciaridae). J. Econ. Entomol. 2020, 113, 1927–1932. [Google Scholar] [CrossRef]
- Shi, C.H.; Zhang, S.; Hu, J.R.; Zhang, Y.J. Effects of non-lethal high-temperature stress on Bradysia odoriphaga (Diptera: Sciaridae) larval development and offspring. Insects 2020, 11, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, G.D.; Xue, M.; Luo, Y.; Ji, G.X.; Liu, F.; Zhao, H.; Sun, X. Effect of short-term heat shock and physiological responses to heat stress in two Bradysia adults, Bradysia odoriphaga and Bradysia difformis. Sci. Rep. 2017, 7, 13381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, G.D.; Wilson, L.T.; Henneberry, T.J. Heliothis virescens (Lepidoptera: Noctuidae): Initiation of Summer diapause. J. Econ. Entomol. 1985, 78, 320–324. [Google Scholar] [CrossRef] [Green Version]
- Nibouche, S. High temperature induced diapause in the cotton bollworm shape Helicoverpa armigera. Entomol. Exp. Appl. 1998, 87, 271–274. [Google Scholar] [CrossRef]
- Ma, C.S.; Hau, B.; Poehling, H.M. The effect of heat stress on the survival of the rose grain aphid, Metopolophium dirhodum (Hemiptera: Aphididae). Eur. J. Entomol. 2004, 101, 327–331. [Google Scholar] [CrossRef]
- Mahroof, R.; Subramanyam, B.; Flinn, P. Reproductive performance of Tribolium castaneum (Coleoptera: Tenebrionidae) exposed to the minimum heat treatment temperature as pupae and adults. J. Econ. Entomol. 2005, 98, 626–633. [Google Scholar] [CrossRef]
- Shi, C.H.; Hu, J.R.; Wei, Q.W.; Yang, Y.T.; Cheng, J.X.; Han, H.L.; Wu, Q.J.; Wang, S.L.; Xu, B.Y.; Su, Q.; et al. Control of Bradysia odoriphaga (Diptera: Sciaridae) by soil solarization. Crop Prot. 2018, 114, 76–82. [Google Scholar] [CrossRef]
- Zhang, A.M.; Liu, X.D.; Zhai, B.P.; Gu, X.Y. Influences of temperature on biological characteristics of the small brown planthopper, Laodelphax striatellus (Fallén) (Hemiptera: Delphacidae). Acta Entomol. Sin. 2008, 51, 640–645. [Google Scholar]
- Chiu, M.C.; Kuo, J.J.; Kuo, M.H. Life stage-dependent effects of experimental heat waves on an insect herbivore. Ecol. Entomol. 2015, 40, 175–181. [Google Scholar] [CrossRef]
- Zhang, S.; Cao, Z.; Wang, Q.L.; Zhang, F.; Liu, T.X. Exposing eggs to high temperatures affects the development, survival and reproduction of Harmonia axyridis. J. Therm. Biol. 2014, 39, 40–44. [Google Scholar] [CrossRef]
- Liang, L.; Zhang, S.; Han, H.L.; Xu, B.Y.; Zhang, Y.J. Effects of male age on its mating ability and female reproduction in Bradysia odoriphaga. J. Environ. Entomol. 2019, 41, 657–663. [Google Scholar]
- Li, W.X.; Yang, Y.T.; Xie, W.; Wu, Q.J.; Xu, B.Y.; Wang, S.L.; Zhang, Y.J. Effects of temperature on the age-stage, two-sex life table of Bradysia odoriphaga (Diptera: Sciaridae). J. Econ. Entomol. 2015, 108, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.R.; Xie, C.; Shi, C.H.; Wang, S.L.; Wu, Q.J.; Li, C.R.; Zhang, Y.J. Effect of sex and air temperature on the flight capacity of Bradysia odoriphaga (Diptera: Sciaridae). J. Econ. Entomol. 2019, 112, 2161–2166. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.S.; Ma, G.; Pincebourde, S. Survive a warming climate: Insect responses to extreme high temperatures. Annu. Rev. Entomol. 2021, 66, 163–184. [Google Scholar] [CrossRef]
- Durak, R.; Dampc, J.; Dampc, J. Role of temperature on the interaction between Japanese quince Chaenomeles japonica and herbivorous insect Aphis pomi (Hemiptera: Aphidoidea). Environ. Exp. Bot. 2020, 176, 104100. [Google Scholar] [CrossRef]
- Pelletier, Y. Determination of the lethal high temperature for the Colorado potato beetle (Coleoptera: Chrysomelidae). Can. Agric. Eng. 1998, 40, 185–189. [Google Scholar]
- Uddin, M.D.K.; Yin, X.W.; Zhang, L. Courtship and mating behavior of the Chinese Chive fly, Bradysia odoriphaga (Diptera: Sciaridae) and evidence of female sex pheromone. Pak. J. Zool. 2016, 48, 1543–1548. [Google Scholar]
- Liang, L.N.; Zhang, W.; Ma, G.; Hoffmann, A.A.; Ma, C.S. A single hot event stimulates adult performance but reduces in the oriental fruit moth, Grapholitha molesta. PLoS ONE 2014, 9, e116339. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Insect cage for adults.

Figure 2.
High-temperature stress affects the emergence rate in female (A) and male (B) Bradysia odoriphaga pupae. Data are mean ± SD of five independent replicates (n = 30 for each replicate, N = 150 treatments). Different bar labels indicate significant differences according to Tukey’s test (p < 0.05). Pupae for control or each treatment were placed in a single, clearly marked Petri dish. Pupal eclosion was observed daily, and pupae were considered dead if eclosion did not begin within 10 days.
Figure 2.
High-temperature stress affects the emergence rate in female (A) and male (B) Bradysia odoriphaga pupae. Data are mean ± SD of five independent replicates (n = 30 for each replicate, N = 150 treatments). Different bar labels indicate significant differences according to Tukey’s test (p < 0.05). Pupae for control or each treatment were placed in a single, clearly marked Petri dish. Pupal eclosion was observed daily, and pupae were considered dead if eclosion did not begin within 10 days.

Figure 3.
High-temperature stress impacts of adult lifespan in Bradysia odoriphaga pupae ((d) = days). (A) indicates unmated female adults; (B) indicates unmated male adults; (C) indicates mated female adults; (D) indicates mated male adults; (E) indicates comparison of lifespan between female and male adults; (F) indicates comparison of lifespan between mated and unmated adults. Data presented are mean ± SD of five replicates. Different bar labels in Figure (A–D) and Figure (E,F) separately indicate significant differences according to Tukey’s test and Student’s t-test (p < 0.05). Female and male adult lifespan was recorded each day once they emerged as adults. Adults were considered dead if they did not move after being touched by an object. The total number (N) of unmated female adults was 146, 143, 142, 142, 132, 130, 129, 127, and 14; the total number (N) of unmated male adults was 144, 140, 139, 137, 128, 125, 117, 116, and 9 after pupae exposure to 25 °C or 34 °C for 1 h, 34 °C for 2 h, 34 °C for 4 h, 37 °C for 1 h, 37 °C for 2 h, 37 °C for 4 h, 40 °C for 1 h, and 40 °C for 2 h, respectively. The total numbers (N) of mated female and male adults were 30 and 30, respectively, under different test conditions.
Figure 3.
High-temperature stress impacts of adult lifespan in Bradysia odoriphaga pupae ((d) = days). (A) indicates unmated female adults; (B) indicates unmated male adults; (C) indicates mated female adults; (D) indicates mated male adults; (E) indicates comparison of lifespan between female and male adults; (F) indicates comparison of lifespan between mated and unmated adults. Data presented are mean ± SD of five replicates. Different bar labels in Figure (A–D) and Figure (E,F) separately indicate significant differences according to Tukey’s test and Student’s t-test (p < 0.05). Female and male adult lifespan was recorded each day once they emerged as adults. Adults were considered dead if they did not move after being touched by an object. The total number (N) of unmated female adults was 146, 143, 142, 142, 132, 130, 129, 127, and 14; the total number (N) of unmated male adults was 144, 140, 139, 137, 128, 125, 117, 116, and 9 after pupae exposure to 25 °C or 34 °C for 1 h, 34 °C for 2 h, 34 °C for 4 h, 37 °C for 1 h, 37 °C for 2 h, 37 °C for 4 h, 40 °C for 1 h, and 40 °C for 2 h, respectively. The total numbers (N) of mated female and male adults were 30 and 30, respectively, under different test conditions.
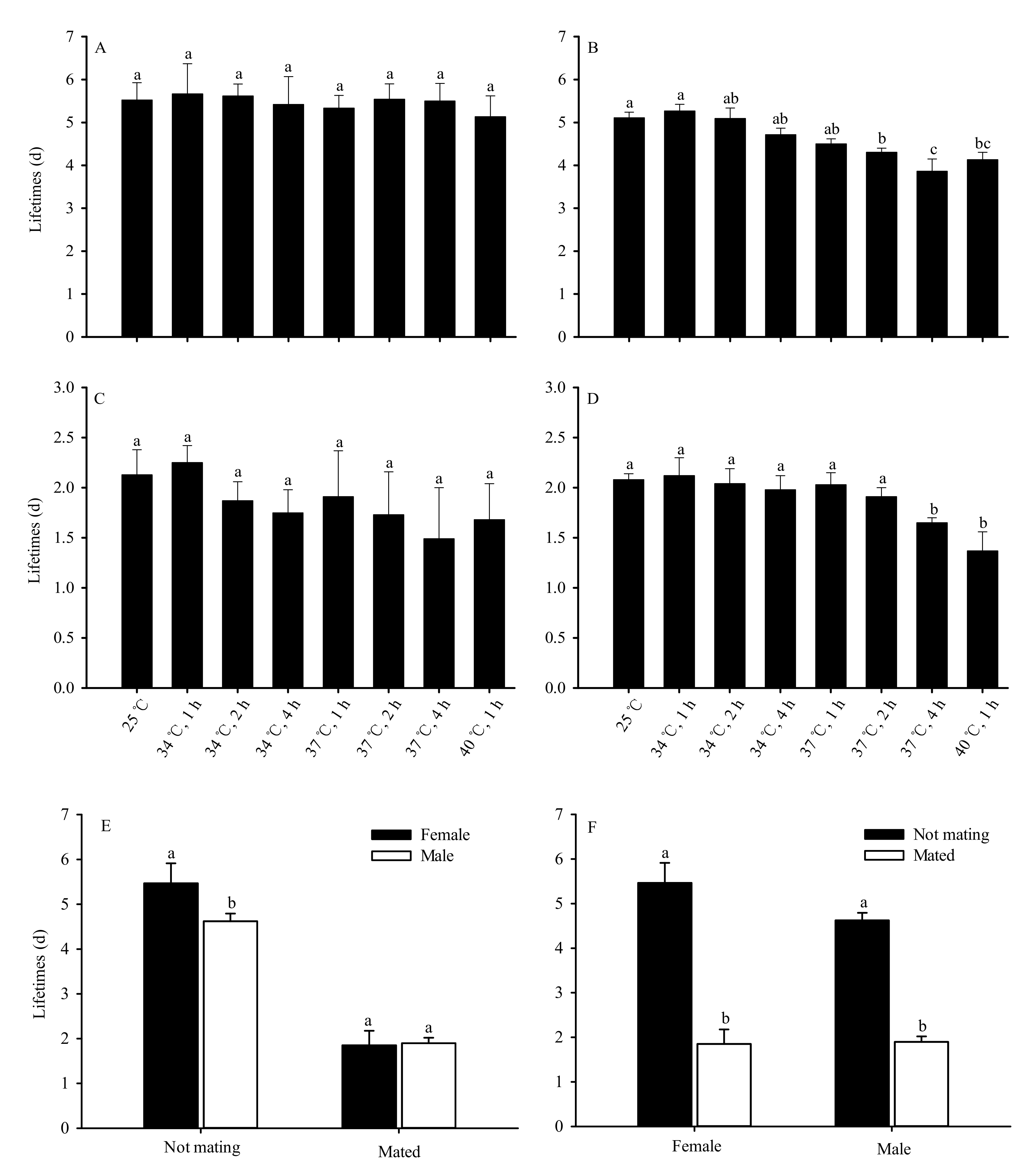
Figure 4.
High-temperature stress impacts on reproductive capacity (A) and egg hatchability (B) for mated female adults in Bradysia odoriphaga pupae. Data are mean ± SD of five replicates. Different bar labels indicate significant differences according to Tukey’s test (p < 0.05). Female and male adults were paired and placed into different Petri dishes (Φ = 60 mm). One pair was placed in each Petri dish, while six pairs were considered a replicate. Microscopy was used to count the number of larvae and eggs. On the condition that some of the eggs in one Petri dish successfully hatched into larvae, the number of eggs in that dish was recorded for subsequent statistical analysis.
Figure 4.
High-temperature stress impacts on reproductive capacity (A) and egg hatchability (B) for mated female adults in Bradysia odoriphaga pupae. Data are mean ± SD of five replicates. Different bar labels indicate significant differences according to Tukey’s test (p < 0.05). Female and male adults were paired and placed into different Petri dishes (Φ = 60 mm). One pair was placed in each Petri dish, while six pairs were considered a replicate. Microscopy was used to count the number of larvae and eggs. On the condition that some of the eggs in one Petri dish successfully hatched into larvae, the number of eggs in that dish was recorded for subsequent statistical analysis.
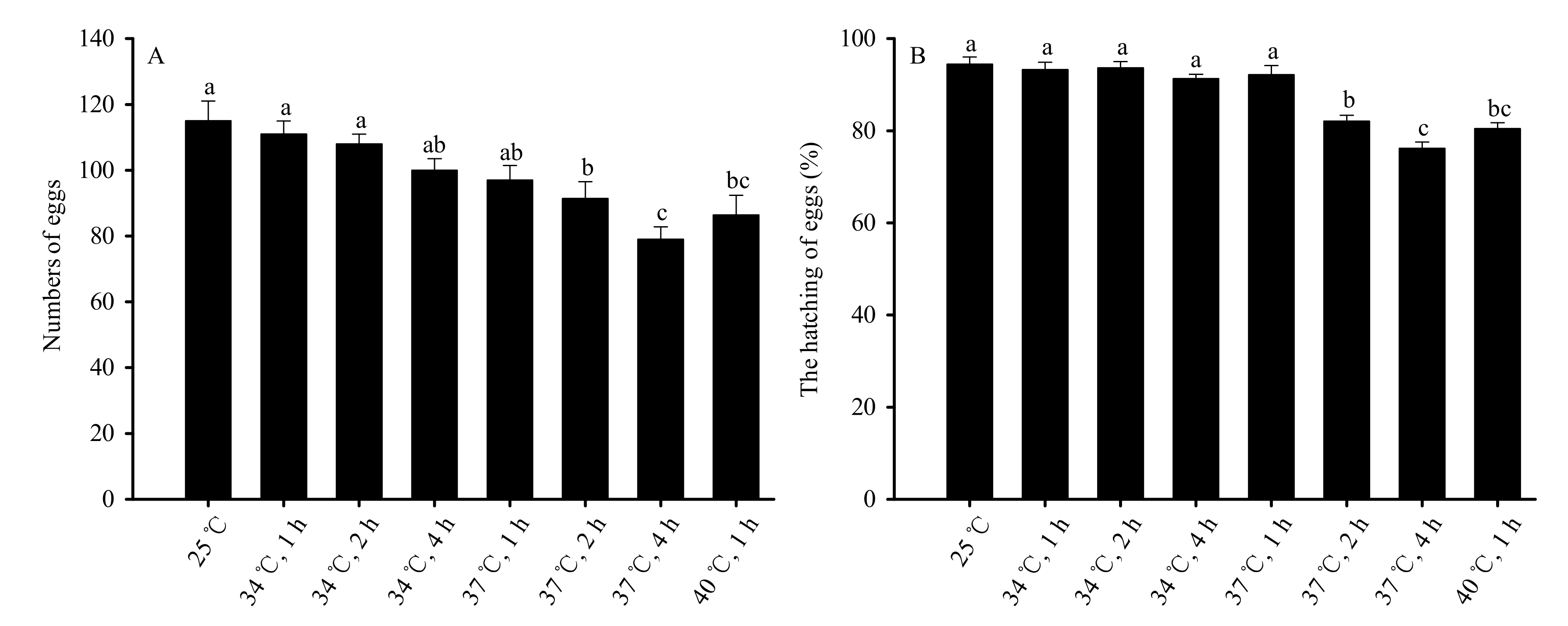
Figure 5.
Impacts on the length of testis and ovaries of unmated male and female adults after short-term high-temperature stress on pupae. Figure (A) represents ovary length; Figure (B) represents testis length. Values are averages ± SD of five replicates. Bars with different letters are significantly difference, based on Tukey’s test (p < 0.05) in each panel. Male and female adults (within 8 h of eclosion) were dissected, and the length of testis and ovaries was measured. The average length of six testis or ovaries in each treatment or the control was considered a replicate and was used for statistical analysis.
Figure 5.
Impacts on the length of testis and ovaries of unmated male and female adults after short-term high-temperature stress on pupae. Figure (A) represents ovary length; Figure (B) represents testis length. Values are averages ± SD of five replicates. Bars with different letters are significantly difference, based on Tukey’s test (p < 0.05) in each panel. Male and female adults (within 8 h of eclosion) were dissected, and the length of testis and ovaries was measured. The average length of six testis or ovaries in each treatment or the control was considered a replicate and was used for statistical analysis.
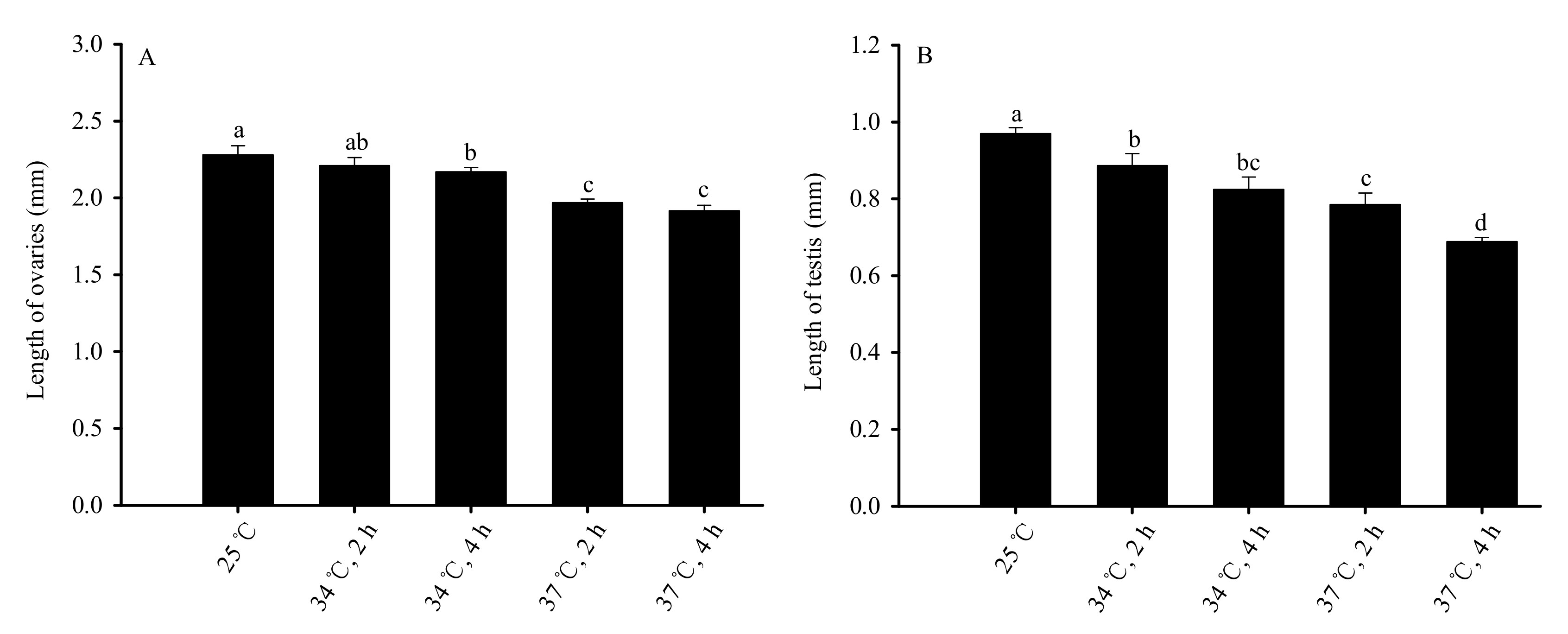
Figure 6.
Impacts on the length of reproductive organs after short-term high-temperature stress on Bradysia odoriphaga unmated adults. (A) represent the length of the ovaries recovery for 1 h; (B) represent the length of the ovaries recovery for 24 h; (C) represent the length of the testis recovery for 1 h; (D) represent the length of the testis recovery for 24 h; (E) represent that the length of the ovaries was compared between recovery for 1 h and 24 h; (F) represent that the length of the testis was compared between recovery for 1 h and 24 h. Data shown are mean ± SD of five replicates. Different bar labels indicate significant differences, based on Tukey’s test (p < 0.05). Female adults were dissected, and the length of ovaries and testis was measured. The average length of six ovaries and six testis from each treatment was separately considered a replicate and was used for statistical analysis.
Figure 6.
Impacts on the length of reproductive organs after short-term high-temperature stress on Bradysia odoriphaga unmated adults. (A) represent the length of the ovaries recovery for 1 h; (B) represent the length of the ovaries recovery for 24 h; (C) represent the length of the testis recovery for 1 h; (D) represent the length of the testis recovery for 24 h; (E) represent that the length of the ovaries was compared between recovery for 1 h and 24 h; (F) represent that the length of the testis was compared between recovery for 1 h and 24 h. Data shown are mean ± SD of five replicates. Different bar labels indicate significant differences, based on Tukey’s test (p < 0.05). Female adults were dissected, and the length of ovaries and testis was measured. The average length of six ovaries and six testis from each treatment was separately considered a replicate and was used for statistical analysis.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hu, J.; Medison, R.G.; Zhang, S.; Ma, P.; Shi, C. Impacts of Non-Lethal High-Temperature Stress on the Development and Reproductive Organs of Bradysia odoriphaga. Insects 2022, 13, 74. https://doi.org/10.3390/insects13010074
AMA Style
Hu J, Medison RG, Zhang S, Ma P, Shi C. Impacts of Non-Lethal High-Temperature Stress on the Development and Reproductive Organs of Bradysia odoriphaga. Insects. 2022; 13(1):74. https://doi.org/10.3390/insects13010074
Chicago/Turabian StyleHu, Jingrong, Rudoviko Galileya Medison, Seng Zhang, Peifang Ma, and Caihua Shi. 2022. "Impacts of Non-Lethal High-Temperature Stress on the Development and Reproductive Organs of Bradysia odoriphaga" Insects 13, no. 1: 74. https://doi.org/10.3390/insects13010074
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.