
An Analysis of the Possible Migration Routes of Oedaleus decorus asiaticus Bey-Bienko (Orthoptera: Acrididae) from Mongolia to China


 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Sources
2.1.1. Insect Information
2.1.2. Meteorological Data
2.2. Analysis of Three-Dimensional Trajectory and Atmospheric Circulation Fields Based on the WRF Model
2.2.1. WRF Model Initial Conditions
2.2.2. Analysis of the Atmospheric Circulation Background Fields of O. decorus asiaticus
2.2.3. The Choice of Moving in Peak Day
3. Results
3.1. Analysis of Population Dynamics under Light
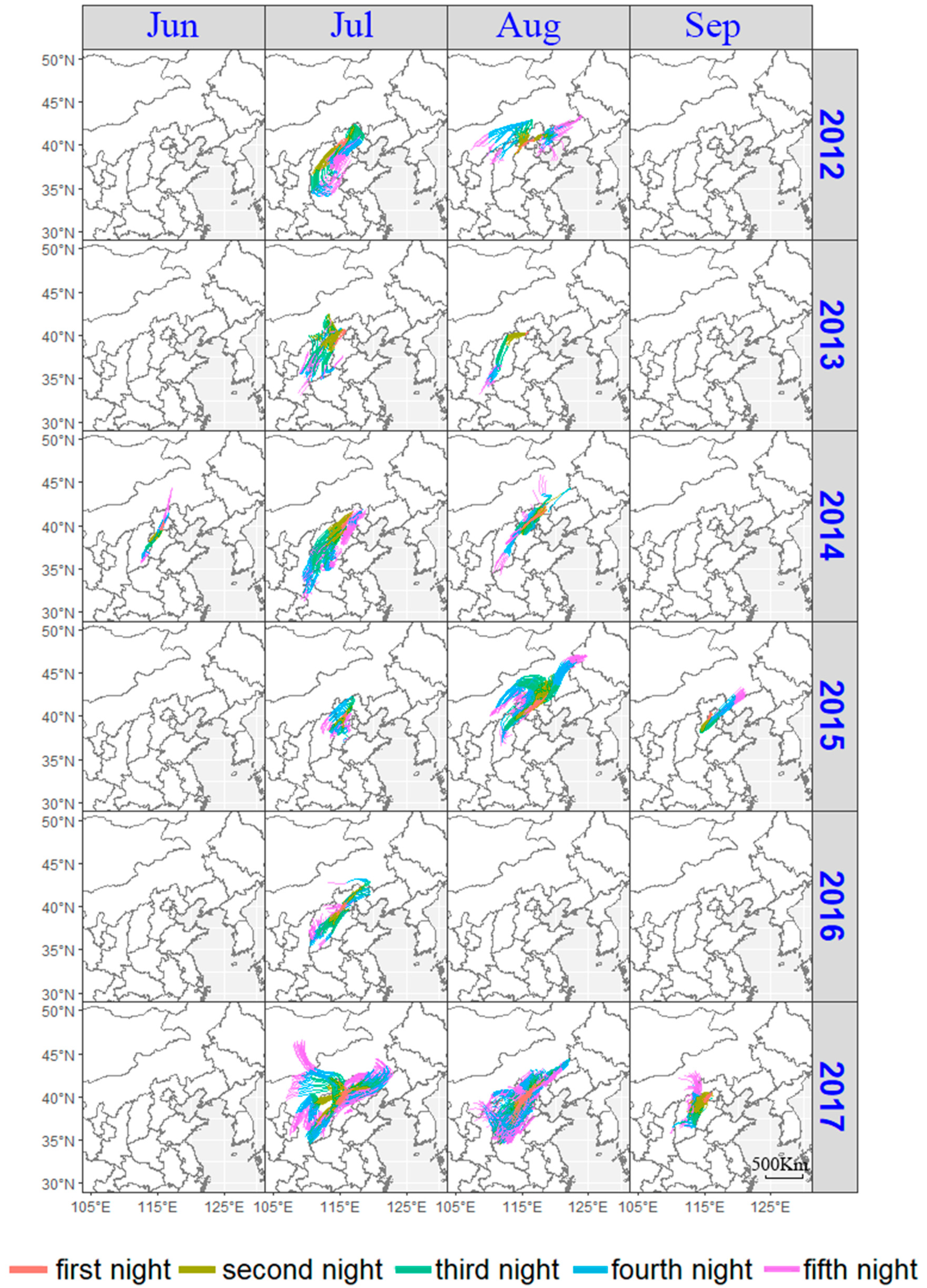
3.2. Analysis of the Possible Origin of Migrating O. decorus asiaticus Adults
3.3. Landing Mechanisms
3.3.1. Landing Mechanisms Affecting O. decorus asiaticus Migration in Yanqing, China
3.3.2. Case Analysis of Moving into the Peak on 8 July 2017
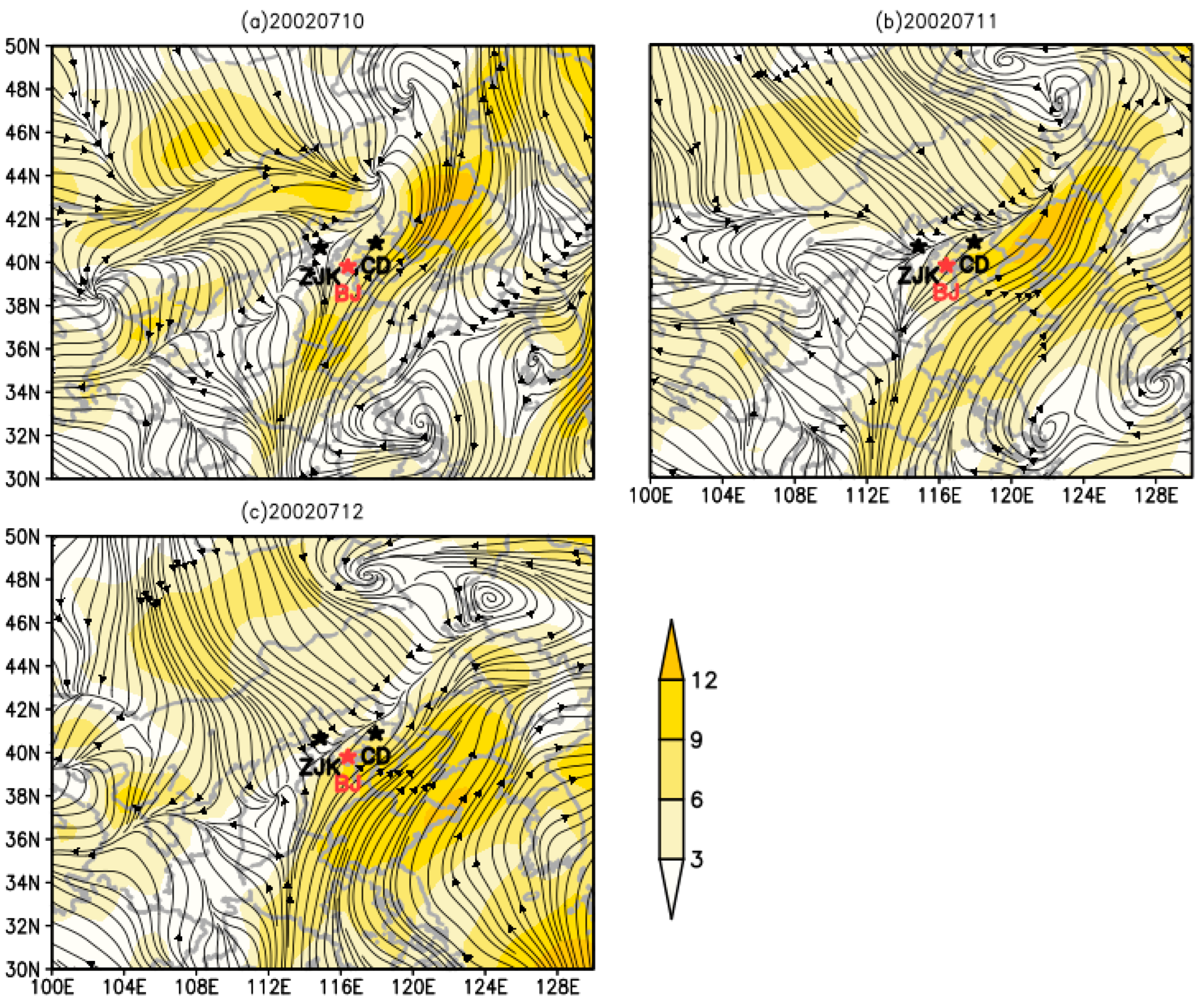
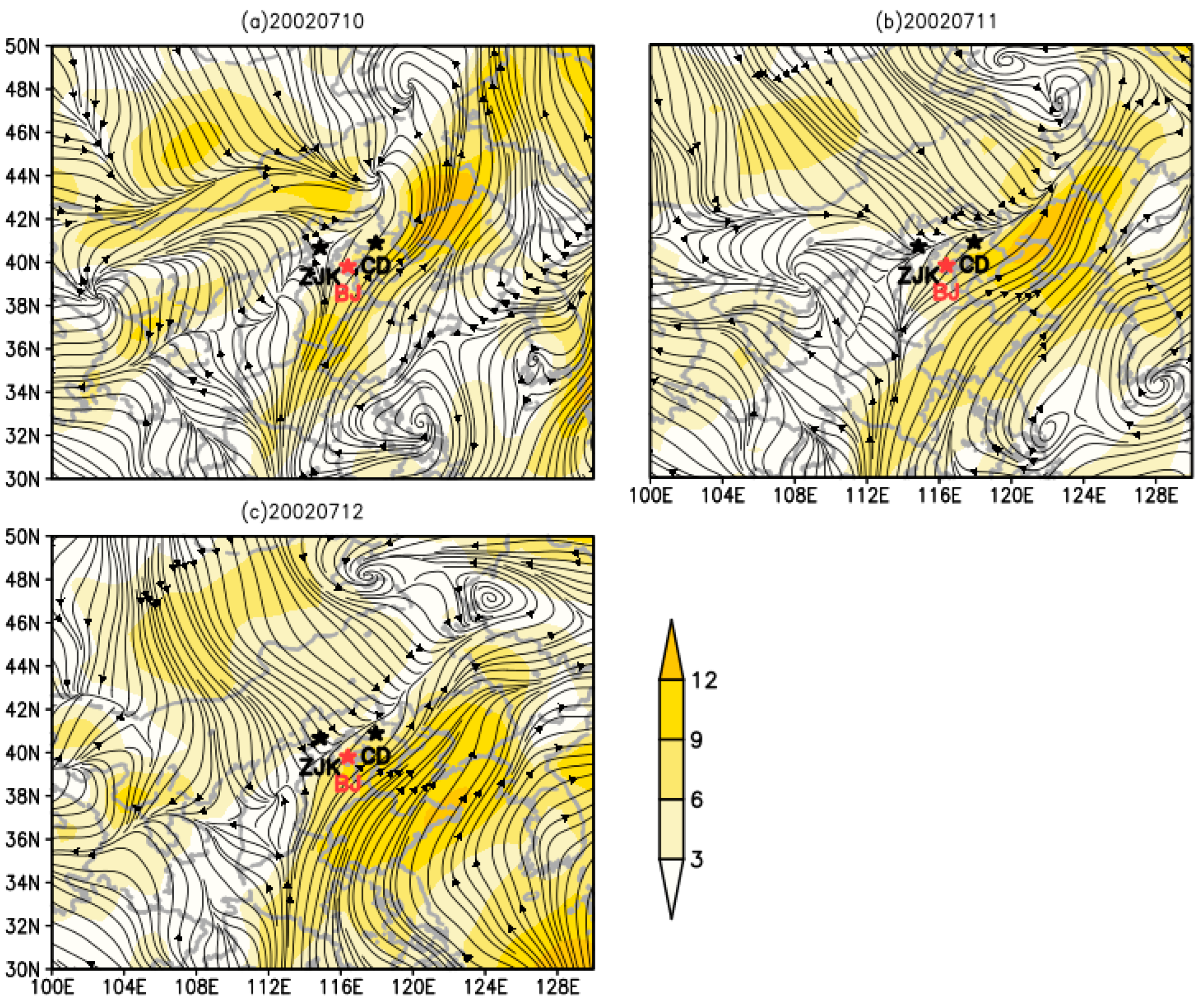
3.4. Dynamic Analysis of O. decorus asiaticus Migration in 2002
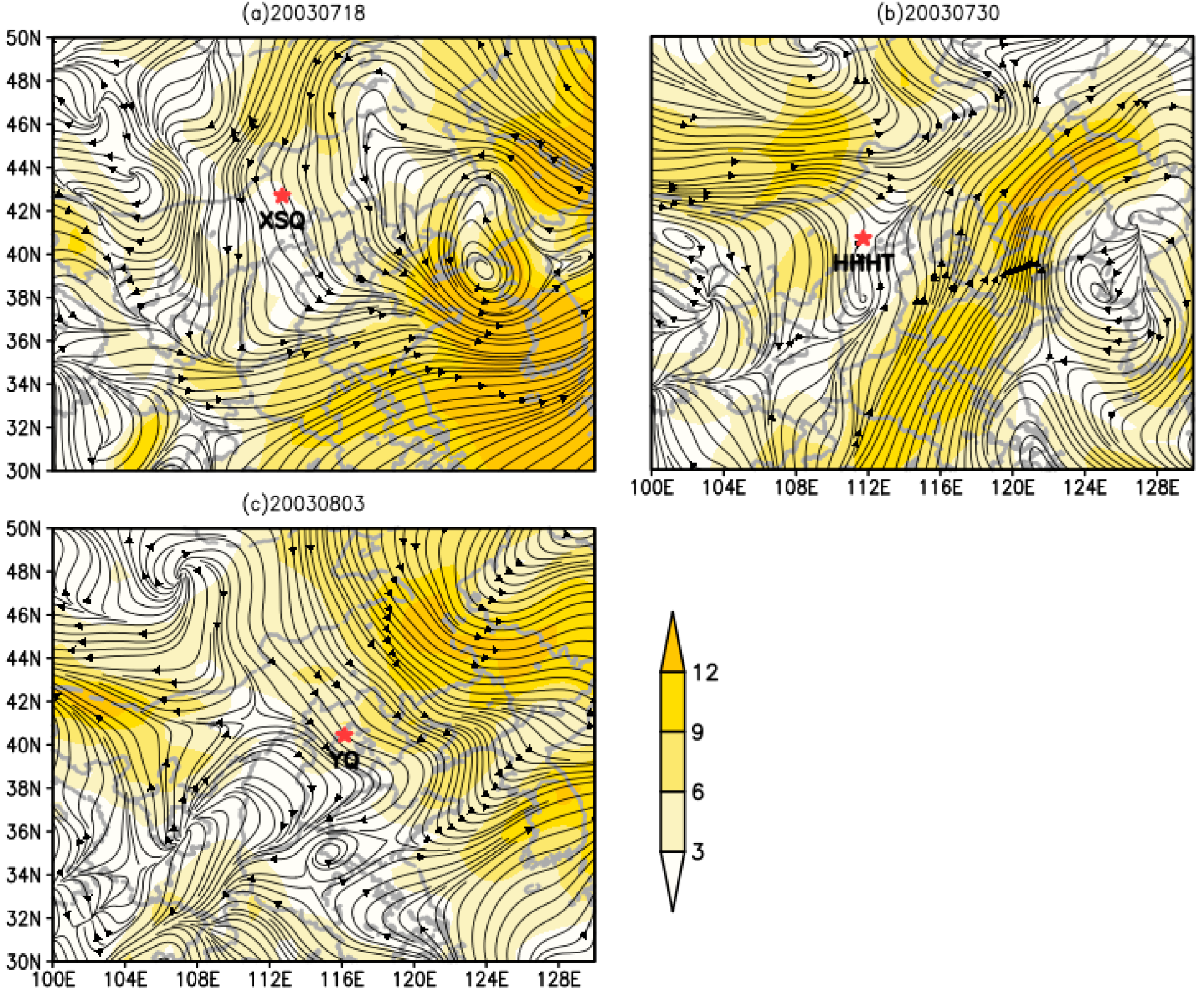
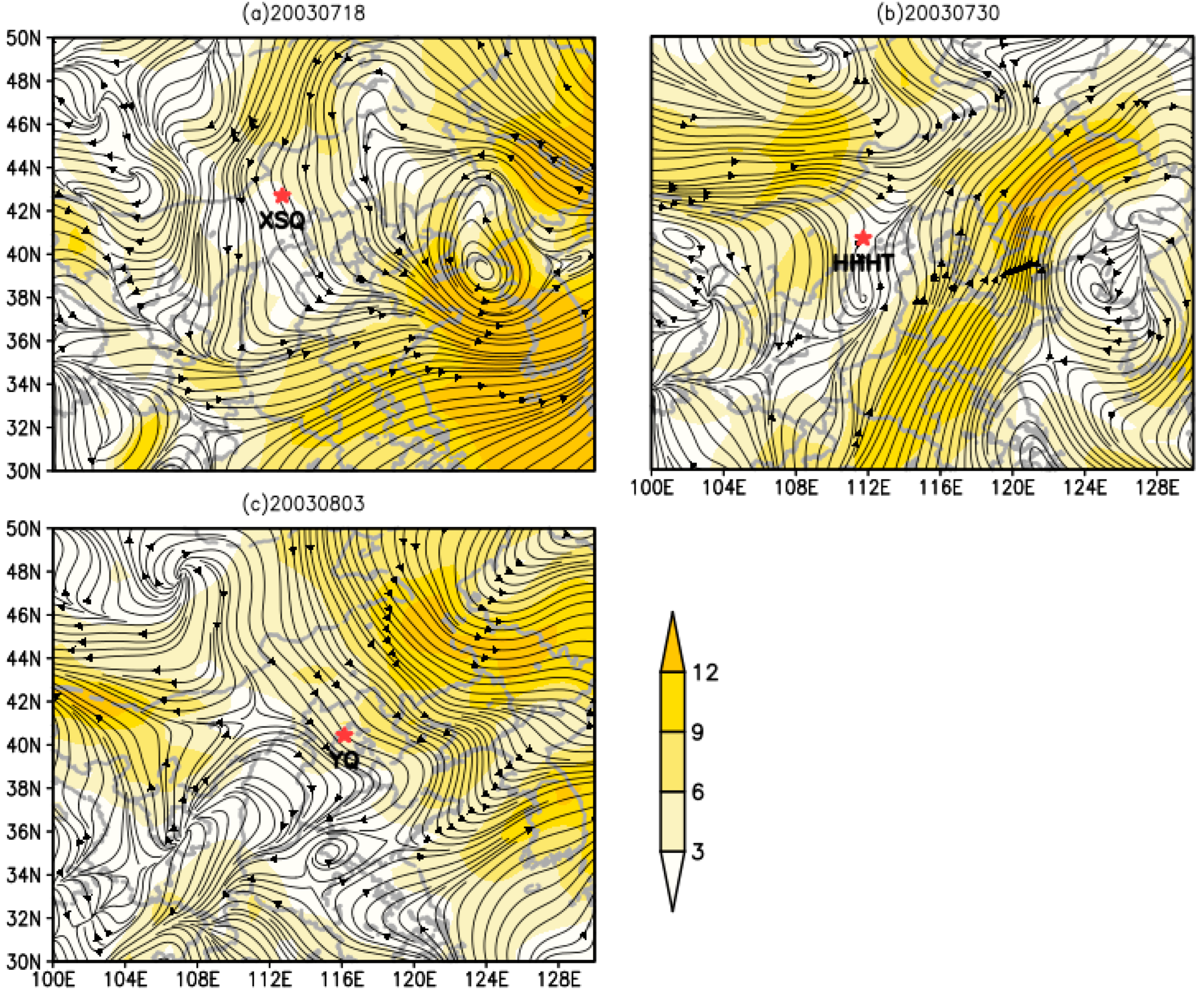
3.5. Dynamic Analysis of O. decorus asiaticus Migration in 2003
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Cui, B.Y.; Huang, X.B.; Li, S.; Hao, K.; Chang, B.H.; Tu, X.B.; Pang, B.P.; Zhang, Z.H. Quercetin Affects the Growth and Development of the Grasshopper Oedaleus asiaticus (Orthoptera: Acrididae). J. Econ. Entomol. 2019, 112, 1175–1182. [Google Scholar] [CrossRef]
- Zhang, L.; Lecoq, M.; Latchininsky, A.; Hunter, D. Locust and grasshopper management. Annu. Rev. Entomol. 2019, 64, 15–34. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.Q.; Wei, Z.Z. Determination of food intake of Oedaleus decorus asiaticus. Entomology. 1989, 26, 8–10. [Google Scholar]
- Jiang, X.; Mai, M.; Zhang, L. Nocturnal Migration of Grasshopper (Acrididae: Oedaleus asiaticus). Acta Geogr. Sin. 2003, 11, 75–77. [Google Scholar]
- Liu, L.; Guo, A.H. Analysis of Meteorological and Ecological Conditions of Grasshopper Infestation in Inner Mongolia in 2004. Meteorol. Appl. 2004, 30, 55–57. [Google Scholar] [CrossRef]
- Jago, N.D.; Ritchie, J.M.; Fishpool, L.D.C.; Rainy, R.C.; Daring, P.; Cheke, A.L. A Migrant Pest in the Sahel: The Senegalese Grasshopper Oedaleus senegalensis: Discussion. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1990, 328, 539–553. [Google Scholar] [CrossRef]
- Qiao, F. Oedaleus decorus asiaticus B.-Bienko, the main force of locust disaster moving northward. Man Biosph. 2005, 3, 25. [Google Scholar]
- Rankin, M.A.; Burchsted, J.C.A. The Cost of Migration in Insects. Annu. Rev. Entom. 1992, 37, 533–559. [Google Scholar] [CrossRef]
- Chapman, J.W.; Nesbit, R.L.; Burgin, L.E.; Reynolds, D.R.; Smith, A.D.; Middleton, D.R.; Hill, J.K. Flight orientation behaviors promote optimal migration trajectories in high-flying insects. Science 2010, 327, 682–685. [Google Scholar] [CrossRef]
- Chapman, J.; Klaassen, R.G.; Drake, V.; Fossette, S.; Hays, G.C.; Metcalfe, J.D.; Reynolds, A.M.; Reynolds, D.R.; Alerstam, T. Animal orientation strategies for movement in flows. Curr. Biol. 2011, 21, 861–870. [Google Scholar] [CrossRef] [Green Version]
- Crummay, F.A.; Atkinson, B.W. Atmospheric influences on light-trap catches of the brown planthopper rice pest. Agri. Forest Meteorol. 1997, 88, 181–197. [Google Scholar] [CrossRef]
- Westbrook, J.K.; Isard, S.A. Atmospheric scales of biotic dispersal. Agric. Meteorol. 1999, 97, 263–274. [Google Scholar] [CrossRef]
- Hu, G.; Bao, Y.X.; Wang, J.Q.; Zhai, B.P. Case studies on the landing mechanisms of the brown planthoppers Nilaparvata lugens (Stal). Acta Ecol. Sin. 2007, 27, 5068–5075. [Google Scholar]
- Farrow, R.A. Population Dynamics of the Australian Plague Locust, Chortoicetes Terminifera (Walker) in Central Western New South Wales Iii. Analysis of Population Processes. Aust. J. Zool. 1982, 30, 569–580. [Google Scholar] [CrossRef]
- Walker, T.J. Migrating Lepidoptera: Are butterflies better than moths? Fla. Entomol. 1980, 63, 79–98. [Google Scholar] [CrossRef]
- Zhang, X.X.; Geng, J.G.; Zhou, W.J. Ecological mechanism of rice leaf roller migration. J. Nanjing Agri. Univer. 1981, 4, 40–51. [Google Scholar]
- Wu, J.C. Effects of light, temperature and food on migration of Cnaphalocrocis medinalis. Acta Entomol. 1985, 28, 398–405. [Google Scholar]
- Dingle, H. Migration: The Biology of Life on the Move; Oxford University Press: Oxford, UK, 1996; pp. 375–413. [Google Scholar]
- Wang, Y.P.; Tu, X.B.; Lin, P.J.; Li, S.; Xu, C.M.; Wang, X.Q.; Reynolds, D.R.; Chapman, J.W.; Zhang, Z.H.; Hu, G. Migratory take-off behaviour of the Mongolian grasshopper Oedaleus asiaticus. Insects 2020, 11, 416. [Google Scholar] [CrossRef]
- Rainey, R.C. Weather and the movement of locust swarms: A new hypothesis. Nature 1951, 168, 1057–1060. [Google Scholar] [CrossRef]
- Wu, Q.L.; Hu, G.; Westbrook, J.K.; Sword, G.A.; Zhai, B.P. An advanced numerical trajectory model tracks a corn earworm moth migration event in Texas, USA. Insects 2018, 9, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y. Biology of Locusts in China; Shandong Science and Technology Press: Shandong, China, 1991; pp. 335–372. [Google Scholar]
- Tu, X.B.; Hu, G.; Fu, X.W.; Zhang, Y.H.; Ma, J.; Wang, Y.P.; Gould, P.; Du, G.L.; Su, H.T.; Zhang, Z.H.; et al. Mass windborne migrations extend the range of the migratory locust in East China. Agric. For. Entomol. 2020, 22, 41–49. [Google Scholar] [CrossRef]
- Hu, G.; Lu, F.; Lu, M.H.; Liu, W.C.; Xu, W.G.; Jiang, X.H.; Zhai, B.P. The influence of typhoon Khanun on the return migration of Nilaparvata lugens in eastern China. PLoS ONE 2013, 8, 57277. [Google Scholar] [CrossRef] [Green Version]
- Drake, V.A.; Farrow, R.A. The nocturnal migration of the Australian plague locust, Chortoicetes terminifera (Walker) (Orthoptera: Acrididae): Quantitative radar observations of a series of northward flights. Bull. Entomol. Res. 1983, 73, 567–585. [Google Scholar] [CrossRef]
- Hu, G.; Lu, M.H.; Tuan, H.A.; Liu, W.C.; Xie, M.C.; McInerney, C.E.; Zhai, B.P. Population dynamics of rice planthoppers, Nilaparvata lugens and Sogatella furcifera (Hemiptera, Delphacidae) in central Vietnam and its effects on their spring migration to China. Bull. Entomol. Res. 2017, 107, 369–381. [Google Scholar] [CrossRef]
- Wang, F.Y.; Yang, F.; Lu, M.H.; Luo, S.Y.; Zhai, B.P.; Lim, K.S.; McInerney, C.E.; Hu, G. Determining the migration duration of rice leaf folder (Cnaphalocrocis medinalis (Guenée)) moths using a trajectory analytical approach. Sci. Rep. 2017, 7, 39853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.X.; Zhou, L.Y.; Cheng, J.Y. Estimation of trajectory analysis parameters of Cnaphalocrocis medinalis in Jianghuai rice area. J. Nanjing Agric. Univ. 1994, 17, 32–38. [Google Scholar]
- Huis, A.V.; Cressman, K.; Magor, J.I. Preventing desert locust plagues: Optimizing management interventions. Entomol. Exp. Appl. 2010, 122, 191–214. [Google Scholar] [CrossRef]
- Qi, G.J.; Lu, F.; Gao, Y.; Liang, J.L.; Lan, R.Q.; Zhang, X.X.; Zhai, B.P.; Lv, L.H. Analysis of a migration process and the source population of Cnaphalocrocis medinalis (Guenée) (Lepidoptera: Pyralidae) in 2010. Acta Entomol. Sin. 2011, 54, 1194–1203. [Google Scholar]
- Hu, G.; Wu, Q.L.; Wu, X.W.; Jiang, Y.Y.; Zeng, J.; Zhai, B.P. Outbreak mechanism of second generation armyworms in northeastern China: A case study in 1978. Chin. J. Appl. Entomol. 2014, 51, 927–994. [Google Scholar]
- Ma, J.; Wang, Y.C.; Hu, Y.Y.; Lu, M.H.; Wan, G.J.; Chen, F.J.; Wan, C.L.; Zhai, B.P.; Hu, G. Brown planthopper Nilaparvata lugens (Stål) was concentrated at the rear of the typhoon Soudelor in Eastern China in August 2015. Insect Sci. 2017, 25, 916–926. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, Y.P.; Wu, M.F.; Gao, B.Y.; Liu, J.; Lee, G.S.; Qtuka, A.; Hu, G. High risk of the fall armyworm invading Japan and the Korean Peninsula via overseas migration. J. Appl. Entomol. 2019, 14, 911–920. [Google Scholar] [CrossRef]
- Zhai, B.P.; Zhang, X.X. Behavior of insects during migration. Acta Zool. Sin. 1993, 4, 440–446. [Google Scholar]
- Shen, H.M. The Study on Oversea Source Area of Nilaparvata lugens (Stal) and Sogatella furcifera (Horvfith) in China; Depaament of Entomology Nanjing Agricultural University: Nanjing, China, 2010. [Google Scholar]
- Shen, H.M.; Lu, J.P.; Zhou, J.Y.; Zhang, X.X.; Cheng, X.N.; Zhai, B.P. Source areas and landing mechanism of early immigration of white-backed planthoppers Sogatella furcifera (Horvbth) in Yunnan, 2009. Acta Eco. Sin. 2011, 31, 4350–4364. [Google Scholar]
- Jiang, G.H.; Tan, H.Q.; Shen, W.Z.; Cheng, X.N.; Chen, R.H. Meteorological conditions for long-distance northward migration of Nilaparvata lugens. Acta Ent. 1981, 03, 17–27. [Google Scholar]
- Bao, Y.X.; Wang, C.H.; Wang, J.Q.; Zhai, B.P. Research on the influence of wind field and precipitation on the landing process of main migratory pests in rice. In Proceedings of the Yangtze River Delta Science and Technology Forum-Yangtze River Delta Meteorological Technology Innovation and Development Forum, Jiangsu, China, 1 October 2006. [Google Scholar]
- Ritchie, J. A taxonomic revision of the genus Oedaleus Fieber (Orthoptera: Acrididae). Bull. Br. Mus. Nat. Hist. Zool. 1981, 42, 83–183. [Google Scholar]
- Ritchie, J. A taxonomic revision of the genus Gastrimargus Saussure (Orthoptera: Acrididae). Bull. Br. Mus. Nat. Hist. Zool. 1982, 44, 239–329. [Google Scholar]
- Ma, C.; Liu, C.; Yang, P.; Kang, L. The complete mitochondrial genomes of two band-winged grasshoppers, Gastrimargus marmoratus and Oedaleus asiaticus. BMC Genom. 2009, 10, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Yang, P.; Jiang, F.; Chapuis, M.P.; Shali, Y.; Sword, G.; Kang, L. Mitochondrial genomes reveal the global phylogeography and dispersal routes of the migratory locust. Mol. Ecol. 2012, 21, 4344–4358. [Google Scholar] [CrossRef]
- Elamin, A.E.H.; Abdalla, A.M.; Naim, A.M.E. Studies on population dynamics of senegalese grasshopper (Oedaleus senegalensis) in Kordofan of Sudan. World 2013, 1, 85–89. [Google Scholar] [CrossRef]
- Xu, F.Z.; Meng, Z.P.; Guo, Y.H.; Shen, J.P.; Han, C.Z. Occurrence and control of Oedaleus decorus asiaticus B.-Bienko in the ecotone of agriculture and animal husbandry in Ulanqab City, China. Inn. Mong. Agric. Sci. Technol. 2005, S2, 384–387. [Google Scholar]
- Yang, X.Y.; Xu, S.Q.; Zhang, L.; Qin, Y.L.; Pan, M.T. Interdecadal Changes of Cyclone Activities in the North of East Asia during Boreal Spring. In Proceedings of the 2018 3rd International Conference on Modelling, Simulation and Applied Mathematics (MSAM 2018), Shanghai, China, 22–23 July 2018; Atlantis Press: Paris, France, 2018; pp. 277–281. [Google Scholar]
- Lee, J.; Son, S.W.; Cho, H.O.; Kim, J.; Cha, D.H.; Gyakum, J.R.; Chen, D. Extratropical cyclones over East Asia: Climatology, seasonal cycle, and long-term trend. Clim. Dyn. 2020, 54, 1131–1144. [Google Scholar] [CrossRef]
- Sergeev, M.G.; Storozhenko, S.Y.; Benediktov, A.A. An annotated check-list of Orthoptera of Tuva and adjacent regions. Part 3. Suborder Caelifera (Acrididae: Gomphocerinae: Gomphocerini; Locustinae). Far East. Entomol. 2020, 402, 1–36. [Google Scholar]
- Farrow, R.A. A modified light-trap for obtaining large samples of night-flying locusts and grasshopper. Austral. Entomol. 1974, 13, 357–360. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Domain1 |
|---|---|
| Location | 40° N, 116° E |
| The number grid points | 99 × 99 |
| Distance between grid points (km) | 30 |
| Layers | 30 |
| Map projection | Lambert |
| Microphysics scheme | WSM6 |
| Longwave radiation scheme | RRTMG |
| Shortwave radiation scheme | RRTMG |
| Surface layer scheme | Monin-Obukhow |
| Land/water surface scheme | Noah |
| Planetary boundary layer scheme | YSU |
| Cumulus parameterization | Tiedtke |
| Forecast time | 72 h |
| Year | Date of First Capture | Date of Final Capture | Duration (d) | Date of Peak Catches (n) | Time of Maximum Capture | Total Catches | |||
|---|---|---|---|---|---|---|---|---|---|
| Number (n) | Time | Number (n) | Time | Number (n) | Time | ||||
| 2012 | 19 July (1) | 20:00–21:00 | 23 August (1) | 01:00–02:00 | 36 | 20 July (4) | 20:00–21:00 | 20:00–21:00 | 11 |
| 2013 | 8 July (1) | 22:00–23:00 | 24 August–25 August (1) | 24:00–01:00 | 49 | 16 August (4) | 20:00–21:00 | 20:00–21:00 | 21 |
| 2014 | 30 June (1) | 22:00–23:00 | 19 August (1) | 21:00–22:00 | 52 | 19 August–20 August (7) | 24:00–01:00 | 24:00–01:00 | 32 |
| 19 August–20 August (9) | 23:00–01:00 | ||||||||
| 2015 | 13 July (1) | 21:00–22:00 | 4 September (2) | 20:00–22:00 | 53 | 15 August (3) | 20:00–21:00 | 20:00–21:00 | 34 |
| 15 August–20 August (8) | 22:00–02:00 | ||||||||
| 2016 | 27 July (1) | 01:00–02:00 | 30 July (1) | 02:00–03:00 | 4 | - | - | - | 2 |
| 2017 | 8 July (3) | 21:00–23:00 | 11 September (2) | 20:00–21:00 | 66 | 15 August–16 August (141) | 20:00–02:00 | 20:00–21:00 | 566 |
| 16 August (1) | 04:00–05:00 | ||||||||
| Duration | Total | Valid Trajectories | Overrange Trajectories |
|---|---|---|---|
| 1 | 672 | 539 | 133 |
| 2 | 4312 | 3210 | 1102 |
| 3 | 3210 | 3083 | 127 |
| 4 | 3083 | 3020 | 63 |
| 5 | 3020 | 2967 | 53 |
| Total | 14,297 | 12,819 | 1478 |
| Year | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trajectories | Valid Trajectories | Overrange Trajectories | Valid Trajectories | Overrange Trajectories | Valid Trajectories | Overrange Trajectories | Valid Trajectories | Overrange Trajectories | Valid Trajectories | Overrange Trajectories | Valid Trajectories | Overrange Trajectories |
| Inner Mongolia | 83 | 1 | 24 | 3 | 43 | 7 | 546 | 23 | 15 | 3 | 1091 | 12 |
| Beijing | 78 | 12 | 1 | 6 | 169 | 33 | 68 | 27 | 17 | 7 | 981 | 178 |
| Ningxia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Jilin | 5 | 0 | 0 | 0 | 1 | 0 | 4 | 0 | 0 | 0 | 0 | 0 |
| Tianjin | 7 | 0 | 0 | 0 | 9 | 0 | 11 | 0 | 0 | 0 | 12 | 0 |
| Shandong | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 0 |
| Shanxi | 139 | 21 | 193 | 9 | 336 | 38 | 142 | 3 | 146 | 11 | 2156 | 113 |
| Hebei | 444 | 78 | 42 | 52 | 598 | 90 | 583 | 157 | 79 | 20 | 3722 | 554 |
| Henan | 39 | 0 | 9 | 0 | 93 | 0 | 0 | 0 | 0 | 0 | 282 | 6 |
| Hubei | 0 | 0 | 0 | 0 | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Chongqing | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Shaanxi | 5 | 0 | 70 | 0 | 33 | 0 | 0 | 0 | 12 | 0 | 137 | 0 |
| Liaoning | 83 | 0 | 0 | 0 | 0 | 0 | 34 | 0 | 4 | 0 | 150 | 0 |
| Heilongjiang | 0 | 0 | 0 | 0 | 0 | 0 | 39 | 0 | 0 | 0 | 0 | 0 |
| Gansu | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 20 | 0 |
| Mongolia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 49 | 0 |
| Sea | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 |
| Total | 890 | 112 | 339 | 70 | 1296 | 168 | 1427 | 210 | 273 | 41 | 8608 | 863 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, S.; Du, G.; Hu, G.; Zhang, Y.; Tu, X.; Zhang, Z. An Analysis of the Possible Migration Routes of Oedaleus decorus asiaticus Bey-Bienko (Orthoptera: Acrididae) from Mongolia to China. Insects 2022, 13, 72. https://doi.org/10.3390/insects13010072
Wang Y, Li S, Du G, Hu G, Zhang Y, Tu X, Zhang Z. An Analysis of the Possible Migration Routes of Oedaleus decorus asiaticus Bey-Bienko (Orthoptera: Acrididae) from Mongolia to China. Insects. 2022; 13(1):72. https://doi.org/10.3390/insects13010072
Chicago/Turabian StyleWang, Yunping, Shuang Li, Guilin Du, Gao Hu, Yunhui Zhang, Xiongbing Tu, and Zehua Zhang. 2022. "An Analysis of the Possible Migration Routes of Oedaleus decorus asiaticus Bey-Bienko (Orthoptera: Acrididae) from Mongolia to China" Insects 13, no. 1: 72. https://doi.org/10.3390/insects13010072