GOBP1 Plays a Key Role in Sex Pheromones and Plant Volatiles Recognition in Yellow Peach Moth, Conogethes punctiferalis (Lepidoptera: Crambidae)

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. RNA Extraction and cDNA Synthesis
2.3. Quantitative Real-Time PCR (qRT-PCR)
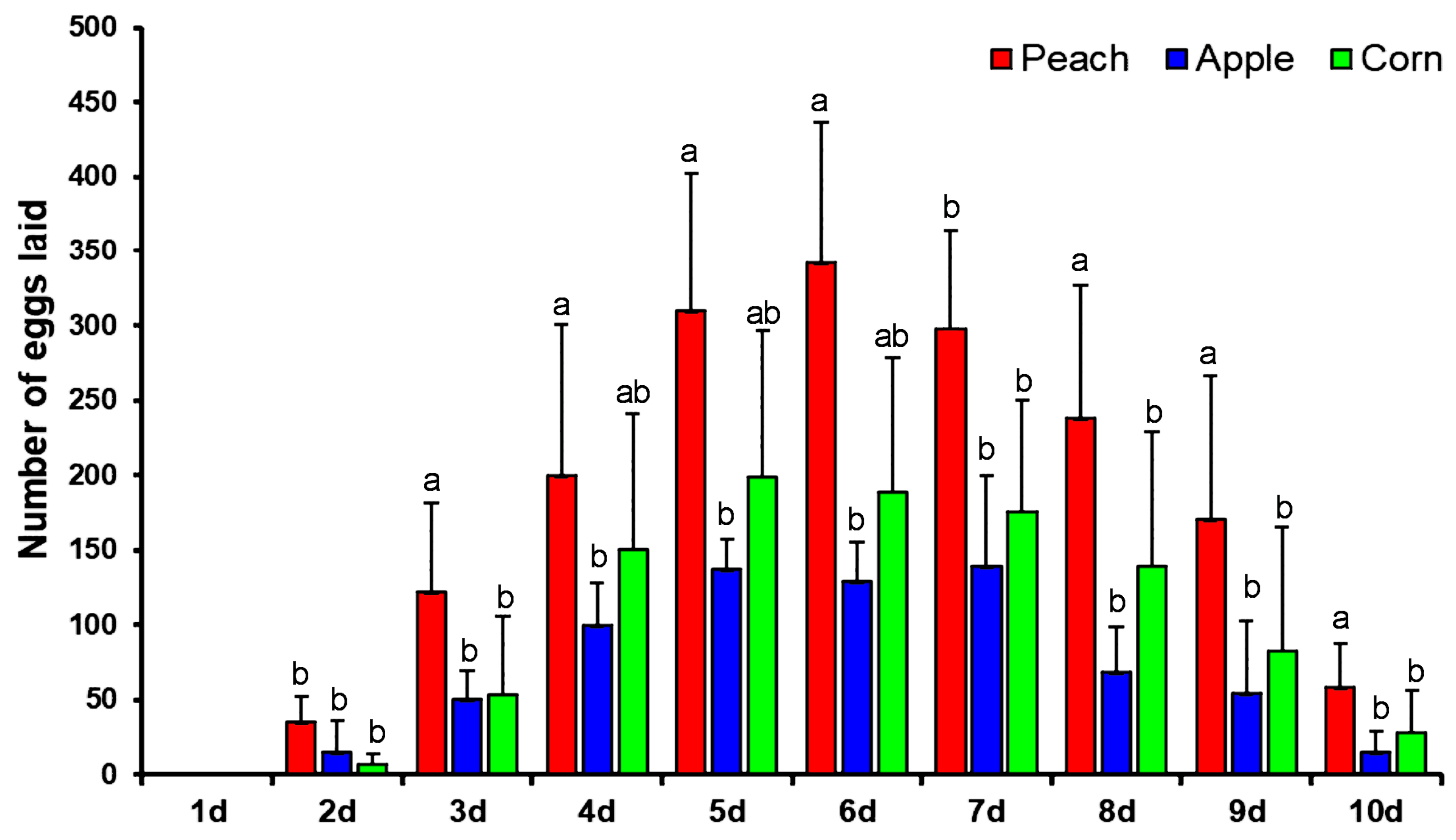
2.4. Oviposition Preference Test
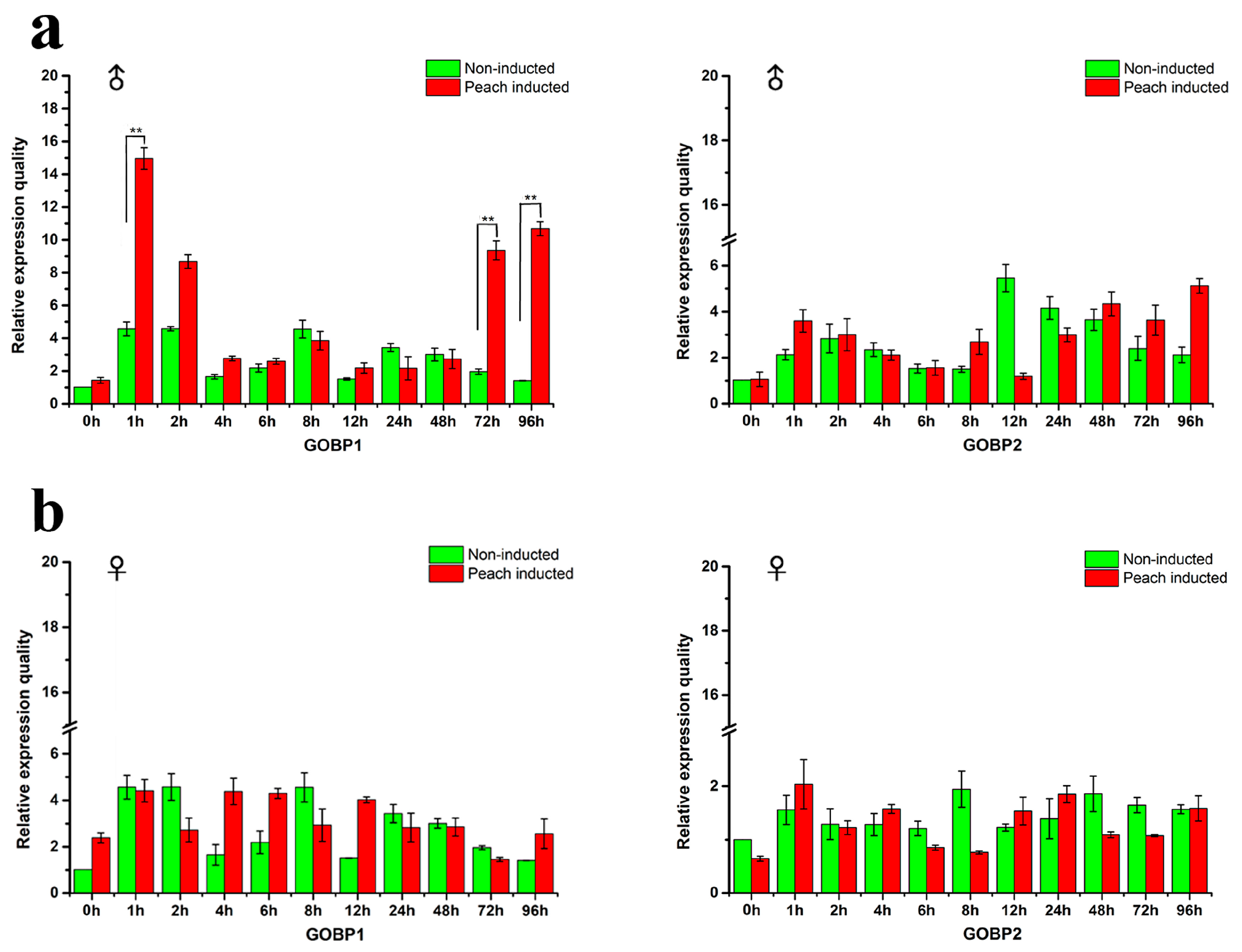
2.5. Effects of Host Stimulation on the Expression Profiles of GOBP Genes
2.6. Preparation of Recombinant GOBPs
2.7. Fluorescence Binding Assays
2.8. CD Spectra, 3D Modeling, and Molecular Docking Studies
3. Results
3.1. qRT-PCR
3.2. Oviposition Preference
3.3. Expression Levels of GOBP1 and GOBP2 Genes
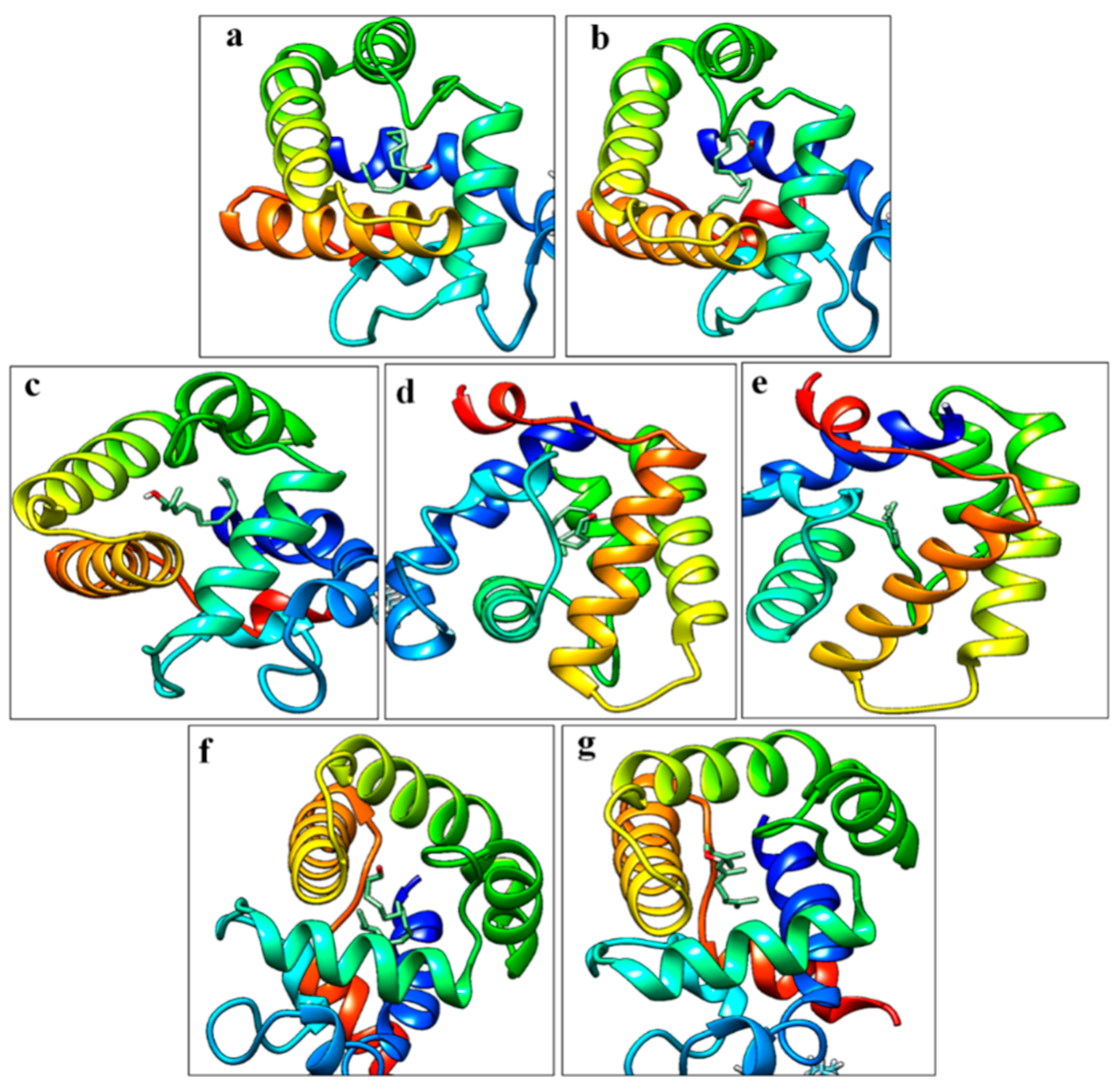
3.4. Heterologous Expression and Structure Analysis GOBPs
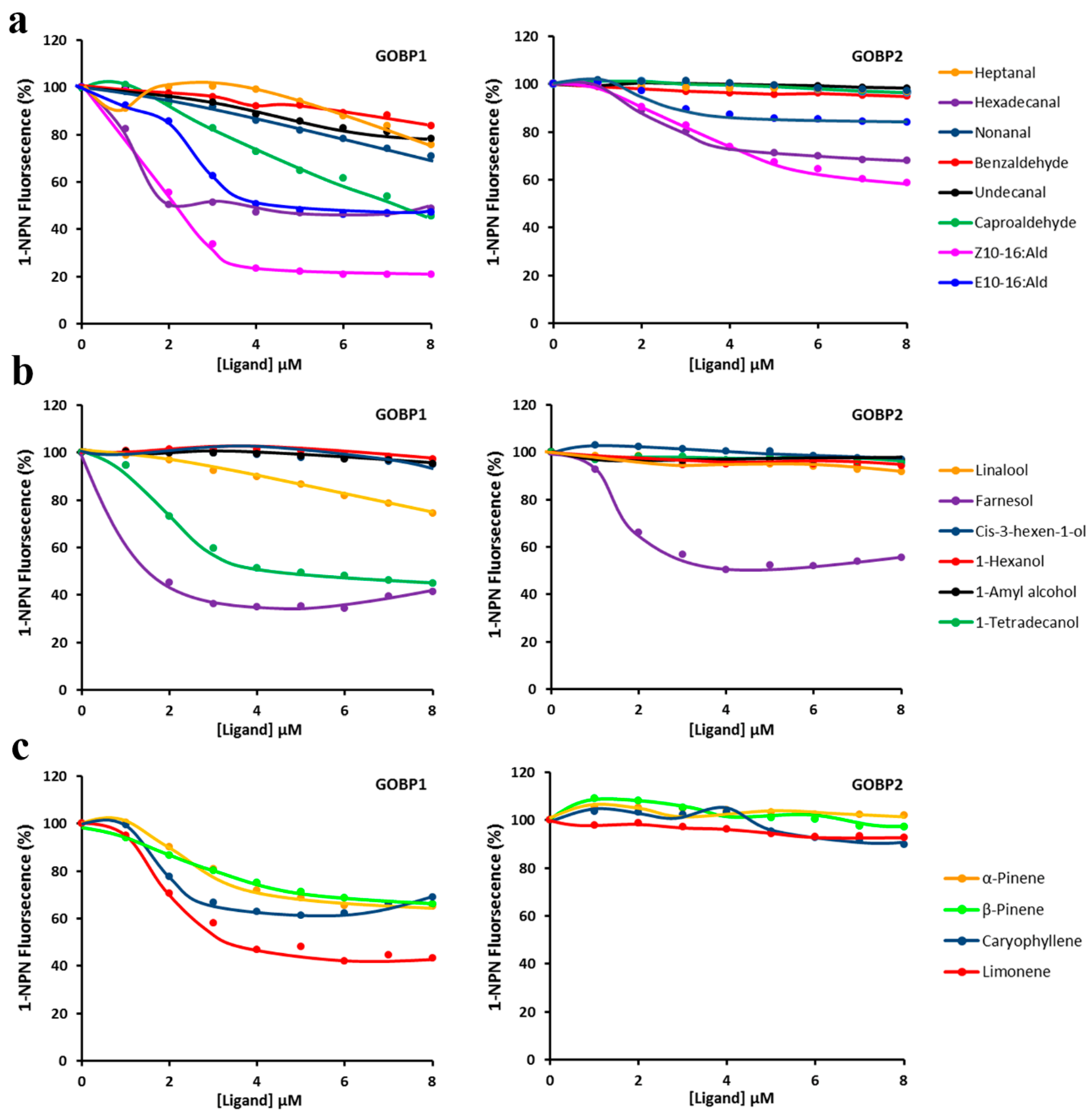
3.5. Flourescent Ligand-Binding Assays
3.6. Molecular Docking Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kaissling, K.E. Pheromone deactivation catalyzed by receptor molecules: A quantitative kinetic model. Chem. Senses 1998, 23, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Krieger, J.; Breer, H. Olfactory reception in invertebrates. Science 1999, 286, 720–723. [Google Scholar] [CrossRef] [PubMed]
- Callahan, F.E.; Vogt, R.G.; Tucker, M.L.; Dickens, J.C.; Mattoo, A.K. High level expression of “male specific” pheromone binding proteins (PBPs) in the antennae of female noctuiid moths. Insect Biochem. Mol. Biol. 2000, 30, 507–514. [Google Scholar] [CrossRef]
- Calvello, M.; Brandazza, A.; Navarrini, A.; Dani, F.R.; Turillazzi, S.; Felicioli, A.; Pelosi, P. Expression of odorant-binding proteins and chemosensory proteins in some Hymenoptera. Insect Biochem. Mol. Biol. 2005, 35, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.J.; Kan, Y.; Antoniw, J.; Pickett, J.A.; Field, L.M. Genome and EST analyses and expression of a gene family with putative functions in insect chemoreception. Chem. Senses. 2006, 31, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Krieger, J.; Gänssle, H.; Raming, K.; Breer, H. Odorant binding proteins of Heliothis virescens. Insect Biochem. Mol. Biol. 1993, 23, 449–456. [Google Scholar] [CrossRef]
- Jia, X.J.; Wang, H.X.; Yan, Z.G.; Zhang, M.Z.; Wei, C.H.; Qin, X.C.; Ji, W.R.; Du, Y.L. Antennal transcriptome and differential expression of olfactory genes in the yellow peach moth, Conogethes punctiferalis (Lepidoptera: Crambidae). Sci. Rep. 2016, 6, 29067. [Google Scholar] [CrossRef] [PubMed]
- Vogt, R.G.; Rybczynski, R.; Lerner, M.R. Molecular cloning and sequencing of general odorant-binding proteins GOBP1 and GOBP2 from the tobacco hawk moth Manduca sexta: Comparisons with other insect OBPs and their signal peptides. J. Neurosci. 1991, 11, 2972–2984. [Google Scholar] [CrossRef] [PubMed]
- Laue, M.; Steinbrecht, R.A.; Ziegelberger, G. Immunocytochemical localization of general odorant-binding protein in olfactory sensilla of the silkmoth Antheraea polyphemus. Naturwissenschaften 1994, 81, 178–180. [Google Scholar] [CrossRef]
- Liu, N.Y.; Yang, F.; Yang, K.; He, P.; Niu, X.H.; Xu, W.; Anderson, A.; Dong, S.L. Two subclasses of odorant-binding proteins in Spodoptera exiguadisplay structural conservation and functional divergence. Insect Mol. Biol. 2014, 24, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Khuhro, S.A.; Liao, H.; Dong, X.T.; Yu, Q.; Yuan, Q.; Dong, S.L. Two general odorant binding proteins display high bindings to both host plant volatiles and sex pheromones in a pyralid moth Chilo suppressalis (Lepidoptera: Pyralidae). J. Asia-Pac. Entomol. 2017, 20, 521–528. [Google Scholar] [CrossRef]
- Ma, M.; Chang, M.M.; Lei, C.L.; Yang, F.L. A garlic substance disrupts odorant-binding protein recognition of insect pheromones released from adults of the angoumois grain moth, Sitotroga cerealella (Lepidoptera: Gelechiidae). Insect Mol. Biol. 2016, 25, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Kubli, E. Sex-peptides: Seminal peptides of the Drosophila male. Cell. Mol. Life Sci. 2003, 60, 1689–1704. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, C.; Dickson, B.J. Sex peptide receptor and neuronal TOR/S6K signaling modulate nutrient balancing in Drosophila. Curr. Biol. 2010, 20, 1000–1005. [Google Scholar] [CrossRef] [PubMed]
- Devaud, J.M.; Acebes, A.; Ramaswami, M.; Ferrús, A. Structural and functional changes in the olfactory pathway of adult Drosophila take place at a critical age. J. Neurobiol. 2003, 56, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Mechaber, W.L.; Capaldo, C.T.; Hildebrand, J.G. Behavioral responses of adult female tobacco hornworms, Manduca sexta, to hostplant volatiles change with age and mating status. J. Insect. Sci. 2002, 2, 5. [Google Scholar] [CrossRef]
- Paula, D.P.; Togawa, R.C.; do Carmo Costa, M.M.; Grynberg, P.; Martins, N.F.; Andow, D.A. Systemic and sex-biased regulation of OBP expression under semiochemical stimuli. Sci. Rep. 2018, 8, 6035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquin-Joly, E.; Legeai, F.; Montagné, N.; Monsempes, C.; François, M.C.; Poulain, L.; Gavory, F.; Larsson, M.C. Candidate chemosensory genes in female antennae of the noctuid moth Spodoptera littoralis. Int. J. Biol. Sci. 2012, 8, 1036–1050. [Google Scholar] [CrossRef]
- Pena, J.; Nadel, H.; Pereira, M.; Smith, D. Tropical Fruit pests and pollinators: Biology, economic importance, natural enemies, and control. In Pollinators and pests of Annona species; Peña, J., Sharp, J., Wysoki, M., Eds.; CABI Publishing: Wallingford, UK, 2002; pp. 197–221. [Google Scholar]
- CABI. Conogethes punctiferalis datasheet. In Crop Protection Compendium; CABI Publishing: Wallingford, UK, 2011. [Google Scholar]
- Chakravarthy, A.; Vasudevkammar, K.K. Monograph on shoot and fruit borer, Conogethes punctiferalis and allied species. In Consortia Research Platform on Borers; ICAR-Indian; Institute of Horticultural Research (IIHR): Bengaluru, Indian, 2015. [Google Scholar]
- Konno, Y.; Honda, N.; Matsumoto, Y. Mechanism of reproductive isolation between the fruit feeding and pin ace feeding type of yellow peach moth, Dichocrocis punctiferalis. Jpn. J. Appl. Entomol. Zool. 1981, 25, 253–258. [Google Scholar] [CrossRef]
- CPCI. Crop Protection Compendium on Internet accessed in 2005; CABI Publishing: Wallingford, UK; Available online: www.cabi.org/cpc.
- Konno, Y.; Arai, K.; Sekiguchi, K.; Matsumoto, Y. (E)-10-Hexadecenal, a sex pheromone component of the yellow peach moth, Dichocrocis punctiferalis Guenée (Lepidoptera: Pyralidae). Appl. Entomol. Zoolog. 1982, 17, 207–217. [Google Scholar] [CrossRef]
- Liu, M.; Tian, Y.; Li, Y. Identification of minor components of the sex pheromone of yellow peach moth Dichocrocis punctiferalis Guenée and field trials. Insect Sci. 1994, 1, 150–155. [Google Scholar] [CrossRef]
- Kyungsaeng, B.; Park, K.C. Insect semiochemical research in Korea: Overview and prospects. Appl. Entomol. Zool. 2005, 40, 13–29. [Google Scholar] [CrossRef]
- Rützler, M.; Zwiebel, L. Molecular biology of insect olfaction: Recent progress and conceptual models. J. Comp. Physiol. A 2005, 191, 777–790. [Google Scholar] [CrossRef]
- Menini, A. The Neurobiology of Olfaction; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2010. [Google Scholar]
- Ge, X.; Ahmed, T.; Zhang, T.; Wang, Z.; He, K.; Bai, S. Binding specificity of two PBPs in the yellow peach moth Conogethes punctiferalis (Guenée). Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Martin, N.; Moore, K.; Musto, C.J.; Linn, C.E. Flight tunnel response of male European corn borer moths to cross-specific mixtures of European and Asian corn borer sex pheromones: Evidence supporting a critical stage in evolution of a new communication system. J. Chem. Ecol. 2016, 42, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Braccini, C.L.; Vega, A.S.; Aráoz, M.V.; Teal, P.E.; Cerrillo, T.; Zavala, J.A.; Fernandez, P.C. Both volatiles and cuticular plant compounds determine oviposition of the willow sawfly Nematus oligospilus on Leaves of Salix spp. (Salicaceae). J. Chem. Ecol. 2015, 41, 985–996. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Rui, C.H.; Yang, D.B.; Wang, Z.Y.; Yuan, H.Z. De novo transcriptome and expression profile analyses of the Asian corn borer (Ostrinia furnacalis) reveals relevant flubendiamide response genes. BMC Genom. 2017, 18, 20. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.; Benes, V.; Garson, J.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.; Shipley, G.; et al. The MIQE guidelines: Minimum information for publication of quantitative Real-Time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Hu, X.J.; Xu, Z.F.; He, L.; Xiao, W. Screening of reference genes for qRT-PCR in Conogethes punctiferails (Lepidoptera: Crambidae). Acta Entomol. Sin. 2017, 11, 1266–1277. [Google Scholar] [CrossRef]
- Glaser, N.; Frérot, B.; Leppik, E.; Monsempes, C.; Claire, C.D.; Ru, B.L.; Lecocq, T.; Calatayud, P.A. Similar differentiation patterns between PBP expression levels and pheromone component ratios in two populations of Sesamia nonagrioides. J. Chem. Ecol. 2014, 40, 923–927. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Liu, N.; Zhu, J.; Zhang, T.; Dong, S. Characterization of two odorant binding proteins in Spodoptera exigua reveals functional conservation and difference. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2017, 213, 20–27. [Google Scholar] [CrossRef]
- Yu, Y.; Ma, F.; Cao, Y.; Zhang, J. Structural and functional difference of pheromone binding proteins in discriminating chemicals in the gypsy moth, Lymantria dispar. Int. J. Biol. Sci. 2012, 8, 979–991. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Q.; Zhang, S.; Luo, J.Y.; Zhu, J.; Cui, J.J.; Dong, S.L. Expression analysis and binding assays in the chemosensory protein gene family Indicate multiple roles in Helicoverpa armigera. J. Chem. Ecol. 2015, 41, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.J.; Robertson, G.; He, X.; Dufour, S.; Hooper, A.M.; Pickett, J.A.; Keep, N.H.; Field, L.M. Characterisation of Bombyx mori odorant-binding proteins reveals that a general odorant-binding protein discriminates between sex pheromone components. J. Mol. Biol. 2009, 389, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Marti-Renom, M.; Stuart, A.; Fiser, A.; Sanchez, R.; Melo, F.; Šali, A. Comparative protein structure modeling of genes and genomes. Annu. Rev. Biophys. Biomol. Struct. 2000, 29, 291–325. [Google Scholar] [CrossRef]
- Laskowski, R.; Rullmannn, J.; MacArthur, M.; Kaptein, R.; Thornton, J. Thornton AQUA and PROCHECK-NMR: Programs for checking the quality of protein structures solved by NMR. J. Biomol. NMR 1996, 8, 477–486. [Google Scholar] [CrossRef]
- Thompson, J.N.; Pellmyr, O. Evolution of oviposition behavior and host preference in lepidoptera. Annu. Rev. Entomol. 1991, 1, 65–89. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Wadhams, L.J.; Woodcock, C.M. Insect host location: A volatile situation. Trends Plant Sci. 2005, 10, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Najar-Rodriguez, A.J.; Galizia, C.G.; Stierle, J.; Dorn, S. Behavioral and neurophysiological responses of an insect to changing ratios of constituents in host plant-derived volatile mixtures. J. Exp. Biol. 2010, 213, 3388–3397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.L.; Zhang, J.X.; Yan, Z.G.; Yang, M.M.; Zhang, M.Z.; Zhang, Z.Y.; Qin, L.; Cao, Q. Host preference and performance of the yellow peach moth (Conogethes punctiferalis) on chestnut cultivars. PLoS ONE 2016, 11, e0157609. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K. Green leaf volatiles: Hydroperoxide lyase pathway of oxylipin metabolism. Curr. Opin. Plant Biol. 2006, 9, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Scala, A.; Allmann, S.; Mirabella, R.; Haring, M.A.; Schuurink, R.C. Green leaf volatiles: A plant’s multifunctional weapon against herbivores and pathogens. Int. J. Mol. Sci. 2013, 14, 17781–17811. [Google Scholar] [CrossRef] [PubMed]
- Gadenne, C.; Renou, M.; Sreng, L. Hormonal control of pheromone responsiveness in the male black cutwormAgrotis ipsilon. Experientia 1993, 49, 721–724. [Google Scholar] [CrossRef]
- TURGEON, J.J.; McNEIL, J.N.; ROELOFSt, W.L. Responsiveness of Pseudaletia unipuncta males to the female sex pheromone. Physiol. Entomol. 1983, 8, 339–344. [Google Scholar] [CrossRef]
- Pearson, G.A.; Schal, C. Electroantennogram responses of both sexes of grape root borer (Lepidoptera: Sesiidae) to synthetic female sex pheromone. Environ. Entomol. 1999, 28, 943–946. [Google Scholar] [CrossRef]
- Reddy, G.V.; Guerrero, A. Behavioral responses of the Diamondback moth, Plutella xylostella, to green leaf volatiles of Brassica oleracea Subsp. Capitata. J. Agric. Food Chem. 2000, 48, 6025–6029. [Google Scholar] [CrossRef]
- Sandler, B.H.; Nikonova, L.; Leal, W.S.; Clardy, J. Sexual attraction in the silkworm moth: Structure of the pheromone-binding-protein–bombykol complex. Chem. Biol. 2000, 7, 143–151. [Google Scholar] [CrossRef]
- Leal, W.S.; Nikonova, L.; Peng, G. Disulfide structure of the pheromone binding protein from the silkworm moth, Bombyx mori. FEBS Lett. 1999, 464, 85–90. [Google Scholar] [CrossRef]
- Scaloni, A.; Monti, M.; Angeli, S.; Pelosi, P. Structural analysis and disulfide-bridge pairing of two odorant-binding proteins from bombyx mori. Biochem. Biophys. Res. Commun. 1999, 266, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Liu, N.Y.; He, P.; Li, Z.Q.; Dong, S.L.; Mu, L.F. Molecular characterization, expression patterns, and ligand-binding properties of two odorant-binding protein genes from Orthaga achatina (butler) (lepidoptera: Pyralidae). Arch. Insect Biochem. Physiol. 2012, 80, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Free, J.; Ferguson, A.; Pickett, J. Evaluation of the various components of the Nasonov pheromone used by clustering honeybees. Physiol. Entomol. 1981, 6, 263–268. [Google Scholar] [CrossRef]
- Schiestl, F.; Ayasse, M. Post-mating odor in females of the solitary bee, Andrena nigroaenea (Apoidea, Andrenidae), inhibits male mating behavior. Behav. Ecol. Sociobiol. 2000, 48, 303–307. [Google Scholar] [CrossRef]
- Bergman, P.; Bergstrom, G. Scent marking, scent origin, and species specificity in male premating behavior of two Scandinavian bumblebees. Can. Field-Naturalist. 1997, 111, 1235–1251. [Google Scholar] [CrossRef]
- Merritt, T.J.; LaForest, S.; Prestwich, G.D.; Quattro, J.M.; Vogt, R.G. Patterns of gene duplication in lepidopteran pheromone binding proteins. J. Mol. Evol. 1998, 46, 272–276. [Google Scholar] [CrossRef]
- Vogt, R.G.; Rogers, M.E.; Franco, M.; Sun, M. A comparative study of odorant binding protein genes: Differential expression of the PBP1-GOBP2 gene cluster in Manduca sexta (Lepidoptera) and the organization of OBP genes in Drosophila melanogaster (Diptera). J. Exp. Biol. 2002, 205, 719–744. [Google Scholar] [CrossRef]
- Liu, N.Y.; Yang, K.; Liu, Y.; Xu, W.; Anderson, A.; Dong, S.L. Two general-odorant binding proteins in Spodoptera litura are differentially tuned to sex pheromones and plant odorants. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2015, 180, 23–31. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, Z.; Wang, H.; Kong, X. Molecular characterization, expression pattern, and ligand-binding property of three odorant binding protein genes from Dendrolimus tabulaeformis. J. Chem. Ecol. 2014, 40, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Chen, X.; Li, B.; Zhang, G.; Li, Y.; Wu, J. Binding properties of general odorant binding proteins from the oriental fruit moth, Grapholita molesta (Busck) (Lepidoptera: Tortricidae). PLoS ONE 2016, 11, e0155096. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | GOBP1 | GOBP2 | ||||
|---|---|---|---|---|---|---|
| Intensity (%) | IC50 (μM) | KD (μM) | Intensity (%) | IC50 (μM) | KD (μM) | |
| ALDEHYDES | ||||||
| Heptanal | 75.78 ± 5.17 | 13.52 ± 3.22 | 5.76 ± 0.64 | 96.80 ± 1.33 | 55.95 ± 1.54 | 23.12 ± 4.68 |
| Hexadecanal | 48.64 ± 4.35 | 1.29 ± 0.14 | 0.55 ± 0.08 | 67.94 ± 1.79 | 21.06 ± 1.83 | 8.70 ± 0.74 |
| Nonanal | 71.04 ± 8.80 | 13.35 ± 2.41 | 5.69 ± 0.36 | 96.99 ± 2.63 | 92.79 ± 3.83 | 38.34 ± 1.09 |
| Benzaldehyde | 83.58 ± 6.61 | 20.62 ± 1.31 | 8.79 ± 0.59 | 94.76 ± 7.32 | 77.12 ± 1.40 | 31.86 ± 3.03 |
| Undecanal | 78.33 ± 2.89 | 16.81 ± 2.41 | 7.17 ± 0.67 | 98.12 ± 1.92 | 96.67 ± 2.41 | 39.94 ± 9.70 |
| n-Hexanal | 45.50 ± 4.58 | 7.31 ± 1.43 | 3.11 ± 0.71 | 96.44 ± 1.50 | 68.07 ± 3.70 | 28.13 ± 0.62 |
| Z10-16:Ald | 20.88 ± 0.48 | 2.82 ± 0.44 | 1.20 ± 0.09 | 58.63 ± 2.72 | 8.65 ± 1.87 | 3.58 ± 0.62 |
| E10-16:Ald | 47.29 ± 7.52 | 4.90 ± 0.54 | 2.09 ± 0.45 | 84.05 ± 2.25 | 21.44 ± 4.15 | 8.86 ± 0.93 |
| ALCOHOLS | ||||||
| Linalool | 74.52 ± 2.42 | 15.71 ± 1.61 | 6.70 ± 0.69 | 91.86 ± 0.71 | 44.85 ± 1.16 | 18.53 ± 0.48 |
| Farnesol | 41.45 ± 2.51 | 0.83 ± 0.09 | 0.35 ± 0.04 | 55.64 ± 1.78 | 3.98 ± 0.36 | 1.64 ± 0.03 |
| Cis-3-hexen-1-ol | 94.99 ± 1.78 | 78.63 ± 0.23 | 33.53 ± 1.21 | 96.93 ± 0.99 | 51.33 ± 1.60 | 21.21 ± 1.14 |
| n-Hexanol | 97.23 ± 0.89 | 60.35 ± 2.44 | 25.73 ± 0.43 | 94.24 ± 1.82 | 95.19 ± 1.68 | 43.88 ± 2.41 |
| 1-Amyl alcohol | 95.2 ± 1.12 | 33.50 ± 1.02 | 14.28 ± 1.74 | 96.54 ± 1.39 | 98.67 ± 2.30 | 75.89 ± 1.39 |
| 1-Tetradecanol | 45.8 ± 0.56 | 4.81 ± 0.72 | 2.05 ± 0.43 | 85.52 ± 1.47 | 54.02 ± 2.56 | 22.32 ± 1.89 |
| OLEFINES | ||||||
| α-Pinene | 64.96 ± 3.26 | 11.10 ± 3.47 | 4.73 ± 0.48 | 98.57 ± 1.13 | 67.95 ± 4.66 | 28.07 ± 4.06 |
| β-Pinene | 66.21 ± 2.96 | 9.94 ± 2.72 | 4.24 ± 0.69 | 97.15 ± 3.95 | 33.70 ± 3.29 | 13.92 ± 1.36 |
| Caryophllene | 68.98 ± 3.16 | 9.09 ± 2.42 | 3.88 ± 0.91 | 89.93 ± 2.15 | 20.98 ± 2.10 | 8.67 ± 1.12 |
| Limonene | 43.27 ± 2.47 | 3.62 ± 0.89 | 1.54 ± 0.39 | 92.62 ± 1.21 | 35.47 ± 1.14 | 14.65 ± 2.78 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, D.; Zhang, T.; Bai, S.; Prabu, S.; He, K.; Dewer, Y.; Wang, Z. GOBP1 Plays a Key Role in Sex Pheromones and Plant Volatiles Recognition in Yellow Peach Moth, Conogethes punctiferalis (Lepidoptera: Crambidae). Insects 2019, 10, 302. https://doi.org/10.3390/insects10090302
Jing D, Zhang T, Bai S, Prabu S, He K, Dewer Y, Wang Z. GOBP1 Plays a Key Role in Sex Pheromones and Plant Volatiles Recognition in Yellow Peach Moth, Conogethes punctiferalis (Lepidoptera: Crambidae). Insects. 2019; 10(9):302. https://doi.org/10.3390/insects10090302
Chicago/Turabian StyleJing, Dapeng, Tiantao Zhang, Shuxiong Bai, Sivaprasath Prabu, Kanglai He, Youssef Dewer, and Zhenying Wang. 2019. "GOBP1 Plays a Key Role in Sex Pheromones and Plant Volatiles Recognition in Yellow Peach Moth, Conogethes punctiferalis (Lepidoptera: Crambidae)" Insects 10, no. 9: 302. https://doi.org/10.3390/insects10090302