The ABC Transporter Components HgdB and HgdC are Important for Glycolipid Layer Composition and Function of Heterocysts in Anabaena sp. PCC 7120


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
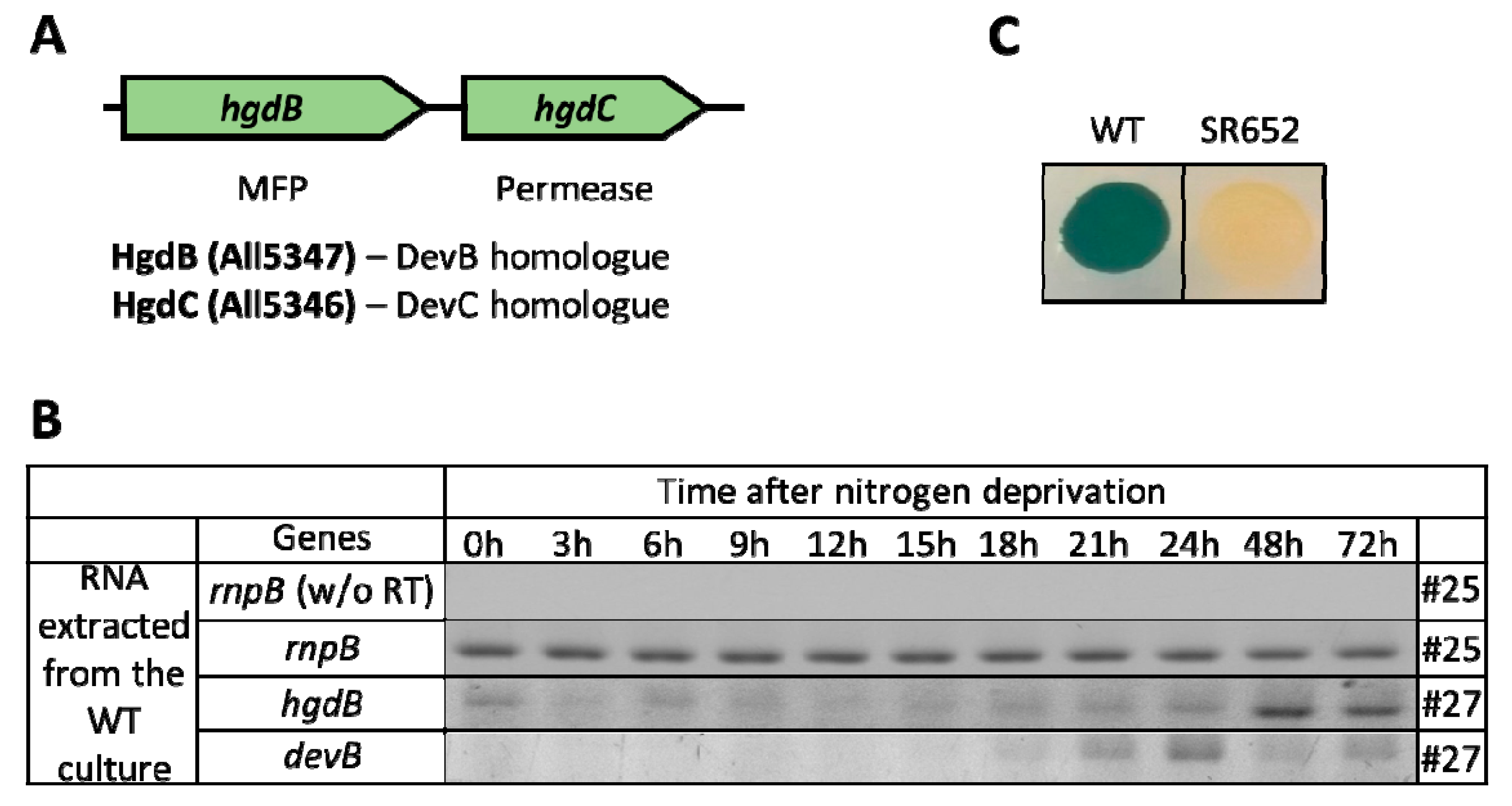
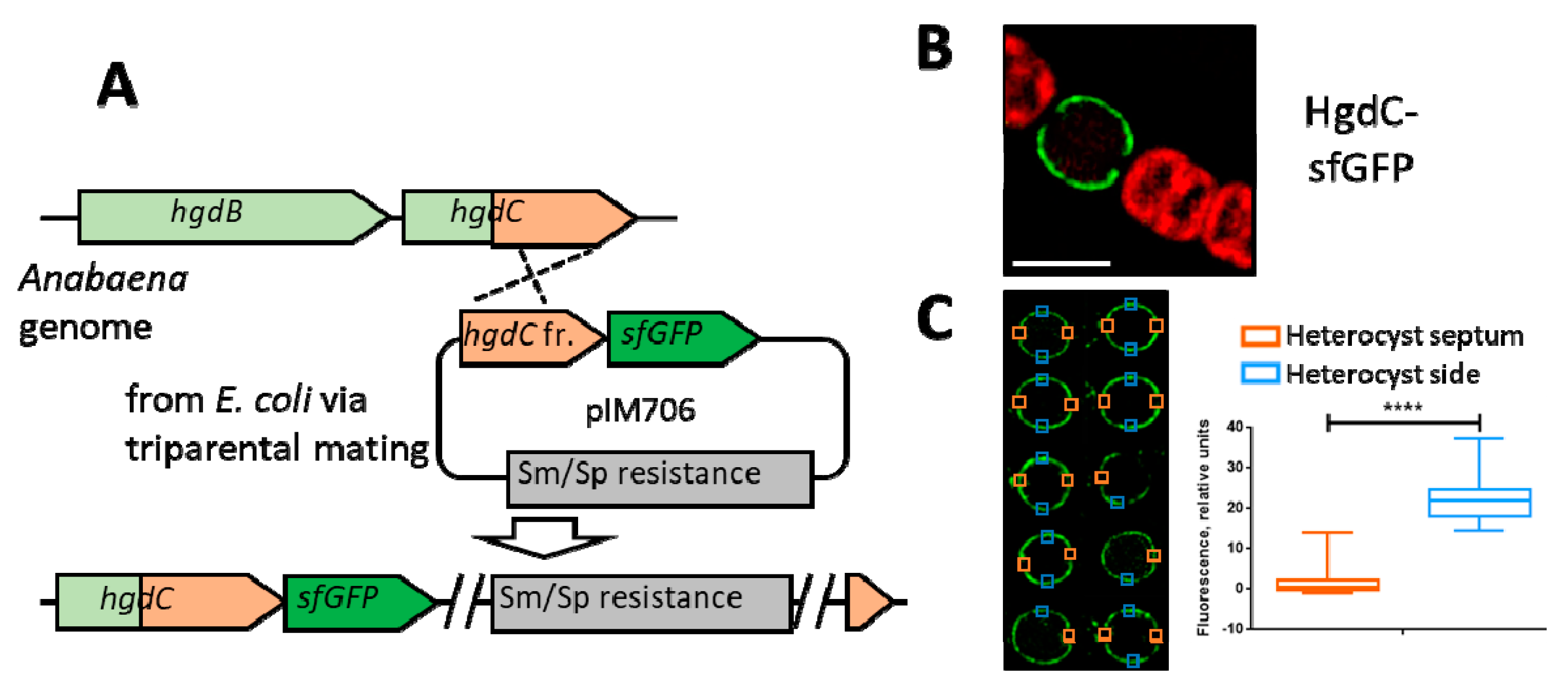
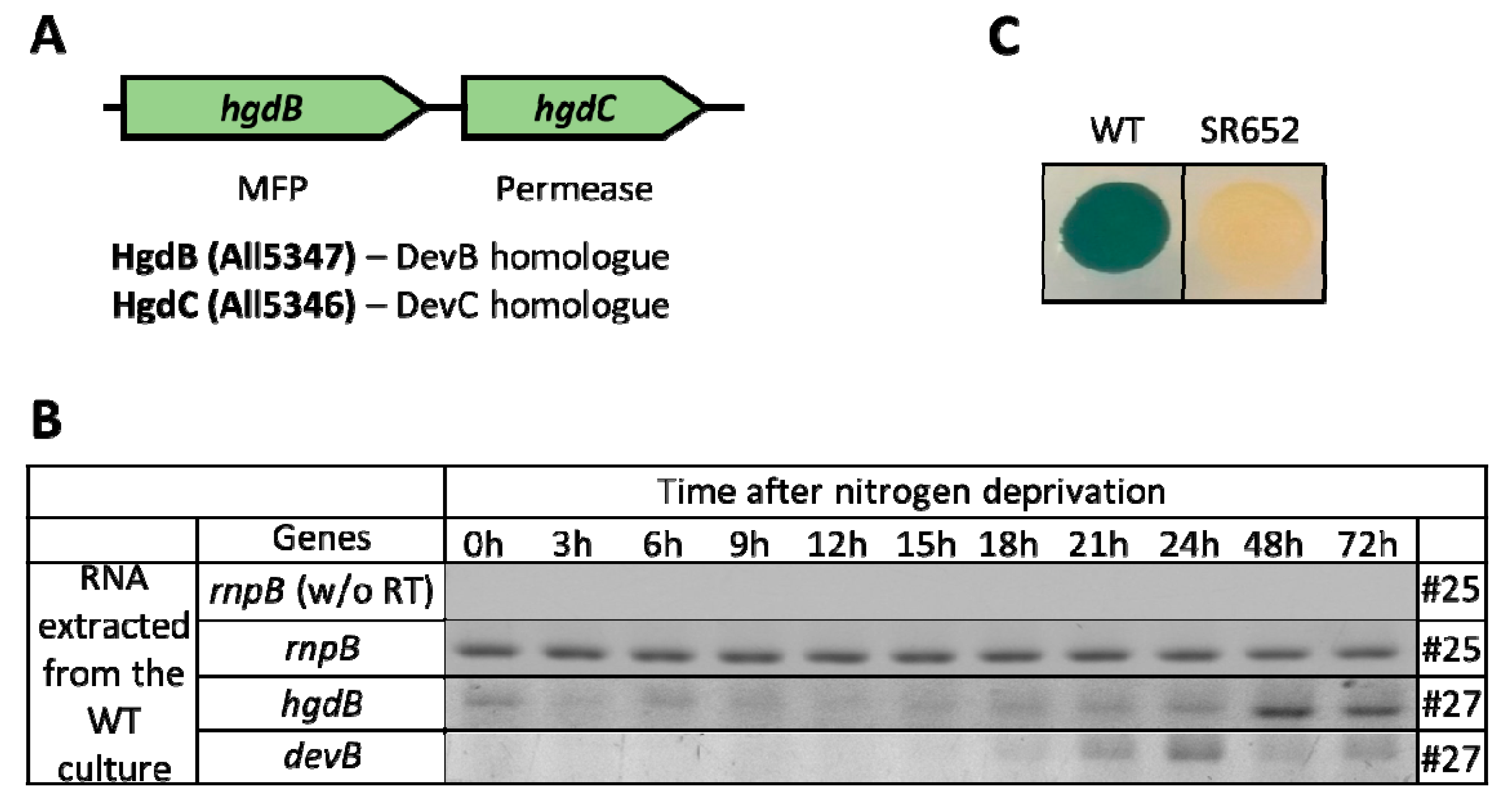
3.1. HgdBC Is Differentially Expressed in Heterocysts and Localizes at the Membrane
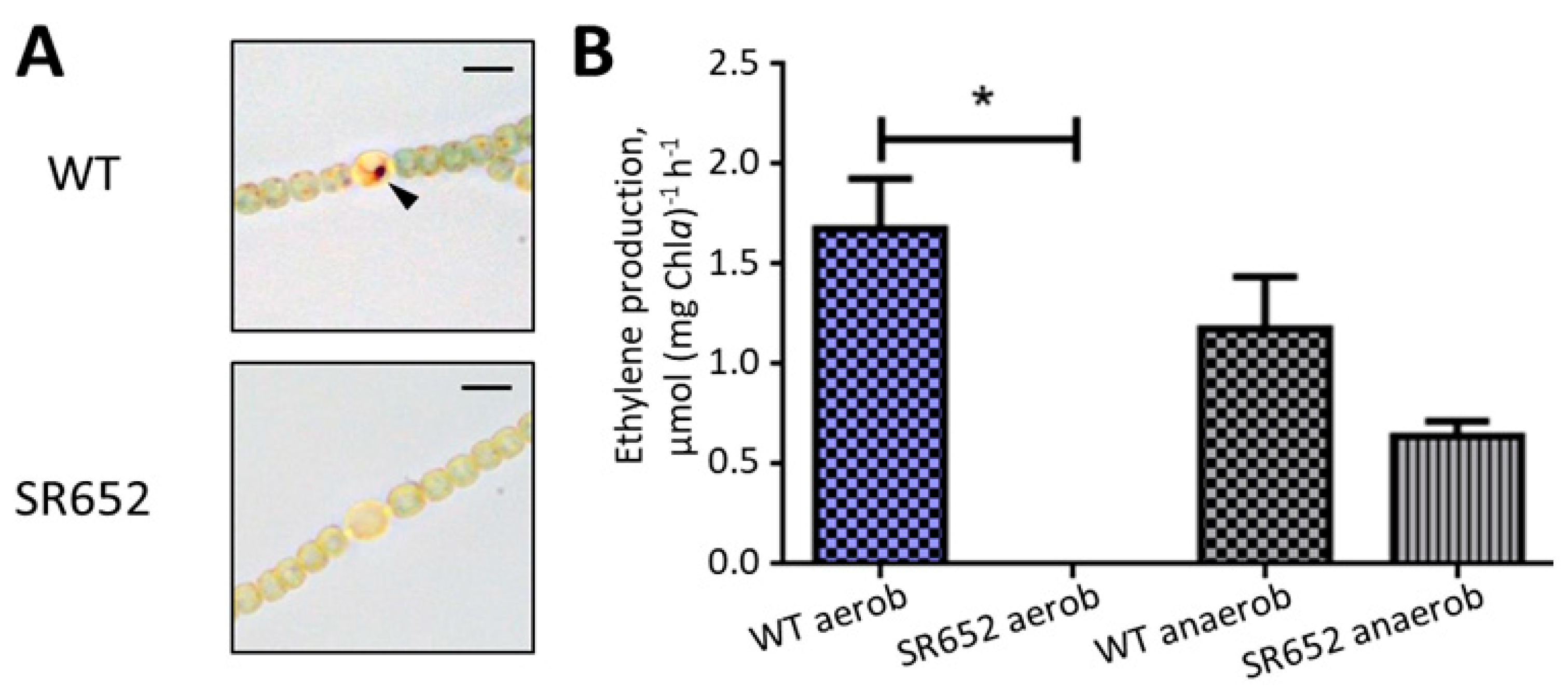
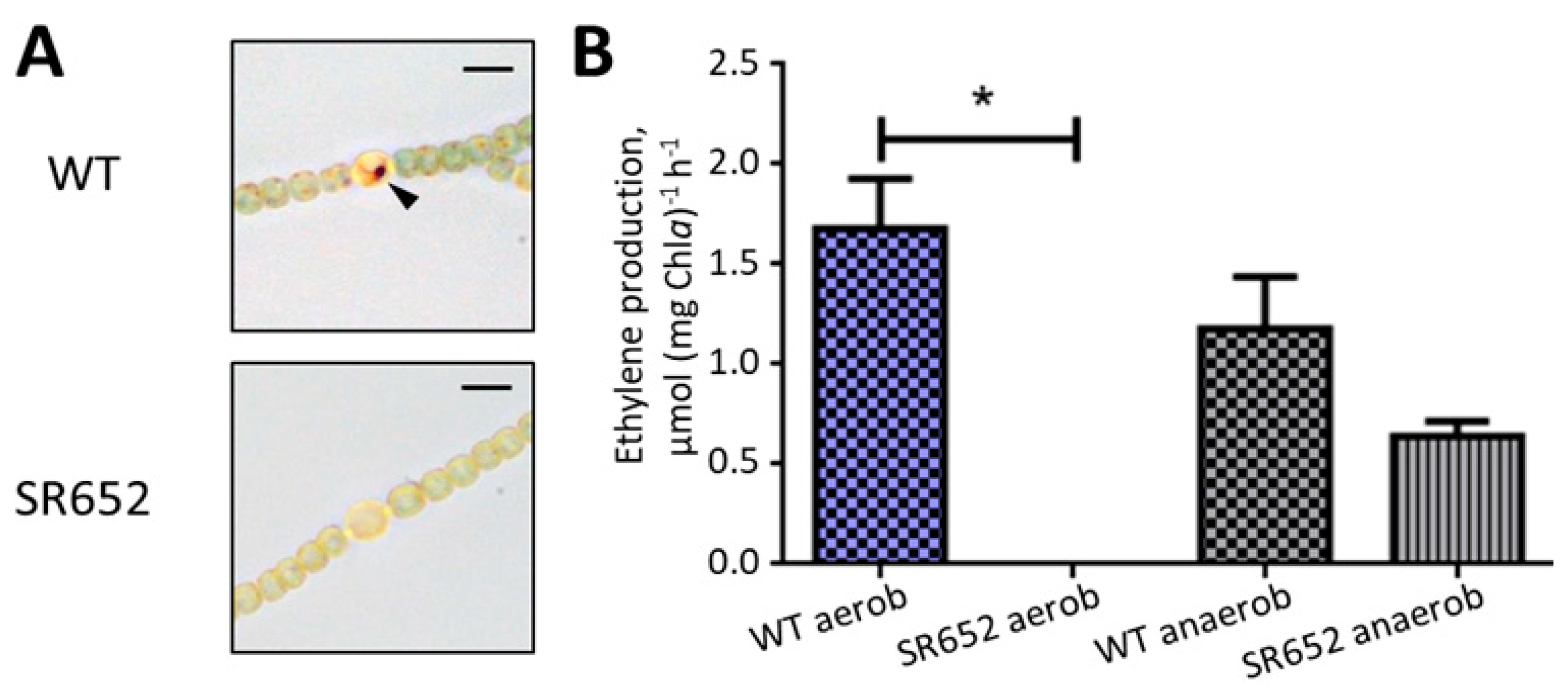
3.2. The HgdB Mutant Is Impaired in Diazotrophic Growth
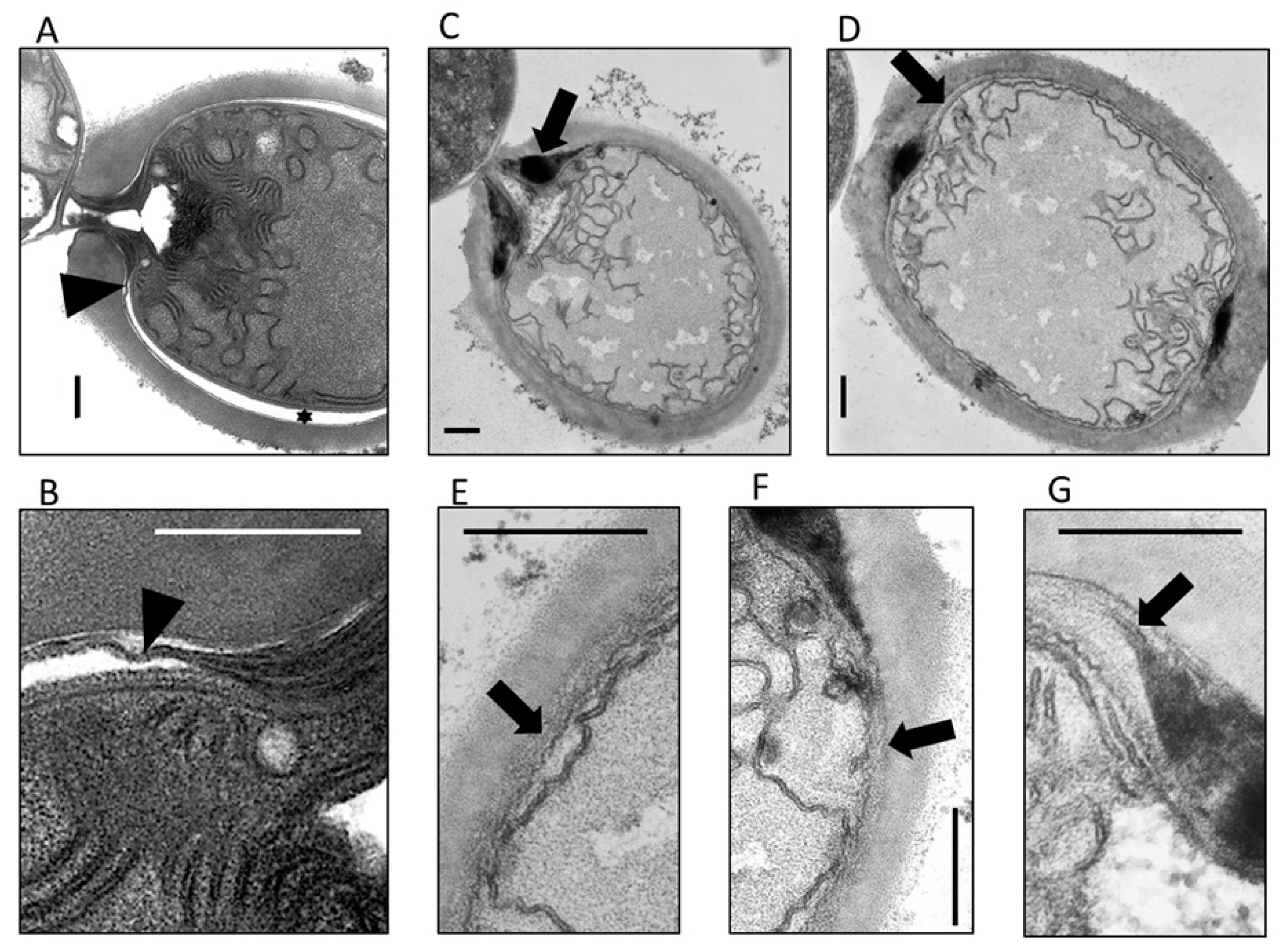
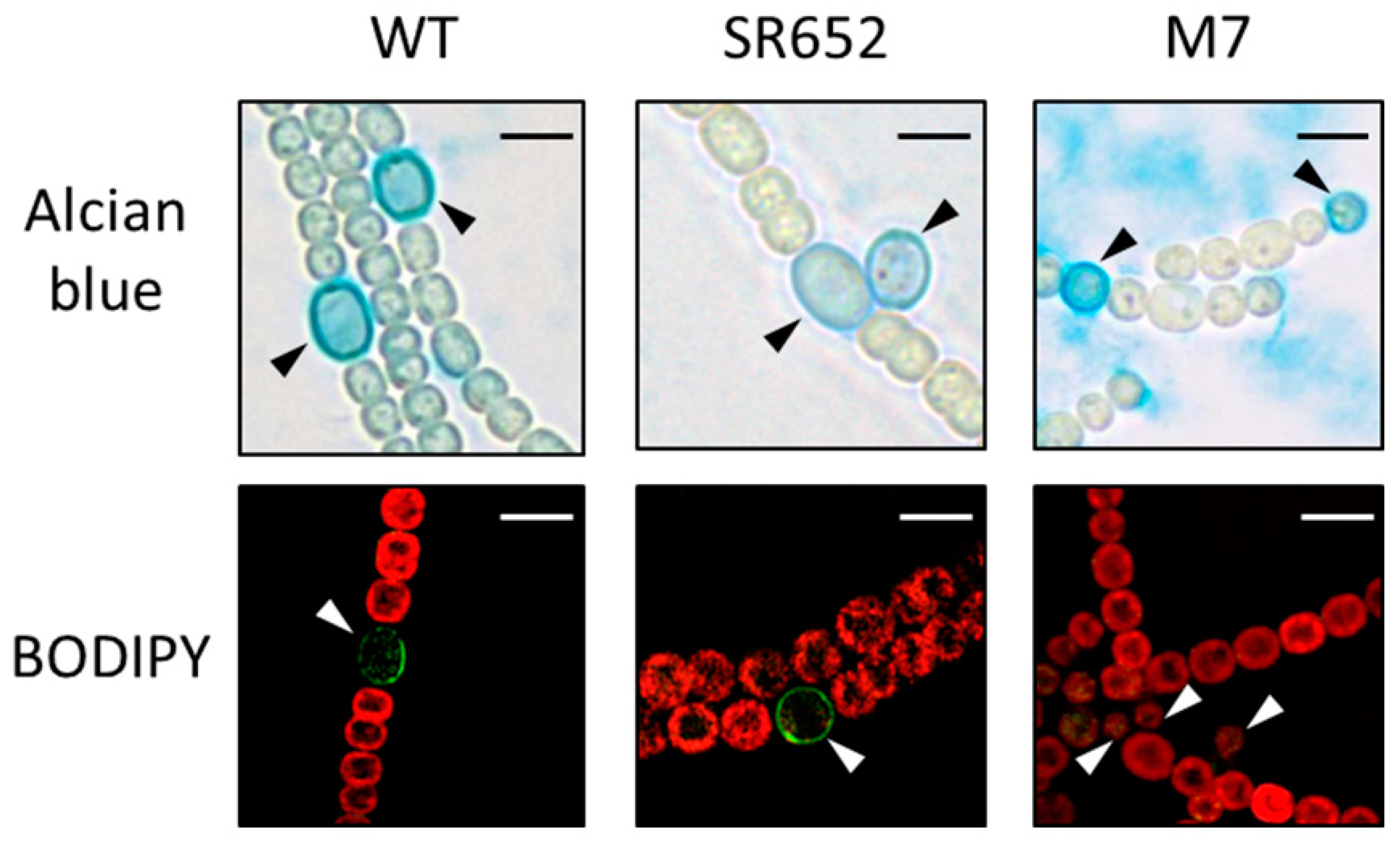
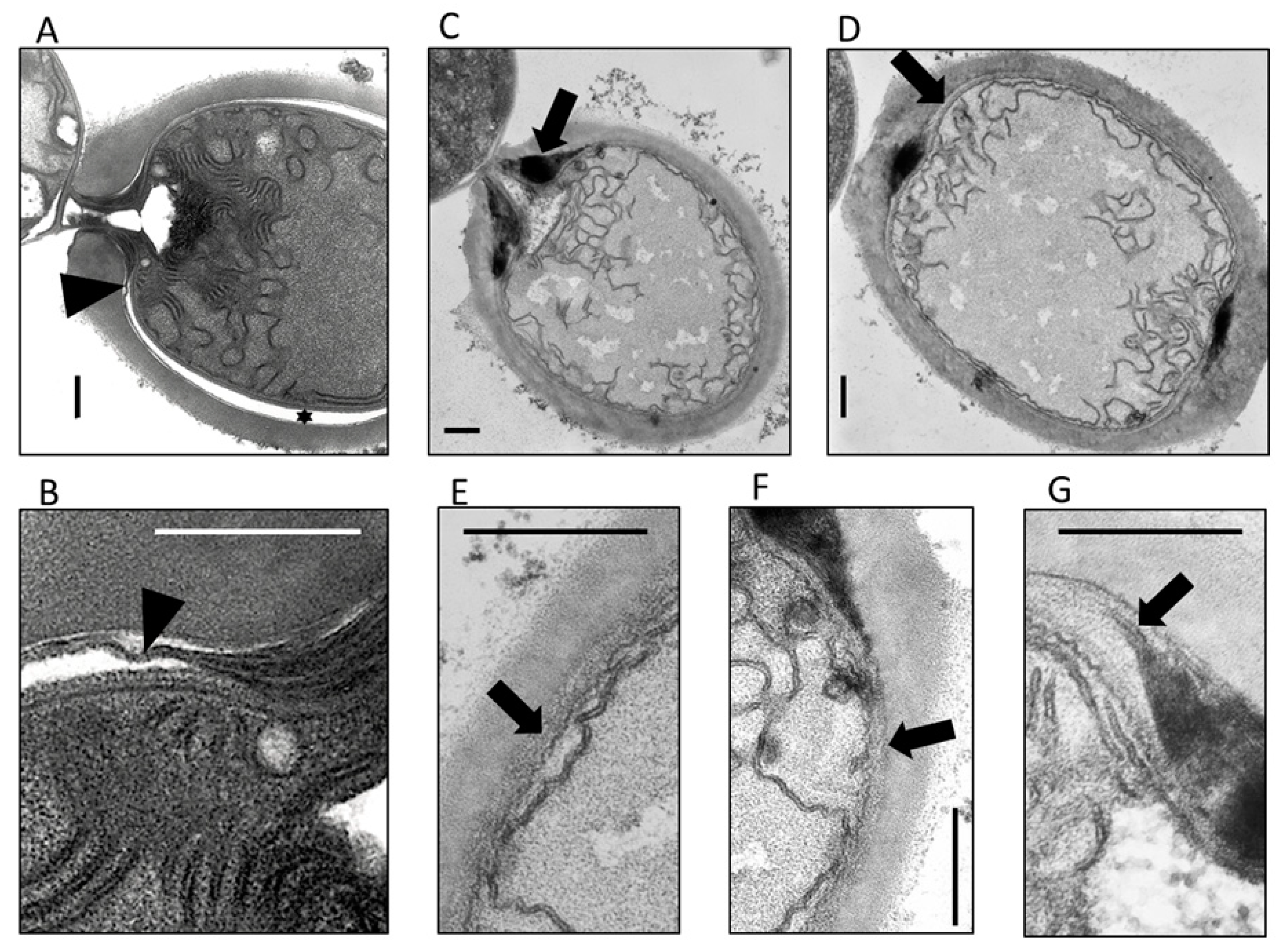
3.3. The HgdB-Mutant Forms an Aberrant Heterocyst Cell Envelope
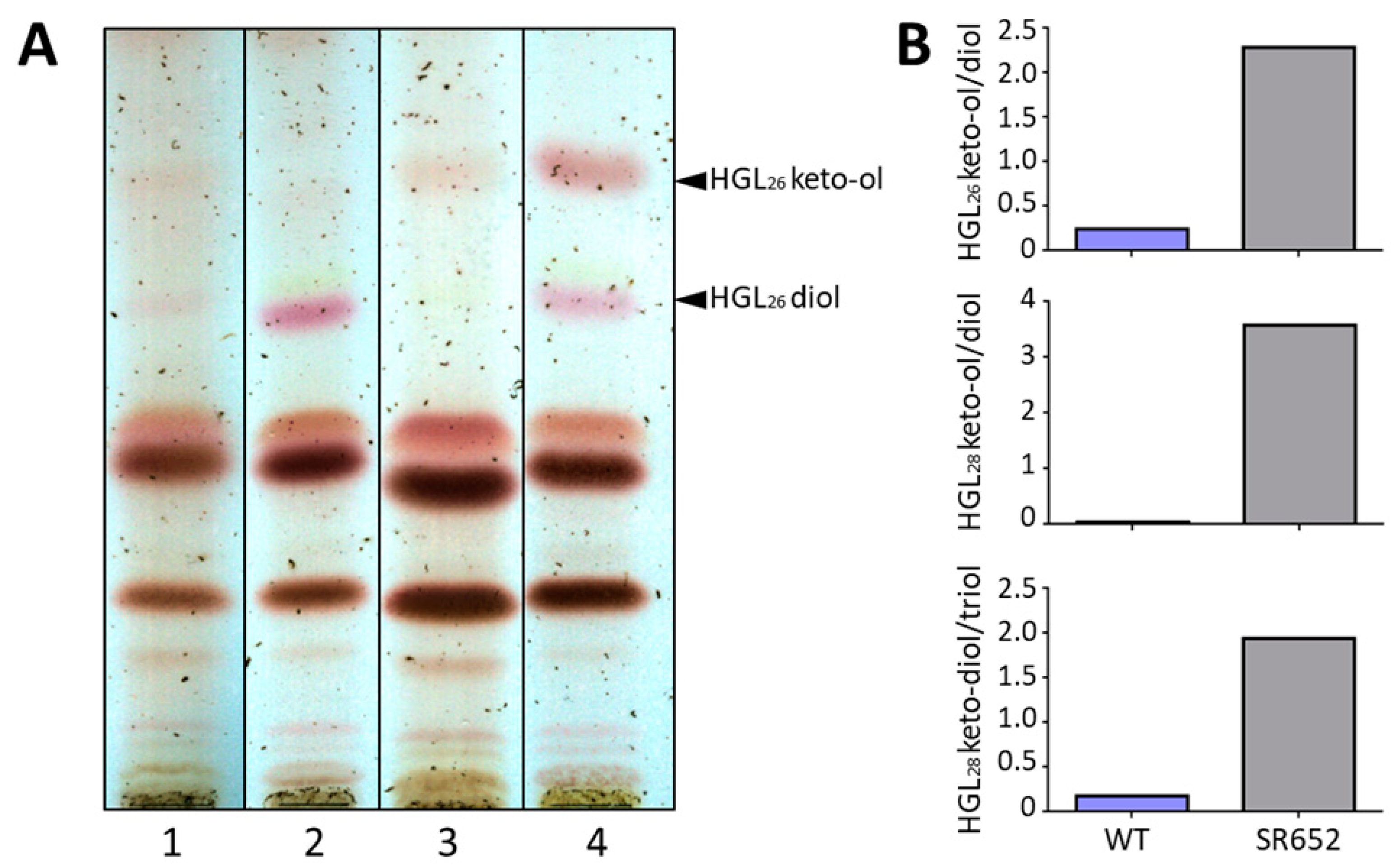
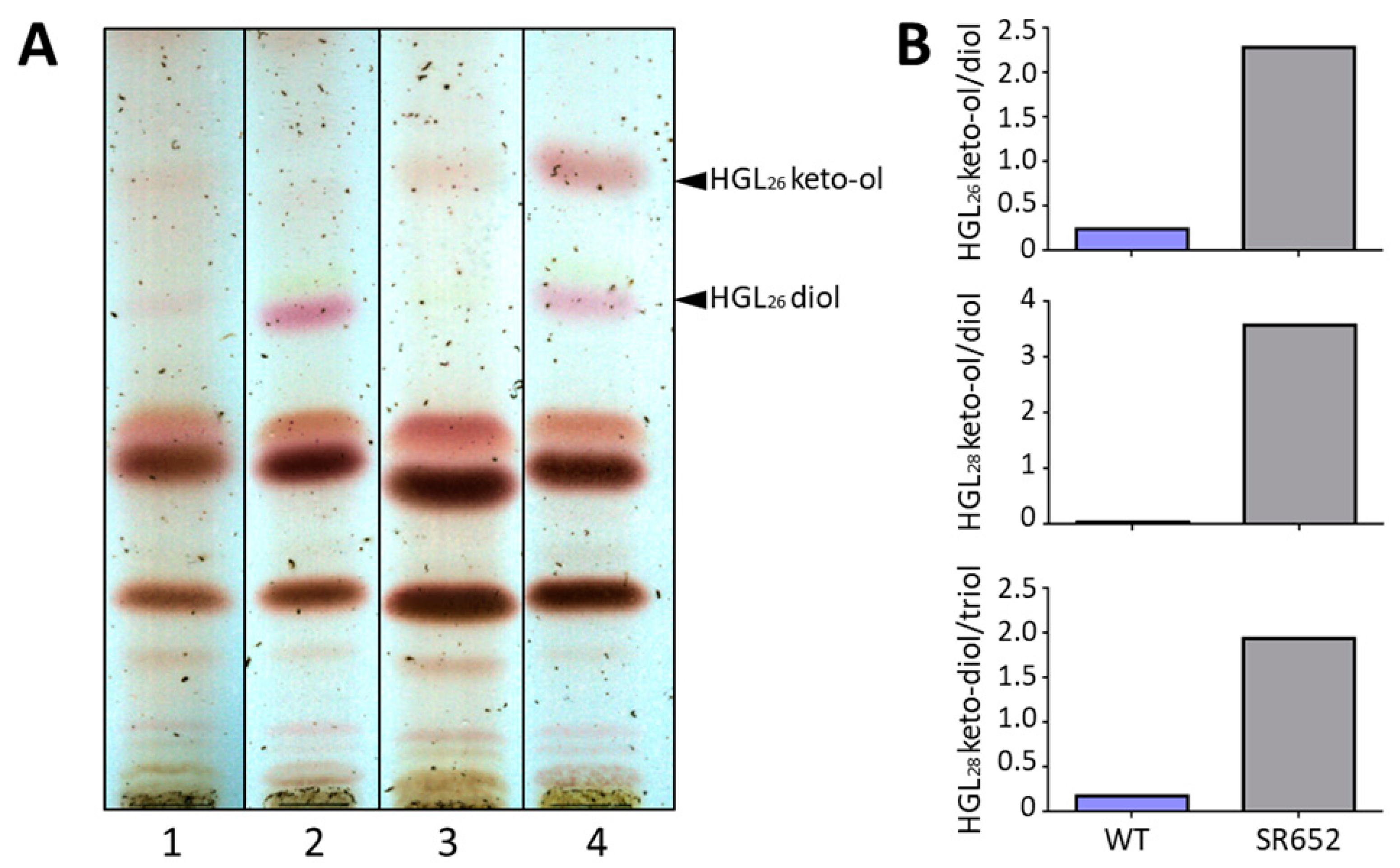
3.4. The Glycolipid Composition Is Affected in the HgdB-Mutant
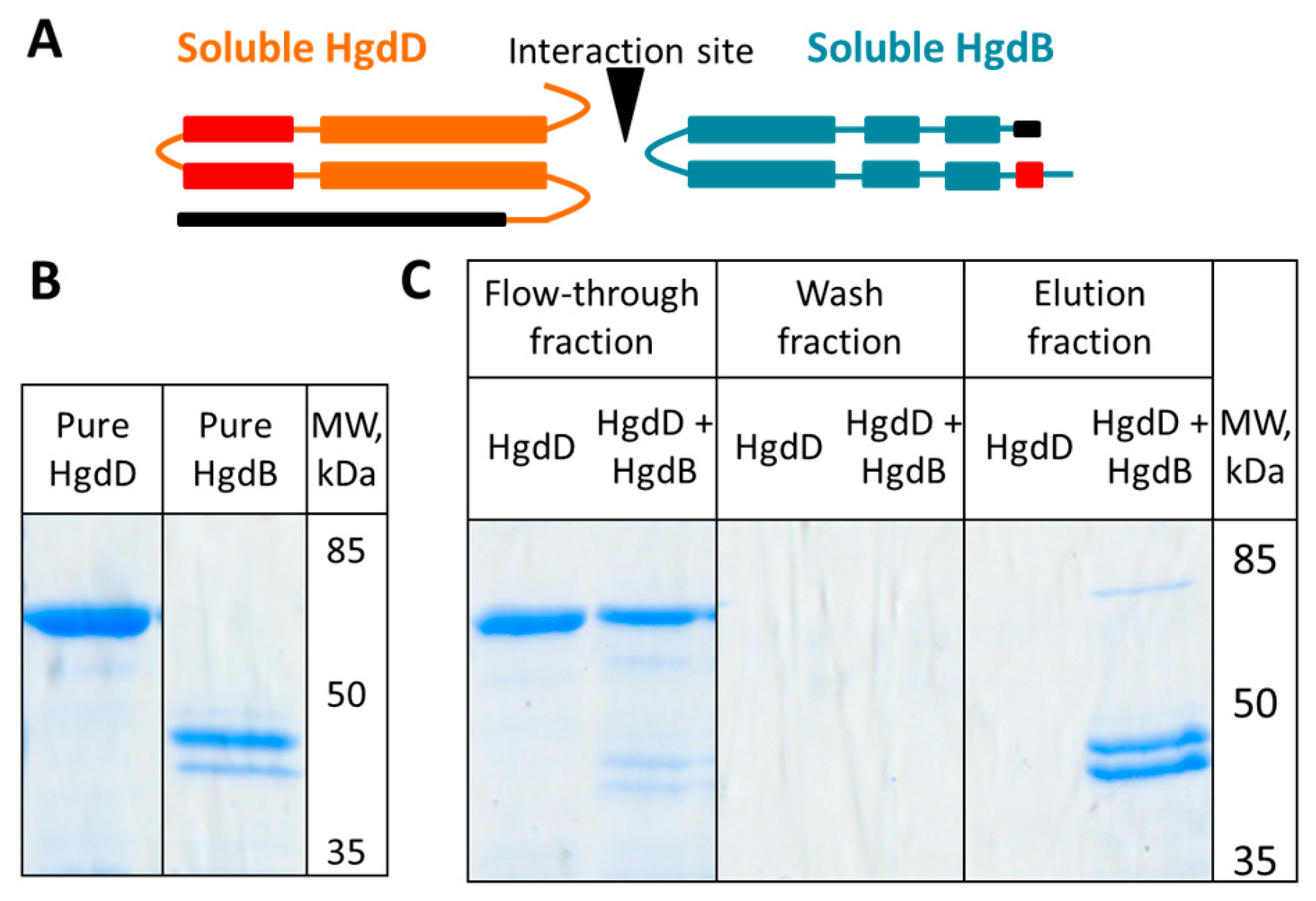
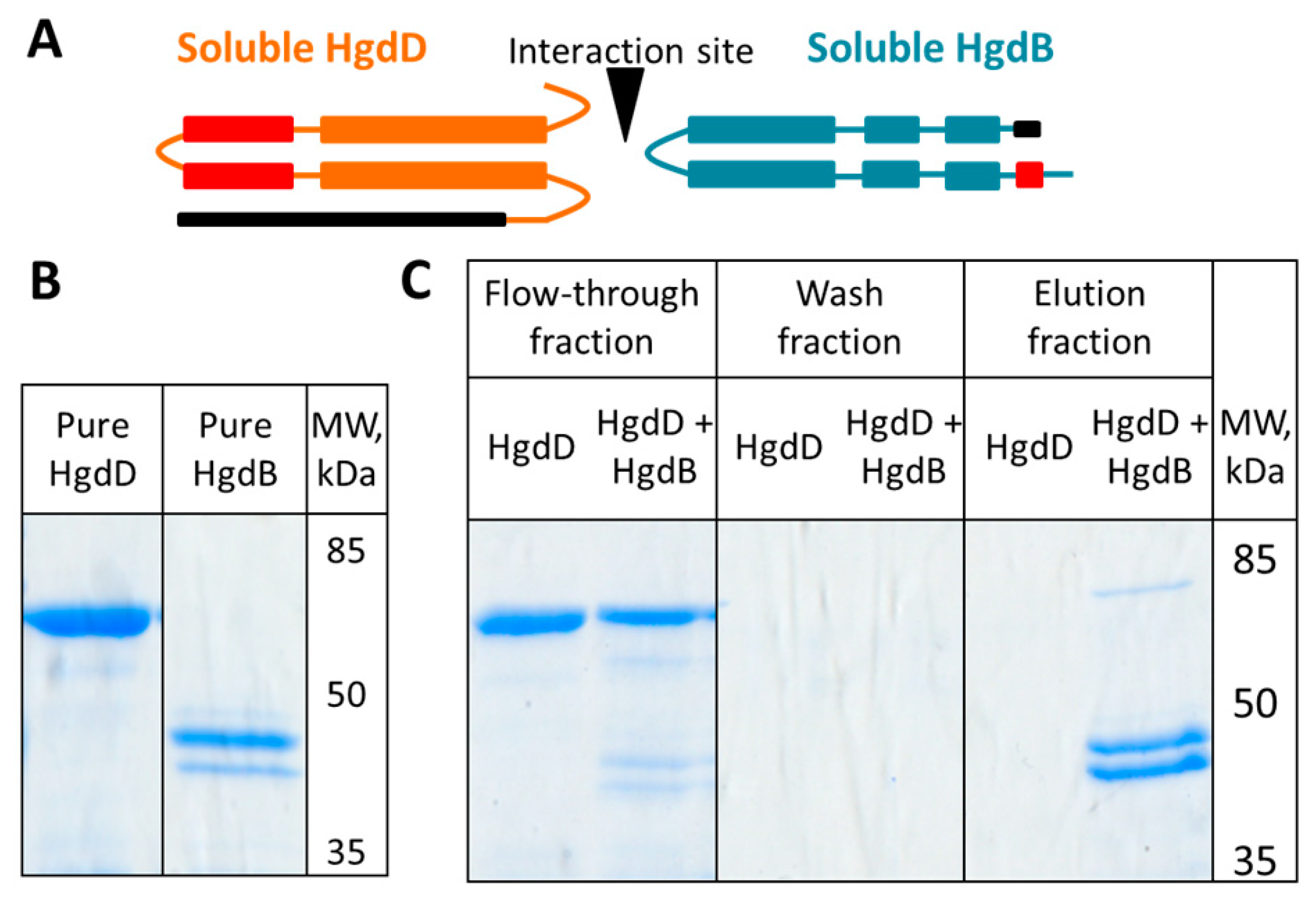
3.5. HgdB Interacts with HgdD in Vitro
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adams, D.G.; Duggan, P.S. Tansley Review No. 107 Heterocyst and akinete di ff erentiation in cyanobacteria. New Phytol. 1999, 144, 3–33. [Google Scholar] [CrossRef]
- Maldener, I.; Summers, M.L.; Sukenik, A. Cellular differentiation in filamentous cyanobacteria. In The Cell Biology of Cyanobacteria; Flores, E., Herrero, A., Eds.; Caister Academic Press: Norfolk, UK, 2014; pp. 263–291. ISBN 978-1-908230-38-6. [Google Scholar]
- Muro-Pastor, A.M.; Hess, W.R. Heterocyst differentiation: From single mutants to global approaches. Trends Microbiol. 2012, 20, 548–557. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Mella-Herrera, R.A.; Golden, J.W. Cyanobacterial heterocysts. Cold Spring Harb. Perspect. Biol. 2010, 2, a000315. [Google Scholar] [CrossRef] [PubMed]
- Wörmer, L.; Cires, S.; Velazquez, D.; Quesada, A.; Hinrichs, K.-U. Cyanobacterial heterocyst glycolipids in cultures and environmental samples: Diversity and biomarker potential. Limnol. Oceanogr. 2012, 57, 1775–1788. [Google Scholar] [CrossRef]
- Bauersachs, T.; Compaoré, J.; Hopmans, E.C.; Stal, L.J.; Schouten, S.; Sinninghe, J.S. Distribution of heterocyst glycolipids in cyanobacteria. Phytochemistry 2009, 70, 2034–2039. [Google Scholar] [CrossRef] [PubMed]
- Gambacorta, A.; Pagnotta, E.; Romano, I.; Sodano, G.; Trincone, A. Heterocyst Glycilipids From Nitrogen-Fixing Cyanobacteria Other Than Nostocaceae. Phytochemistry 1998, 48, 801–805. [Google Scholar] [CrossRef]
- Gambacorta, A.; Trincone, A.; Soriente, A.; Sodano, G. Chemistry of glycolipids from the heterocysts of nitrogen-fixing cyanobacteria. Curr. Top. Phytochem. 1999, 2, 145–150. [Google Scholar]
- Soriente, A.; Gambacorta, A.; Trincone, A.; Sili, C.; Vincenzini, M.; Sodano, G. Heterocyst glycolipids of the cyanobacterium Cyanospira rippkae. Phytochemistry 1993, 33, 393–396. [Google Scholar] [CrossRef]
- Bale, N.J.; Hopmans, E.C.; Zell, C.; Sobrinho, R.L.; Kim, J.-H.; Damste, J.S.S.; Villareal, T.A.; Schouten, S. Long chain glycolipids with pentose head groups as biomarkers for marine endosymbiotic heterocystous cyanobacteria. Org. Geochem. 2015, 81, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Gambacorta, A.; Soriente, A.; Trincone, A.; Sodano, G. Biosynthesis of the heterocyst glycolipids in the cyanobacterium Anabaena Cylindrica. Phytochemistry 1995, 39, 771–774. [Google Scholar] [CrossRef]
- Schouten, S.; Villareal, T.A.; Hopmans, E.C.; Mets, A.; Swanson, K.M.; Damste, J.S.S. Endosymbiotic heterocystous cyanobacteria synthesize different heterocyst glycolipids than free-living heterocystous cyanobacteria. Phytochemistry 2013, 85, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Delepelaire, P. Type I secretion in gram-negative bacteria. Biochim. Biophys. Acta Mol. Cell Res. 2004, 1694, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Zgurskaya, H.I. Multicomponent drug efflux complexes: Architecture and mechanism of assembly. Future Microbiol. 2009, 4, 919–932. [Google Scholar] [CrossRef] [PubMed]
- Shvarev, D.; Maldener, I. ATP-binding cassette transporters of the multicellular cyanobacterium Anabaena sp. PCC 7120: A wide variety for a complex lifestyle. FEMS Microbiol. Lett. 2018, 365, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Staron, P.; Forchhammer, K.; Maldener, I. Novel ATP-driven pathway of glycolipid export involving TolC protein. J. Biol. Chem. 2011, 286, 38202–38210. [Google Scholar] [CrossRef] [PubMed]
- Staron, P.; Forchhammer, K.; Maldener, I. Structure-function analysis of the ATP-driven glycolipid efflux pump DevBCA reveals complex organization with TolC/HgdD. FEBS Lett. 2014, 588, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Moslavac, S.; Nicolaisen, K.; Mirus, O.; Al Dehni, F.; Pernil, R.; Flores, E.; Maldener, I.; Schleiff, E. A TolC-like protein is required for heterocyst development in Anabaena sp. strain PCC 7120. J. Bacteriol. 2007, 189, 7887–7895. [Google Scholar] [CrossRef] [PubMed]
- Moslavac, S.; Bredemeier, R.; Mirus, O.; Granvogl, B.; Eichacker, L.A.; Schleiff, E. Proteomic Analysis of the Outer Membrane of Anabaena sp. Strain PCC 7120. J. Proteome Res. 2005, 4, 1330–1338. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, G.; Arnold, M.; Hannus, S.; Maldener, I. The DevBCA exporter is essential for envelope formation in heterocysts of the cyanobacterium Anabaena sp. strain PCC 7120. Mol. Microbiol. 1998, 27, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Maldener, I.; Fiedler, G.; Ernst, A.; Fernandez-piias, F.; Wolkl, C.P. Characterization of devA, a Gene Required for the Maturation of Proheterocysts in the Cyanobacterium Anabaena sp. Strain PCC 7120. J. Bacteriol. 1994, 176, 7543–7549. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.; Huang, G.; Lechno-Yossef, S.; Wolk, C.P.; Kaneko, T.; Tabata, S. Clustered genes required for synthesis and deposition of envelope glycolipids in Anabaena sp. strain PCC 7120. Mol. Microbiol. 2005, 58, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Fan, Q.; Lechno-Yossef, S.; Wojciuch, E.; Wolk, C.P.; Kaneko, T.; Tabata, S. Clustered Genes Required for the Synthesis of Heterocyst Envelope Polysaccharide in Anabaena sp. Strain PCC 7120. J. Bacteriol. 2005, 187, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Staron, P.; Maldener, I. All0809/8/7 is a DevBCA-like ABC-type efflux pump required for diazotrophic growth in Anabaena sp. PCC 7120. Microbiology 2012, 158, 2537–2545. [Google Scholar] [CrossRef] [PubMed]
- Staron, P. Structural and Functional Characterization of the ATP-Driven Glycolipid-Efflux Pump DevBCA-TolC and Its Homologues in the Filamentous Cyanobacterium Anabaena sp. PCC 7120; Eberhard-Karls Universität Tübingen: Tübingen, Germany, 2012. [Google Scholar]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Wolk, C.P.; Vonshak, A.; Kehoe, P.; Elhai, J. Construction of shuttle vectors capable of conjugative transfer from Escherichia coli to nitrogen-fixing filamentous cyanobacteria. Proc. Natl. Acad. Sci. USA 1984, 81, 1561–1565. [Google Scholar] [CrossRef] [PubMed]
- Elhai, J.; Wolk, C.P. Conjugal Transfer of DNA to Cyanobacteria. Methods Enzymol. 1988, 167, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.G.; Young, L.; Chuang, R.-Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Pédelacq, J.-D.; Cabantous, S.; Tran, T.; Terwilliger, T.C.; Waldo, G.S. Engineering and characterization of a superfolder green fluorescent protein. Nat. Biotechnol. 2006, 24, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Perez, R.; Wörmer, L.; Sass, P.; Maldener, I. A highly asynchronous developmental program triggered during germination of dormant akinetes of filamentous diazotrophic cyanobacteria. FEMS Microbiol. Ecol. 2018, 94, fix131. [Google Scholar] [CrossRef] [PubMed]
- Lehner, J.; Berendt, S.; Dörsam, B.; Pérez, R.; Forchhammer, K.; Maldener, I. Prokaryotic multicellularity: A nanopore array for bacterial cell communication. FASEB J. 2013, 27, 2293–2300. [Google Scholar] [CrossRef] [PubMed]
- McKinney, R.E. Staining bacterial polysaccharides. J. Bacteriol. 1953, 66, 453–454. [Google Scholar] [PubMed]
- Fay, P.; Kulasooriya, S.A. Tetrazolium reduction and nitrogenase activity in heterocystous blue-green algae. Arch. Mikrobiol. 1972, 87, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Winkenbach, F.; Wolk, C.P.; Jost, M. Lipids of membranes and of the cell envelope in heterocysts of a blue-green alga. Planta 1972, 107, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Mackinney, G. Absorption of Light by Chlorophyll Solutions. J. Biol. Chem. 1941, 140, 315–322. [Google Scholar]
- Wörmer, L.; Lipp, J.S.; Schröder, J.M.; Hinrichs, K.U. Application of two new LC-ESI-MS methods for improved detection of intact polar lipids (IPLs) in environmental samples. Org. Geochem. 2013, 59, 10–21. [Google Scholar] [CrossRef]
- Bauersachs, T.; Hopmans, E.C.; Compaore, J.; Stal, L.J.; Schouten, S.; Damste, J.S.S. Rapid analysis of long-chain glycolipids in heterocystous cyanobacteria using high-performance liquid chromatography coupled to electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2009, 23, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Bornikoel, J.; Staiger, J.; Madlung, J.; Forchhammer, K.; Maldener, I. LytM factor Alr3353 affects filament morphology and cell–cell communication in the multicellular cyanobacterium Anabaena sp. PCC 7120. Mol. Microbiol. 2018, 108, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, C.F.; Pacheco, C.C.; Tamagnini, P.; Oliveira, P. Identification of inner membrane translocase components of TolC-mediated secretion in the cyanobacterium Synechocystis sp. PCC 6803. Environ. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Luque, I.; Flores, E.; Herrero, A. Molecular mechanism for the operation of nitrogen control in cyanobacteria. EMBO J. 1994, 13, 2862–2869. [Google Scholar] [PubMed]
- Vázquez-Bermúdez, M.F.; Flores, E.; Herrero, A. Analysis of binding sites for the nitrogen-control transcription factor NtcA in the promoters of Synechococcus nitrogen-regulated genes. Biochim. Biophys. Acta Gene Struct. Expr. 2002, 1578, 95–98. [Google Scholar] [CrossRef]
- Fiedler, G.; Muro-Pastor, A.M.; Flores, E.; Maldener, I. NtcA-dependent expression of the devBCA operon, encoding a heterocyst-specific ATP-binding cassette transporter in Anabaena spp. J. Bacteriol. 2001, 183, 3795–3799. [Google Scholar] [CrossRef] [PubMed]
- Ernst, A.; Black, T.; Cai, Y.; Panoff, J.; Tiwari, D.N.; Wolk, C.P. Synthesis of Nitrogenase in Mutants of the Cyanobacterium Anabaena sp. Strain PCC 7120 Affected in Heterocyst Development or Metabolism. J. Bacteriol. 1992, 174, 6025–6032. [Google Scholar] [CrossRef] [PubMed]
- Hahn, A.; Stevanovic, M.; Mirus, O.; Schleiff, E. The TolC-like protein HgdD of the cyanobacterium Anabaena sp. PCC 7120 is involved in secondary metabolite export and antibiotic resistance. J. Biol. Chem. 2012, 287, 41126–41138. [Google Scholar] [CrossRef] [PubMed]
- Awai, K.; Lechno-Yossef, S.; Wolk, C.P. Heterocyst Envelope Glycolipids. In Lipids in Photosynthesis: Essential and Regulatory Functions; Wada, H., Murata, N., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 179–202. ISBN 978-90-481-2863-1. [Google Scholar]
- Jones, P.M.; O’Mara, M.L.; George, A.M. ABC transporters: A riddle wrapped in a mystery inside an enigma. Trends Biochem. Sci. 2009, 34, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Flores, E.; Herrero, A.; Wolk, C.P.; Maldener, I. Is the periplasm continuous in filamentous multicellular cyanobacteria? Trends Microbiol. 2006, 14, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-Y.; Lin, G.-M.; Xing, W.-Y.; Zhang, C.-C. Diversity of Growth Patterns Probed in Live Cyanobacterial Cells Using a Fluorescent Analog of a Peptidoglycan Precursor. Front. Microbiol. 2018, 9, 791. [Google Scholar] [CrossRef] [PubMed]
- Walsby, A.E. The permeability of heterocysts to the gases nitrogen and oxygen. Proc. R. Soc. B Biol. Sci. 1985, 226, 345–366. [Google Scholar] [CrossRef]
- Soriente, A.; Sodano, G.; Cambacorta, A.; Trincone, A. Structure of the “heterocyst glycolipids” of the marine cyanobacterium Nodularia harveyana. Tetrahedron 1992, 48, 5375–5384. [Google Scholar] [CrossRef]
- Bauersachs, T.; Stal, L.J.; Grego, M.; Schwark, L. Temperature induced changes in the heterocyst glycolipid composition of N2 fixing heterocystous cyanobacteria. Org. Geochem. 2014, 69, 98–105. [Google Scholar] [CrossRef]
- Valladares, A.; Maldener, I.; Muro-Pastor, A.M.; Flores, E.; Herrero, A. Heterocyst development and diazotrophic metabolism in terminal respiratory oxidase mutants of the cyanobacterium Anabaena sp. strain PCC 7120. J. Bacteriol. 2007, 189, 4425–4430. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Wang, L.; Jeanjean, R.; Zhang, C.C. PrpJ, a PP2C-type protein phosphatase located on the plasma membrane, is involved in heterocyst maturation in the cyanobacterium Anabaena sp. PCC 7120. Mol. Microbiol. 2007, 64, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Li, J.H.; Cheng, Y.; Wang, L.; Chen, W.L.; Zhang, C.C. Two genes encoding protein kinases of the HstK family are involved in synthesis of the minor heterocyst-specific glycolipid in the cyanobacterium Anabaena sp. strain PCC 7120. J. Bacteriol. 2007, 189, 5075–5081. [Google Scholar] [CrossRef] [PubMed]
- Awai, K.; Wolk, C.P. Identifcation of the glycosyl transferase required for synthesis of the principal glycolipid characteristic of heterocysts of Anabaena sp. strain PCC 7120. FEMS Microbiol. Lett. 2007, 266, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Stevanovic, M.; Hahn, A.; Nicolaisen, K.; Mirus, O.; Schleiff, E. The components of the putative iron transport system in the cyanobacterium Anabaena sp. PCC 7120. Environ. Microbiol. 2012, 14, 1655–1670. [Google Scholar] [CrossRef] [PubMed]
- Hahn, A.; Stevanovic, M.; Mirus, O.; Lytvynenko, I.; Pos, K.M.; Schleiff, E. The outer membrane TolC-like channel HgdD is part of tripartite resistance-nodulation-cell division (RND) efflux systems conferring multiple-drug resistance in the cyanobacterium Anabaena sp. PCC7120. J. Biol. Chem. 2013, 288, 31192–31205. [Google Scholar] [CrossRef] [PubMed]
- Flaherty, B.; Nieuwerburgh, F.; Head, S.; Golden, J. Directional RNA deep sequencing sheds new light on the transcriptional response of Anabaena sp. strain PCC 7120 to combined-nitrogen deprivation. BMC Genom. 2011, 12, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maldener, I.; Hannus, S.; Kammerer, M. Description of five mutants of the cyanobacterium Anabaena sp. strain PCC 7120 affected in heterocyst differentiation and identification of the transposon-tagged genes. FEMS Microbiol. Lett. 2003, 224, 205–213. [Google Scholar] [CrossRef]
- Khudyakov, I.; Wolk, C.P. hetC, a gene coding for a protein similar to bacterial ABC protein exporters, is involved in early regulation of heterocyst differentiation in Anabaena sp. strain PCC 7120. J. Bacteriol. 1997, 179, 6971–6978. [Google Scholar] [CrossRef] [PubMed]
- Holland, D.; Wolk, C.P. Identification and characterization of hetA, a gene that acts early in the process of morphological differentiation of heterocysts. J. Bacteriol. 1990, 172, 3131–3137. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shvarev, D.; Nishi, C.N.; Wörmer, L.; Maldener, I. The ABC Transporter Components HgdB and HgdC are Important for Glycolipid Layer Composition and Function of Heterocysts in Anabaena sp. PCC 7120. Life 2018, 8, 26. https://doi.org/10.3390/life8030026
Shvarev D, Nishi CN, Wörmer L, Maldener I. The ABC Transporter Components HgdB and HgdC are Important for Glycolipid Layer Composition and Function of Heterocysts in Anabaena sp. PCC 7120. Life. 2018; 8(3):26. https://doi.org/10.3390/life8030026
Chicago/Turabian StyleShvarev, Dmitry, Carolina N. Nishi, Lars Wörmer, and Iris Maldener. 2018. "The ABC Transporter Components HgdB and HgdC are Important for Glycolipid Layer Composition and Function of Heterocysts in Anabaena sp. PCC 7120" Life 8, no. 3: 26. https://doi.org/10.3390/life8030026