
Acetate Metabolism in Anaerobes from the Domain Archaea

Department of Biochemistry and Molecular Biology, Pennsylvania State University, University Park, PA 16802, USA
Life 2015, 5(2), 1454-1471; https://doi.org/10.3390/life5021454
Submission received: 16 April 2015
/
Accepted: 1 June 2015
/
Published: 9 June 2015
(This article belongs to the Special Issue Archaea: Evolution, Physiology, and Molecular Biology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Acetate and acetyl-CoA play fundamental roles in all of biology, including anaerobic prokaryotes from the domains Bacteria and Archaea, which compose an estimated quarter of all living protoplasm in Earth’s biosphere. Anaerobes from the domain Archaea contribute to the global carbon cycle by metabolizing acetate as a growth substrate or product. They are components of anaerobic microbial food chains converting complex organic matter to methane, and many fix CO2 into cell material via synthesis of acetyl-CoA. They are found in a diversity of ecological habitats ranging from the digestive tracts of insects to deep-sea hydrothermal vents, and synthesize a plethora of novel enzymes with biotechnological potential. Ecological investigations suggest that still more acetate-metabolizing species with novel properties await discovery.
1. Introduction
Acetate and acetyl-CoA play a prominent role in the metabolism of all three phylogenetic domains of life, including anaerobic prokaryotes from the domains Bacteria and Archaea, which contribute to an estimated quarter of all living protoplasm in Earth’s biosphere [1]. Anaerobes from the domain Archaea play significant roles in the global carbon cycle by metabolizing acetate as a growth substrate or product. Many anaerobes also fix CO2 into cell material via synthesis of acetyl-CoA [2].
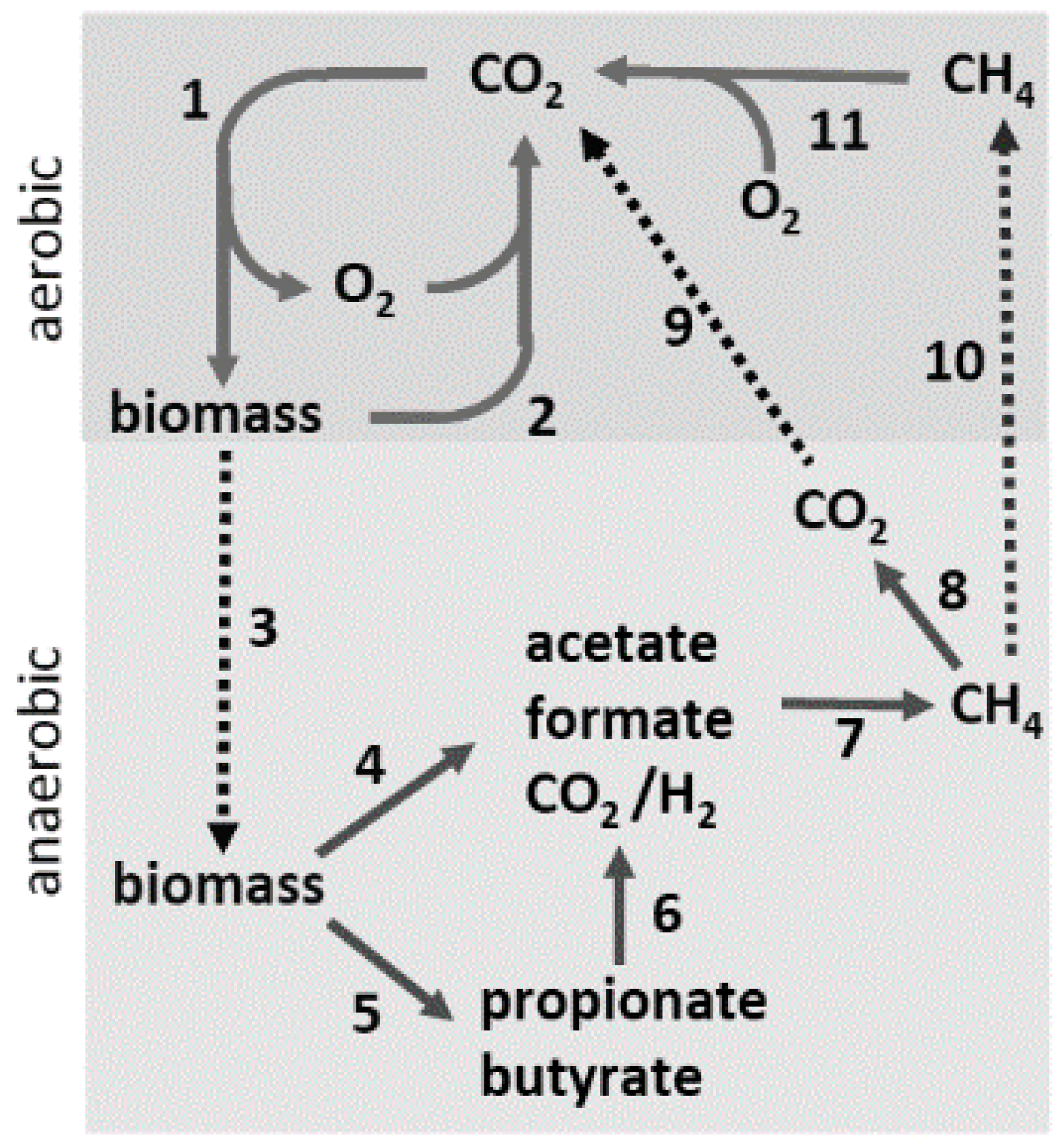
In anaerobic environments where terminal electron acceptors (Fe(III), Mn(IV), SO42−, S0, NO3−) are absent, anaerobes from both prokaryotic domains convert complex organic matter to CH4 and CO2, providing an essential link in the global carbon cycle (Figure 1). In Step 1, CO2 is incorporated into biomass driven primarily by photosynthesis. In environments where O2 is abundant, microbes oxidize the biomass, producing CO2 that re-enters the carbon cycle (Step 2). A significant portion of the biomass is diverted to an assortment of anaerobic habitats devoid of terminal electron acceptors (Step 3), where anaerobic microbial food chains, comprised of at least three distinct metabolic groups, digest the biomass to CO2 and CH4 (Steps 4–7). The fermentative group converts the complex biomass primarily into, acetate along with lesser amounts of volatile fatty acids, H2, and CO2 (Steps 4 and 5). The fatty acids are converted chiefly into acetate plus either formate or H2 by the acetogenic group (Step 6). Thus, acetate emerges as the principal product of the fermentative and acetogenic groups. The CH4-producing (methanogen) group is sub-divided into acetate-utilizing (acetoclastic) and CO2-reducing species. Acetoclastic species convert the methyl group of acetate to CH4 and the carbonyl group to CO2. The CO2-reducing species reduce CO2 to CH4 with electrons derived from H2 or formate. At least two-thirds of the CH4 produced derives from acetate, the central intermediary in anaerobic microbial food chains. A portion of the CH4 is oxidized to CO2 (Step 8) by associations of anaerobes that utilize sulfate, nitrate, manganese, or iron as terminal electron acceptors [3]. The CO2 and remaining CH4 diffuse into aerobic zones (Steps 9 and 10), where O2-requiring methanotrophic microbes oxidize the CH4 to CO2 (Step 11), completing the carbon cycle. Anaerobes also participate in chemoautotrophic habitats, where they fix carbon dioxide in catabolic and anabolic pathways. Although the fermentative and acetogenic groups are largely populated with characterized isolates from the domain Bacteria, all characterized methanogens are classified in the domain Archaea. Acetate-utilizing anaerobes classified in the domain Archaea also proliferate in environments where terminal electron acceptors are abundant and obtain energy through anaerobic respiration, converting acetate to CO2.
Figure 1.
The global carbon cycle. Solid lines indicate steps in the cycle (see text) and dotted lines indicate transfer of material between aerobic and anaerobic environments.
Figure 1.
The global carbon cycle. Solid lines indicate steps in the cycle (see text) and dotted lines indicate transfer of material between aerobic and anaerobic environments.
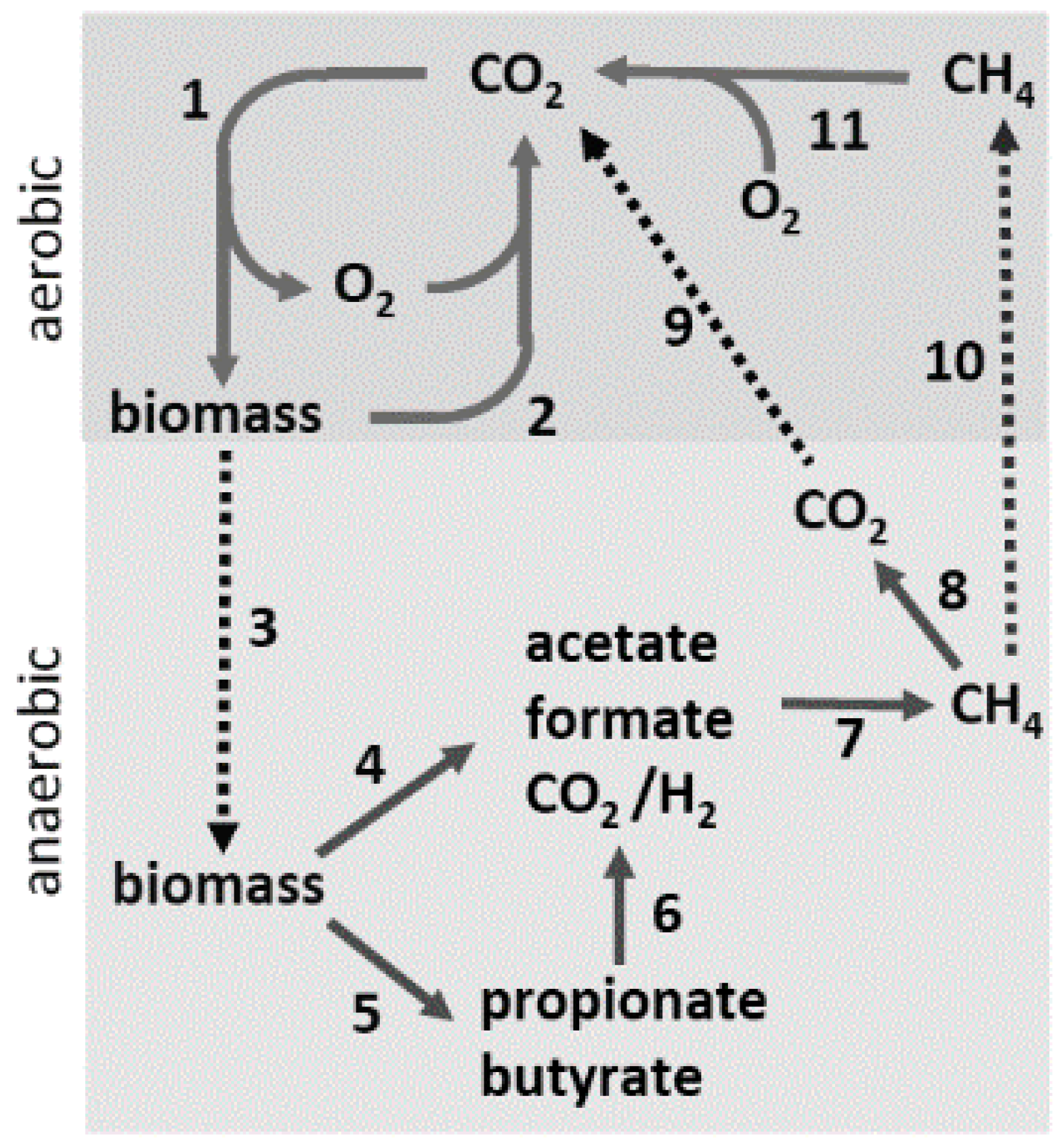
This review encompasses the role of acetate, which has the greatest influence on the ecology of environments, in the energy conversion pathways of anaerobes from the domain Archaea; however, acetate and acetyl-CoA also play a prominent role in the biosynthetic pathways of anaerobes from the domain Archaea [2].
2. Acetate Production
2.1. Heterotrophic Energy-Converting Pathways Producing Acetate
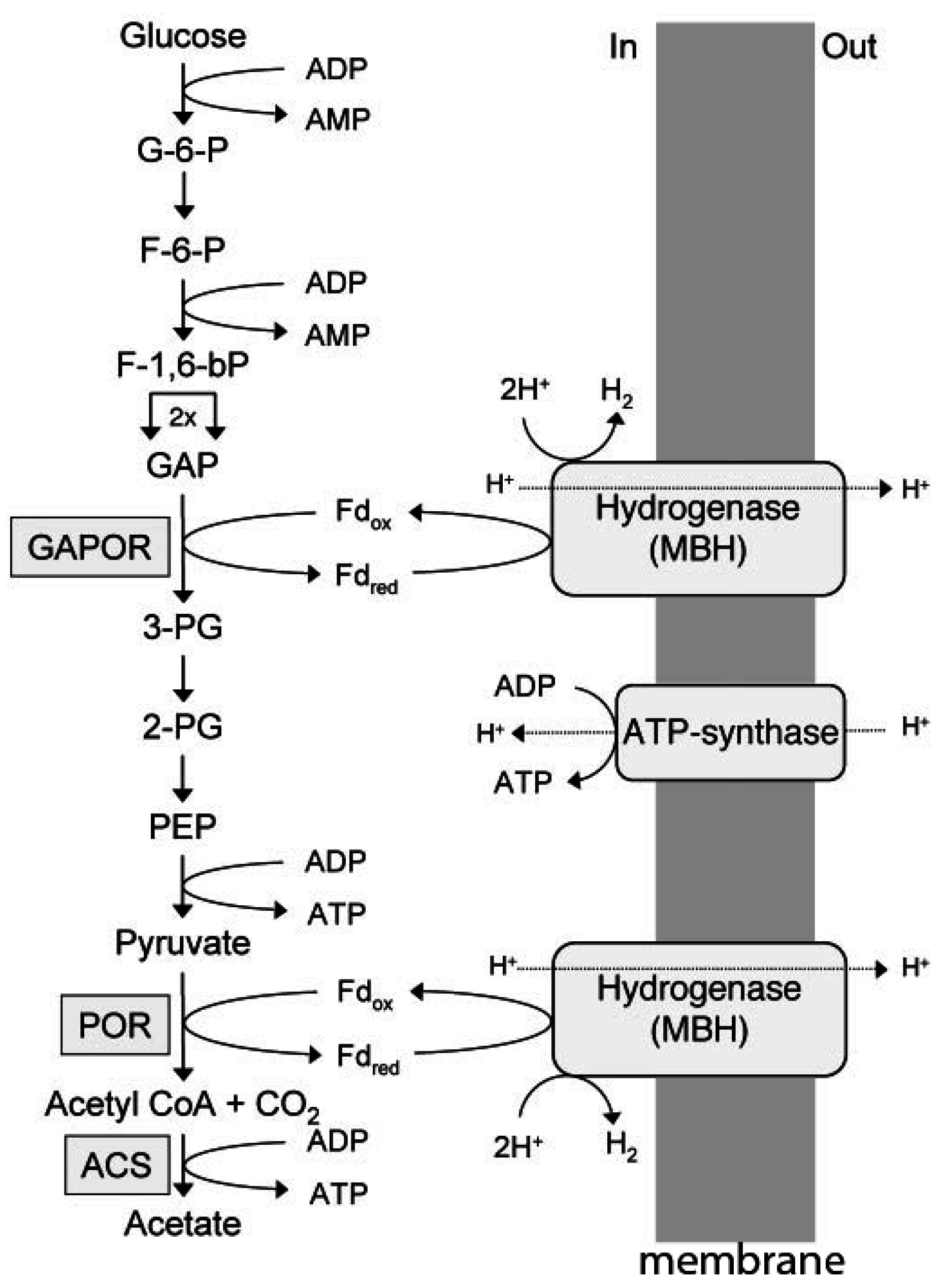
Investigations of heterotrophic hyperthermophilic species from the domain Archaea have revealed pathways that deviate substantially from pathways in heterotrophic organisms from the domain Bacteria. Pyrococcus furiosus, for example, grows at 100 °C and ferments carbohydrates to acetate, CO2, and H2 by an unusual Emden–Meyerhof pathway involving the novel enzymes ADP-dependent glucokinase, ADP-dependent phosphofruktokinase, glyceraldehyde-3-phosphate ferredoxin oxidoreductase (GAPOR), phosphoenolpyruvate synthase, pyruvate: Ferredoxin oxidoreductase (POR), and ADP-forming acetyl-CoA synthetase (Figure 2) [4]. Other heterotrophic acetate-producing hyperthermophiles that utilize one or more of these enzymes include species from the genera Thermococcus, Desulfurococcus, Staphylothermus, and Archaeoglobus [5,6,7,8]. In the glycolytic pathway exemplified by P. furiosus (Figure 2), ferredoxin reduced by GAPOR and POR is re-oxidized by a membrane-bound hydrogenase that generates an ion gradient driving ATP synthesis [4]. GAPOR is an oxygen-sensitive homomonomer with a molecular mass of 63 kDa, and contains a pterin cofactor, one tungsten, and six iron atoms per monomer [9]. Pyruvate is oxidized to acetyl-CoA, catalyzed by pyruvate: Ferredoxin oxidoreductase (POR). The enzyme has a molecular mass of 100 kDa and is comprised of three subunits (45, 31, and 24 kDa), and contains thiamine pyrophosphate (TPP) and two ferredoxin-type [4Fe-4S] clusters [10]. The enzyme requires CoASH but not TPP for pyruvate oxidation activity. The POR also catalyzes the formation of acetaldehyde from pyruvate in a CoA-dependent reaction, although the cofactor plays a structural rather than catalytic role [11]. Acetyl-CoA is converted to acetate by an ADP-forming acetyl-CoA synthetase, which generates ATP by substrate level phosphorylation [12,13]. Conversion of acetyl-CoA to acetate by acetyl-CoA synthetases is characteristic of the domain Archaea, in contrast to the domain Bacteria, in which phosphotransacetylase and acetate kinase predominate [14]. A reaction mechanism has been proposed for the heterotetrameric (α2β2) enzyme from P. furiosus that follows a four-step mechanism including transient phosphorylation of two active site histidine residues (Equations (1)–(4)) [15].
E + acetyl-CoA + Pi = E·acetyl~P + CoASH
E·acetyl~P = acetate + E-His257α~P
E-His257α~P = E-His71β~P
E-His71β~P + ADP = ATP + E
Archaeoglobus fulgidus is an example of a heterotrophic hyperthermophile from the domain Archaea that utilizes terminal electron acceptors in place of reducing protons and producing H2. A strain of A. fulgidus grows with starch as the sole source of carbon and energy by an incomplete oxidation of glucose to acetate and CO2, utilizing a modified Embden–Meyerhof pathway resembling that of P. furiosus [8]. However, in contrast to H2-producing P. furiosus, electrons derived from the oxidation are transferred to sulfate, producing sulfide (Equation (5)):
C6H12O6 + H2SO4 = 2CO2 + 2C2H4O2 + H2S + 2H2O.
Figure 2.
Electron transport and energy conversion during glucose catabolism by P. furiosus. G-6-P, glucose 6-phosphate; F-6-P, fructose 6-phosphate; F-1,6-bP, fructose 1,6-bisphosphate; GAP, glyceraldehyde phosphate, GAPOR, GAP: Ferredoxin oxidoreductase; 3-PG, 3-phosphoglycerate; 2-PG, 2-phosphoglycerate; PEP, phosphoenolpyruvate; POR, pyruvate: Ferredoxin oxidoreductase; ACS, acetyl-CoA synthase. Reproduced with permission [4]. Copyright (2006) National Academy of Sciences, USA.
Figure 2.
Electron transport and energy conversion during glucose catabolism by P. furiosus. G-6-P, glucose 6-phosphate; F-6-P, fructose 6-phosphate; F-1,6-bP, fructose 1,6-bisphosphate; GAP, glyceraldehyde phosphate, GAPOR, GAP: Ferredoxin oxidoreductase; 3-PG, 3-phosphoglycerate; 2-PG, 2-phosphoglycerate; PEP, phosphoenolpyruvate; POR, pyruvate: Ferredoxin oxidoreductase; ACS, acetyl-CoA synthase. Reproduced with permission [4]. Copyright (2006) National Academy of Sciences, USA.

2.2. Chemolithotrophic Energy-Converting Pathways Producing Acetate
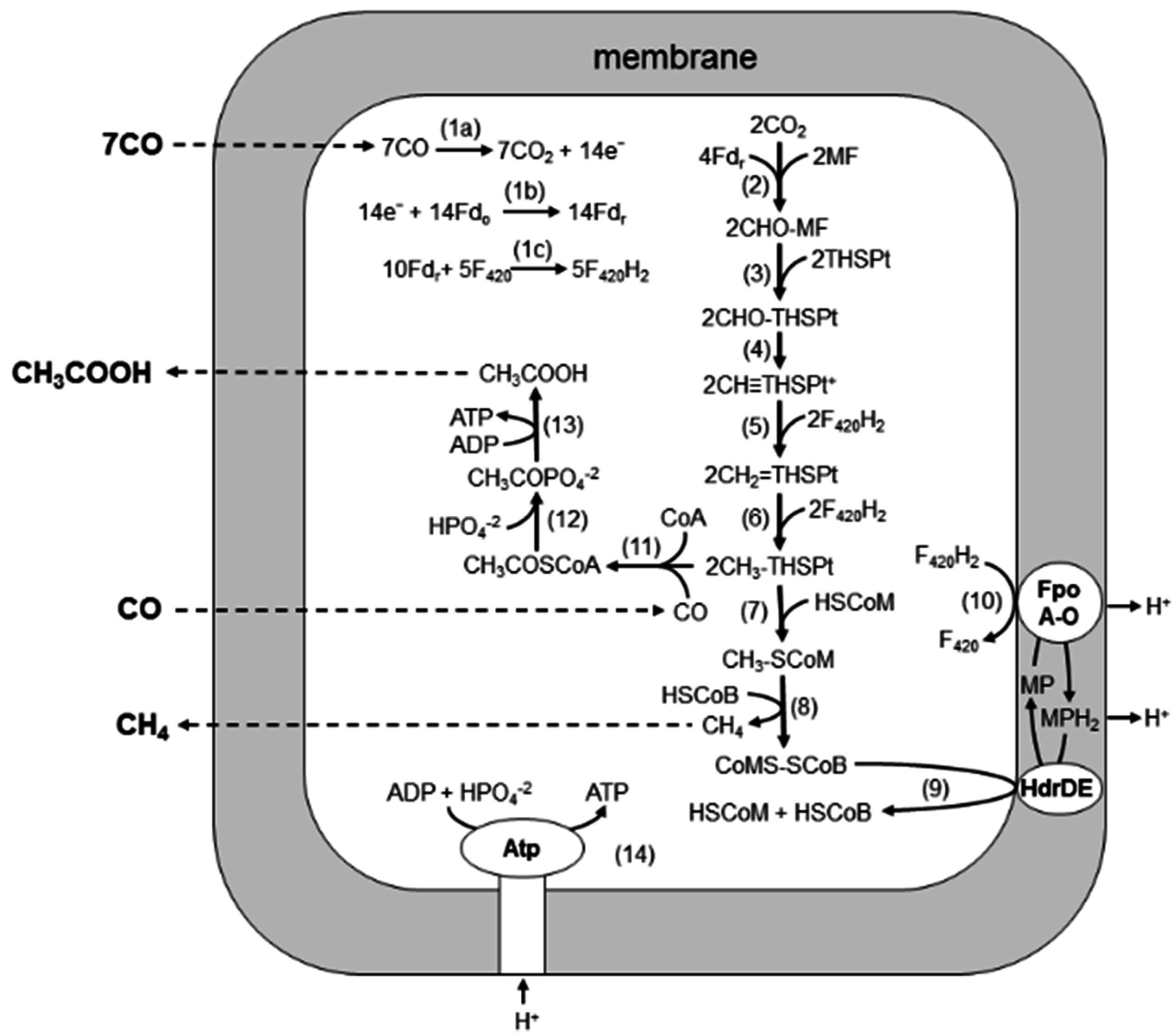
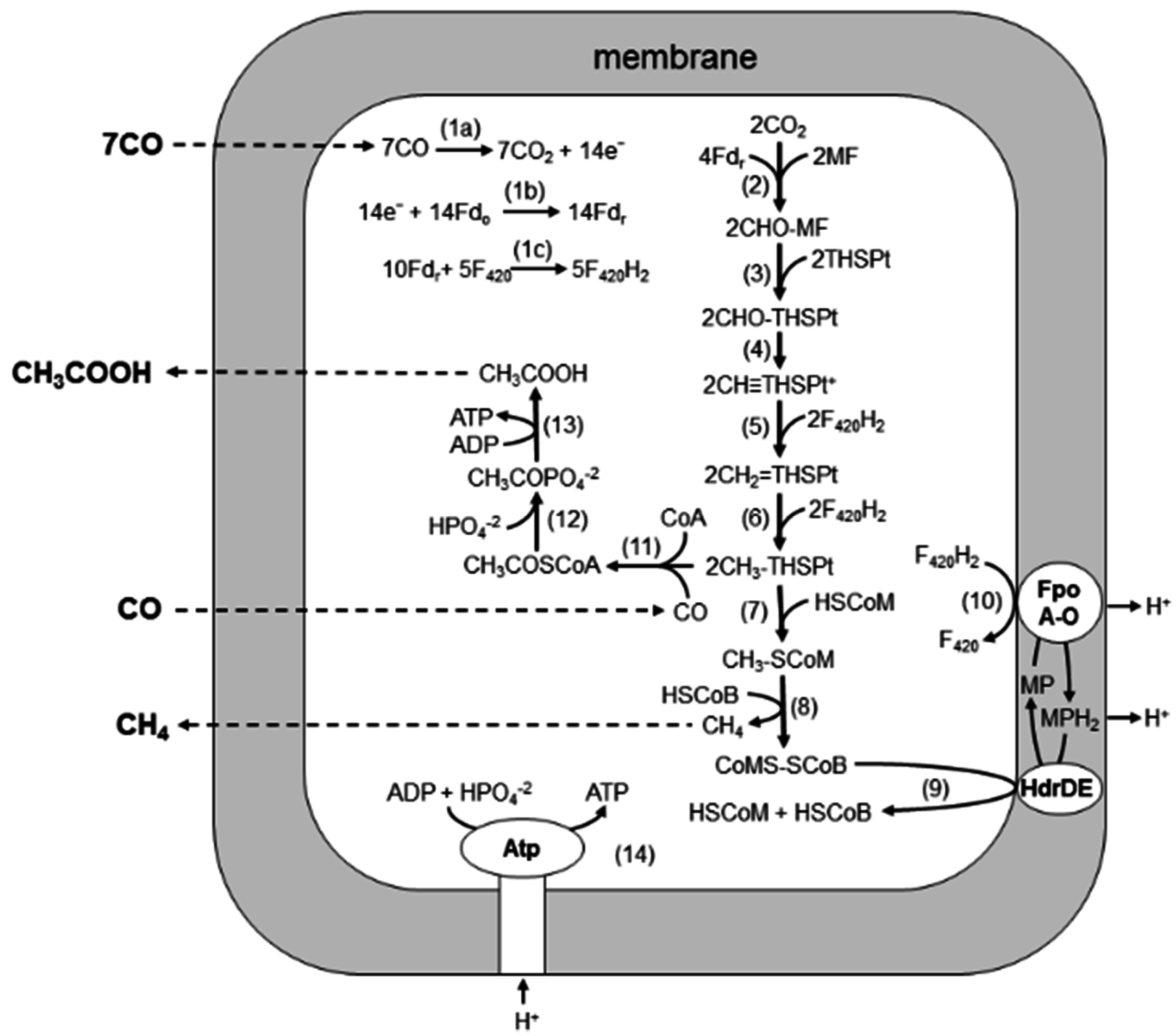
Biochemical and quantitative proteomic analyses of M. acetivorans revealed a pathway for CO-dependent growth of M. acetivorans in which both acetate and CH4 are products [16,17] (Figure 3). In the pathway, electrons derived from the oxidation of CO (Reaction 1a–c) are used to reduce CO2 to a methyl group attached to THSPt (Reactions 2–6), similar to the pathway of obligate H2-utilizing CO2-reducing methanogens, which are the subject of several reviews recommended to the reader [18,19,20,21]. The CO dehydrogenase/acetyl-CoA synthetase CODH/ACS is proposed to oxidize CO-reducing ferredoxin, which donates electrons in the first reductive step (Reaction 2). It is possible that ferredoxin also donates electrons to FpoF, catalyzing the reduction of coenzyme F420 to F420H2 (Reaction 1c) that donates electrons to complete the reduction of CO2 to the methyl level (Reactions 5 and 6). Indeed, it was recently shown that the FpoF of Methanosarcina mazei catalyzes reaction 1c [22] and a homolog in M. acetivorans (encoded by MA3732) is upregulated 11-fold in CO- vs. acetate-grown cells [16]. The methyl group of methyl-THSPt is reduced to CH4 by reactions common to all methanogenic pathways (Reactions 7–9), albeit with two exceptions. First, a membrane-bound complex (FpoA-O) oxidizes F420H2 and transfers electrons to HdrDE mediated by MP, which generates a proton gradient (Reactions 9 and 10) driving ATP synthesis (Reaction 14). Second, transfer of the methyl group of methyl-THSPt to HS-CoM (Reaction 7) is thought to be catalyzed by two enzymes, the membrane-bound Mtr complex coupled to Na+ translocation, common to all methanogenic pathways; and a soluble enzyme (CmtA) unique to the CO-dependent CO2 reduction pathway [23]. Acetate is a prominent product in addition to CH4. Levels of CODH/ACS, phosphotransacetylase, and acetate kinase are prominent in CO-grown cells [16], supporting a route to acetate wherein CODH/ACS synthesizes acetyl-CoA with the methyl group of methyl-THSPt, CO, and CoA-SH (Reaction 11), which are further converted to acetate by phosphotransacetylase and acetate kinase (Reactions 12 and 13) with the production of ATP. Thus, ATP is synthesized via both substrate level and chemiosmotic mechanisms.
A. fulgidus uses essentially the same pathway during CO-dependent growth, employing similar enzymes and electron carriers [24,25,26,27,28], except for conversion of acetyl-CoA to acetate, which is catalyzed by either an AMP- or ADP-forming acetyl-CoA synthetase [29,30,31]. A. fulgidus does not produce methane, and acetate is the sole product in the absence of sulfate.
Figure 3.
Pathway for conversion of CO to acetate and CH4 by M. acetivorans. See text. Fdo, oxidized ferredoxin; Fdr, reduced ferredoxin; F420, coenzyme F420; MF, methanofuran; THSPt, tetrahydrosarcinapterin; HSCoM, coenzyme M; HSCoB, coenzyme B; Fpo, F420H2 dehydrogenase complex; MP, methanophenazine; Hdr, heterodisulfide reductase. Reproduced from [16] with permission. Copyright (2006) National Academy of Sciences, USA.
Figure 3.
Pathway for conversion of CO to acetate and CH4 by M. acetivorans. See text. Fdo, oxidized ferredoxin; Fdr, reduced ferredoxin; F420, coenzyme F420; MF, methanofuran; THSPt, tetrahydrosarcinapterin; HSCoM, coenzyme M; HSCoB, coenzyme B; Fpo, F420H2 dehydrogenase complex; MP, methanophenazine; Hdr, heterodisulfide reductase. Reproduced from [16] with permission. Copyright (2006) National Academy of Sciences, USA.
3. Acetate Utilization
3.1. Acetotrophic Energy-Converting Pathways Producing Methane
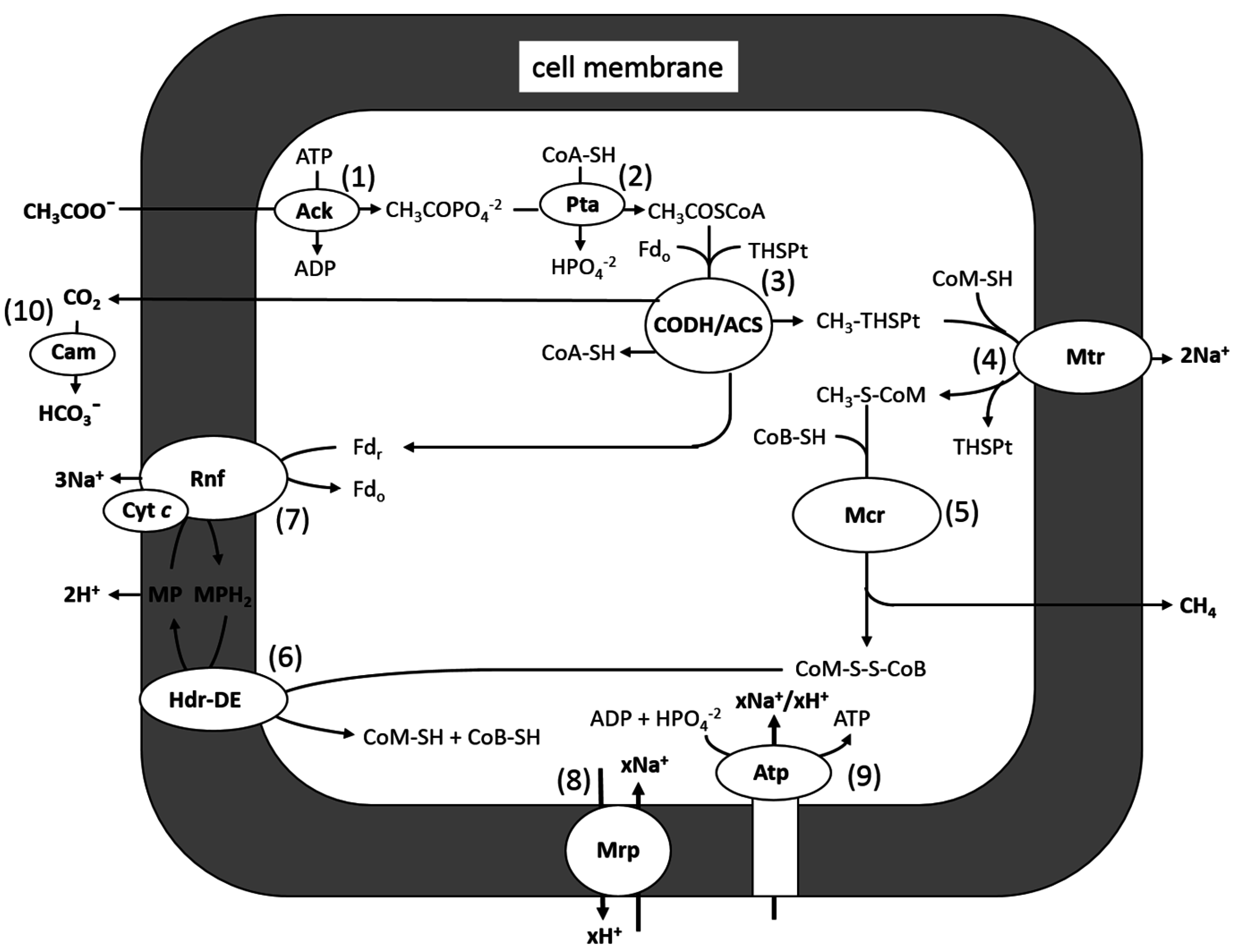
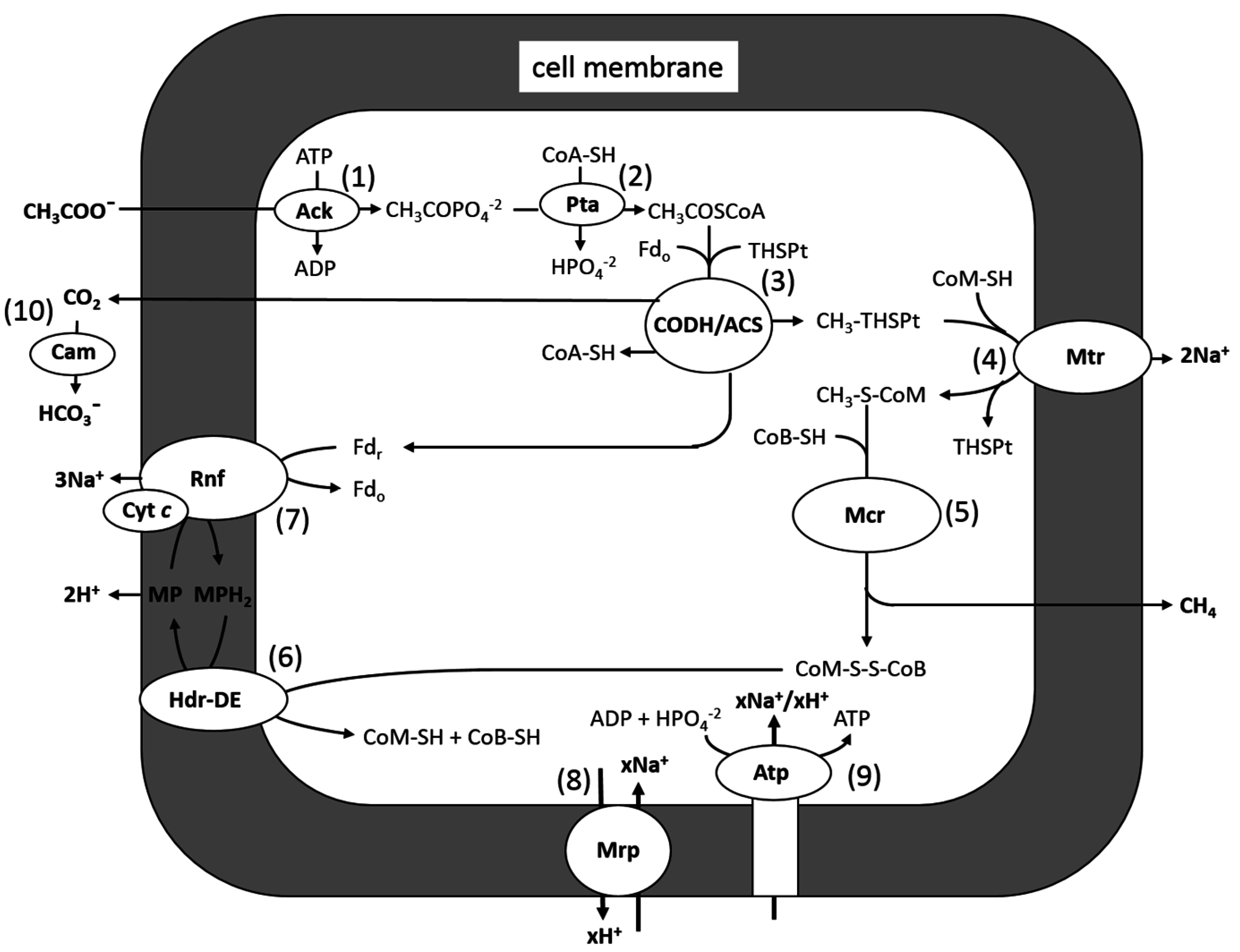
Methane-producing species from the genera Methanosarcina and Methanosaeta are the only known acetoclastic genera in the domain Archaea. The majority of studies on the acetoclastic pathway have involved Methanosarcina species, for which M. acetivorans is a model (Figure 4). Carbon transfer reactions in the pathway can be divided into two parts: (1) Reactions 1–3: activation to acetyl-CoA and cleaving the C-C bond of the acetyl group, yielding CH3-THSPt and CO2, which are unique to the pathway; and (2) Reactions 4–6: reducing the methyl group of CH3-THSPt to CH4, which is common to all methanogenic pathways. Methanogenesis, and by inference reactions common to all methanogenic pathways, is thought to have evolved soon after the origin of life, approximately 3.75 billion years ago [32,33]. However, evolution of the acetoclastic pathway is proposed to have evolved approximately 250–300 million years ago during the end-Permian carbon cycle that contributed to the mass extinction of that period [34]. Reactions 2 and 3, catalyzed by acetate kinase and CO dehydrogenase/acetyl-CoA synthase CODH/ACS, are thought to be ancient enzymes evolving soon after the origin of life [35,36]. Homologs of acetate kinase and phosphotransacetylase are key enzymes in energy-yielding pathways of fermentative and acetogenic species from the domain Bacteria, converting acetyl-CoA to ATP and acetate. These observations are consistent with the evolution of the aceticlastic pathway by horizontal gene transfer of enzymes catalyzing Reactions 1–3 from the domain Bacteria and grafting on to Reactions 4–6, common to more ancient methanogenic pathways. Reactions 4 and 5, and the enzymes catalyzing them, are the subject of recent reviews that are recommended to the reader [37,38].
Figure 4.
Pathway of aceticlastic methanogenesis in M. acetivorans. Ack, acetate kinase; Pta, phosphotransacetylase; CoA-SH, coenzyme A; THSPt, tetrahydrosarcinapterin; Fdr, reduced ferredoxin; Fdo, oxidized ferredoxin; Cdh, CO dehydrogenase/acetyl-CoA synthase; CoM-SH, coenzyme M; Mtr, methyl-THSPt:CoM-SH methyltransferase; CoB-SH, coenzyme B; MP, methanophenazine; Hdr-DE, heterodisulfide reductase; Rnf, Rnf complex; Mrp, Mrp complex; Atp, ATP synthase. Modified from [39].
Figure 4.
Pathway of aceticlastic methanogenesis in M. acetivorans. Ack, acetate kinase; Pta, phosphotransacetylase; CoA-SH, coenzyme A; THSPt, tetrahydrosarcinapterin; Fdr, reduced ferredoxin; Fdo, oxidized ferredoxin; Cdh, CO dehydrogenase/acetyl-CoA synthase; CoM-SH, coenzyme M; Mtr, methyl-THSPt:CoM-SH methyltransferase; CoB-SH, coenzyme B; MP, methanophenazine; Hdr-DE, heterodisulfide reductase; Rnf, Rnf complex; Mrp, Mrp complex; Atp, ATP synthase. Modified from [39].
The first crystal structure for any acetate kinase was from the acetoclastic methanogen Methanosarcina thermophila, which revealed properties suggesting that the enzyme is the founding member of the Acetate and Sugar Kinase/Hsc70/Actin (ASKHA) superfamily of phosphotransferases [35]. Structural and biochemical investigations established a direct in-line mechanism in which the carboxylate anion attacks the γ-phosphate of ATP enabling transfer of the phosphate group [40,41,42,43,44,45,46,47,48]. The first crystal structure of any phosphotransacetylase was also from M. thermophila, the structural and biochemical analyses of which indicate a mechanism that proceeds through base-catalyzed abstraction of the HS-CoA thiol proton and subsequent nucleophilic attack of −S-CoA on the carbonyl carbon of acetyl phosphate [49].
The CODH/ACS complex [50,51,52,53,54,55,56,57,58,59] cleaves the C-C and C-S bonds of acetyl-CoA, transferring the methyl group to THSPt and oxidizing the carbonyl group to CO2 with transfer of electrons to ferredoxin [60,61,62] (Reaction 3). The same enzyme complex functions in reverse to synthesize acetyl-CoA for cell carbon in CO2-reducing methanogens and oxidizes exogenous CO in the pathway of CO conversion to CH4, CO2, and acetate in M. acetivorans. Of ancient origin, primitive ancestors of CODH/ACS likely played a central role in the early evolution of life [36,63,64]. Although a two-subunit enzyme has been purified and characterized from an acetate-utilizing species of the genus Methanosaeta [54,57,58,59], the majority of mechanistic studies have been with the five-subunit (αβγδε) complexes from the acetate-utilizing species M. thermophila and Methanosarcina barkeri. The complexes are resolvable into three components containing the αε, γδ, or β subunit(s) [65]. The αε component catalyzes the oxidation of CO and reduction of ferredoxin [60,66]. The crystal structure from M. barkeri shows a α2ε2 arrangement with the α subunit containing four 4Fe-4S clusters and a NiFe3S4 cluster bridged to an exogenous iron atom called the “C” cluster [67]. Two of the 4Fe-4S clusters are postulated to function in electron transport from the active site “C” cluster to ferredoxin. The structure suggests coupling between CO bound to the nickel and H2O/OH− bound to the exogenous iron in the C=O bond-forming step leading to the oxidized product CO2. The structure also shows a gas channel extending from the “C” cluster to the protein surface with the potential to interface with the β component containing the “A” cluster, thus catalyzing acetyl-CoA cleavage and carbonyl group conversion to CO [68,69,70,71]. Although the structure is unknown, spectroscopic investigations indicate that the “A” cluster is comprised of a 4Fe-4S center bridged to a binuclear Ni-Ni site [70,72], similar to the homolog from an acetate-producing species from the domain Bacteria that synthesizes acetyl-CoA [73]. A mechanism is proposed in which transfer of an electron from “C” to “A” maintains the reduced catalytically active Ni(I) redox state of “A” [74]. Both kinetic and EPR approaches support the fact that alterations in the Ni coordination environment of the “A” cluster promote C−C bond cleavage, dependent on changes in the protein conformation from the open to closed state [75,76]. Moreover, CO is proposed to be an inhibitor of C-C bond cleavage; thus, control over C−C bond cleavage in concert with containment of CO in the gas channel explains the requirement for tight coupling of the decarbonylation reaction for efficient transfer of CO to “C” for oxidation [75]. The γδ component transfers the methyl group of acetyl-CoA to THSPt, involving a corrinoid cofactor and an iron-sulfur cluster [55,77,78,79], although it has yet to be determined which of the two subunits interact with THSPt. Spectroscopic EPR analyses indicate that the corrinoid cofactor is maintained in the base-off state with a of −486 mV for the Co2+/1+ couple that facilitates reduction of Co2+ to Co1+ required for methylation of the corrinoid [77]. The analysis also identified a 4Fe-4S cluster with a midpoint potential close to the Co2+/1+ couple, suggesting that the cluster is involved in reducing Co2+.
The conversion of acetate to CH4 and CO2 provides only a marginal amount of energy available for growth (ΔG0' = −36 kJ/CH4) that is spent on the ATP consumed in the activation to acetyl-CoA (Figure 4), which illustrates the importance of cells maximizing thermodynamic efficiency. A theoretical analysis of acetate-grown M. barkeri indicates that transfer of CH4 and CO2 into the gaseous phase contributes to the driving force of growth [80]. Thus it has been proposed that a carbonic anhydrase (Cam) from M. thermophila is located outside the cell membrane, where it converts CO2 to membrane-impermeable HCO3− (Reaction 10, Figure 4), thereby facilitating removal of CO2 from the cytoplasm [81]. Cam from M. thermophila is the archetype of an independently evolved class (γ class) of carbonic anhydrases that contains Fe2+ in the active site, contrary to all prokaryotic carbonic anhydrases, which contain zinc [82,83]. Structural and biochemical analyses [84] support a two-step ping pong mechanism, shown in Equations (6) and (7), where E represents enzyme residues, M is metal, and B is the buffer.
E-Fe2+-OH− + CO2 = E-Fe2+-HCO3−
E-Fe2+-HCO3− + H2O = E-Fe2+-H2O + HCO3−
E-Fe2+-H2O = H+-E-Fe2+-OH−
H+-E-Fe2+-OH− + B = E-Fe2+-OH− + BH+
In Step 1 a lone pair of electrons on the metal-bound hydroxide attacks CO2-producing metal-bound bicarbonate (Equation (6a)), which is subsequently displaced by water (Equation (6b)). In Step 2 a proton is extracted from the metal-bound water (Equation (7a)), and then transferred to the buffer (Equation (7b)).
ATP is synthesized by a chemiosmotic mechanism. Ferredoxin accepts electrons derived from the oxidation of the carbonyl group of acetyl-CoA by CODH/ACS in both H2-dependent and H2-independent acetotrophic methanogens. Both types also obtain energy for growth by coupling electron transfer from ferredoxin to the heterodisulfide CoM-S-S-CoB, with translocation of ions generating a gradient that drives ATP synthesis catalyzed by an A1A0-type ATP synthase [85,86,87,88]. The reduced ferredoxin of H2-dependent species donates electrons to a membrane-bound hydrogenase (Ech), evolving H2 and translocating protons [89,90,91]. It is proposed that a hydrogenase (Vho) reoxidizes H2 and donates electrons to a quinone-like electron carrier called methanophenazine (MP) [92]. The MP donates electrons to the heterodisulfide reductase HdrDE concomitant with translocation of two protons contributing to the gradient. An additional two protons are translocated by the Vho. However, most acetotrophic methanogens are H2 independent [93,94,95,96,97,98,99,100,101,102,103,104] and likely evolved a different strategy for oxidizing ferredoxin and reducing CoM-S-S-CoB typified by M. acetivorans [105,106] (Figure 4). The genome does not encode a functional Ech; instead, acetate-grown cells preferentially synthesize a Rnf complex [87] similar to the six-subunit Rnf complexes in microbes from the domain Bacteria [107,108,109,110,111,112,113]. A Δrnf strain is unable to grow with acetate, confirming that the complex is essential [114]. Unlike all other characterized Rnf complexes, the contiguous genes encoding the six-subunit core complex from M. acetivorans are co-transcribed with a gene encoding a multiheme cytochrome c abundant in membranes of acetate-grown cells [87]. A topology model [39] predicts roles for each of the six core subunits and cytochrome c wherein MP mediates electron transfer between cytochrome c and HdrDE, translocating a pair of protons [39]. It has been shown that the Rnf complex translocates Na+ (Reaction 7), joining the Na+ translocating methyl transfer (Reaction 4) [114]. Thus, both Na+ and H+ gradients are generated during growth with acetate. Notably, the A1A0-type ATP synthase of M. acetivorans is dependent on both Na+ and H+ gradients [115]. A multisubunit Na+/H+ antiporter (Mrp) is proposed to adjust the Na+/H+ ratio (Reaction 8) optimal for ATP synthesis by the A1A0-type ATP synthase (Reaction 9) [116].
A genome-wide analysis of Methanosaeta thermophila has revealed genes encoding enzymes catalyzing reactions in the pathway of acetate to CH4 identical to Methanosarcina species except for the activation of acetate to acetyl-CoA, which is catalyzed by an AMP- and PPi-forming acetyl-CoA synthetase [117,118,119,120]. The synthetase has a several-fold lower Km for acetate (0.4 mM) than acetate kinase from M. thermophila (22 mM) [118,121], a result consistent with Methanosaeta species dominating over Methanosarcina species in environments where acetate is in low concentrations. Genes encoding Ech and Rnf are absent in the genome of Methanosaeta, suggesting an unknown alternative electron transport pathway and mechanism for energy conservation [117].
3.2. Acetotrophic Energy-Converting Pathways Reducing Exogenous Electron Acceptors
Acetate-utilizing prokaryotes from the domain Archaea also obtain energy by anaerobic respiration. Ferroglobus placidus and Geoglobus ahangari are hyperthermophiles growing at 85 °C by oxidizing acetate to CO2, only with Fe(III) serving as the electron acceptor [122]. Geoglobus acetivorans is another hyperthermophile growing optimally at 81 °C and utilizing acetate in addition to formate, pyruvate, fumarate, malate, propionate, butyrate, succinate, glycerol, stearate, palmitate, peptone, and yeast extract as electron donors for Fe(III) reduction. The organism is also able to grow with H2 as the electron donor and Fe(III) as an electron acceptor without the need for organic substances [123]. Hyperthermophilic Thermococcus species have also been implicated in oxidizing acetate and reducing Fe(III) [124].
4. Concluding Remarks
Although heterotrophic acetate-producing hyperthermophiles are abundantly documented, no hyperthermophilic acetoclastic methanogen has been described that presents a disconnect in the ecology of these two metabolic groups for which cognate mesophiles function syntrophically in anaerobic microbial food chains, converting complex biomass to CH4. One possibility is that hyperthermophilic temperatures are a thermodynamic barrier to the conversion of acetate to CH4. Additionally, although it appears that heterotrophic organisms from the domain Archaea (Thaumarchaea) proliferate in mesothermal anaerobic environments [125], isolates and details of their metabolism are largely unknown. Thus, the finding that anaerobic respiratory acetate-oxidizing species are found in hyperthermophilic environments suggests the possibility that acetate-producing heterotrophs like P. furiosus supply acetate in a two-component microbial food chain converting complex organics to CO2.
Acknowledgments
Funding for research performed in the author’s laboratory derived from: National Science Foundation No. 0820734; Department of Energy, Energy Biosciences Program No. DE-FG02-95ER20198, and the National Institutes of Health No. GM44661.
Conflicts of Interest
The author declares no conflict of interest.
References
- Whitman, W.B.; Coleman, D.C.; Wiebe, W.J. Prokaryotes: The Unseen Majority. Proc. Natl. Acad. Sci. USA 1998, 95, 6578–6583. [Google Scholar] [CrossRef] [PubMed]
- Berg, I.A.; Kockelkorn, D.; Ramos-Vera, W.H.; Say, R.F.; Zarzycki, J.; Hugler, M.; Alber, B.E.; Fuchs, G. Autotrophic Carbon Fixation in Archaea. Nat. Rev. Microbiol. 2010, 8, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Thauer, R.K. Functionalization of Methane in Anaerobic Microorganisms. Angew. Chem. Int. Ed. Engl. 2010, 49, 6712–6713. [Google Scholar] [CrossRef] [PubMed]
- Sapra, R.; Bagramyan, K.; Adams, M.W. A Simple Energy-Conserving System: Proton Reduction Coupled to Proton Translocation. Proc. Natl. Acad. Sci. USA 2003, 100, 7545–7550. [Google Scholar] [CrossRef] [PubMed]
- Selig, M.; Xavier, K.B.; Santos, H.; Schonheit, P. Comparative Analysis of Embden-Meyerhof and Entner-Doudoroff Glycolytic Pathways in Hyperthermophilic Archaea and the Bacterium Thermotoga. Arch. Microbiol. 1997, 167, 217–232. [Google Scholar] [PubMed]
- Perevalova, A.A.; Svetlichny, V.A.; Kublanov, I.V.; Chernyh, N.A.; Kostrikina, N.A.; Tourova, T.P.; Kuznetsov, B.B.; Bonch-Osmolovskaya, E.A. Desulfurococcus Fermentans Sp. Nov., A Novel Hyperthermophilic Archaeon from a Kamchatka Hot Spring, and Emended Description of the Genus Desulfurococcus. Int. J. Syst. Evol. Microbiol. 2005, 55, 995–999. [Google Scholar] [CrossRef] [PubMed]
- Anderson, I.J.; Dharmarajan, L.; Rodriguez, J.; Hooper, S.; Porat, I.; Ulrich, L.E.; Elkins, J.G.; Mavromatis, K.; Sun, H.; Land, M.; et al. The Complete Genome Sequence of Staphylothermus Marinus Reveals Differences in Sulfur Metabolism among Heterotrophic Crenarchaeota. BMC Genomics 2009, 10. [Google Scholar] [CrossRef] [PubMed]
- Labes, A.; Schonheit, P. Sugar Utilization in the Hyperthermophilic, Sulfate-Reducing Archaeon Archaeoglobus Fulgidus Strain 7324: Starch Degradation to Acetate and CO2 via a Modified Embden-Meyerhof Pathway and Acetyl-Coa Synthetase (Adp-Forming). Arch. Microbiol. 2001, 176, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Mukund, S.; Adams, M.W. Glyceraldehyde-3-Phosphate Ferredoxin Oxidoreductase, a Novel Tungsten-Containing Enzyme with a Potential Glycolytic Role in the Hyperthermophilic Archaeon Pyrococcus furiosus. J. Biol. Chem. 1995, 270, 8389–8392. [Google Scholar] [CrossRef] [PubMed]
- Blamey, J.M.; Adams, M.W.W. Purification and characterization of pyruvate ferredoxin oxidoreductase from the hyperthermophilic archaeon Pyrococcus furiosus. Biochim. Biophys. Acta 1993, 1161, 19–27. [Google Scholar] [CrossRef]
- Ma, K.; Hutchins, A.; Sung, S.J.; Adams, M.W. Pyruvate Ferredoxin Oxidoreductase from the Hyperthermophilic Archaeon, Pyrococcus furiosus, Functions as a Coa-Dependent Pyruvate Decarboxylase. Proc. Natl. Acad. Sci. USA 1997, 94, 9608–9613. [Google Scholar] [CrossRef] [PubMed]
- Musfeldt, M.; Selig, M.; Schonheit, P. Acetyl Coenzyme a Synthetase (ADP Forming) from the Hyperthermophilic Archaeon Pyrococcus furiosus: Identification, Cloning, Separate Expression of the Encoding Genes, Acdai and Acdbi, in Escherichia Coli, and in vitro Reconstitution of the Active Heterotetrameric Enzyme from Its Recombinant Subunits. J. Bacteriol. 1999, 181, 5885–5888. [Google Scholar] [PubMed]
- Glasemacher, J.; Bock, A.K.; Schmid, R.; Schonheit, P. Purification and Properties of Acetyl-Coa Synthetase (Adp-Forming), an Archaeal Enzyme of Acetate Formation and Atp Synthesis, from the Hyperthermophile Pyrococcus furiosus. Eur. J. Biochem. 1997, 244, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Schafer, T.; Selig, M.; Schonheit, P. Acetyl-Coa Synthetase (Adp Forming) in Archaea, a Novel Enzyme Involved in Acetate Formation and Atp Synthesis. Arch. Microbiol. 1993, 159, 72–83. [Google Scholar] [CrossRef]
- Brasen, C.; Schmidt, M.; Grotzinger, J.; Schonheit, P. Reaction Mechanism and Structural Model of ADP-Forming Acetyl-Coa Synthetase from the Hyperthermophilic Archaeon Pyrococcus furiosus: Evidence for a Second Active Site Histidine Residue. J. Biol. Chem. 2008, 283, 15409–15418. [Google Scholar] [CrossRef] [PubMed]
- Lessner, D.J.; Li, L.; Li, Q.; Rejtar, T.; Andreev, V.P.; Reichlen, M.; Hill, K.; Moran, J.J.; Karger, B.L.; Ferry, J.G. An Unconventional Pathway for Reduction of CO2 to Methane in Co-Grown Methanosarcina acetivorans Revealed by Proteomics. Proc. Natl. Acad. Sci. USA 2006, 103, 17921–17926. [Google Scholar] [CrossRef] [PubMed]
- Rother, M.; Oelgeschlager, E.; Metcalf, W.M. Genetic and Proteomic Analyses of Co Utilization by Methanosarcina acetivorans. Arch. Microbiol. 2007, 188, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Ferry, J.G. Fundamentals of Methanogenic Pathways That Are Key to the Biomethanation of Complex Biomass. Curr. Opin. Biotechnol. 2011, 22, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Ferry, J.G. How to Make a Living Exhaling Methane. Annu. Rev. Microbiol. 2010, 64, 453–473. [Google Scholar] [CrossRef] [PubMed]
- Thauer, R.K.; Kaster, A.K.; Seedorf, H.; Buckel, W.; Hedderich, R. Methanogenic Archaea: Ecologically Relevant Differences in Energy Conservation. Nat. Rev. Microbiol. 2008, 6, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Whitman, W.B. Metabolic, Phylogenetic, and Ecological Diversity of the Methanogenic Archaea. Ann. N. Y. Acad. Sci. 2008, 1125, 171–189. [Google Scholar] [CrossRef] [PubMed]
- Welte, C.; Deppenmeier, U. Re-Evaluation of the Function of the F420 Dehydrogenase in Electron Transport of Methanosarcina Mazei. FEBS J. 2011, 278, 1277–1287. [Google Scholar] [CrossRef] [PubMed]
- Vepachedu, V.R.; Ferry, J.G. Role of the Fused Corrinoid/Methyl Transfer Protein Cmta during Co-Dependent Growth of Methanosarcina acetivorans. J. Bacteriol. 2012, 194, 4161–4168. [Google Scholar] [CrossRef] [PubMed]
- Moller-Zinkhan, D.; Thauer, R.K. Anaerobic Lactate Oxidation to 3 CO2 by Archaeoglobus Fulgidus via The Carbon Monoxide Dehydrogenase Pathway. Demonstration of the Acetyl-Coa Carbon-Carbon Cleavage Reaction in Cell Extracts. Arch. Microbiol. 1990, 153, 215–218. [Google Scholar] [CrossRef]
- Gorris, L.; Voet, A.; van der Drift, C. Structural Characteristics of Methanogenic Cofactors in the Non-Methanogenic Archaebacterium Archaeoglobus fulgidus. Biofactors 1991, 3, 29–35. [Google Scholar] [PubMed]
- Schworer, B.; Breitung, J.; Klein, A.R.; Stetter, K.O.; Thauer, R.K. Formylmethanofuran: Tetrahydromethanopterin Formyltransferase and N5,N10-Methylenetetrahydromethanopterin Dehydrogenase from the Sulfate-Reducing Archaeoglobus Fulgidus: Similarities with the Enzymes from Methanogenic Archaea. Arch. Microbiol. 1993, 159, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Kunow, J.; Linder, D.; Stetter, K.O.; Thauer, R.K. F420h2:Quinone Oxidoreductase from Archaeoglobus Fulgidus. Characterization of a Membrane-Bound Multisubunit Complex Containing Fad and Iron-Sulfur Clusters. Eur. J. Biochem. 1994, 223, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.R.; Reed, D.W.; Millstein, J.H.; Hartzell, P.L.; Grahame, D.A.; Demoll, E. Acetyl-Coa Decarbonylase/Synthase Complex from Archaeoglobus fulgidus. Arch. Microbiol. 1998, 169, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Ingram-Smith, C.; Smith, K.S. Amp-Forming Acetyl-Coa Synthetases in Archaea Show Unexpected Diversity in Substrate Utilization. Archaea 2007, 2, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Henstra, A.M.; Dijkema, C.; Stams, A.J. Archaeoglobus Fulgidus Couples CO Oxidation to Sulfate Reduction and Acetogenesis with Transient Formate Accumulation. Environ. Microbiol. 2007, 9, 1836–1841. [Google Scholar] [CrossRef] [PubMed]
- Musfeldt, M.; Schonheit, P. Novel Type of Adp-Forming Acetyl Coenzyme a Synthetase in Hyperthermophilic Archaea: Heterologous Expression and Characterization of Isoenzymes from the Sulfate Reducer Archaeoglobus fulgidus and the Methanogen Methanococcus jannaschii. J. Bacteriol. 2002, 184, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Battistuzzi, F.U.; Feijao, A.; Hedges, S.B. A Genomic Timescale of Prokaryote Evolution: Insights into the Origin of Methanogenesis, Phototrophy, and the Colonization of Land. BMC Evol. Biol. 2004, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, Y.; Yamada, K.; Yoshida, N.; Maruyama, S.; Isozaki, Y. Evidence from Fluid Inclusions for Microbial Methanogenesis in the Early Archaean Era. Nature 2006, 440, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Rothman, D.H.; Fournier, G.P.; French, K.L.; Alm, E.J.; Boyle, E.A.; Cao, C.; Summons, R.E. Methanogenic Burst in the End-Permian Carbon Cycle. Proc. Natl. Acad. Sci. USA. 2014, 111, 5462–5467. [Google Scholar] [CrossRef] [PubMed]
- Buss, K.A.; Cooper, D.R.; Ingram-Smith, C.; Ferry, J.G.; Sanders, D.A.; Hasson, M.S. Urkinase: Structure of Acetate Kinase, a Member of the Askha Superfamily of Phosphotransferases. J. Bacteriol. 2001, 183, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, P.A.; Chang, B. The Evolution of Acetyl-Coa Synthase. Orig. Life Evol. Biosph. 2001, 31, 403–434. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, G.; Thauer, R.K. The Na+ Translocating Methyltransferase Complex from Methanogenic Archaea. Biochim. Biophys. Acta 2001, 1505, 28–36. [Google Scholar] [CrossRef]
- Shima, S.; Warkentin, E.; Thauer, R.K.; Ermler, U. Structure and Function of Enzymes Involved in the Methanogenic Pathway Utilizing Carbon Dioxide and Molecular Hydrogen. J. Biosci. Bioeng. 2002, 93, 519–530. [Google Scholar] [CrossRef]
- Suharti, S.; Wang, M.; de Vries, S.; Ferry, J.G. Characterization of the RNFB and RNFG Subunits of the RNF Complex from the Archaeon Methanosarcina acetivorans. PLoS ONE 2014, 9, E97966. [Google Scholar] [CrossRef] [PubMed]
- Gorrell, A.; Lawrence, S.H.; Ferry, J.G. Structural and Kinetic Analyses of Arginine Residues in the Active-Site of the Acetate Kinase from Methanosarcina thermophila. J. Biol. Chem. 2005, 280, 10731–10742. [Google Scholar] [CrossRef] [PubMed]
- Miles, R.D.; Gorrell, A.; Ferry, J.G. Evidence for a Transition State Analog, Mgadp-Aluminum Fluoride-Acetate, in Acetate Kinase from Methanosarcina thermophila. J. Biol. Chem. 2002, 277, 22547–22552. [Google Scholar] [CrossRef] [PubMed]
- Ingram-Smith, C.; Gorrell, A.; Lawrence, S.H.; Iyer, P.; Smith, K.; Ferry, J.G. Identification of the Acetate Binding Site in the Methanosarcina thermophila Acetate Kinase. J. Bacteriol. 2005, 187, 2386–2394. [Google Scholar] [CrossRef] [PubMed]
- Miles, R.D.; Iyer, P.P.; Ferry, J.G. Site-Directed Mutational Analysis of Active Site Residues in the Acetate Kinase from Methanosarcina thermophila. J. Biol. Chem. 2001, 276, 45059–45064. [Google Scholar] [CrossRef] [PubMed]
- Ingram-Smith, C.; Barber, R.D.; Ferry, J.G. The Role of Histidines in the Acetate Kinase from Methanosarcina thermophila. J. Biol. Chem. 2000, 275, 33765–33770. [Google Scholar] [CrossRef] [PubMed]
- Singh-Wissmann, K.; Miles, R.D.; Ingram-Smith, C.; Ferry, J.G. Identification of Essential Arginines in the Acetate Kinase from Methanosarcina thermophila. Biochemistry 2000, 39, 3671–3677. [Google Scholar] [CrossRef] [PubMed]
- Singh-Wissmann, K.; Ingram-Smith, C.; Miles, R.D.; Ferry, J.G. Identification of Essential Glutamates in the Acetate Kinase From Methanosarcina thermophila. J. Bacteriol. 1998, 180, 1129–1134. [Google Scholar] [PubMed]
- Gorrell, A.; Ferry, J.G. Investigation of the Methanosarcina thermophila Acetate Kinase Mechanism by Fluorescence Quenching. Biochemistry 2007, 46, 14170–14176. [Google Scholar] [CrossRef] [PubMed]
- Ingram-Smith, C.; Wharton, J.; Reinholz, C.; Doucet, T.; Hesler, R.; Smith, K. The Role of Active Site Residues in Atp Binding and Catalysis in the Methanosarcina thermophila Acetate Kinase. Life 2015, 5, 861–871. [Google Scholar] [CrossRef] [PubMed]
- Iyer, P.P.; Lawrence, S.H.; Luther, K.B.; Rajashankar, K.R.; Yennawar, H.P.; Ferry, J.G.; Schindelin, H. Crystal Structure of Phosphotransacetylase from the Methanogenic Archaeon Methanosarcina thermophila. Structure 2004, 12, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Abbanat, D.R.; Ferry, J.G. Synthesis of Acetyl-Coa by the Carbon Monoxide Dehydrogenase Complex from Acetate-Grown Methanosarcina thermophila. J. Bacteriol. 1990, 172, 7145–7150. [Google Scholar] [PubMed]
- Raybuck, S.A.; Ramer, S.E.; Abbanat, D.R.; Peters, J.W.; Orme-Johnson, W.H.; Ferry, J.G.; Walsh, C.T. Demonstration of Carbon-Carbon Bond Cleavage of Acetyl Coenzyme a by Using Isotopic Exchange Catalyzed by the Co Dehydrogenase Complex from Acetate-Grown Methanosarcina thermophila. J. Bacteriol. 1991, 173, 929–932. [Google Scholar] [PubMed]
- Terlesky, K.C.; Nelson, M.J.K.; Ferry, J.G. Isolation of an Enzyme Complex with Carbon Monoxide Dehydrogenase Activity Containing a Corrinoid and Nickel from Acetate-Grown Methanosarcina thermophila. J. Bacteriol. 1986, 168, 1053–1058. [Google Scholar] [PubMed]
- Eggen, R.I.L.; Vankranenburg, R.; Vriesema, A.J.M.; Geerling, A.C.M.; Verhagen, M.F.J.M.; Hagen, W.R.; Devos, W.M. Carbon Monoxide Dehydrogenase from Methanosarcina Frisia Gö1. Characterization of the Enzyme and the Regulated Expression of Two Operon-Like Cdh Gene Clusters. J. Biol. Chem. 1996, 271, 14256–14263. [Google Scholar] [PubMed]
- Jetten, M.S.M.; Hagen, W.R.; Pierik, A.J.; Stams, A.J.M.; Zehnder, A.J.B. Paramagnetic Centers and Acetyl-Coenzyme a CO Exchange Activity of Carbon Monoxide Dehydrogenase from Methanothrix soehngenii. Eur. J. Biochem. 1991, 195, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Grahame, D.A. Substrate and Cofactor Reactivity of A Carbon Monoxide Dehydrogenase Corrinoid Enzyme Complex. Stepwise Reduction of Iron Sulfur and Corrinoid Centers, the Corrinoid Co2+/1+ Redox Midpoint Potential, and Overall Synthesis of Acetyl-Coa. Biochemistry 1993, 32, 10786–10793. [Google Scholar] [CrossRef] [PubMed]
- Grahame, D.A.; Demoll, E. Substrate and Accessory Protein Requirements and Thermodynamics of Acetyl-Coa Synthesis and Cleavage in Methanosarcina barkeri. Biochemistry 1995, 34, 4617–4624. [Google Scholar] [CrossRef] [PubMed]
- Jetten, M.S.M.; Stams, A.J.M.; Zehnder, A.J.B. Purification and Characterization of an Oxygen-Stable Carbon Monoxide Dehydrogenase of Methanothrix soehngenii. FEBS Lett. 1989, 181, 437–441. [Google Scholar] [CrossRef]
- Eggen, R.I.L.; Geerling, A.C.M.; Jetten, M.S.M.; Devos, W.M. Cloning, Expression, and Sequence Analysis of the Genes for Carbon Monoxide Dehydrogenase of Methanothrix soehngenii. J. Biol. Chem. 1991, 266, 6883–6887. [Google Scholar] [PubMed]
- Jetten, M.S.M.; Pierik, A.J.; Hagen, W.R. Epr Characterization of a High-Spin System in Carbon Monoxide Dehydrogenase from Methanothrix soehngenii. Eur. J. Biochem. 1991, 202, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Terlesky, K.C.; Ferry, J.G. Ferredoxin Requirement for Electron Transport from the Carbon Monoxide Dehydrogenase Complex to a Membrane-Bound Hydrogenase in Acetate-Grown Methanosarcina thermophila. J. Biol. Chem. 1988, 263, 4075–4079. [Google Scholar] [PubMed]
- Terlesky, K.C.; Ferry, J.G. Purification and Characterization of a Ferredoxin from Acetate-Grown Methanosarcina thermophila. J. Biol. Chem. 1988, 263, 4080–4082. [Google Scholar] [PubMed]
- Fischer, R.; Thauer, R.K. Ferredoxin-Dependent Methane Formation from Acetate in Cell Extracts of Methanosarcina barkeri (Strain Ms). FEBS Lett. 1990, 269, 368–372. [Google Scholar] [CrossRef]
- Ferry, J.G.; House, C.H. The Stepwise Evolution of Early Life Driven by Energy Conservation. Mol. Biol. Evol. 2006, 23, 1286–1292. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Russell, M.J. On the Origin of Biochemistry at an Alkaline Hydrothermal Vent. Philos. Trans. R. Soc. B 2006, 362, 1887–1926. [Google Scholar] [CrossRef] [PubMed]
- Abbanat, D.R.; Ferry, J.G. Resolution of Component Proteins in an Enzyme Complex from Methanosarcina thermophila Catalyzing the Synthesis or Cleavage of Acetyl-Coa. Proc. Natl. Acad. Sci. USA 1991, 88, 3272–3276. [Google Scholar] [CrossRef] [PubMed]
- Clements, A.P.; Ferry, J.G. Cloning, Nucleotide Sequence, and Transcriptional Analyses of the Gene Encoding a Ferredoxin from Methanosarcina thermophila. J. Bacteriol. 1992, 174, 5244–5250. [Google Scholar] [PubMed]
- Gong, W.; Hao, B.; Wei, Z.; Ferguson, D.J., Jr.; Tallant, T.; Krzycki, J.A.; Chan, M.K. Structure of the A2e2 Ni-Dependent Co Dehydrogenase Component of the Methanosarcina barkeri Acetyl-Coa Decarbonylase/Synthase Complex. Proc. Natl. Acad. Sci. USA 2008, 105, 9558–9563. [Google Scholar] [CrossRef] [PubMed]
- Murakami, E.; Ragsdale, S.W. Evidence for Intersubunit Communication during Acetyl-Coa Cleavage by the Multienzyme Co Dehydrogenase/Acetyl-Coa Synthase Complex from Methanosarcina thermophila Evidence That the Beta Subunit Catalyzes C-C and C-S Bond Cleavage. J. Biol. Chem. 2000, 275, 4699–4707. [Google Scholar] [CrossRef] [PubMed]
- Grahame, D.A.; Demoll, E. Partial Reactions Catalyzed by Protein Components of the Acetyl-Coa Decarbonylase Synthase Enzyme Complex from Methanosarcina barkeri. J. Biol. Chem. 1996, 271, 8352–8358. [Google Scholar] [PubMed]
- Funk, T.; Gu, W.W.; Friedrich, S.; Wang, H.X.; Gencic, S.; Grahame, D.A.; Cramer, S.P. Chemically Distinct Ni Sites in the A-Cluster in Subunit Beta of the Acetyl-Coa Decarbonylase/Synthase Complex from Methanosarcina thermophila: Ni L-Edge Absorption And X-Ray Magnetic Circular Dichroism Analyses. J. Am. Chem. Soc. 2004, 126, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Gencic, S.; Grahame, D.A. Nickel in Subunit B of the Acetyl-Coa Decarbonylase/Synthase Multienzyme Complex in Methanogens. J. Biol. Chem. 2003, 278, 6101–6110. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.W.; Gencic, S.; Cramer, S.P.; Grahame, D.A. The A-Cluster in Subunit Beta of the Acetyl-Coa Decarbonylase/Synthase Complex from Methanosarcina thermophila: Ni and Fe K-Edge Xanes and Exafs Analyses. J. Am. Chem. Soc. 2003, 125, 15343–15351. [Google Scholar] [CrossRef] [PubMed]
- Ragsdale, S.W. Nickel and the Carbon Cycle. J. Inorg. Biochem. 2007, 101, 1657–1666. [Google Scholar] [CrossRef] [PubMed]
- Gencic, S.; Grahame, D.A. Two Separate One-Electron Steps in the Reductive Activation of the a Cluster in Subunit Beta of the Acds Complex in Methanosarcina thermophila. Biochemistry 2008, 47, 5544–5455. [Google Scholar] [CrossRef] [PubMed]
- Gencic, S.; Duin, E.C.; Grahame, D.A. Tight Coupling of Partial Reactions in the Acetyl-Coa Decarbonylase/Synthase (Acds) Multienzyme Complex from Methanosarcina thermophila: Acetyl C-C Bond Fragmentation at the a Cluster Promoted by Protein Conformational Changes. J. Biol. Chem. 2010, 285, 15450–15463. [Google Scholar] [CrossRef] [PubMed]
- Gencic, S.; Kelly, K.; Ghebreamlak, S.; Duin, E.C.; Grahame, D.A. Different Modes of Carbon Monoxide Binding to Acetyl-Coa Synthase and the Role of a Conserved Phenylalanine in the Coordination Environment of Nickel. Biochemistry 2013, 52, 1705–1716. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, P.E.; Lu, W.P.; Ragsdale, S.W.; Ferry, J.G. Characterization of the Metal Centers of the Corrinoid/Iron- Sulfur Component of the Co Dehydrogenase Enzyme Complex from Methanosarcina thermophila by Epr Spectroscopy and Spectroelectrochemistry. J. Biol. Chem. 1993, 268, 325–329. [Google Scholar] [PubMed]
- Grahame, D.A. Catalysis of Acetyl-Coa Cleavage and Tetrahydrosarcinapterin Methylation by a Carbon Monoxide Dehydrogenase-Corrinoid Enzyme Complex. J. Biol. Chem. 1991, 266, 22227–22233. [Google Scholar] [PubMed]
- Maupin-Furlow, J.; Ferry, J.G. Characterization of the Cdhd and Cdhe Genes Encoding Subunits of the Corrinoid Iron-Sulfur Enzyme of the Co Dehydrogenase Complex from Methanosarcina thermophila. J. Bacteriol. 1996, 178, 340–346. [Google Scholar] [PubMed]
- Liu, J.S.; Marison, I.W.; von Stockar, U. Microbial Growth by a Net Heat Up-Take: A Calorimetric and Thermodynamic Study on Acetotrophic Methanogenesis by Methanosarcina barkeri. Biotechnol. Bioeng. 2001, 75, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Alber, B.E.; Ferry, J.G. A Carbonic Anhydrase from the Archaeon Methanosarcina thermophila. Proc. Natl. Acad. Sci. USA 1994, 91, 6909–6913. [Google Scholar] [CrossRef] [PubMed]
- Macauley, S.R.; Zimmerman, S.A.; Apolinario, E.E.; Evilia, C.; Hou, Y.; Ferry, J.G.; Sowers, K.R. The Archetype G-Class Carbonic Anhydrase (Cam) Contains Iron when Synthesized in vivo. Biochemistry 2009, 48, 817–819. [Google Scholar] [CrossRef] [PubMed]
- Tripp, B.C.; Bell, C.B.; Cruz, F.; Krebs, C.; Ferry, J.G. A Role for Iron in an Ancient Carbonic Anhydrase. J. Biol. Chem. 2004, 279, 6683–6687. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, S.A.; Ferry, J.G. Proposal for a Hydrogen Bond Network in the Active Site of the Prototypic G-Class Carbonic Anhydrase. Biochemistry 2006, 45, 5149–5157. [Google Scholar] [CrossRef] [PubMed]
- Hovey, R.; Lentes, S.; Ehrenreich, A.; Salmon, K.; Saba, K.; Gottschalk, G.; Gunsalus, R.P.; Deppenmeier, U. DNA Microarray Analysis of Methanosarcina Mazei Go1 Reveals Adaptation to Different Methanogenic Substrates. Mol. Genet. Genomics 2005, 273, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, Q.; Rohlin, L.; Kim, U.; Salmon, K.; Rejtar, T.; Gunsalus, R.P.; Karger, B.L.; Ferry, J.G. Quantitative Proteomic and Microarray Analysis of the Archaeon Methanosarcina acetivorans Grown With Acetate versus Methanol. J. Proteome Res. 2007, 6, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, L.; Rejtar, T.; Lessner, D.J.; Karger, B.L.; Ferry, J.G. Electron Transport in the Pathway of Acetate Conversion to Methane in the Marine Archaeon Methanosarcina acetivorans. J. Bacteriol. 2006, 188, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Pisa, K.Y.; Weidner, C.; Maischak, H.; Kavermann, H.; Muller, V. The Coupling Ion in the Methanoarchaeal ATP Synthases: H+ vs. Na+ in the A1A0 ATP Synthase from the Archaeon Methanosarcina mazei Gö1. FEMS Microbiol. Lett. 2007, 277, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Hedderich, R. Energy-Converting [Nife] Hydrogenases from Archaea and Extremophiles: Ancestors of Complex I. J. Bioenerg. Biomembr. 2004, 36, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Meuer, J.; Kuettner, H.C.; Zhang, J.K.; Hedderich, R.; Metcalf, W.W. Genetic Analysis of the Archaeon Methanosarcina barkeri Fusaro Reveals a Central Role for ECH Hydrogenase and Ferredoxin in Methanogenesis and Carbon Fixation. Proc. Natl. Acad. Sci. USA 2002, 99, 5632–5637. [Google Scholar] [CrossRef] [PubMed]
- Heiden, S.; Hedderich, R.; Setzke, E.; Thauer, R.K. Purification of a Cytochrome-B Containing H2-Heterodisulfide Oxidoreductase Complex from Membranes of Methanosarcina barkeri. Eur. J. Biochem. 1993, 213, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Deppenmeier, U. The Membrane-Bound Electron Transport System of Methanosarcina Species. J. Bioenerg. Biomembr. 2004, 36, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, S.; Ueno, A.; Naganuma, T.; Kaneko, K. Methanosarcina subterranea sp. nov., a methanogenic archaeon isolated from a deep subsurface diatomaceous shale formation of northernmost japan. Int. J. Syst. Evol. Microbiol. 2015, 65, 1167–1171. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.; Schirmack, J.; Ganzert, L.; Morozova, D.; Mangelsdorf, K. Methanosarcina soligelidi sp. nov., a desiccation and freeze-thaw resistant methanogenic archaeon isolated from a siberian permafrost-affected soil. Int. J. Syst. Evol. Microbiol. 2013, 64, 3478–3484. [Google Scholar]
- Von Klein, D.; Arab, H.; Volker, H.; Thomm, M. Methanosarcina baltica, sp. nov., a novel methanogen isolated from the Gotland Deep of the Baltic Sea. Extremophiles 2002, 6, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Blotevogel, K.H.; Fischer, U. Transfer of Methanococcus Frisius to the Genus Methanosarcina as Methanosarcina Frisia Comb. Nov. Int. J. Syst. Bacteriol. 1989, 39, 91–92. [Google Scholar] [CrossRef]
- Zinder, S.H.; Sowers, K.R.; Ferry, J.G. Methanosarcina thermophila sp. nov., a Thermophilic, Acetotrophic, Methane-Producing Bacterium. Int. J. Syst. Bacteriol. 1985, 35, 522–523. [Google Scholar] [CrossRef]
- Shimizu, S.; Upadhye, R.; Ishijima, Y.; Naganuma, T. Methanosarcina horonobensis sp. nov., a methanogenic archaeon isolated from a deep subsurface miocene formation. Int. J. Syst. Evol. Microbiol. 2011, 61, 2503–2507. [Google Scholar] [CrossRef] [PubMed]
- Lyimo, T.J.; Pol, A.; den Camp, H.J.O.; Harhangi, H.R.; Vogels, G.D. Methanosarcina semesiae sp. nov., a dimethylsulfide-utilizing methanogen from mangrove sediment. Int. J. Syst. Evol. Microbiol. 2000, 50, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Cabrol, N.; Villemur, R.; Perrier, J.; Jacob, F.; Foullet, B.; Chambon, P. Isolation of a methanogenic bacterium, Methanosarcina sp. strain FR, for its abiilty to degrade high concentrations of perchloroethylene. Can. J. Microbiol. 1998, 44, 1142–1147. [Google Scholar] [CrossRef]
- Zhilina, T.N.; Zavarzin, G.A. Comparative cytology of Methanosarcinae and description of Methanosarcina vacuolata sp. nova. Microbiology 1979, 48, 223–228. [Google Scholar]
- Sowers, K.R.; Baron, S.F.; Ferry, J.G. Methanosarcina acetivorans sp. nov., An Acetotrophic Methane-Producing Bacterium Isolated from Marine Sediments. Appl. Environ. Microbiol. 1984, 47, 971–978. [Google Scholar] [PubMed]
- Ganzert, L.; Schirmack, J.; Alawi, M.; Mangelsdorf, K.; Sand, W.; Hillebrand-Voiculescu, A.; Wagner, D. Methanosarcina spelaei sp. nov., a Methanogenic Archaeon Isolated from a Floating Biofilm of a Subsurface Sulphurous Lake. Int. J. Syst. Evol. Microbiol. 2014, 64, 3478–3484. [Google Scholar] [CrossRef] [PubMed]
- Simankova, M.V.; Parshina, S.N.; Tourova, T.P.; Kolganova, T.V.; Zehnder, A.J.; Nozhevnikova, A.N. Methanosarcina lacustris sp. nov., a New Psychrotolerant Methanogenic Archaeon from Anoxic Lake Sediments. Syst. Appl. Microbiol. 2001, 24, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, L.; Rejtar, T.; Lessner, D.J.; Karger, B.L.; Ferry, J.G. Electron Transport in the Pathway of Acetate Conversion to Methane in the Marine Archaeon Methanosarcina acetivorans. J. Bacteriol. 2006, 188, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Tomb, J.F.; Ferry, J.G. Electron Transport in Acetate-Grown Methanosarcina acetivorans. BMC Microbiol. 2011, 11. [Google Scholar] [CrossRef] [PubMed]
- Hreha, T.N.; Mezic, K.G.; Herce, H.D.; Duffy, E.D.; Bourges, A.; Pryshchep, S.; Juarez, O.; Barquera, B. The Complete Topology of the RNF Complex from Vibrio Cholerae. Biochemistry 2015, 54, 2443–2455. [Google Scholar] [CrossRef] [PubMed]
- Saeki, K.; Kumagai, H. The Rnf Gene Products in Rhodobacter Capsulatus Play an Essential Role in Nitrogen Fixation during Anaerobic Dmso-Dependent Growth in the Dark. Arch. Microbiol. 1998, 169, 464–467. [Google Scholar] [CrossRef] [PubMed]
- Muller, V.; Imkamp, F.; Biegel, E.; Schmidt, S.; Dilling, S. Discovery of a Ferredoxin: Nad+-Oxidoreductase (RNF) in Acetobacterium Woodii: A Novel Potential Coupling Site in Acetogens. Ann. N. Y. Acad. Sci. 2008, 1125, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Seedorf, H.; Fricke, W.F.; Veith, B.; Bruggemann, H.; Liesegang, H.; Strittmatter, A.; Miethke, M.; Buckel, W.; Hinderberger, J.; Li, F.; et al. The Genome of Clostridium kluyveri, a Strict Anaerobe with Unique Metabolic Features. Proc. Natl. Acad. Sci. USA 2008, 105, 2128–2133. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Kohler, J.; Hurek, T.; Reinhold-Hurek, B. A Novel Regulatory Role of the RNF Complex of Azoarcus SP. Strain Bh72. Mol. Microbiol. 2011, 83, 408–422. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, P.L.; Zhang, T.; Dar, S.A.; Leang, C.; Lovley, D.R. The RNF Complex of Clostridium Ljungdahlii Is a Proton-Translocating Ferredoxin: Nad+ Oxidoreductase Essential for Autotrophic Growth. mBio 2012, 4. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.; Kuehl, J.V.; Price, M.N.; Ray, J.; Deutschbauer, A.M.; Arkin, A.P.; Stahl, D.A. The Energy-Conserving Electron Transfer System Used by Desulfovibrio alaskensis Strain G20 During Pyruvate Fermentation Involves Reduction of Endogenously Formed Fumarate and cytoplasmic and membrane-bound complexes, Hdr-Flox and RNF. Environ. Microbiol. 2014, 16, 3463–3486. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, K.; Welte, C.; Deppenmeier, U.; Muller, V. Electron Transport During Aceticlastic Methanogenesis by Methanosarcina acetivorans Involves a Sodium-Translocating RNF Complex. FEBS J. 2012, 279, 4444–4452. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, K.; Leone, V.; Faraldo-Gomez, J.D.; Muller, V. Promiscuous Archaeal ATP Synthase Concurrently Coupled to Na+ And H+ Translocation. Proc. Natl. Acad. Sci. USA 2012, 109, 947–952. [Google Scholar] [CrossRef] [PubMed]
- Jasso-Chavez, R.; Apolinario, E.E.; Sowers, K.R.; Ferry, J.G. MRPA Functions in Energy Conversion during Acetate-Dependent Growth of Methanosarcina acetivorans. J. Bacteriol. 2013, 195, 3987–3994. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.S.; Ingram-Smith, C. Methanosaeta, the Forgotten Methanogen? Trends Microbiol. 2007, 7, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.; Welte, C.; Deppenmeier, U. Acetate Activation in Methanosaeta Thermophila: Characterization of the Key Enzymes Pyrophosphatase and Acetyl-Coa Synthetase. Archaea 2012, 2012, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jetten, M.S.M.; Stams, A.J.M.; Zehnder, A.J.B. Isolation and Characterization of Acetyl-Coenzyme a Synthetase from Methanothrix soehngenii. J. Bacteriol. 1989, 171, 5430–5435. [Google Scholar] [PubMed]
- Eggen, R.I.L.; Geerling, A.C.M.; Boshoven, A.B.P.; Devos, W.M. Cloning, Sequence Analysis, and Functional Expression of the Acetyl Coenzyme a Synthetase Gene from Methanothrix soehngenii in Escherichia Coli. J. Bacteriol. 1991, 173, 6383–6389. [Google Scholar] [PubMed]
- Aceti, D.J.; Ferry, J.G. Purification and Characterization of Acetate Kinase from Acetate-Grown Methanosarcina thermophila. J. Biol. Chem. 1988, 263, 15444–15448. [Google Scholar] [PubMed]
- Tor, J.M.; Kashefi, K.; Lovley, D.R. Acetate Oxidation Coupled to Fe(III) Reduction in Hyperthermophilic Microorganisms. Appl. Environ. Microbiol. 2001, 67, 1363–1365. [Google Scholar] [CrossRef] [PubMed]
- Slobodkina, G.B.; Kolganova, T.V.; Querellou, J.; Bonch-Osmolovskaya, E.A.; Slobodkin, A.I. Geoglobus Acetivorans sp. nov., an Iron(III)-Reducing Archaeon from a Deep-Sea Hydrothermal Vent. Int. J. Syst. Evol. Microbiol. 2009, 59, 2880–2883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slobodkin, A.; Campbell, B.; Cary, S.C.; Bonch-Osmolovskaya, E.; Jeanthon, C. Evidence for the Presence of Thermophilic Fe(III)-Reducing Microorganisms in Deep-Sea Hydrothermal Vents at 13 Degrees N (East Pacific Rise). FEMS Microbiol. Ecol. 2001, 36, 235–243. [Google Scholar] [PubMed]
- Seyler, L.M.; Mcguinness, L.M.; Kerkhof, L.J. Crenarchaeal Heterotrophy in Salt Marsh Sediments. ISME J. 2014, 8, 1534–1543. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ferry, J.G. Acetate Metabolism in Anaerobes from the Domain Archaea. Life 2015, 5, 1454-1471. https://doi.org/10.3390/life5021454
AMA Style
Ferry JG. Acetate Metabolism in Anaerobes from the Domain Archaea. Life. 2015; 5(2):1454-1471. https://doi.org/10.3390/life5021454
Chicago/Turabian StyleFerry, James G. 2015. "Acetate Metabolism in Anaerobes from the Domain Archaea" Life 5, no. 2: 1454-1471. https://doi.org/10.3390/life5021454