Integrating Dendrochronology, Climate and Satellite Remote Sensing to Better Understand Savanna Landscape Dynamics in the Okavango Delta, Botswana



Abstract
:1. Introduction
1.1 Study Area

2. Materials and Methods
2.1. Precipitation Data
2.2. Regional Dendrochronology


| Species | Common Name | n | Avg. DBH (cm) | Measurable Ring Structure |
|---|---|---|---|---|
| Acacia tortilis | Umbrella thorn | 3 | 20.7 | no (rings inconsistently distinct so not measureable**) |
| Colophospermum mopane | Mopane | 3 | 41.1 | no |
| Pterocarpus angolensis | Mokwa | 3 | 45 | yes |
| Brachystegia sp. | 2 | 33.7 | no (rings inconsistently distinct so not measureable) | |
| Lonchocarpus nelsii | Kalahari apple leaf | 2 | 29.2 | no |
| Acacia nigrescens | knobthorn | 1 | - | no core extracted—too dense |
| Baikiaea plurijuga | Rhodesian teak | 1 | 43.4 | no |
| Boscia albitrunca | Shepherd’s tree | 1 | 41.9 | no |
| Burkea africana | Wild Seringa | 1 | - | yes |
| Croton magalobotrys | Large Fever-berry | 1 | 41.5 | no |
| Lonchocarpus capassa | Raintree | 1 | >50 | no |

| Species | Location | Correlated Climate Variable (Ring Widths and Climate) | Longest Chronology (Years) | Chronology Sample Depth (#Stems) | Authors |
|---|---|---|---|---|---|
| Pterocarpus angolensis | Zimbabwe | Summer rainfall; ENSO; Decadal droughts | 201 | 36 | Therrell et al. 2006 [31] |
| Zimbabwe | Rainfall (+) | 120 | 14 to 39 | Stahle et al. 1999 [30] | |
| Namibia | Rainfall (+); Temperature (−); ENSO | 79 | 4 to 5 | Fichtler et al. 2004 [28] | |
| Burkea africana | Namibia | Rainfall (+); Temperature (−); ENSO | 110 | 5 to 12 | Fichtler et al. 2004 [28] |
| Brachystegia spiciformis | Namibia, Zambia, Tanzania | Rainfall (+); ENSO (−) | 149 | 9 to 34 | Trouet et al. 2010 [34] |
| Zambia | December & January rainfall (+); Temperature (−) | 63 | 13 to 16 | Trouet et al. 2006 [33] | |
| Isoberlinia tomentosa | Tanzania | Rainfall; Temperature SOI-value | 38 | 8 | Trouet et al. 2001 [32] |
2.3. CASA Model to Derive NPP
2.3.1. Remote Sensing Data
2.3.2. Climate Data
2.3.3. Land Cover and Soil Data
2.3.4. CASA Model Description and Evaluation
3. Results and Discussion
3.1. Precipitation Change

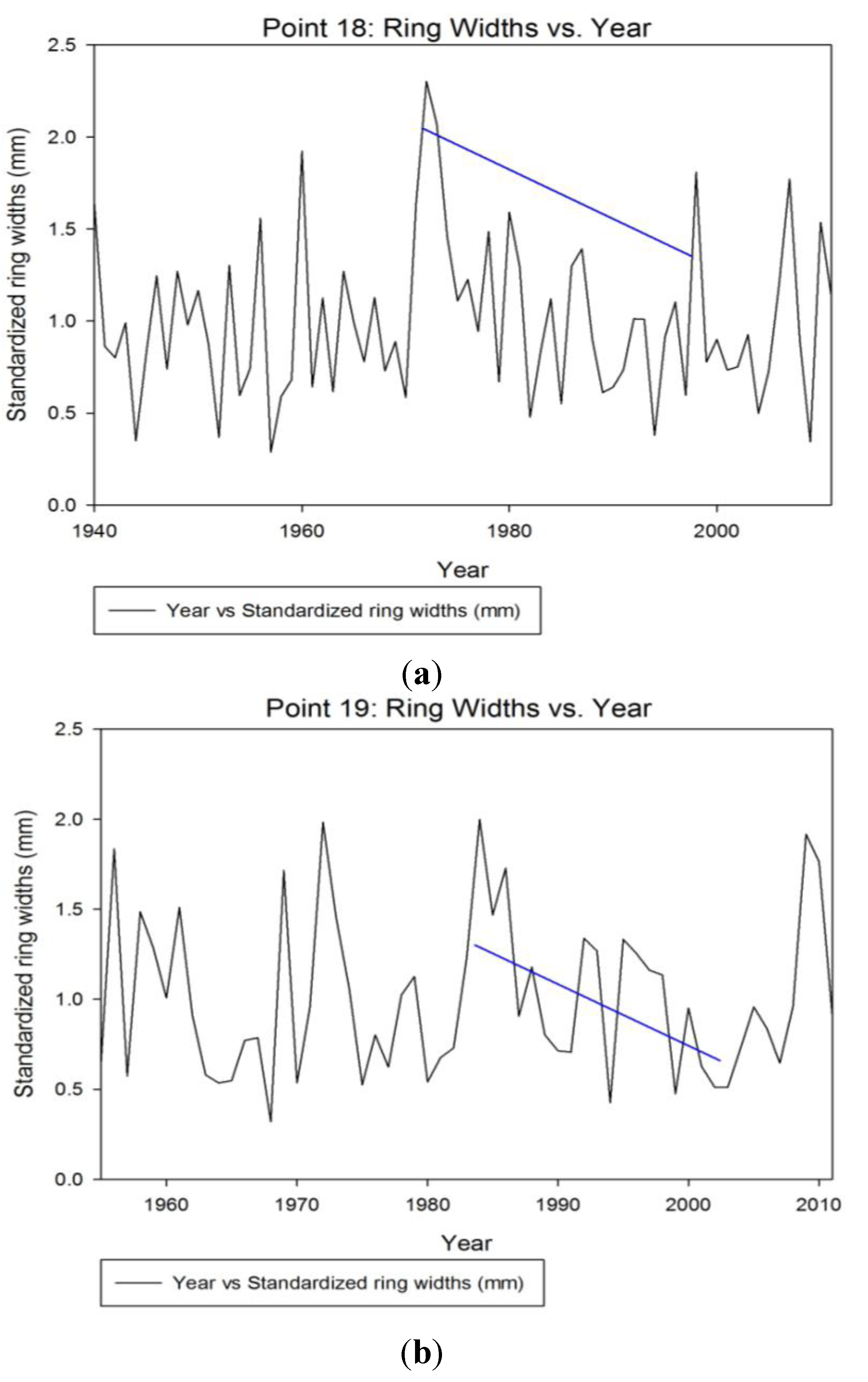
3.2. Dendrochronology
3.2.1. Comparison to the Literature and to Available Published Tree Core data

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3.2.2. Tree Ring Analysis from Sampled Cores

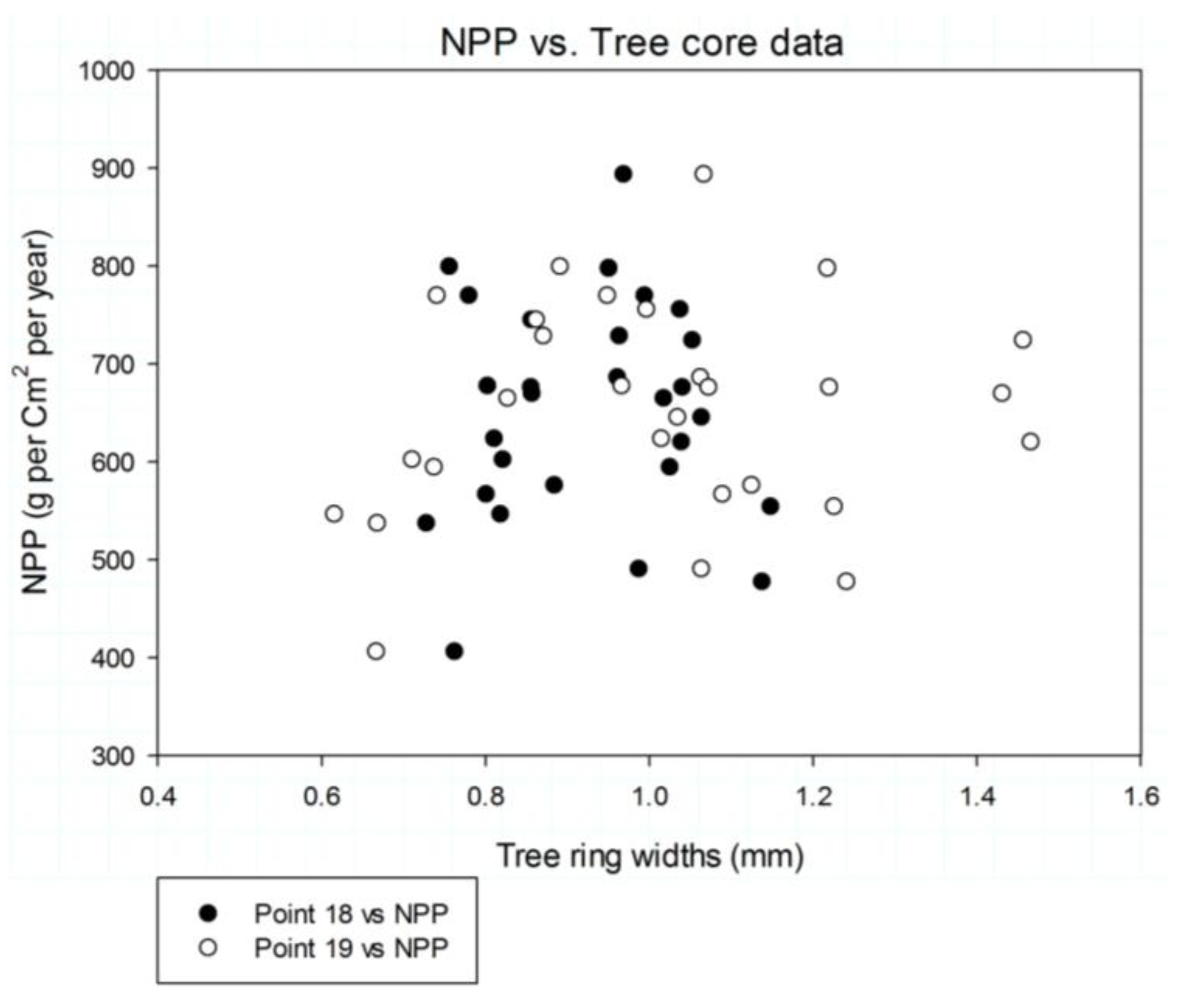
3.3. NPP Analysis


4. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Intergovernmental Panel on Climate Change (CCI), The Physical Science Basis, Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007.
- Davis, M.B. Climate Change and the Survival of Forest Species. In The Earth in Transition: Patterns and Processes of Biotic Impoverishment; Woodwell, G.M., Ed.; Cambridge University Press: Cambridge, UK, 1990; pp. 99–110. [Google Scholar]
- Malcolm, J.R.; Markham, A.; Neilson, R.P.; Garaci, M. Estimated migration rates under scenarios of global climate change. J. Biogeogr. 2002, 29, 835–849. [Google Scholar] [CrossRef]
- Houghton, J. Global Warming: The Complete Briefing; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Delcourt, P.A.; Delcourt, H.R. Late-quaternary dynamics of temperate forests: Applications of paleoecology to issues of global environmental change. Quat. Sci. Rev. 1987, 6, 129–146. [Google Scholar]
- Davis, M.B.; Shaw, R.G. Range shifts and adaptive responses to quaternary climate change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef]
- Scott, L. Late quaternary forest history in Venda, Southern Africa. Rev. Paleobot. Palynol. 1987, 53, 1–10. [Google Scholar] [CrossRef]
- Scott, L. Climatic conditions in Southern Africa since the last glacial maximum, inferred from pollen analysis. Paleogeogr. Paleoclim. Paleoecol. 1989, 70, 345–353. [Google Scholar] [CrossRef]
- Dupont, L.M.; Caley, T.; Kim, J-H.; Castaneda, I.; Malaize, B.; Giraudeau, J. Glacial-interglacial vegetation dynamics in South Eastern Africa depend on sea surface temperature variations in the West Indian Ocean. Clim. Past Discuss. 2011, 7, 2261–2296. [Google Scholar] [CrossRef]
- Ekblom, A. Forest-savanna dynamics in the coastal lowland of southern Mozambique since C. AD 1400. The Holocene 2008, 18, 1247–1257. [Google Scholar] [CrossRef]
- Scott, L.; Bousman, C.B. Palynological analysis of hyrax middens from Southern Africa. Paleogeogr. Paleoclim. Paleoecol. 1990, 76, 367–379. [Google Scholar] [CrossRef]
- Kambatuku, J.R.; Cramer, M.D.; Ward, D. Intraspecific competition between shrubs in a semi-arid savanna. Plant Ecol. 2011, 212, 1701–1713. [Google Scholar]
- Scholes, R.; Walker, B. An African Savanna: Synthesis of the Nylsvley Study; Cambridge University Press: New York, NY, USA, 1993. [Google Scholar]
- Hanan, N.; Lehmann, C. Tree-Grass. Interactions in Savannas: Paradigms, Contradictions, and Conceptual Models. In Ecosystem Function in Savannas; Hill, M., Hanan, N., Eds.; Taylor and Francis Group: Boca Raton, FL, USA, 2010; pp. 39–53. [Google Scholar]
- Ellis, J.E.; Swift, D.M. Stability of African pastoral ecosystems: Alternate paradigms and implications for development. J. Range Manag. 1988, 41, 450–459. [Google Scholar] [CrossRef]
- Zen, N.; Neelin, D. The role of vegetation-climate interaction and interannual variability in shaping the African savanna. J. Climate 2000, 13, 2665–2670. [Google Scholar] [CrossRef]
- Meyer, K.M.; Wiegand, K.; Ward, D.; Moustakas, A. The rhythm of savanna patch dynamics. J. Ecol. 2007, 95, 1306–1315. [Google Scholar] [CrossRef]
- Gillson, L.; Hoffman, M.T. Rangeland ecology in a changing world. Science 2007, 315, 53–54. [Google Scholar] [CrossRef]
- Blaikie, P.; Brookfield, H. Land Degradation and Society; Methuen: London, UK, 1987. [Google Scholar]
- Johnson, D.L.; Lewis, L. Land Degradation: Creation and Destruction; Blackwell: Oxford, UK, 2004. [Google Scholar]
- Thomas, D.S.G.; Twyman, C. Good or bad rangeland? Hybrid knowledge, science and local understandings of vegetation dynamics in the Kalahari. Land Degrad. Dev. 2004, 15, 215–231. [Google Scholar] [CrossRef]
- Ringrose, S.; Matbeson, W.; Tempest, F.; Boyle, T. The development and causes of range degradation features in south-east Botswana using multi-temporal Landsat MSS imagery. Photogramm. Eng. Remote Sens. 1990, 56, 1253–1262. [Google Scholar]
- Barnes, R.F.W. The effect of elephant browsing on woodlands in a Tanzanian National Park: Measurements, models and management. J. Appl. Ecol. 1983, 20, 521–539. [Google Scholar] [CrossRef]
- Baxter, P.W.J.; Getz, W.M. A model-framed evaluation of elephant effects on tree and fire dynamics in African Savannas. Ecol. Appl. 2005, 15, 1331–1341. [Google Scholar] [CrossRef]
- Ntumi, C.P.; van Varde, R.J.; Fairall, N.; de Boer, W.F. Use of space and habitat by elephants in the Maputo Elephant Reserve, Mozambique. South. Afr. J. Wildl. Res. 2005, 35, 139–146. [Google Scholar]
- Campo-Bescos, M.; Munoz-Carpena, R.; Kaplan, D.; Southworth, J.; Zhu, L.; Waylen, P. Beyond precipitation: Physiographic gradients dictate the relative importance of environmental drivers on savanna vegetation. PLOS One 2013, 8, e72348. [Google Scholar]
- Sankaran, M.; Hanan, N.; Scholes, R.J.; Ratnam, J.; Augustine, D.J.; Cade, B.S.; Gignoux, J.; Higgins, S.I.; Roux, X.L.; Ludwig, F.; et al. Determinants of woody cover in African savannas. Nature 2005, 438, 1038. [Google Scholar]
- Fichtler, E.; Trouet, V.; Beeckman, H.; Coppin, P.; Worbes, M. Climatic signals in tree rings of Burkea Africana. Trees 2004, 18, 442–451. [Google Scholar]
- Rozendaal, D.M.A.; Zuidema, P.A. Dendroecology in the tropics: A review. Trees 2010, 25, 3–16. [Google Scholar] [CrossRef]
- Stahle, D.W.; Mushove, P.T.; Cleaveland, M.K.; Roig, F.; Haynes, G.A. Management implications of annual growth rings in Pterocarpus angolensis from Zimbabwe. Forest Ecol. Manag. 1999, 124, 217–229. [Google Scholar] [CrossRef]
- Therrell, M.D.; Stahle, D.W.; Ries, L.P.; Shugart, H.H. Tree-ring reconstructed rainfall variability in Zimbabwe. Clim. Dyn. 2006, 26, 677–685. [Google Scholar] [CrossRef]
- Trouet, V.; Haneca, K.; Coppin, P.; Beeckman, H. Tree ring analysis of Brachystegia spiciformis and Isoberlinia tomentosa: Evaluation of the ENSO-signal in the Miombo woodland of Eastern Africa. IAWA J. 2001, 22, 385–399. [Google Scholar] [CrossRef]
- Trouet, V.; Coppin, P.; Beeckman, H. Annual growth ring patterns in Brachystegia spiciformis reveal influence of precipitation on tree growth. Biotropica 2006, 38, 375–382. [Google Scholar] [CrossRef]
- Trouet, V.; Esper, J.; Beeckman, H. Climate/growth relationships of Brachystegia spiciformis from the miombo woodland in south central Africa. Dendrochronologia 2010, 28, 161–171. [Google Scholar] [CrossRef]
- Gieske, A. Modelling outflow from the Jao/Boro River system in the Okavango Delta, Botswana. J. Hydrol. 1997, 193, 214–239. [Google Scholar] [CrossRef]
- McCarthy, T.S. The great inland deltas of Africa. J. Afr. Earth Sci. 1993, 17, 275–291. [Google Scholar] [CrossRef]
- Ellery, K.; Ellery, W.N.; Rogers, K.H.; Walker, B.H. Water depth and biotic insulation: major determinants of backswamp plant community composition. Wetl. Ecol. Manag. 1991, 1, 149–162. [Google Scholar]
- Hare, S.R.; Mantua, N.J. Empirical evidence for North Pacific regime shifts in 1977 and 1989. Prog. Oceanogr. 2000, 47, 1030–1045. [Google Scholar]
- Mason, S. El Niño, climate change and Southern African climate. Environmetrics 2001, 12, 327–345. [Google Scholar] [CrossRef]
- Chavez, F.P.; Ryan, J.; Lluta-Cota, S.E.; Ñiquen, M. From anchovies to sardines and back: Multidecadal change in the Pacific Ocean. Science 2003, 299, 217–221. [Google Scholar]
- Shi, G.; Ribbe, J.; Cai, W.; Cowan, T. Multidecadal variability in the transmission of ENSO signals to the Indian Ocean. Geophys. Res. Lett. 2007, 34, L09706. [Google Scholar] [CrossRef]
- Manatsa, D.; Matarira, C.H.; Mukwada, G. Relative Impacts of ENSO and Indian Ocean dipol/zonal mode on east SADC rainfall. Int. J. Climatol. 2011, 31, 558–577. [Google Scholar] [CrossRef]
- Ropelewski, C.F.; Halpert, M.S. Halpert global and regional scale precipitation patterns associated with the El Niño/Southern Oscillation. Mon. Weather Rev. 1987, 115, 1606–1626. [Google Scholar] [CrossRef]
- Waylen, P.; Henworth, S. A note on the timing of precipitation variability in Zimbabwe as related to the Southern Oscillation. Int. J. Climatol. 1996, 16, 1137–1148. [Google Scholar] [CrossRef]
- Gaughan, A.E.; Waylen, P.R. Spatial and temporal precipitation variability in the Okavango-Kwando-Zambezi catchment, Southern Africa. J. Arid Environ. 2012, 82, 19–30. [Google Scholar] [CrossRef]
- Matsuura, K.; Willmott, C. Terrestrial Air Temperature and Precipitation: 1900–2006 Gridded Monthly Time Series, Version 1.01; University of Delaware: Newark, DE, USA, 2007. [Google Scholar]
- Speer, J.H. Fundamentals of Tree-Ring Research; University of Arizona Press: Tucson, AZ, USA, 2010. [Google Scholar]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull 1983, 43, 69–78. [Google Scholar]
- Cook, E.R.; Holmes, R.L. Program ARSTAN Users Manual; Laboratory of Tree-Ring Research, University of Arizona: Tucson, AZ, USA, 1984. [Google Scholar]
- Potter, C.S.; Randerson, J.T.; Field, C.B.; Matson, P.A.; Vitousek, P.M.; Mooney, H.A.; Klooster, S.A. Terrestrial ecosystem production: a process model based on global satellite and surface data. Glob. Biogeochem. Cy. 1993, 7, 811–841. [Google Scholar] [CrossRef]
- Fensholt, R.; Rasmussen, K.; Kaspersen, P.; Huber, S.; Horion, S.; Swinnen, E. Assessing land degradation/recovery in the African Sahel from long-term earth observation based primary productivity and precipitation relationships. Remote Sens. 2013, 5, 664–686. [Google Scholar] [CrossRef]
- Tucker, C.; Pinzon, J.; Brown, M.; Slayback, D.; Pak, E.; Mahoney, R.; Vermote, E.; El Saleous, N. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int. J. Remote Sens. 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- Cook, B.; Pau, S. A global assessment of long-term greening and browning trends in pasture lands using the GIMMS LAI3g dataset. Remote Sens. 2013, 5, 2492–2512. [Google Scholar] [CrossRef]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations—The CRU TS3.10 dataset. Int. J. Climatol. 2013. [Google Scholar] [CrossRef]
- European Commission (JRC). Global Land Cover 2000 Database. Available online: http://bioval.jrc.ec.europa.eu/products/glc2000/glc2000.php (accessed on 18 March 2013).
- Field, C.B.; Randerson, C.J.T.; Malmström, M. Global net primary production: combining ecology and remote sensing. Remote Sens. Environ. 1995, 51, 74–97. [Google Scholar] [CrossRef]
- Gower, S.T.; Kirschbaum, A.A. BigFoot Field Data for North American Sites,1999–2003. Data Set; Oak Ridge National Laboratory Distributed Active Archive Center: Oak Ridge, TN, USA, 2008. [Google Scholar]
- Potter, C.; Klooster, S.; Genovese, V. Net primary production of terrestrial ecosystems from 2000 to 2009. Clim. Chang. 2012, 115, 365–378. [Google Scholar] [CrossRef]
- Stahle, D.G.H.; Klimowicz, J.; Muleya, S. Pterocarpus angolensis Chronology, Zimbabewe; International Tree-Ring Data Bank (ITRDB). Available online: http://www.ncdc.noaa.gov/paleo/treering.html (accessed on 18 March 2013).
- Webster, P.J. The role of hydrological processes in ocean-atmosphere interactions. Rev. Geophys. 1994, 32, 427–476. [Google Scholar] [CrossRef]
- Mushove, P.; Stahle, D.W.; Cleaveland, M.K. Pterocarpus angolensis Chronology, Zimbabewe; International Tree-Ring Data Bank (ITRDB). Available online: http://www.ncdc.noaa.gov/paleo/treering.html (accessed on 18 August 2013).
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Southworth, J.; Rigg, L.; Gibbes, C.; Waylen, P.; Zhu, L.; McCarragher, S.; Cassidy, L. Integrating Dendrochronology, Climate and Satellite Remote Sensing to Better Understand Savanna Landscape Dynamics in the Okavango Delta, Botswana. Land 2013, 2, 637-655. https://doi.org/10.3390/land2040637
Southworth J, Rigg L, Gibbes C, Waylen P, Zhu L, McCarragher S, Cassidy L. Integrating Dendrochronology, Climate and Satellite Remote Sensing to Better Understand Savanna Landscape Dynamics in the Okavango Delta, Botswana. Land. 2013; 2(4):637-655. https://doi.org/10.3390/land2040637
Chicago/Turabian StyleSouthworth, Jane, Lesley Rigg, Cerian Gibbes, Peter Waylen, Likai Zhu, Shannon McCarragher, and Lin Cassidy. 2013. "Integrating Dendrochronology, Climate and Satellite Remote Sensing to Better Understand Savanna Landscape Dynamics in the Okavango Delta, Botswana" Land 2, no. 4: 637-655. https://doi.org/10.3390/land2040637
APA StyleSouthworth, J., Rigg, L., Gibbes, C., Waylen, P., Zhu, L., McCarragher, S., & Cassidy, L. (2013). Integrating Dendrochronology, Climate and Satellite Remote Sensing to Better Understand Savanna Landscape Dynamics in the Okavango Delta, Botswana. Land, 2(4), 637-655. https://doi.org/10.3390/land2040637