
Decline of the Maurepas Swamp, Pontchartrain Basin, Louisiana, and Approaches to Restoration


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
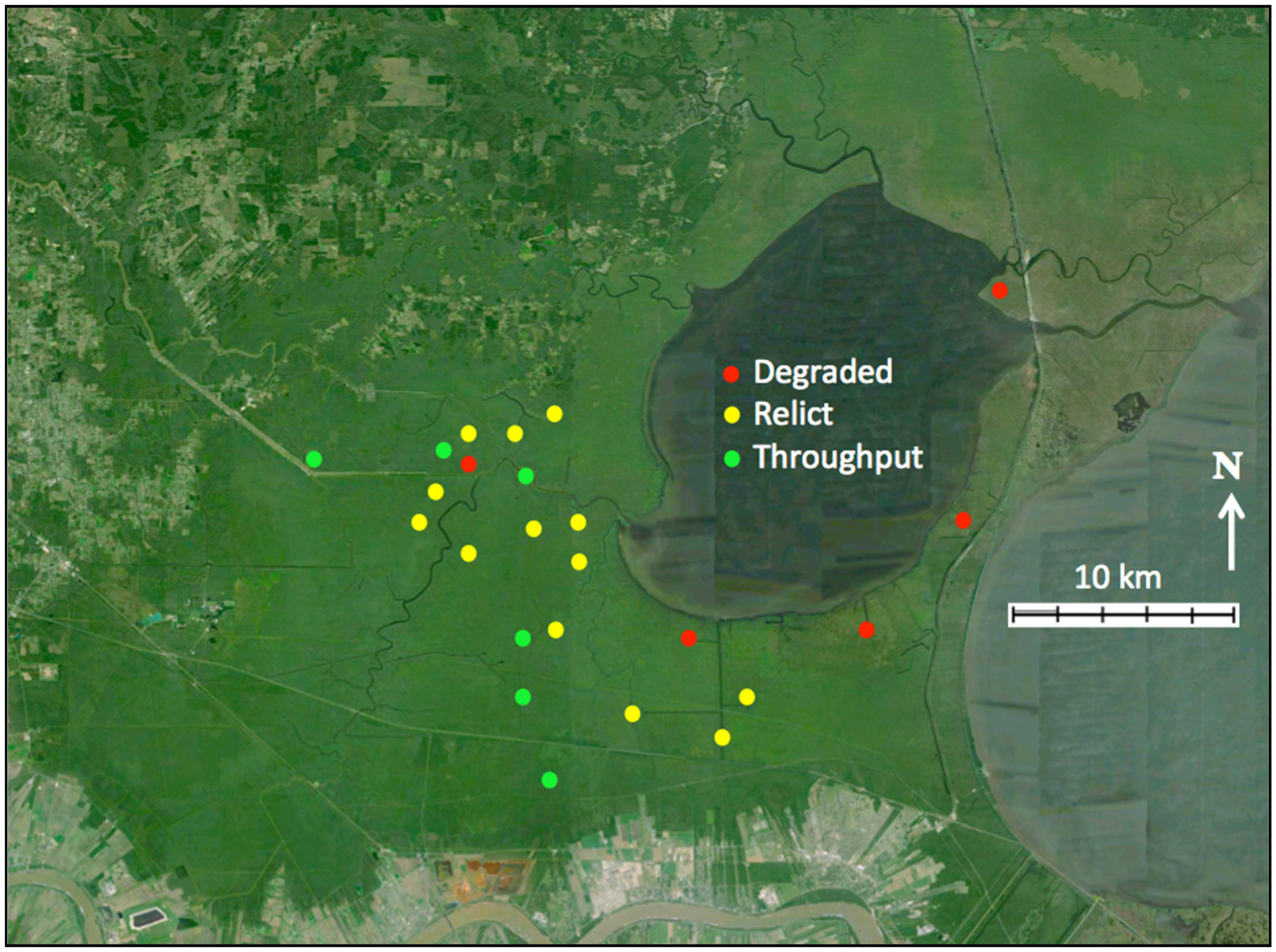
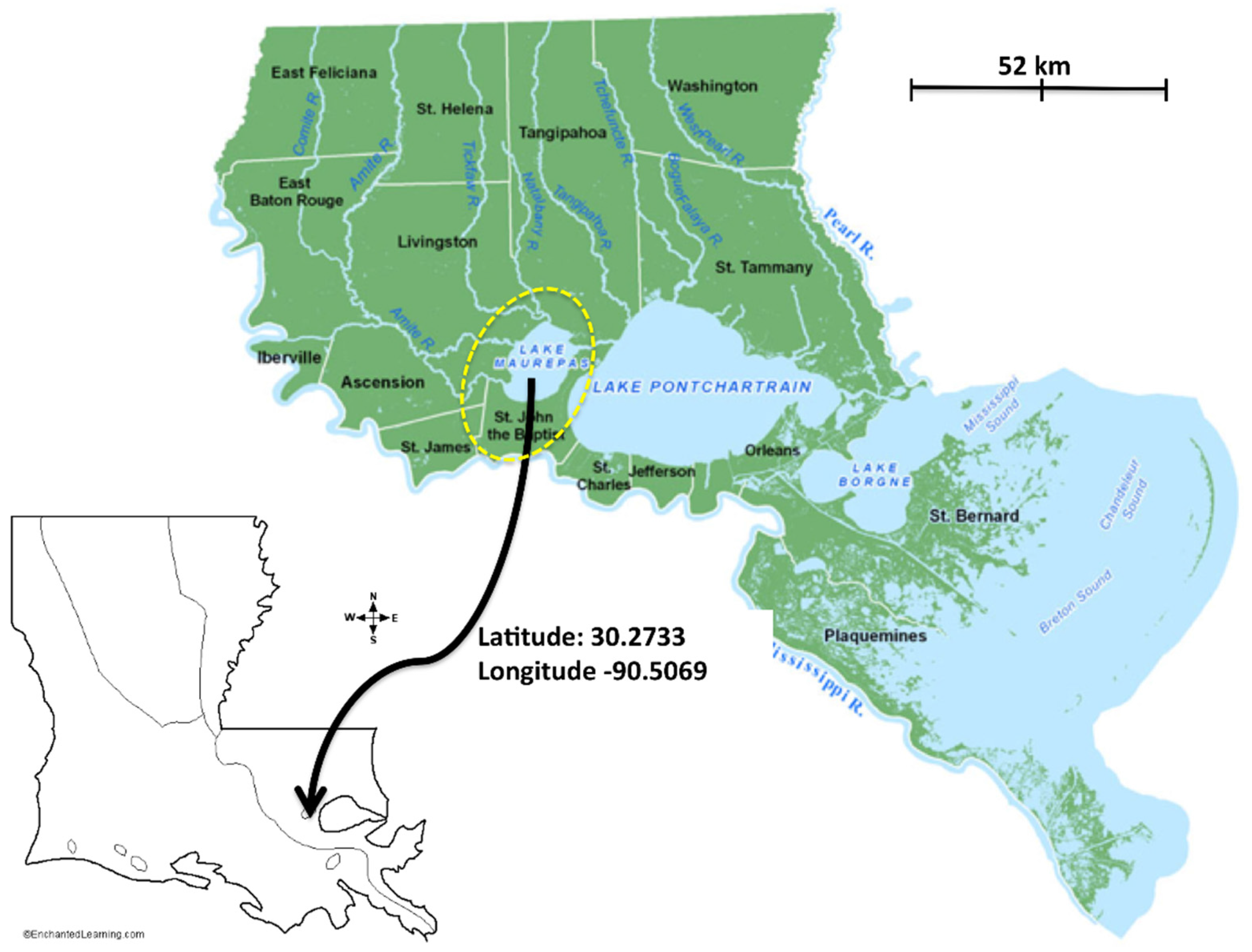
2.1. Study Sites
2.2. Environmental Variables
2.3. Herbaceous Vegetation
2.4. Forest Vegetation
2.5. Sediment Elevation Change
2.6. Mapping Habitat Types
2.7. Statistical Analysis
3. Results
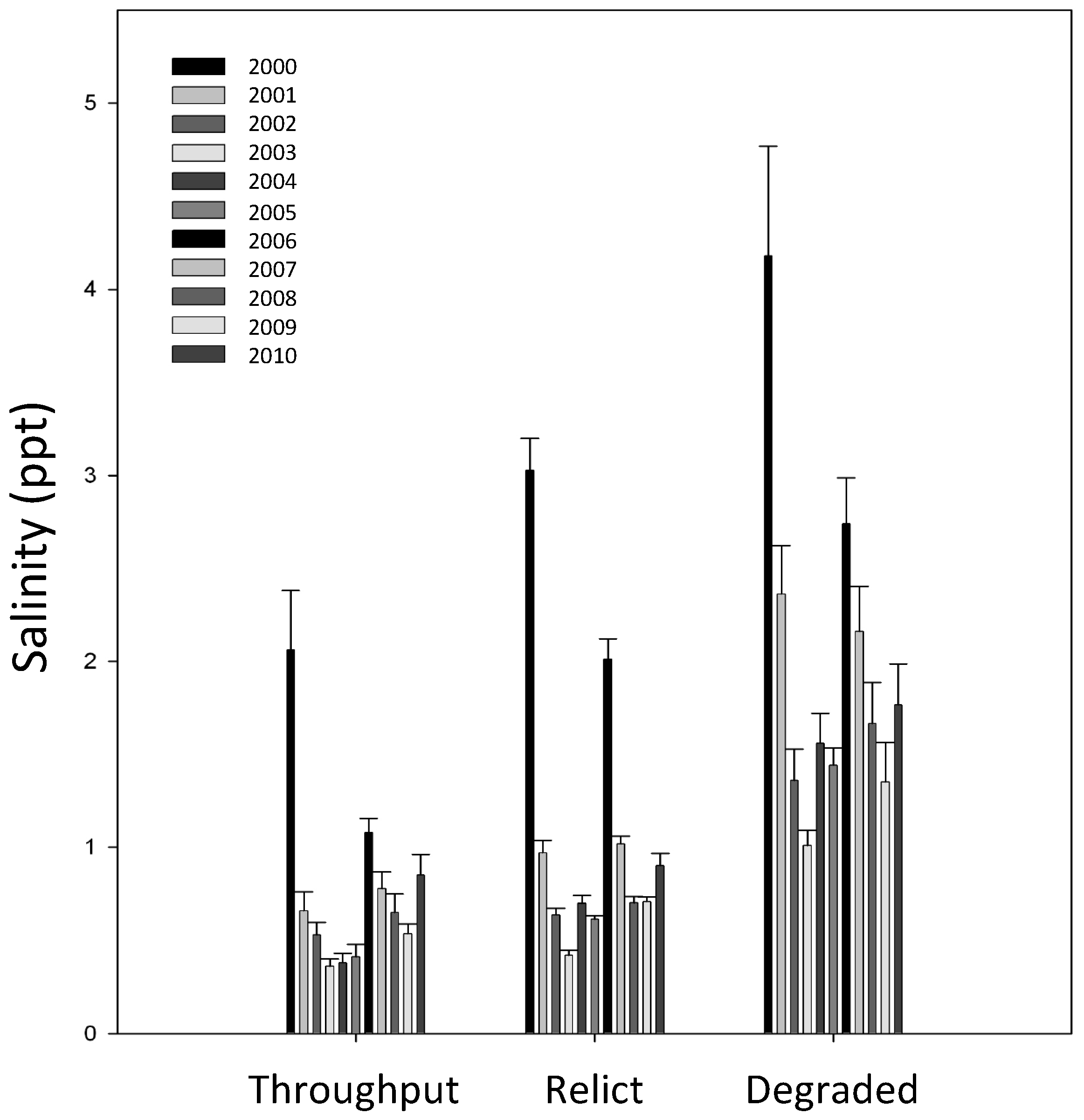
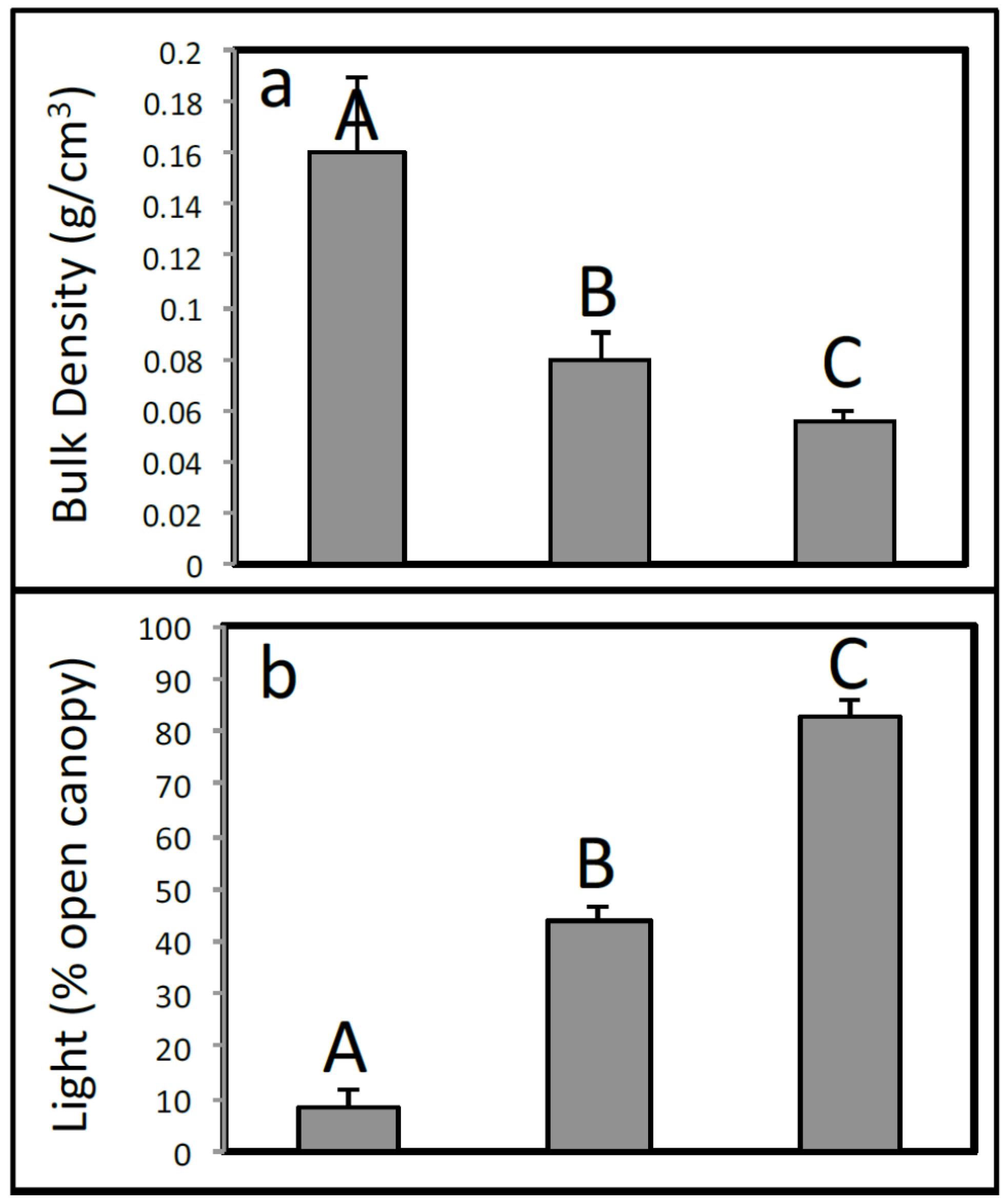
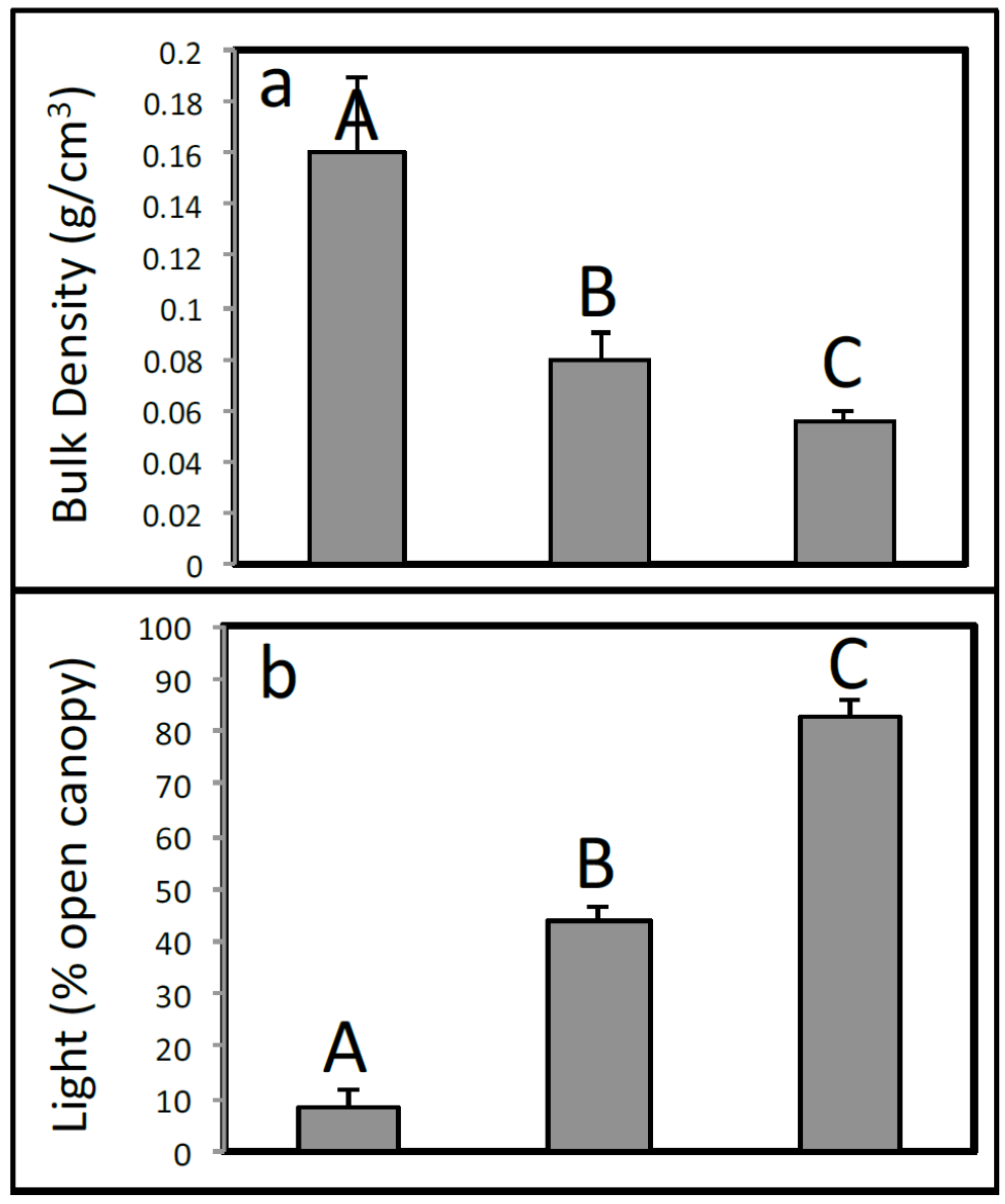
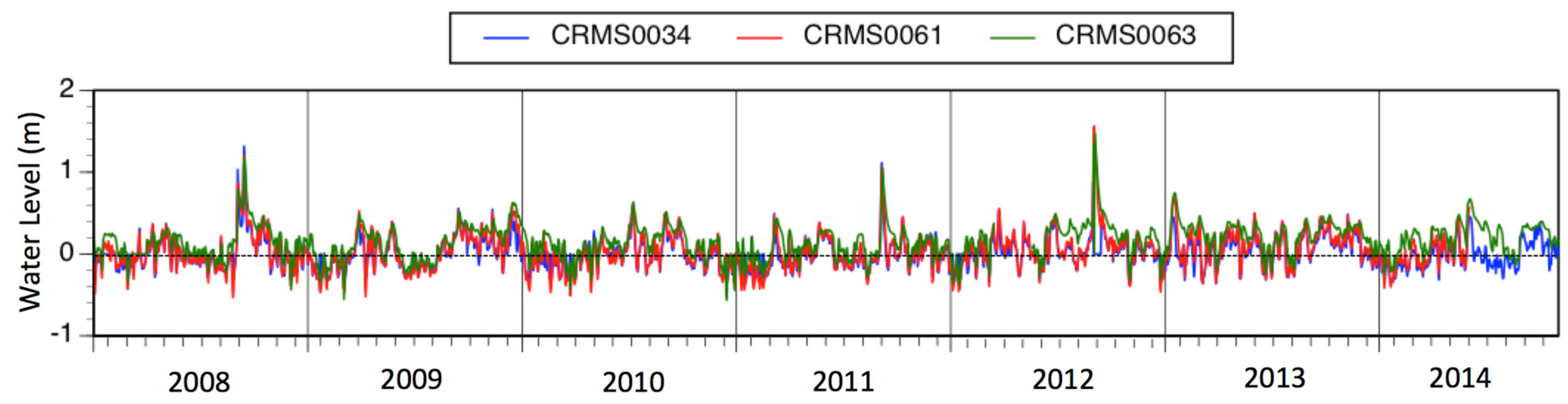
3.1. Environmental Variables
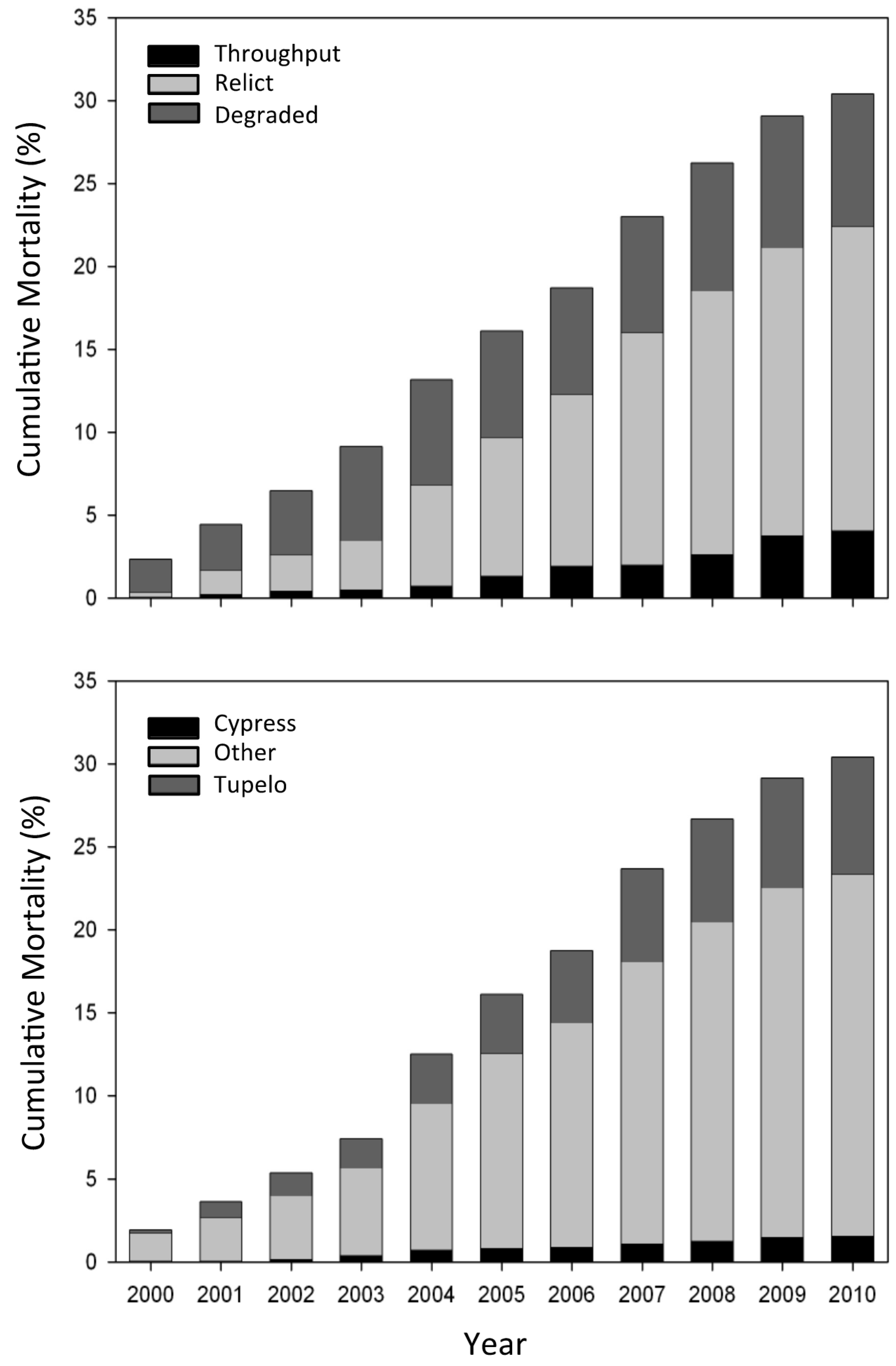
3.2. Tree Mortality
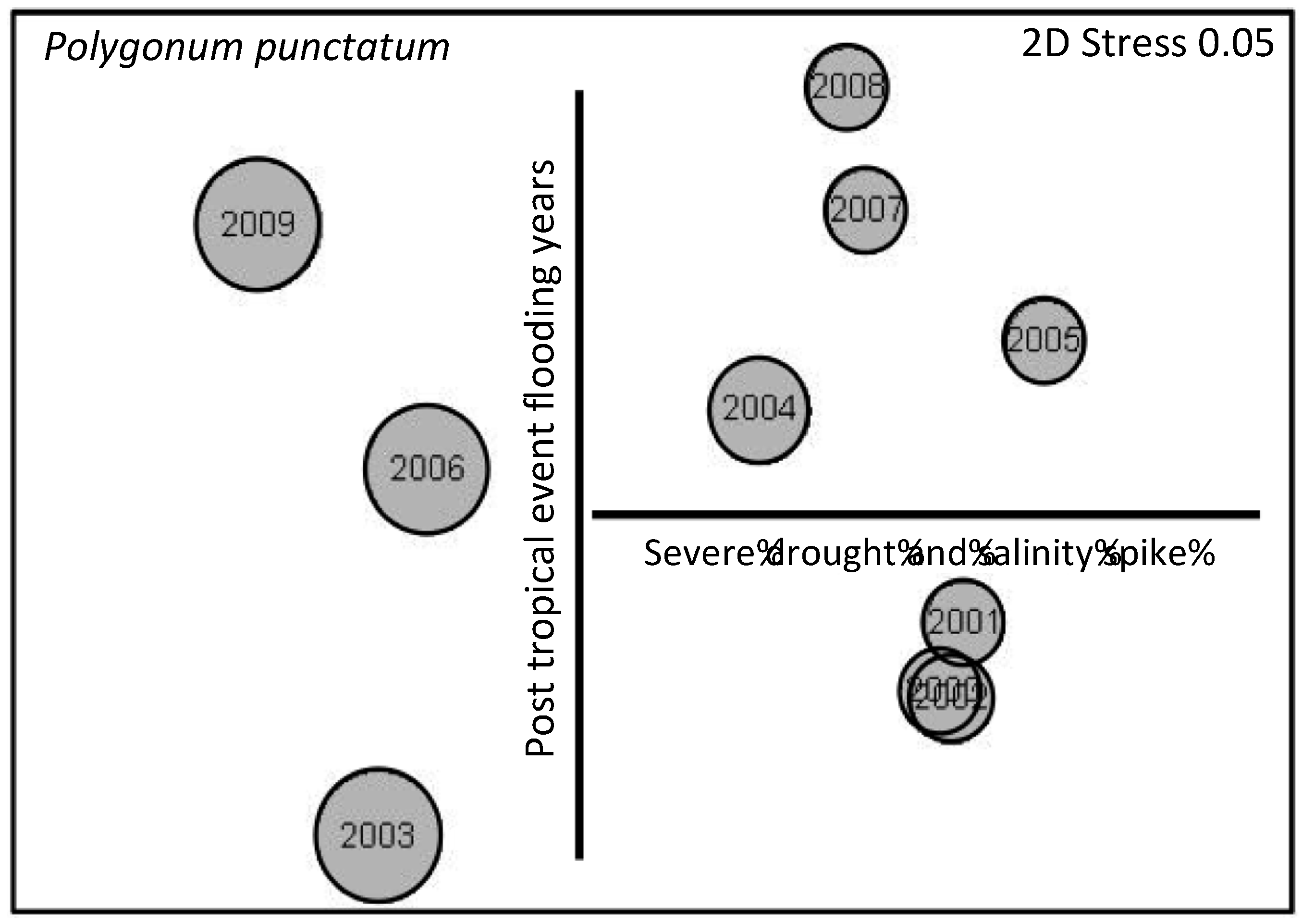
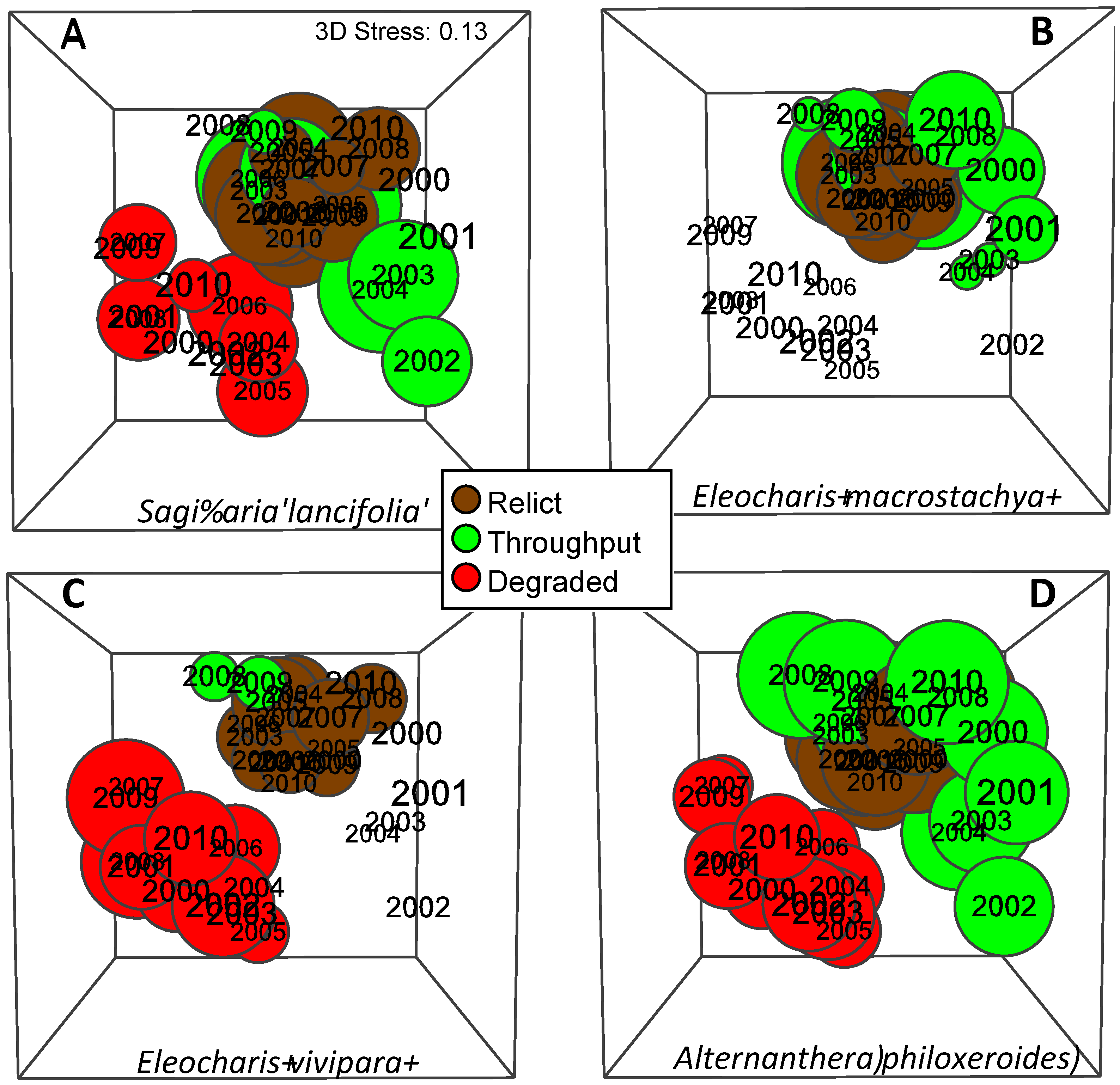
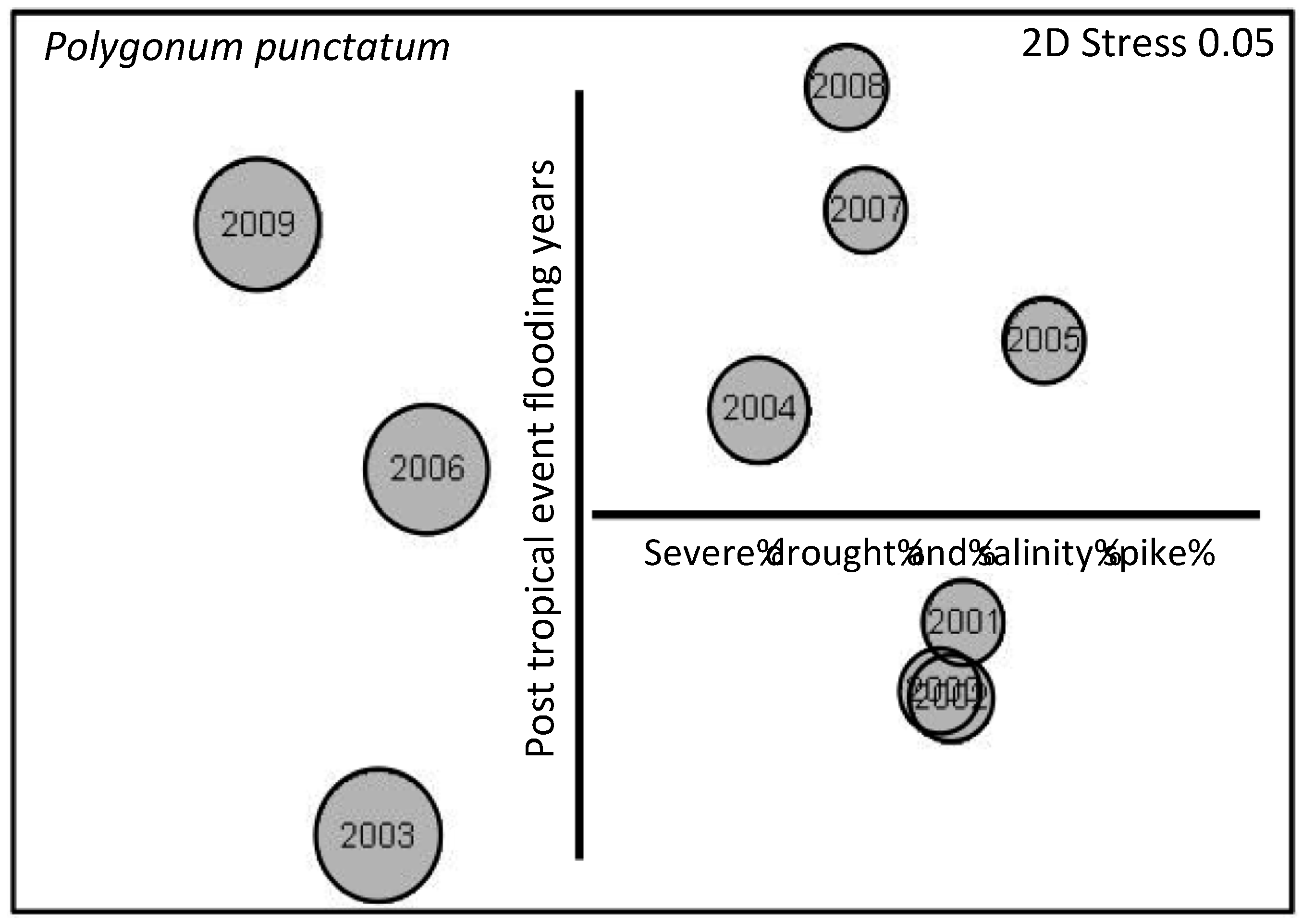
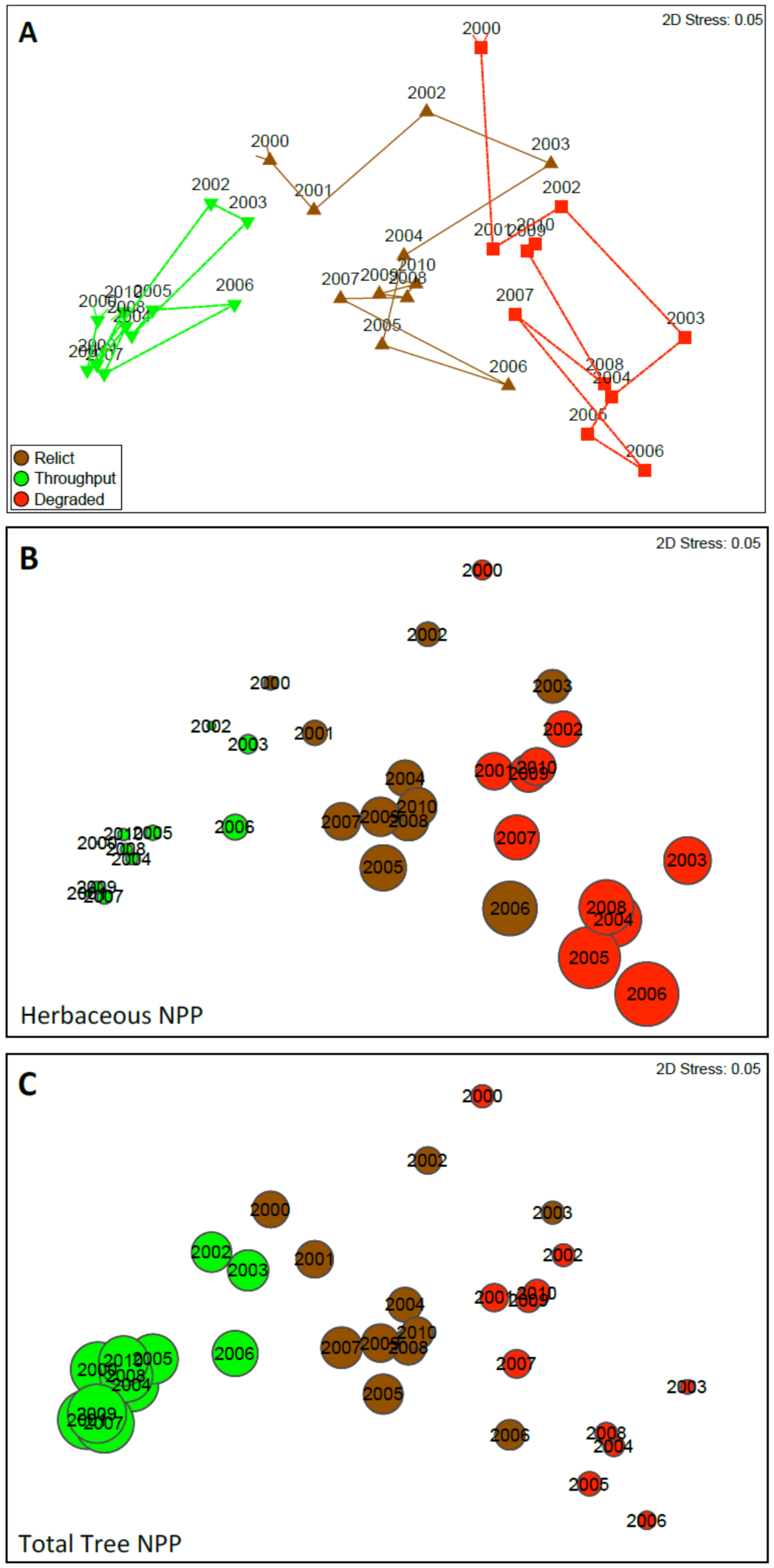
3.3. Herbaceous Vegetation
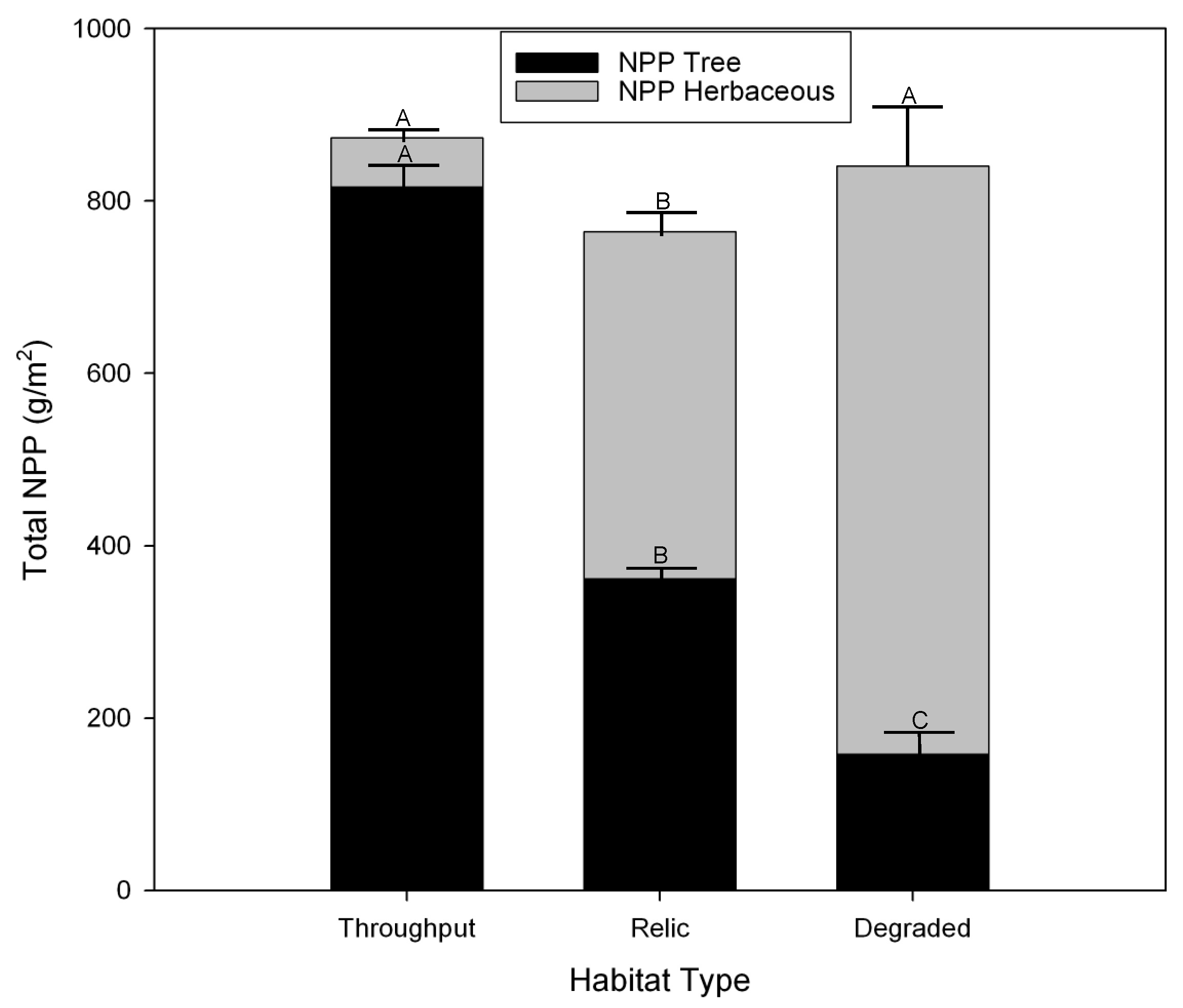
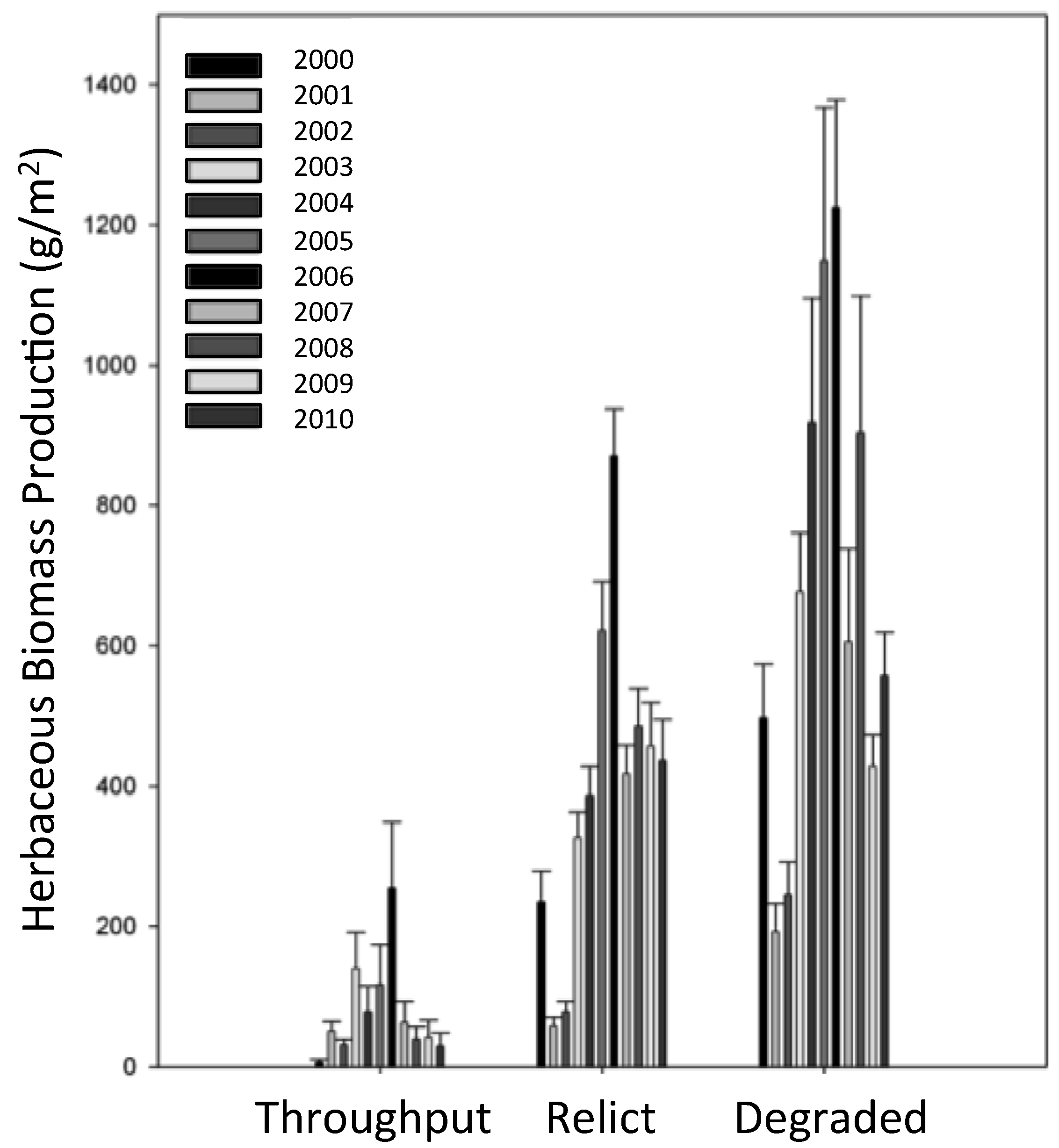
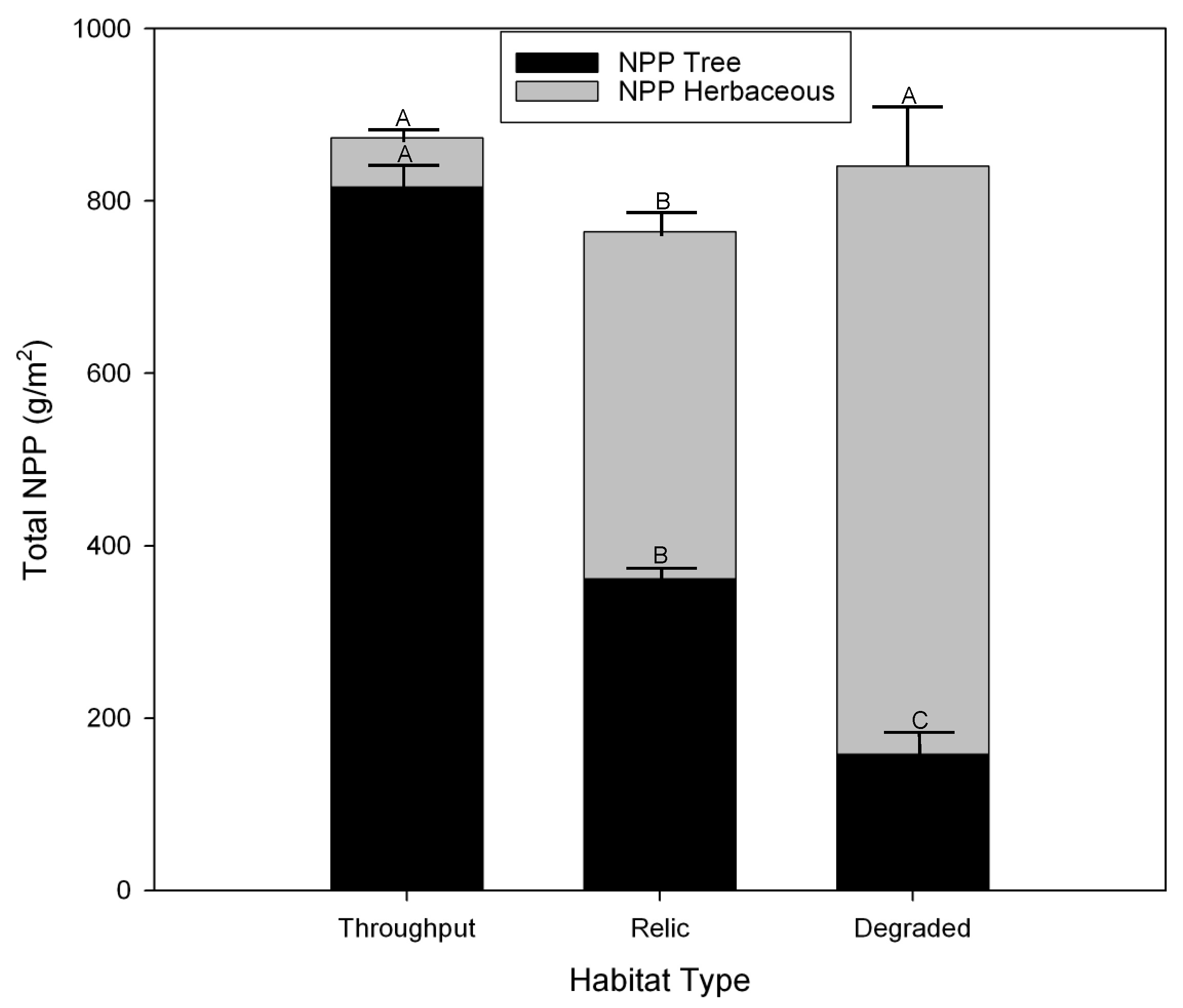
3.4. Herbaceous Vegetation Production
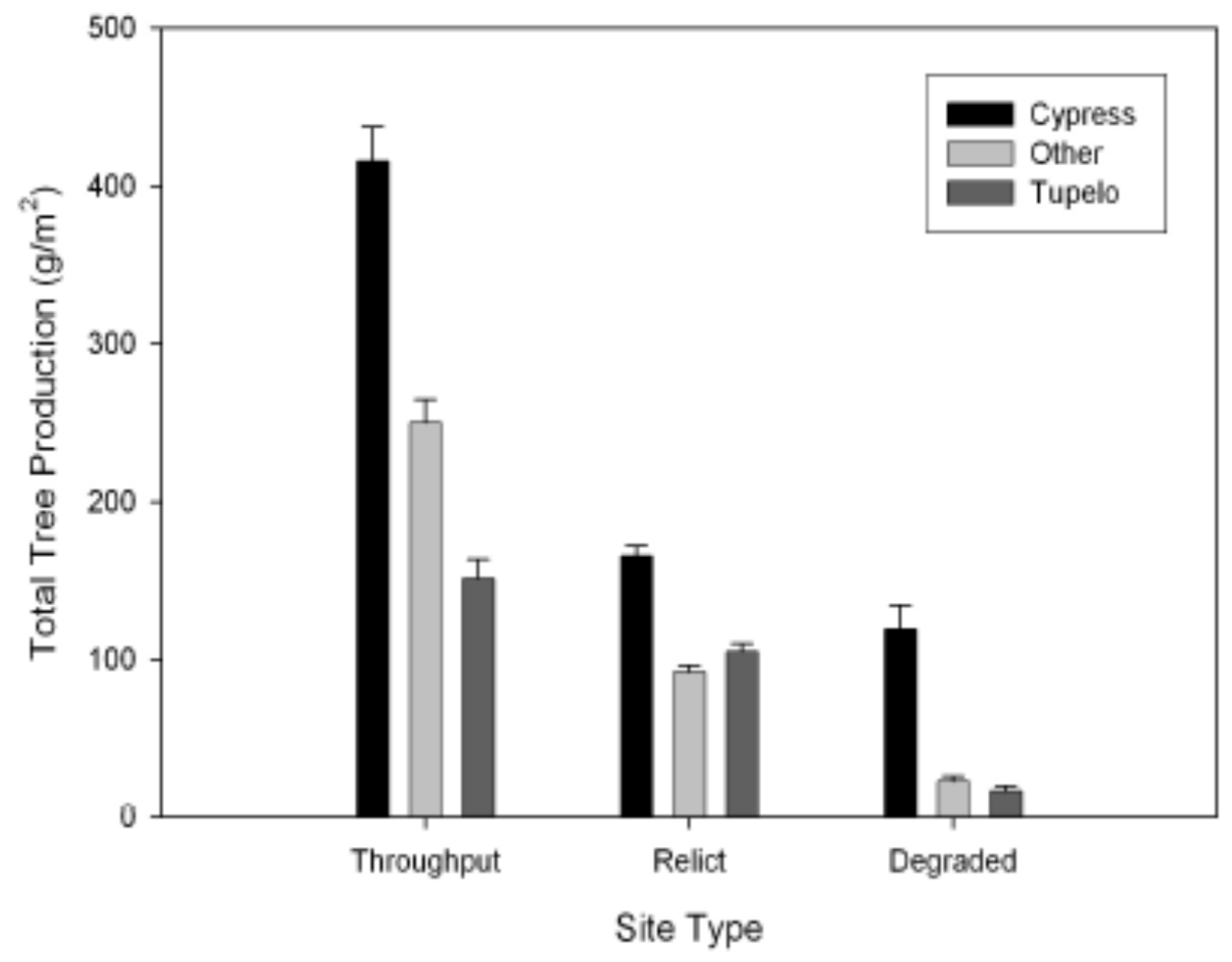
3.5. Tree Production
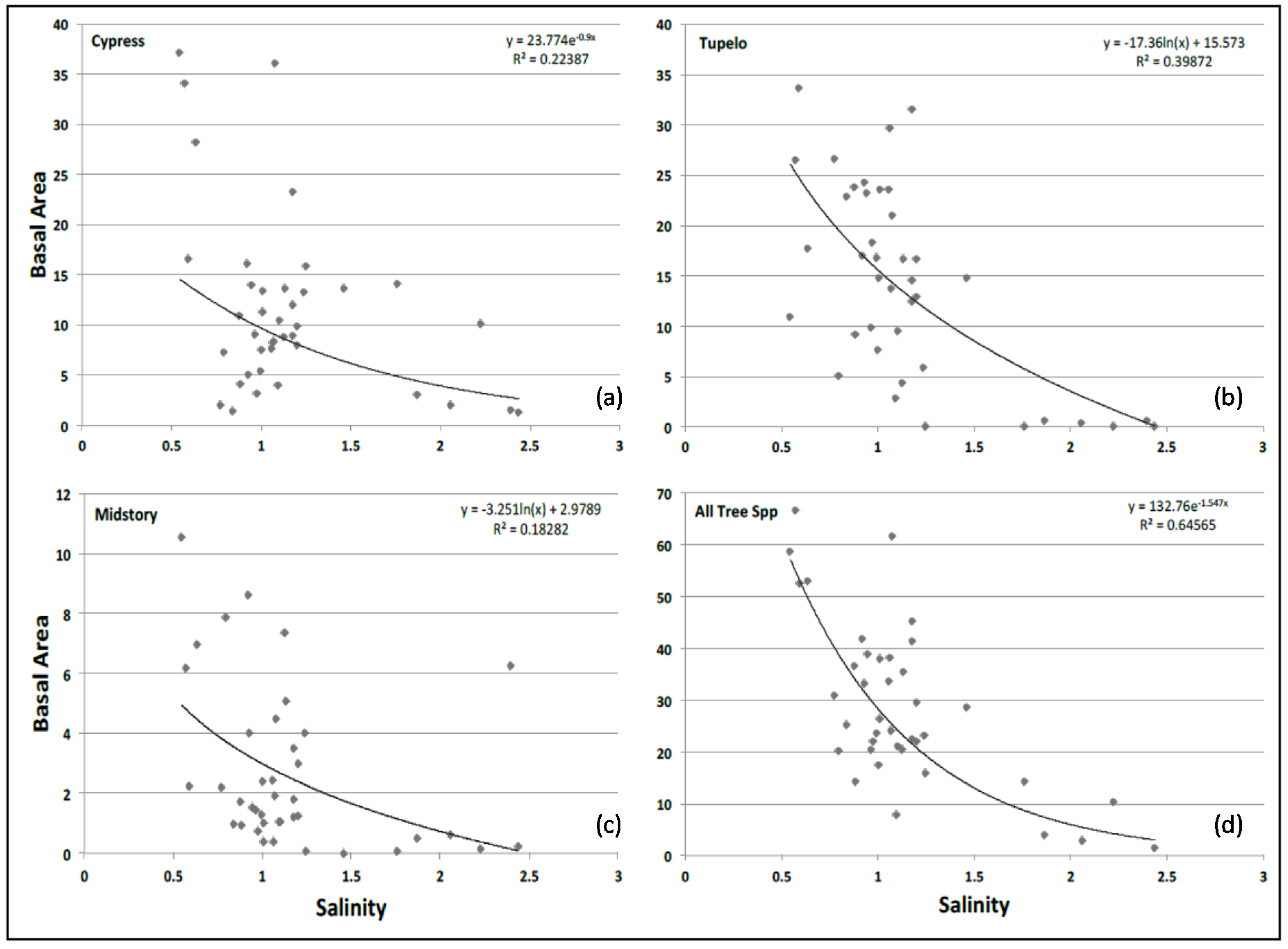
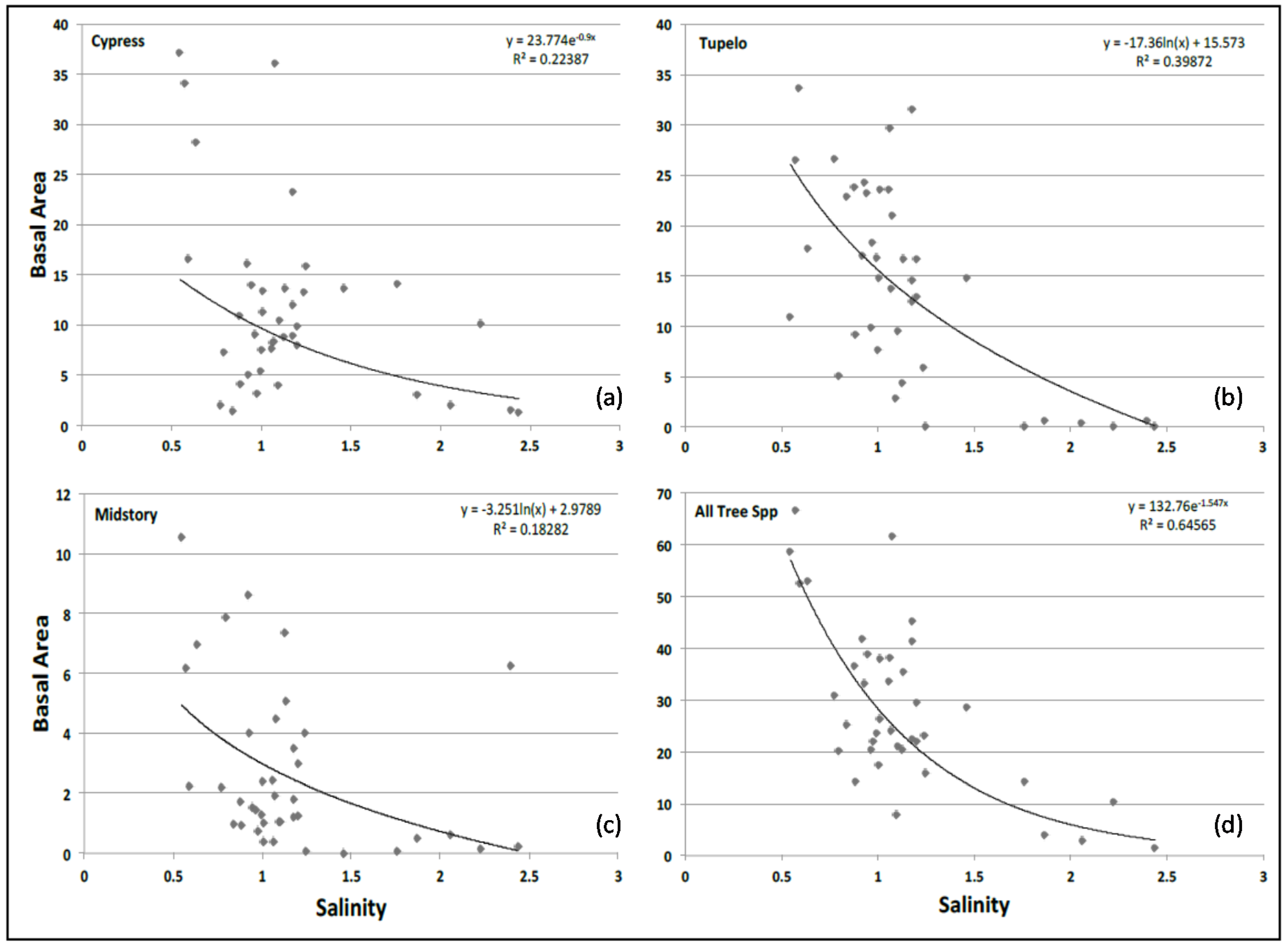
3.6. Basal Area and Salinity
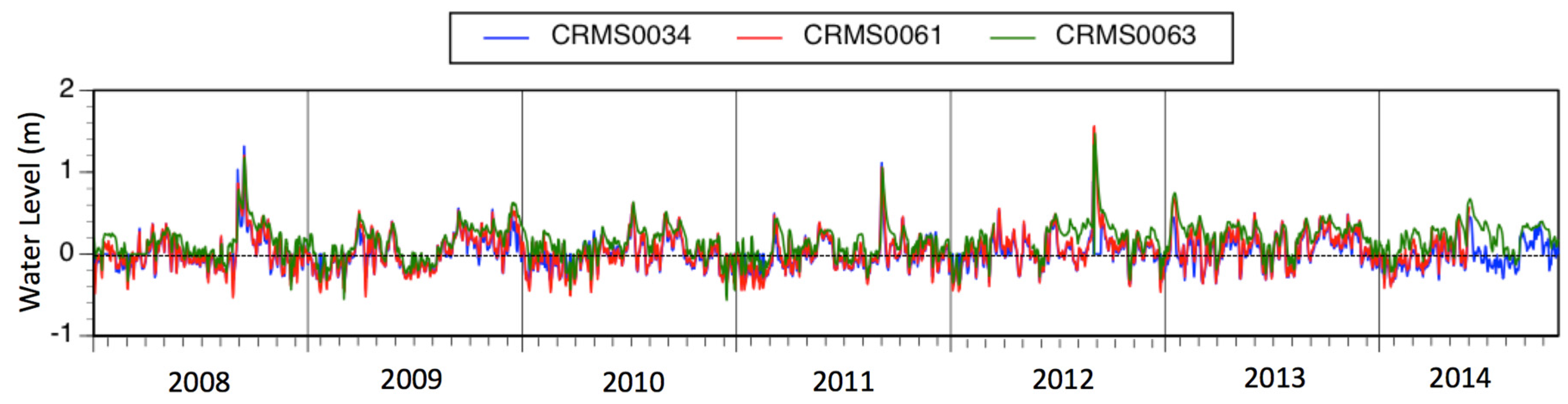
3.7. Surface Elevation Change
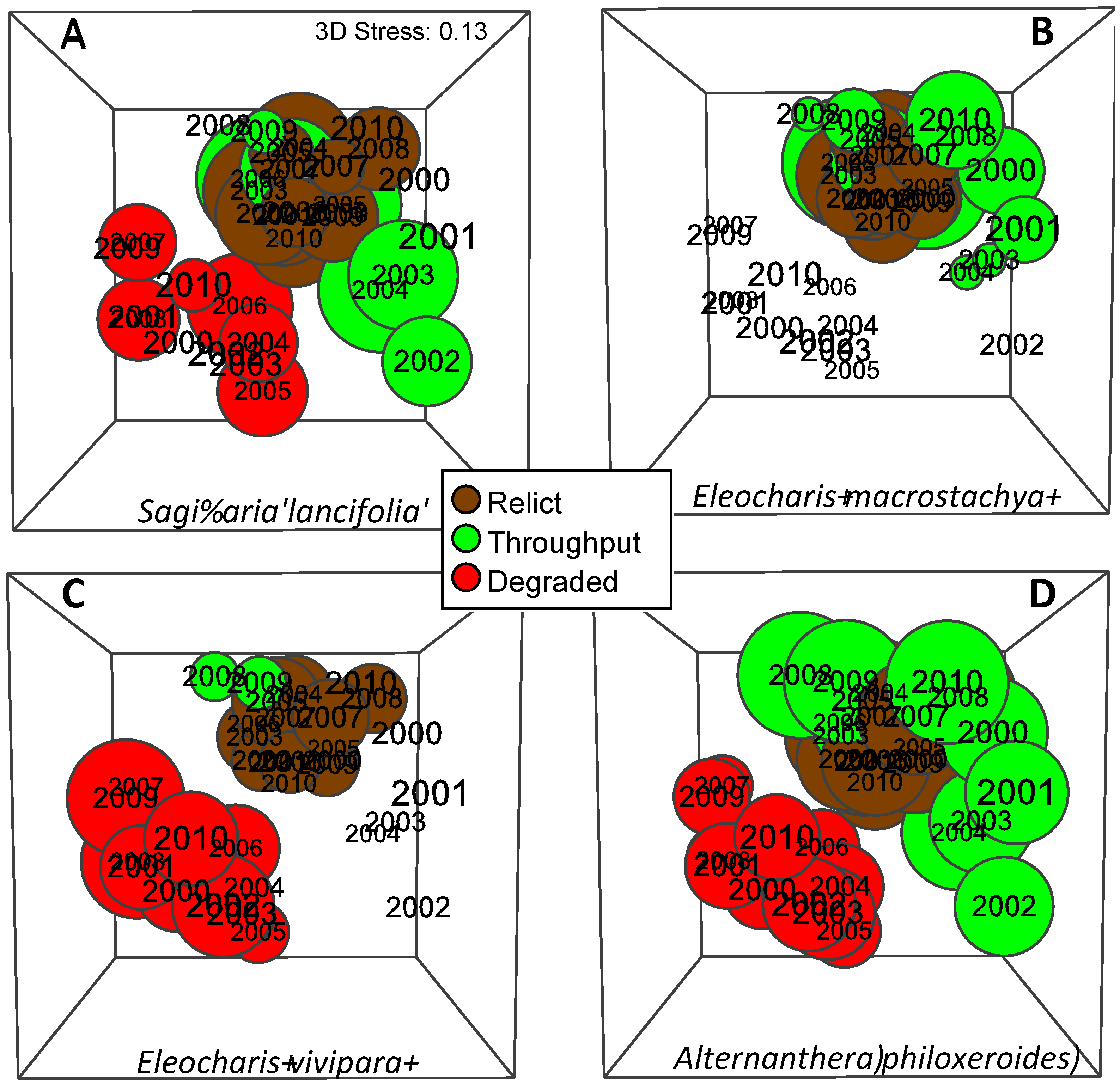
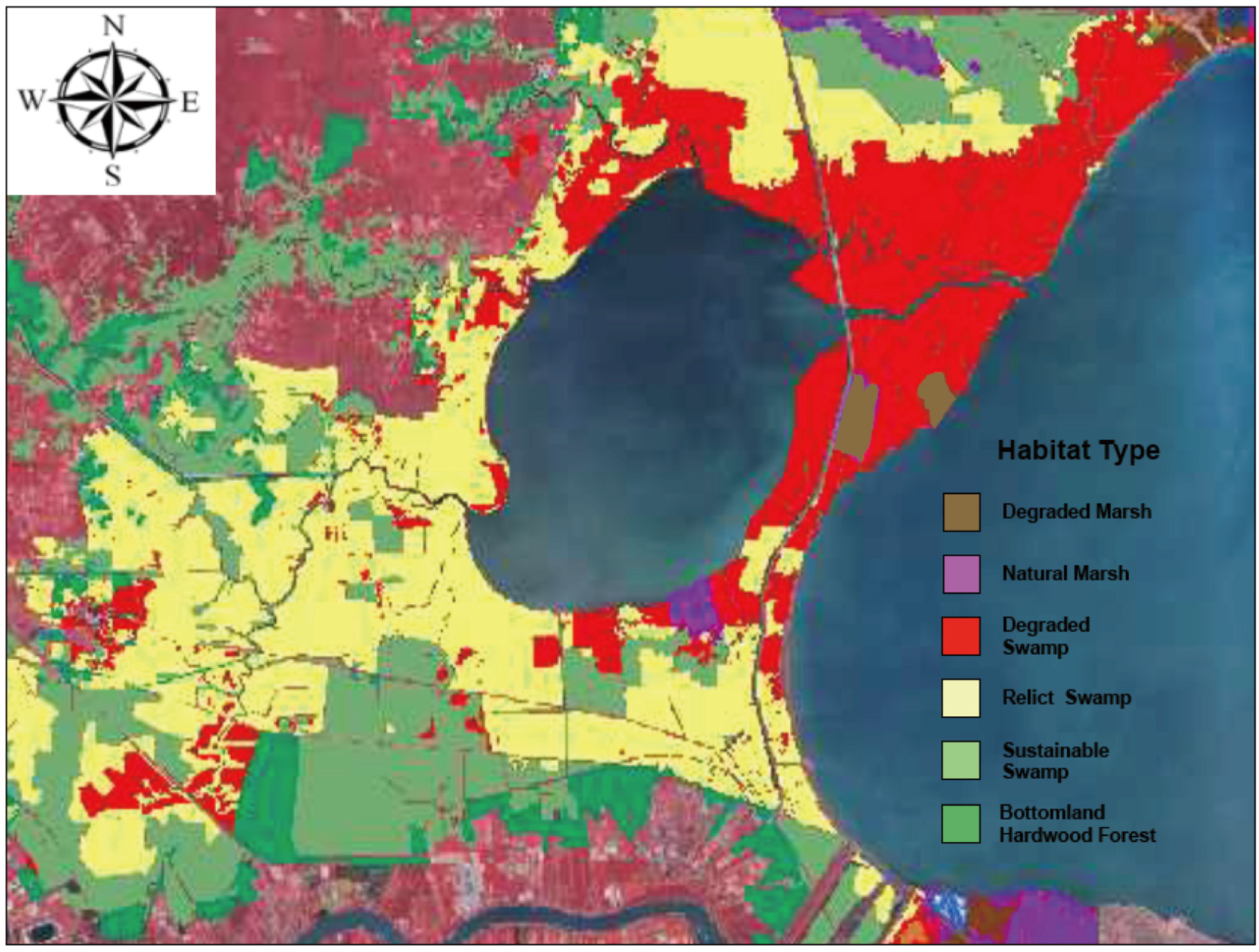
3.8. Habitat Classification
4. Discussion
5. Restoration of the Maurepas Swamp
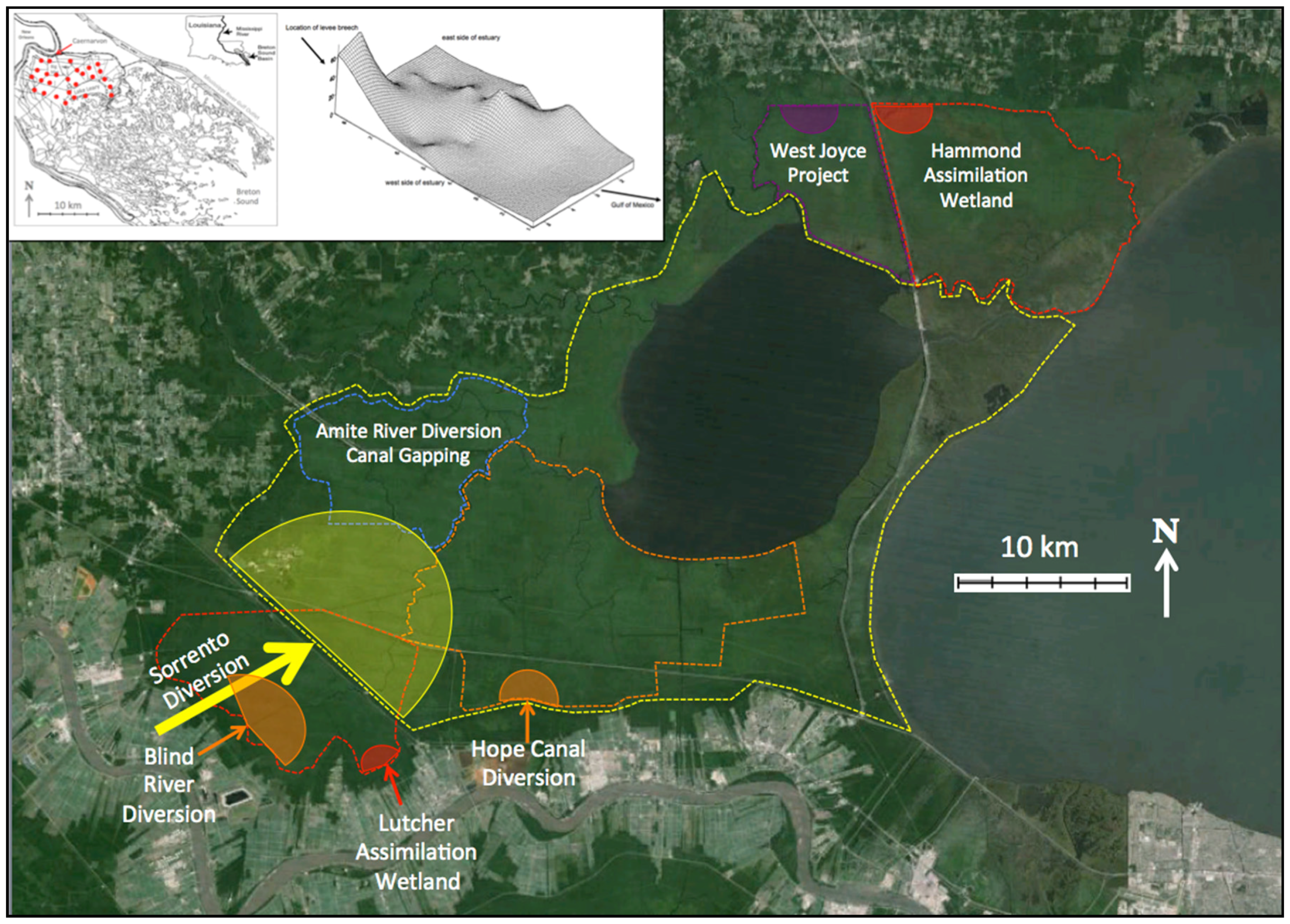
5.1. Approaches to Restoration
5.2. Impediments to Restoration
5.2.1. Saltwater Intrusion and Sea Level Rise.
5.2.2. Hydrologic Isolation
5.2.3. Nutrient Limitation
5.2.4. Herbivory
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Day, J.W.; Kemp, G.P.; Freemen, A.M.; Muth, D.P. Perspectives on the Restoration of the Mississippi Delta; Springer: Dordrecht, The Netherlands, 2014; p. 194. [Google Scholar]
- Barras, J.A.; Bourgeois, P.E.; Handley, L.R. Land Loss in Coastal Louisiana, 1956–1990; National Wetlands Research Center Open File Report 94-01; National Biological Survey: Lafayette, LA, USA, 1994. [Google Scholar]
- Barras, J.A.; Bernier, J.C.; Morton, R.A. Land Area Change in Coastal Louisiana—A Multidecadal Perspective (from 1956 to 2006); U.S. Geological Survey Scientific Investigations Map 3019, scale 1:250,000; US Department of the Interior, US Geological Survey: Lafayette, LA, USA, 2008; p. 14.
- Britsch, L.D.; Dunbar, J.B. Land loss rates: Louisiana Coastal Plain. J. Coast. Res. 1993, 9, 324–338. [Google Scholar]
- Couvillion, B.R.; Barras, J.A.; Steyer, G.D.; Sleavin, W.; Fischer, M.; Beck, H.; Trahan, N.; Griffin, B.; Heckman, D. Land Area Change in Coastal Louisiana from 1932 to 2010; US Geological Survey Scientific Investigation Map 3164, scale 1:265,000; US Department of the Interior, US Geological Survey: Reston, VA, USA, 2011; p. 12.
- Day, J.W.; Shaffer, G.P., Jr.; Britsch, L.D.; Hawes, S.R.; Reed, D.J.; Cahoon, D. Pattern and process of land loss in the Mississippi delta: A spatial and temporal analysis of wetland habitat change. Estuaries 2000, 23, 425–438. [Google Scholar] [CrossRef]
- Day, J.W., Jr.; Boesch, D.F.; Clairain, E.F.; Kemp, G.P.; Laska, S.B.; Mitsch, W.J.; Orth, K.; Mashriqui, H.; Reed, D.R.; Shabman, L.; et al. Restoration of the Mississippi Delta: Lessons from Hurricanes Katrina and Rita. Science 2007, 315, 1679–1684. [Google Scholar] [CrossRef] [PubMed]
- CPRA. Louisiana’s Comprehensive Master Plan for a Sustainable Coast; Coastal Protection and Restoration Authority: Baton Rouge, LA, USA, 2012. [Google Scholar]
- Chambers, J.L.; Conner, W.H.; Day, J.W., Jr.; Faulkner, S.P.; Gardiner, E.S.; Hughes, M.S.; Keim, R.F.; King, S.L.; Miller, C.A.; Nyman, J.A.; et al. Conservation, Protection, and Utilization of Louisiana’s Coastal Wetland Forests; Final report to the Governor of Louisiana from the Coast Wetland Forest Conservation and Use Science Working Group: Baton Rouge, LA, USA, 2005; p. 121. [Google Scholar]
- Shaffer, G.P.; Wood, W.B.; Hoeppner, S.S.; Perkins, T.E.; Zoller, J.A.; Kandalepas, D. Degradation of baldcypress-water tupelo swamp to marsh and open water in southeastern Louisiana, USA: An irreversible trajectory? J. Coast. Res. 2009, 54, 152–165. [Google Scholar] [CrossRef]
- Shaffer, G.P.; Day, J.W.; Mack, S.; Kemp, G.P.; van Heerden, I.; Poirrier, M.A.; Westpahl, K.A.; FitzGerald, D.; Milanes, A.; Morris, C.; et al. The MRGO navigation project: A massive human-induced environmental, economic, and storm disaster. J. Coast. Res. 2009, 54, 206–224. [Google Scholar] [CrossRef]
- Day, J.W.; Hunter, R.; Keim, R.; DeLaune, R.; Shaffer, G.; Evers, E.; Reed, D.; Brantley, C.; Kemp, P.; Day, J.; et al. Ecological response of forested wetlands with and without large-scale Mississippi River input: Implications for management. Ecol. Eng. 2012, 46, 57–67. [Google Scholar] [CrossRef]
- Hunter, R.G.; Day, J.W., Jr.; Lane, R.R.; Lindsey, J.; Day, J.N.; Hunter, M.G. Impacts of secondarily treated municipal effluent on a freshwater forested wetland after 60 years of discharge. Wetlands 2009, 29, 363–371. [Google Scholar] [CrossRef]
- Batker, D.; de la Torre, I.; Costanza, R.; Swedeen, P.; Day, J.; Boumans, R.; Bagstad, K. Gaining Ground: Wetlands, Hurricanes and the Economy: The Value of Restoring the Mississippi River Delta; Earth Economics, Inc.: Tacoma, WA, USA, 2010; p. 98. [Google Scholar]
- Costanza, R.; d’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- LDNR. Coast 2050: Toward a Sustainable Coastal Louisiana; Louisiana Coastal Wetlands Conservation and Restoration Task Force and the Wetlands Conservation and Restoration Authority. Louisiana Department of Natural Resources: Baton Rouge, LA, USA, 1998; p. 161. [Google Scholar]
- Lopez, J.A. The Environmental History of human-induced impacts to the Lake Pontchartrain Basin in Southeastern Louisiana since European settlement-1718 to 2002. J. Coast. Res. 2009, 54, 1–11. [Google Scholar] [CrossRef]
- Saucier, R.T. Recent Geomorphic History of the Ponchartrain Basin; Louisiana State University, Coastal Studies Series 9: Baton Rouge, LA, USA, 1963; p. 114. [Google Scholar]
- Conner, W.H.; Day, J.W. Productivity and composition of a baldcypress-water tupelo site and a bottomland hardwood site in a Louisiana swamp. Am. J. Bot. 1976, 63, 1354–1364. [Google Scholar] [CrossRef]
- Conner, W.H.; Day, J.W. Diameter growth of Taxodium distichum (L.) Rich. and Nyssa aquatica L. from 1979–1985 in four Louisiana swamp stands. Am. Mid. Nat. 1992, 127, 209–299. [Google Scholar] [CrossRef]
- Hoeppner, S.S. Feasibility and Project Benefits of a Diversion into the Degraded Cypress–Tupelo Swamp in the Southern Lake Maurepas Wetlands, Lake Pontchartrain Basin, Louisiana. Master’s Thesis, Southeastern Louisiana University, Hammond, LA, USA, 2002. [Google Scholar]
- Hoeppner, S.S.; Shaffer, G.P.; Perkins, T.E. Through droughts and hurricanes: Tree mortality, forest structure, and biomass production in a coastal swamp targeted for restoration in the Mississippi River Deltaic. For. Ecol. Manag. 2008, 256, 937–948. [Google Scholar] [CrossRef]
- Pezeshki, S.R.; DeLaune, R.D.; Patrick, W.H.J. Flooding and saltwater intrusion: Potential effects on survival and productivity of wetland forests along the U.S. Gulf Coast. For. Ecol. Manag. 1990, 33/34, 287–301. [Google Scholar] [CrossRef]
- Shaffer, G.P.; Perkins, T.E.; Hoeppner, S.S.; Howell, S.; Benard, T.H.; Parsons, A.C. Ecosystem Health of the Maurepas Swamp: Feasibility and Projected Benefits of a Freshwater Diversion; Final Report; Environmental Protection Agency, Region 6: Dallas, TX, USA, 2003; p. 95. [Google Scholar]
- Morton, R.; Barras, J. Hurricane impacts on coastal wetlands: A half-century record of storm-generated features from southern Louisiana. J. Coast. Res. 2011, 27, 27–43. [Google Scholar] [CrossRef]
- Howes, N.C.; FitzGerald, D.M.; Hughes, Z.J.; Georgiou, I.Y.; Kulp, M.A.; Miner, M.D.; Smith, J.M.; Barras, J.A. Hurricane-induced failure of low salinity wetlands. Proc. Natl. Acad. Sci. USA 2010, 107, 14014–14019. [Google Scholar] [CrossRef] [PubMed]
- Trettin, C.C.; Jorgensen, M.F. Carbon Cycling in Wetland Forest Soils. In The Potential of U.S. Forest Soils to Sequester Carbon and Mitigate the Greenhouse Effect; Kimble, J.M., Heath, L., Birdsey, R.A., Lai, R., Eds.; CRC Press: Boca Raton, FL, USA, 2003; pp. 311–331. [Google Scholar]
- Lopez, J.A. The multiple lines of defense strategy to sustain coastal Louisiana. J. Coast. Res. 2009, 54, 186–197. [Google Scholar] [CrossRef]
- Clark, A.; Phillips, D.R.; Frederick, D.J. Weight, Volume, and Physical Properties of Major Hardwood Species in the Gulf and Atlantic Coastal Plains; USDA Forest Service Research Paper SE-250; Southeastern Forest Experimental Station: Asheville, NC, USA, 1985. [Google Scholar]
- Muzika, R.M.; Gladden, J.B.; Haddock, J.D. Structural and Functional Aspects of Succession in South-eastern Floodplain Forests Following a Major Disturbance. Am. Midl. Nat. 1987, 117, 1–9. [Google Scholar] [CrossRef]
- Scott, M.L.; Sharitz, R.R.; Lee, L.C. Disturbance in a Cypress-Tupelo Wetland: An interaction between thermal loading and hydrology. Wetlands 1985, 5, 53–68. [Google Scholar] [CrossRef]
- Cahoon, D.R.; Marin, P.E.; Black, B.K.; Lynch, J.C. A method for measuring vertical accretion, elevation, and compaction of soft, shallow-water sediments. J. Sed. Res. 2000, 70, 1250–1253. [Google Scholar] [CrossRef]
- Lane, R.R.; Day, J.W.; Kemp, G.P.; Mashriqui, H.S.; Day, J.N.; Hamilton, A. Potential Nitrate Removal from a Mississippi River Diversion into the Maurepas Swamps. Ecol. Eng. 2003, 20, 237–249. [Google Scholar] [CrossRef]
- NOAA. Tides & Currents, Mean Sea Level Trend, Grand Isle, Louisiana: National Oceanic and Atmospheric Administration, On-line Material. Available online: http://tidesandcurrents.noaa.gov/sltrends/sltrends_station.shtml?id=8761724 (accessed on 23 March 2015).
- Wilkinson, L. SYSTAT 10.2 for Windows; SYSTAT Software, Inc.: Chicago, IL, USA, 2001. [Google Scholar]
- SAS Institute Inc. SAS 9.1.3 Help and Documentation; SAS Institute Inc.: Cary, NC, USA, 2000–2004. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, Fourth Edition; Prentice Hall: Upper Saddle River, NJ, USA, 1996; p. 662. [Google Scholar]
- Kruskal, J.B. Multidimensional scaling by optimizing goodness of fit to a nonmetric hypothesis. Psychometrika 1964, 29, 1–27. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Primer 6; PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Thomson, D.A.; Shaffer, G.P.; McCorquodale, J.A. A potential interaction between sea-level rise and global warming: Implications for coastal stability in the Mississippi River deltaic plain. Glob. Plan. Chang. 2002, 32, 49–59. [Google Scholar] [CrossRef]
- Cormier, N.; Krauss, K.W.; Conner, W.H. Periodicity in stem growth and litterfall in tidal freshwater forested wetlands: Influence of salinity and drought on nitrogen recycling. East Coasts 2013, 36, 533–546. [Google Scholar] [CrossRef]
- Hackney, C.T.; Avery, G.B.; Leonard, L.A.; Posey, M.; Alpin, T. Biological, Chemical, and Physical Characteristics of Tidal Freshwater Swamp Forests of the Lower Cape Fear River/Estuary, North Carolina. In Ecology of Tidal Freshwater Forested Wetlands of the Southeastern United States; Conner, W.H., Doyle, T.W., Krauss, K.W., Eds.; Springer: New York, NY, USA, 2007; pp. 183–221. [Google Scholar]
- Krauss, K.W.; Duberstein, J.A.; Doyle, T.W.; Conner, W.H.; Day, R.H.; Inabinette, L.W. Site condition, stand structure, and growth of baldcypress along tidal/non-tidal salinity gradients. Wetlands 2009, 29, 505–519. [Google Scholar] [CrossRef]
- Chabreck, R.H. Vegetation, Water and Soil Characteristics of the Louisiana Coastal Region; Bulletin 664; Louisiana State University Agricultural Experimental Station: Baton Rouge, LA, USA, 1972; p. 72. [Google Scholar]
- Penfound, W.T.; Hathaway, E.S. Plant communities in the marshlands of Southeastern Louisiana. Ecol. Mono. 1938, 8, 1–56. [Google Scholar] [CrossRef]
- Conner, W.H.; McLeod, K.W.; McCarron, J.K. Flooding and salinity effects on growth and survival of four common forested wetland species. Wetl. Ecol. Manag. 1997, 5, 99–109. [Google Scholar] [CrossRef]
- Pezeshki, S.R.; DeLaune, R.D.; Patrick, W.H., Jr. Assessment of saltwater intrusion impact on gas exchange behavior of Louisiana Gulf Coast wetland species. Wetl. Ecol. Manag. 1989, 1, 21–30. [Google Scholar] [CrossRef]
- Keim, R.F.; Dean, T.J.; Chambers, J.L.; Conner, W.H. Stand density relationships in baldcypress. For. Sci. 2010, 56, 336–343. [Google Scholar]
- Reineke, L.H. Perfecting a stand density index for even-aged forests. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]
- Rybczyk, J.M.; Cahoon, D.R. Estimating the potential for submergence for two wetlands in the Mississippi River Delta. Estuaries 2002, 35, 985–998. [Google Scholar] [CrossRef]
- Brantley, C.G.; Day, J.W.; Lane, R.R.; Hyfield, E.; Day, J.N.; Ko, J.Y. Primary production, nutrient dynamics, and accretion of a coastal freshwater forested wetland assimilation system in Louisiana. Ecol. Eng. 2008, 34, 7–22. [Google Scholar] [CrossRef]
- Hunter, R.G.; Shaffer, G.P.; Lane, R.R.; Day, J.W. Analysis of Long-Term Productivity and Nutrient Removal at Assimilation Wetlands in Coastal Louisiana; LDEQ OC No. 855–400144: Baton Rouge, LA, USA, 2015; p. 98. [Google Scholar]
- Shaffer, G.; Day, J.; Hunter, R.; Lane, R.; Lundberg, C.; Wood, B.; Hillmann, E.; Day, J.; Strickland, E.; Kandalepas, D. System response, nutria herbivory, and vegetation recovery of a wetland receiving secondarily-treated effluent in coastal Louisiana. Ecol. Eng. 2015, 79, 120–131. [Google Scholar] [CrossRef]
- Effler, R.S.; Goyer, R.A.; Lenhard, G.J. Baldcypress and water tupelo responses to insect defoliation and nutrient augmentation in Maurepas Swamp, Louisiana, USA. For. Ecol. Manag. 2006, 236, 295–304. [Google Scholar] [CrossRef]
- Brown, C.A.; Montz, G.N. Baldcypress and the Tree Unique, the Wood Eternal; Claitor’s Publishing Division: Baton Rouge, LA, USA, 1986; p. 139. [Google Scholar]
- Conner, W.H.; Gosselink, J.G.; Parrondo, R.T. Comparison of the vegetation of three Louisiana swamp sites with different flooding regimes. Am. J. Bot. 1981, 68, 320–331. [Google Scholar] [CrossRef]
- Hesse, I.D.; Day, J.W.; Doyle, T.W. Long-term growth enhancement of baldcypress (Taxodium distichum) from municipal wastewater application. Environ. Manag. 1998, 22, 119–127. [Google Scholar] [CrossRef]
- Day, J.; Ko, J.Y.; Rybczyk, J.; Sabins, D.; Bean, R.; Berthelot, G.; Brantley, C.; Breaux, A.; Cardoch, L.; Conner, W.; et al. The use of wetlands in the Mississippi Delta for wastewater assimilation: A review. Ocean Coast. Manag. 2004, 47, 671–691. [Google Scholar] [CrossRef]
- Hillmann, E.R. The Implications of Nutrient Loading on Deltaic Wetlands. Master’s Thesis, Southeastern Louisiana University, Hammond, LA, USA, 2011. [Google Scholar]
- Carter, M.R.; Burns, L.A.; Cavinder, T.R.; Dugger, K.R.; Fore, P.L.; Hicks, D.B.; Revills, H.L.; Schmidt, T.W. Ecosystems Analysis of the Big Cypress Swamp and Estuaries; USEPA Region IV, South Florida Ecological Study: Miami, FL, USA, 1973. [Google Scholar]
- Conner, W.H.; Duberstein, J.A.; Day, J.W., Jr.; Hutchinson, S. Forest community changes along a flooding/elevation gradient in a south Louisiana forested wetland from 1986–2009. Wetlands 2014, 34, 803–814. [Google Scholar] [CrossRef]
- Megonigal, J.P.; Conner, W.H.; Kroeger, S.; Sharitz, R.R. Aboveground production in southeastern floodplain forests: A test of the subsidy-stress hypothesis. Ecology 1997, 78, 370–384. [Google Scholar] [CrossRef]
- Schlesinger, W.H. Community structure, dyes, and nutrient ecology in the Okenfenokee cypress swamp-forest. Ecol. Mono. 1978, 48, 43–65. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Day, J.W., Jr.; Gilliam, J.W.; Groffman, P.M.; Hey, D.L.; Randall, G.W.; Wang, N. Reducing Nutrient Loads, Especially Nitrate–Nitrogen, to Surface Water, Groundwater, and the Gulf of Mexico; Topic 5 Report for the Integrated Assessment on Hypoxia in the Gulf of Mexico. NOAA Coastal Ocean Program Decision Analysis Series No. 19; NOAA Coastal Ocean Program: Silver Spring, MD, USA, 1999; p. 111. [Google Scholar]
- Taylor, J.R. Community Structure and Primary Productivity of Forested Wetlands in Western Kentucky. Ph.D. Thesis, University of Louisville, Louisville, KY, USA, 1985; p. 139. [Google Scholar]
- U.S. Army Corps of Engineers. Louisiana Coastal Protection and Restoration Final Technical Report; USACE: New Orleans, LA, USA, 2009; p. 94. [Google Scholar]
- Hunter, R.G.; Day, J.W., Jr.; Lane, R.R.; Lindsey, J.; Day, J.N.; Hunter, M.G. Nutrient removal and loading rate analysis of Louisiana forested wetlands assimilating treated municipal effluent. Environ. Manag. 2009, 44, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, G.P.; Day, J.W., Jr. Use of Freshwater Resources to Restore Baldcypress–Water Tupelo Swamps in the Upper Lake Pontchartrain Basin; White Paper; Louisiana Department of Wildlife and Fisheries: Baton Rouge, LA, USA, 2007; p. 44. [Google Scholar]
- Nyman, J. Integrating successional ecology and the delta lobe cycle in wetland research and restoration. East Coasts 2014. [Google Scholar] [CrossRef]
- Paola, C.; Twilley, R.R.; Edmonds, D.A.; Kim, W.; Mohrig, D.; Parker, G.; Viparelli, E.; Voller, V.R. Natural processes in delta restoration: Application to the Mississippi delta. Annu. Rev. Mar. Sci. 2010, 3, 67–91. [Google Scholar] [CrossRef] [PubMed]
- Twilley, R.R.; Rivera-Monroy, R. Sediment and nutrient tradeoffs in restoring Mississippi river delta: Restoration vs. eutrophication. Cont. Water Resour. Ed. 2009, 141, 39–44. [Google Scholar] [CrossRef]
- Lee Wilson and Associates, Inc. Diversion into the Maurepas Swamps. A complex project under the Coastal Wetlands Planning, Protection, and Restoration Act; Final report prepared for the U.S.; Environmental Protection Agency, Region 6: Dallas, TX, USA, 2001; p. 59, Contract No. 68-06-0067 WA#5-02. [Google Scholar]
- Condrey, R.E.; Hoffman, P.E.; Evers, D.E. The Last Naturally Active Delta Complexes of the Mississippi River (LNDM): Discovery and Implications. In Perspectives on the Restoration of the Mississippi Delta; Day, J.W., Kemp, G.P., Freemen, A.M., Muth, D.P., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 33–50. [Google Scholar]
- Davis, D.W. Crevasses on the lower course of the Mississippi River. CoastZone 1993, 1, 360–378. [Google Scholar]
- Welder, F.A. Processes of Deltaic Sedimentation in the Lower Mississippi River; Louisiana State University, Coastal Studies Institute Technical Report: Baton Rouge, LA, USA, 1959; pp. 1–90. [Google Scholar]
- Bentley, S.J.; Freeman, A.M.; Wilson, C.S.; Cable, J.E.; Giosan, L. Using What We Have: Optimizing Sediment Management in Mississippi River Delta Restoration to Improve the Economic Viability of the Nation. In Perspectives on the Restoration of the Mississippi Delta; Day, J.W., Kemp, G.P., Freemen, A.M., Muth, D.P., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 85–97. [Google Scholar]
- Nittrouer, J.A.; Best, J.L.; Brantley, C.; Cash, R.W.; Czapiga, M.; Kumar, P.; Parker, G. Mitigating land loss in coastal Louisiana by controlled diversion of Mississippi River sand. Nat. Geosci. 2012, 5, 534–537. [Google Scholar] [CrossRef]
- Friedlingstein, P.; Andrew, R.M.; Rogelj, J.; Peters, G.P.; Canadell, J.D.; Knutti, R.; Luderer, G.; Raupach, M.R.; Schaeffer, M.; van Vuuren, D.P.; et al. Persistent growth of CO2 emissions and implications for reaching climate targets. Nat. Geosci. 2014, 7, 709–715. [Google Scholar] [CrossRef]
- Myers, R.S.; Shaffer, G.P.; Llewellyn, D.W. Baldcypress (Taxodium distichum (L.) Rich.) restoration in southeast Louisiana: The relative effects of herbivory, flooding, competition, and macronutrients. Wetlands 1994, 15, 141–148. [Google Scholar] [CrossRef]
- Beville, S.L. The Efficacy of a Small-Scale Freshwater Diversion for Restoration of a Swamp in Southeastern Louisiana. Master’s Thesis, Southeastern Louisiana University, Hammond, LA, USA, 2002; p. 86. [Google Scholar]
- Lundberg, C.J. Using Secondarily Treated Sewage Effluent to Restore the Baldcypress-water Tupelo Swamps of the Lake Pontchartrain basin: A Demonstration Study. Master’s Thesis, Southeastern Louisiana University, Hammond, LA, USA, 2008; p. 85. [Google Scholar]
- Lundberg, C.J.; Shaffer, G.P.; Wood, W.B.; Day, J.W., Jr. Growth rates of baldcypress (Taxodium distichum) seedlings in a treated effluent assimilation marsh. Ecol. Eng. 2011, 37, 549–553. [Google Scholar] [CrossRef]
- Lowery, G.H., Jr. The Mammals of Louisiana and its adjacent Waters; Louisiana State University Press: Baton Rouge, LA, USA, 1974. [Google Scholar]
- Blair, R.M.; Langlinais, M.J. Nutria and swamp rabbits damage baldcypress seedlings. J. For. 1960, 58, 388–389. [Google Scholar]
- Goyer, R.A.; Lenhard, C.G. A new insect pest threatens baldcypress. Agricultura 1988, 31, 16–17. [Google Scholar]
- Goyer, R.A.; Chambers, J. Evolution of Insect Defoliation in Baldcypress and its Relationship to Flooding; USDI. N BS. Biological Sciences Report 8: Lafayette, LA, USA, 1996. [Google Scholar]
- Nachod, L.H.; Kucera, D.R. Observations of the Forest Tent Caterpillar in South Louisiana; Insect and Disease Report; Louisiana Office of Forestry: Woodworth, LA, USA, 1971; p. 2. [Google Scholar]
- Nachod, L.H. Spring Defoliation by Forest Insects in Louisiana; Insect and disease Report; Louisiana Office of Forestry: Woodworth, LA, USA, 1977; p. 2. [Google Scholar]
- Allen, J.A.; Conner, W.H.; Goyer, R.A.; Chambers, J.L.; Krauss, K.W. Freshwater Forested Wetlands and Global Climate Change. In Vulnerability of Coastal Wetlands in the Southeastern United States; Biological Science Report USGS/BRD/BSR-1998–0002: Honolulu, HI, USA, 1998. [Google Scholar]
- Souther-Effler, R.F. Interactions of Insect Herbivory and Multiple Abiotic Stress Agents on Two Wetland Tree Species in Southeast Louisiana Swamps. Ph.D. Thesis, Louisiana State University and Agricultural and Mechanical College, Baton Rouge, LA, USA, 2004. [Google Scholar]
- Touliatos, P.; Roth, E. Hurricanes and trees—Ten lessons from Camille. J. For. 1971, 69, 285–289. [Google Scholar]
- Gresham, C.A.; Williams, T.M.; Lipscomb, D.J. Hurricane Hugo wind damage to southeastern U.S. Coastal forest tree species. Biotropica 1991, 23, 420–426. [Google Scholar] [CrossRef]
- Putz, F.E.; Sharitz, R.R. Hurricane damage to old-growth forest in Conaree Swamp National Monument. Can. J. For. Res. 1991, 21, 1765–1770. [Google Scholar] [CrossRef]
- Doyle, T.W.; Keeland, B.D.; Gorman, L.E.; Johnson, D.J. Structural impact of Hurricane Andrew on forested wetlands of the Atchafalaya Basin in south Louisiana. JCR 1995, 21, 354–364. [Google Scholar]
- Williams, K.; Pinzon, Z.S.; Stumpf, R.P.; Raabe, E.A. Sea-Level Rise and Coastal Forests on the Gulf of Mexico; USGS-99-441; U.S. Department of the Interior: Washington, DC, USA, 1999; p. 63. [Google Scholar]








© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaffer, G.P.; Day, J.W.; Kandalepas, D.; Wood, W.B.; Hunter, R.G.; Lane, R.R.; Hillmann, E.R. Decline of the Maurepas Swamp, Pontchartrain Basin, Louisiana, and Approaches to Restoration. Water 2016, 8, 101. https://doi.org/10.3390/w8030101
Shaffer GP, Day JW, Kandalepas D, Wood WB, Hunter RG, Lane RR, Hillmann ER. Decline of the Maurepas Swamp, Pontchartrain Basin, Louisiana, and Approaches to Restoration. Water. 2016; 8(3):101. https://doi.org/10.3390/w8030101
Chicago/Turabian StyleShaffer, Gary P., John W. Day, Demetra Kandalepas, William B. Wood, Rachael G. Hunter, Robert R. Lane, and Eva R. Hillmann. 2016. "Decline of the Maurepas Swamp, Pontchartrain Basin, Louisiana, and Approaches to Restoration" Water 8, no. 3: 101. https://doi.org/10.3390/w8030101
APA StyleShaffer, G. P., Day, J. W., Kandalepas, D., Wood, W. B., Hunter, R. G., Lane, R. R., & Hillmann, E. R. (2016). Decline of the Maurepas Swamp, Pontchartrain Basin, Louisiana, and Approaches to Restoration. Water, 8(3), 101. https://doi.org/10.3390/w8030101