1. Introduction
One of the greatest human-induced pressures on freshwater fish worldwide is the disruption of river connectivity, which can affect passage for many migratory species [
1,
2,
3]. Culverts pose one such connectivity barrier, and continue to be constructed in ways that affect fish passage worldwide [
4,
5]. Aside from disrupting downstream migrations through behavioural deterrents [
6,
7], culvert pipes can restrict upstream fish passage through perching [
8] and/or by creating a velocity barrier if within-pipe water velocity is beyond the swimming capability of the species seeking upstream passage [
9,
10]. In addition to perching and high within-pipe velocities, passage success can also be strongly influenced by culvert length and/or gradient, and various interacting combinations of these factors [
11,
12].
While provision of fish passage through culverts has been investigated with a variety of methods, e.g., [
13,
14], designs or retrofits can be expensive and are often overlooked in favour of less costly options [
4]. Common designs adopted in larger culverts involve the installation of internal weirs and/or baffles which reduce water velocities and provide resting areas for fish, e.g., [
13]. This is not possible in long, small diameter culverts because of logistical issues related to accessing internal sections for retrofitting, maintenance concerns, and/or the potential to compromise hydraulic capacity [
15].
In a recent study, David
et al. [
8] found that upstream migratory post-larval banded kokopu
Galaxias fasciatus (Gray 1842) were successful in climbing vertical mussel spat ropes, overcoming a simulated perched culvert in laboratory trials. It was concluded that mussel spat rope may offer a cost effective solution for enabling the passage of “climbing” fish species past and through perched culverts. Within culverts, ropes may facilitate fish passage for “climbing” species by increasing the potential wetted margin [
16], and for “swimming” species by reducing barrel water velocities and increasing hydraulic complexity [
17]. In this laboratory study we investigate the potential for mussel spat ropes to improve passage of redfin bully
Gobiomorphus huttoni (Ogilby 1894) of various life-stage through culvert pipes where uniform barrel velocities and pipe gradient were expected to restrict passage for this species.
2. Methods
G. huttoni (N = 150, size = 24–73 mm), a widespread amphidromous New Zealand eleotrid fish, were sourced (August 2011) from a 200 m stretch of the Uretara Stream, Katikati, Bay of Plenty, New Zealand (37°57' S, 175°91' E). All fish were captured with a battery powered backpack electric fishing machine (Kainga EFM300, NIWA Instrument Systems, Christchurch, New Zealand). Fish were held for two weeks prior to running trials and were held within a 900 L holding tank for the duration of the trials and fed at the end of each day.
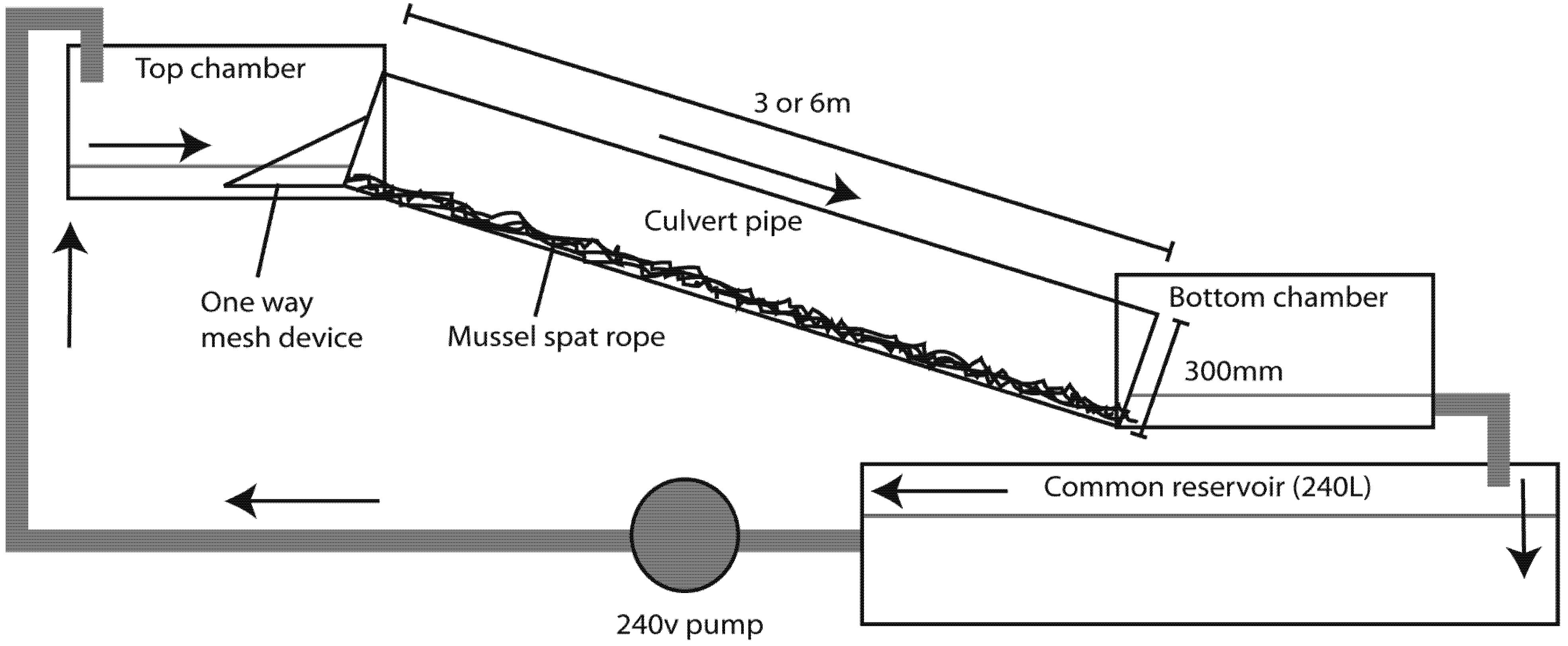
An experimental, simulated culvert system (
Figure 1) was constructed in the laboratory consisting of two trial setups of different lengths of 350 mm diameter culvert pipe; 3 m and 6 m. Although there are a range of culvert pipe sizes used in field situations (often larger than those used here), pipes of this size and smaller are common across New Zealand [
18]. Each trial ran on a pair-wise basis consisting of two pipes; a control pipe (no rope) and a treatment pipe, which was fitted with two parallel, equal lengths of mussel spat rope (used for the collection of mussel spat for aquaculture). The mussel spat rope used, “Super Xmas Tree” (14 mm core, Donaghys Industries, Christchurch, New Zealand), was identical to that used by David
et al. [
8]. While strands of rope project from the inner core increasing the total surface area, the inner core of individual ropes represent 0.16% of the internal area of each pipe. Each pipe conveyed water from an upper chamber down a 10° slope to a lower chamber. Two rocks were placed in each of the bottom chambers to provide resting areas for fish. A cone-shaped mesh device was installed at the top of each culvert to retain any fish that had successfully ascended the setup.
A re-circulating system driven by a 240 V Onga Pump (Model 413, Pentair Water New Zealand Ltd., Rongotea, New Zealand) delivered water from a common 240 L reservoir, to the top of each roped and un-roped trial chamber. Flow volume was equivalent and consistent between control and treatment for each trial (3 m trial = 0.9 ± 0.008 L s−1, t = 0.49, p = 0.63; 6 m trial = 1.04 ± 0.02 L s−1, t = 0, p = 1) and throughout all replicates. A constant depth of ~90–100 mm was maintained in each bottom chamber by recycling water from the lower chambers through a 50 mm drainage outlet at a consistent point on all four chambers, and re-delivering it to the common reservoir. At the end of each day reservoir water was flushed with clean, treated, tap water, and left at least 12 hour before the next trial. Velocities were assessed by measuring the time taken for dye to travel the length of pipes and converted to m s−1, which provides a simple index of velocity within the pipe and enables comparison between treatments. Water temperatures were unregulated throughout the trial and were measured at the beginning and end of each trial in the bottom chambers using an Eutech Instruments ECScan pocket meter.
Figure 1.
Schematic diagram of experimental culvert system assessing the effects of mussel spat rope on passage success of redfin bully Gobiomorphus huttoni. Figure illustrates one of the treatment systems with mussel spat rope attached. Each 3 or 6 m length was paired with treatment (rope) and control systems which were run through trials without rope. Arrows indicate direction of flow.
Figure 1.
Schematic diagram of experimental culvert system assessing the effects of mussel spat rope on passage success of redfin bully Gobiomorphus huttoni. Figure illustrates one of the treatment systems with mussel spat rope attached. Each 3 or 6 m length was paired with treatment (rope) and control systems which were run through trials without rope. Arrows indicate direction of flow.
Two replicates were run each day (am/pm) between 7 am and 5 pm, each running for three hours. Twenty trials (10 m × 3 m; 10 m × 6 m) were completed over a three-week period, from late August to mid-September 2011. Sixty fish (30 individuals each of control or treatment) of variable size were used in each replicate, having been randomly selected from the common holding tank. Fish were used once per day. Fish were recorded either as reaching the top, being in ascent, or remaining in the bottom chamber at the end of each three-hour trial. To assess whether fish size was important for passage, the total length of each fish in their respective position post-trial (i.e., bottom, in ascent, top) was measured for the 6 m trials.
All statistical analyses were carried out on raw data after testing for normality (Shapiro-Wilks:
W = 0.97,
p = 0.12). Where displayed, means are accompanied by one standard error. One-way ANOVA was performed in R 2.13.1 [
19] to determine differences in the number (all trials) and size of fish (6 m trials) between the three positions post-trial. Tukey’s HSD criterion was used post-hoc to assess pair-wise differences between the three positions. Linear regression was performed in R 2.13.1 [
19] to assess whether temperature changed over the duration of the trial period. Two-tailed t-tests were performed on treatment-pipe data to compare ascent success and temperature between 3 m and 6 m trials.
3. Results and Discussion
No fish successfully ascended, or were found to be in ascent of the un-roped (control) 3 m or 6 m culverts, compared to varying success rates in the treatment pipes. High water velocity was assumed to be the main factor limiting passage success. Mean water velocities were slower in the pipes fitted with mussel spat rope (0.68 ± 0.03 ms
−1) than without (1.76 ± 0.04 ms
−1;
t = 22.44,
p < 0.0001) and clear differences were visually evident of the effect of mussel spat rope on the hydrodynamics between the two treatments (
Figure 2). Observations during trials indicated that fish were unable to sustain prolonged bursts of swimming in pipes without ropes. These findings agree with previous work which suggested that in non-retrofitted situations, water velocities were the primary factor limiting the successful ascent of inanga
Galaxias maculatus (Jenyns 1842) and
G. huttoni [
16]. Conversely, in rope treatments, fish were easily able to access the pipes and at times could be observed resting at the base of the pipe and beneath the ropes prior to their ascent. The lower velocities exiting roped culverts may also reduce potential for downstream scouring which typically leads to culvert perching [
8].
Figure 2.
Different flow hydrodynamics between (a) control (no rope); and (b) treatment (rope) pipes, used to test the effect of mussel spat rope on redfin bully Gobiomorphus huttoni passage through simulated laboratory culverts.
Figure 2.
Different flow hydrodynamics between (a) control (no rope); and (b) treatment (rope) pipes, used to test the effect of mussel spat rope on redfin bully Gobiomorphus huttoni passage through simulated laboratory culverts.
The mean proportion of fish reaching the top chamber for the combined (3 m and 6 m) roped trials was greater than the proportion of fish found either in ascent or remaining in the bottom chamber (
Table 1), which Tukey’s HSD criterion indicated were not different from each other. There was a significantly greater success rate of fish passage in 6 m (0.69 ± 0.09) compared to 3 m (0.35 ± 0.11) roped trials (
t = −2.36,
p = 0.03), which was likely related to higher ambient and water temperatures in the 6m trials (
t = −2.44,
p = 0.025) as this increased over the study period (
F1,18 = 14.26,
p = 0.001,
R2 = 0.44,
y = 12.9 + 0.13
x). Previous research has shown that temperatures below 12 °C can reduce the migratory activity of fish compared to warmer water around 16 °C [
20]. Although fish learning in association with temperature may also have contributed to higher passage success in 6m roped trials in the current study, our design did not allow us to differentiate the relative importance of these two factors. Nevertheless, exploring and controlling each of these factors would be an interesting and informative area for future research.
Table 1.
Mean (±1 Standard Error) proportion of redfin bully Gobiomorphus huttoni in either the top chamber, ascending the culvert, or in the bottom chamber of 3 m and 6 m treatment pipes post trial, in a simulated culvert system with mussel spat ropes installed (n = 10 replicates of each length; 30 fish per replicate; control and treatment run for every replicate).
Table 1.
Mean (±1 Standard Error) proportion of redfin bully Gobiomorphus huttoni in either the top chamber, ascending the culvert, or in the bottom chamber of 3 m and 6 m treatment pipes post trial, in a simulated culvert system with mussel spat ropes installed (n = 10 replicates of each length; 30 fish per replicate; control and treatment run for every replicate).
| Pipe length | Top | On rope | Bottom | F | p | Degrees of freedom |
|---|
| 3 m | 0.35 (0.11) | 0.12 (0.03) | 0.52 (0.11) | 4.62 | 0.02 | 2, 27 |
| 6 m | 0.69 (0.09) | 0.26 (0.07) | 0.05 (0.02) | 24.45 | <0.0001 | 2, 27 |
| Combined | 0.52 (0.08) | 0.26 (0.04) | 0.28 (0.08) | 6.26 | 0.004 | 2, 57 |
Irrespective of the mechanism(s) allowing passage success between roped trials, it is important to recognise that not a single bully was able to successfully negotiate either the 3 or 6 m culverts without ropes present. Assumptions were made prior to the trial that re-use of
G. huttoni would not influence ascent success based on results of a similar trial involving re-use of this species on five occasions [
16]. Limited evidence from this study suggests “learning” may have been a factor in their success but this was confounded by a correlation with temperature. There was no evidence to indicate an effect of fish length [length (mm): top = 48.39 ± 0.41, middle = 46.99 ± 0.54, bottom = 49.95 ± 1.41;
F2,326 = 1.9,
p = 0.15]. The locomotory mode of
G. huttoni appears to be dependent on gradient [
16], and larger fish in general tend to swim rather than climb when ascending high velocity/gradient water flows [
21,
22]. Although observations of swimming mode were not possible, successful ascents of this fish throughout the size range used (24–73 mm) indicates that culverts fitted with spat ropes can facilitate passage for both migrant juveniles and non-migrant adults. However, more research would be required to assess whether these ropes would assist larger migratory fish such as adult salmonids.
When designing culverts, it is important to consider the passage requirements and locomotory style of a broad range of species [
23]. If retrofitting of culverts is required to provide for fish passage, the installation of mussel spat ropes may offer a relatively inexpensive solution compared to other methods such as baffles [
8,
15], particularly in long, difficult to access culverts such as those commonly found in urban areas. Ropes also have a low hydrodynamic profile which may be advantageous in situations where culvert capacity may be compromised by the retrofitting of other flow roughening devices [
15]. A recent field trial run over three years in a high energy stream on New Zealand’s east coast indicated no evidence of culvert blockage or retention of debris by ropes installed there [
17].
River fragmentation resulting from inappropriate crossing designs is a widespread international problem for diadromous fish [
3,
4]. The high success of
G. huttoni of varying size (and thus life-stage) through laboratory culverts with mussel spat rope present compared to no success in pipes without rope, suggests mussel spat rope could potentially be utilised by fish with varying locomotory styles. Trials assessing different configurations of rope (such as knotting), pipe length and gradient on passage of other swimming species would be a fruitful area for future research, as well as field testing their application in long culverts. These findings, along with those of David
et al. [
8] and David and Hamer [
17], imply that mussel spat ropes could be an effective and cost effective tool for improving passage through both perched and non-perched culvert barriers in New Zealand and worldwide.




{kind=link}
{kind=link}