The Effects of Antibiotics on Microbial Community Composition in an Estuary Reservoir during Spring and Summer Seasons

1
School of Environmental Science and Engineering, Shanghai Jiao Tong University, Shanghai 200240, China
2
NUS Environmental Research Institute (NERI), National University of Singapore, Singapore 138602, Singapore
3
Department of Civil and Environmental Engineering, National University of Singapore, Singapore 138602, Singapore
*
Author to whom correspondence should be addressed.
Water 2018, 10(2), 154; https://doi.org/10.3390/w10020154
Submission received: 23 November 2017
/
Revised: 24 December 2017
/
Accepted: 17 January 2018
/
Published: 6 February 2018
(This article belongs to the Special Issue Antimicrobial Resistance in Environmental Waters)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The increased antibiotic pollutants in aquatic environments pose severe threats on microbial ecology due to their extensive distribution and antibacterial properties. A total of 16 antibiotics including fluoroquinolones (FQs) (ofloxacin (OFX), ciprofloxacin (CFX), norfloxacin (NFX)), Sulfonamides (SAs) (sulfamonomethoxine (SMM), sulfadiazine (SDZ), sulfaquinoxaline (SQX)), Tetracyclines (TCs) (tetracycline (TC), doxycycline (DC)), β-lactams (penicillin G (PEN G), penicillin V (PEN V), cefalexin (LEX)), Macrolides (MLs) (erythromycin-H2O (ETM), tylosin (TYL)) and other antibiotics (Polymix-B (POL), Vancomycin (VAN), Lincomycin (LIN)) were detected in the surface water of the Qingcaosha Reservoir. Multivariate statistical analysis indicated that both water quality and physicochemical indexes have less contributions on variations of these antibiotics, suggesting the concentrations of antibiotics inside the reservoir are mainly affected by upstream runoff and anthropic activity along the river. Antibiotics including TYL, PEN G and ETM showed significant correlations with variations of bacterial community composition, and closely connected with various gram-negative bacteria in co-occurrence/exclusion patterns of the network, suggesting these bacterial taxa play important roles in the course of migration and transformation of related antibiotics. In conclusion, further research is required to evaluate the potential risk of genetic transfer of resistance to related bacteria induced by long-term exposure to low levels of antibiotics in the environment.
1. Introduction
Estuary reservoirs as important water sources for cities in estuarine areas of the world, are strongly influenced both by estuarine ecologic changes and anthropic activities in surrounding areas [1,2,3]. In recent years, due to the extensive use of antibiotics in the world, the ecological environments in estuarine areas are facing serious ecological threats. Research showed that almost 80% to 90% of these compounds were excreted into the environments after initial metabolisms in organisms [4,5]. Finally, high levels of antibiotics were discharged into estuarine ecosystems via river runoff and sewage outfalls from land-based multiple sources, which have negative effects on human health and the safety of estuarine ecosystems [6,7].
In estuarine aquatic ecosystems, bacterial communities play important roles during the microbial food web that recycles and consumes organic matters [8]. Additionally, studies indicate that the bacterial community composition in aquatic ecosystems was strongly affected by environmental factors such as changes of hydrological conditions and different water trophic statuses [9,10]. Although antibiotics in aquatic ecosystems have been widely studied in recent years, and much has been achieved in the fields of toxic effects of antibiotics on aquatic plants and animals [11,12,13], still less is known about how the antibiotics affect the bacterial community composition in aquatic ecosystems.
The Qingcaosha (QCS) Reservoir is the largest estuary reservoir of China, located at the Yangtze estuary near Shanghai. The reservoir covers a total area of 66.27 km2, with a depth ranging from 2.5 m to 13.5 m. The main function of the reservoir is compensating for drinking water shortages in Shanghai city, inputting water with high turbidity from the Yangtze River estuary and outputting clean water to the water plant after the self-purification inside the reservoir [14,15,16]. Recently, it has been reported that multi-antibiotics were detected in the Yangtze estuary, even in the tap water of Shanghai [17,18,19]. However, still only few studies have discussed the presence of antibiotics inside the QCS Reservoir, especially the influences of these compounds on bacterial community changes, as well as the effect on co-occurrence patterns of cyanobacterial and non-cyanobacterial taxa within the aquatic ecosystem.
This study aims to utilise a systematic method that encompasses the measurement of water quality, antibiotic concentrations and bacterial community composition data sets to evaluate the relationships between water environmental parameters, concentration levels of antibiotics and bacterial community variations in the QCS Reservoir. Especially, it aims to determine how these antibiotics affect bacterial community composition (including co-occurrence patterns between cyanobacteria and non-cyanobacterial taxa) in the estuary aquatic ecosystem during spring and summer season.
2. Materials and Methods
2.1. Sampling Sites and In Situ Measurements
Three locations, sites 1 to 3, were selected as sampling points in the reservoir representing inlet, internal and exit conditions, respectively (Figure S1). To better understand the spatial (sites) and temporal (months) variations of antibiotic concentrations, as well as their co-occurrence pattern with and impact on bacteria and cyanobacteria in surface water of this estuary reservoir during the warm season (from spring to summer) [20,21], we collected water samples from April to September 2014 at a depth of 0.5 m below the surface. All water samples were collected in triplicate, stored in sterile polypropylene bottles and processed immediately upon arrival at the laboratory. For physical and chemical detections (water quality indices, physiochemical parameters and antibiotics), we processed and measured two technical replicates for each of the biological replicates; while for high-throughput sequencing, triplicate samples for each site were combined by equal volume and processed for sequencing.
2.2. Physic-Chemical Parameters and Environmental Factors
Water quality and chemical parameters including water temperature, dissolved oxygen (DO) and pH were measured in situ by using a multi-parameter water quality analyzer (Multi3410, WTW Company, Weilheim, Germany). Other physic-chemical parameters include electrical conductivity (EC), turbidity, total phosphorus (TP), total nitrogen (TN), total carbon (TC), total organic carbon (TOC), inorganic carbon (IC) and ammonium nitrogen (NH4+-N) were analyzed according to standard methods for water and wastewater monitoring analysis [22]. The concentrations of chlorophyll-a (chl-a), which represented phytoplankton biomass, were measured using a Phytoplankton Analyzer (PHYTO-PAM) (Heinz Walz GMBH, Effeltrich, Germany) [23]. After filtering samples through 0.45 μm cellulose acetate membranes, the concentrations of K+, Na+, Ca2+, Mg2+, Al3+ and Si4+ ions were detected by inductively coupled plasma (ICP) spectroscopy [15]. The F−, Cl− and SO42− ions contents were detected using Metrohm 830 Ion Chromatography (Metrohm AG, Herisau, Switzerland) [15].
2.3. Antibiotics Detection and Analysis
Based on the results of our preliminary experiment within QCS Reservoir in an earlier stage, we referred to related researches about antibiotics in the Yangtze estuary [17,18,19], and antibiotics including the fluoroquinolones (FQs) (such as ofloxacin (OFX), ciprofloxacin (CFX), norfloxacin (NFX)), Sulfonamides (SAs) (such as sulfamonomethoxine (SMM), sulfadiazine (SDZ), sulfaquinoxaline (SQX)), Tetracyclines (TCs) (such as tetracycline (TC), doxycycline (DC)), β-lactams (such as penicillin G (ETM), penicillin V (PEN V), cefalexin (LEX)), Macrolides (MLs) (such as erythromycin-H2O (ETM), tylosin (TYL)) and other common antibiotics (such as Polymix-B (POL), Vancomycin (VAN), Lincomycin (LIN)) were selected as target compounds in our research. The antibiotic standards and internal standards (purities of all the chemicals are >98%) were purchased from Dr. Ehrenstorfer GmbH (Augsburg, Germany), and the detection methods of antibiotics were according to our colleague Yue Jiang’s research, and the further information about preparation, extraction, sample detection and analysis can be found in the Supplementary Methods.
2.4. Genome DNA Extraction
For genomic Deoxyribonucleic acid (DNA) extraction, a total of 500 mL water sample was filtrated through 0.22 μm cellulose acetate filter membranes, and DNA was extracted directly from the same amount of membranes using a E.Z.N.A. ™ Water DNA Kit (OMEGA bio-tek, Houston, TX, USA) according to the manufacturer’s instruction.
2.5. The 16S rRNA Gene Sequencing via (Polymerase Chain Reaction) PCR Amplification
To determine the diversity and variability of the bacterial community at different sites of the reservoir, we ran PCR for 18 water samples using a primer set targeting the V4 region of the 16S rRNA gene. This primer set (515F (5′-GTGCCAGCMGCCGCGGTAA-3′) and 806R (5′-GGACTACVSGGGTATCTAAT-3′)) exhibited fewer biases and more accurate taxonomic and phylogenetic information for individual bacterial taxa [24]. The details of PCR amplification procedures can be found in Supplementary Methods.
After purification, the PCR product were sent to Personal Biotechnology Co., Ltd. (Shanghai, China) for paired-end amplicon sequencing (2 × 150 bp) using Illumina MiSeq (Illumina, San Diego, CA, USA). Raw data were processed according to procedures as described previously [25], using the Quantitative Insights into Microbial Ecology (QIIME) pipeline (version 1.7.0, http://qiime.org/) for quality control (Supplementary Methods). The raw sequencing dataset is available for download from NCBI Sequence Read Achieve under BioProject PRJNA397386.
2.6. Statistical Analysis
The average value and standard deviation of each biotic and abiotic parameter were calculated using Microsoft Excel 2016 (Microsoft, Redmond, WA, USA). Before the multivariate statistical analysis, the relative abundance of bacterial OTUs (Operational Taxonomic Units) were square-root transformed to reduce the disturbance of highly abundant OTUs in analysis progress, while the environmental data sets (water quality indices, physiochemical parameters and antibiotics) were normalized using means and standard deviations of the variables.
The similarity matrices of biological and physicochemical characteristics in samples were constructed based on Bray−Curtis similarity and Euclidean distance, respectively. RELATE-BEST was used to evaluate the relationships between environmental factors and bacterial OTUs, and distance-based linear models (DistLM) were created to model and assess the contribution of each of the environmental variables and antibiotics to variations of microbial community composition by using PRIMER v6 and PERMANOVA+ (PRIMER-E Ltd., Plymouth, UK) [26]. Furthermore, the nonmetric multidimensional scaling (NMDS) was conducted to investigate the temporal and spatial variations of bacterial community composition. Alpha-diversity including Chao1, Ace, Chao/Ace, Simpson and Shannon indices between samples were calculated using the observed number of OTUs [27].
Highly abundant OTUs, which contributed to >20% reads for at least four samples and ≥1% reads in all the samples, were selected for further correlation analysis. It is noted that among these selected bacterial OTUs, no alternation was made to the original abundance within any sample. The degree of correlations between environmental variables, antibiotics and highly abundant OTUs across the whole sampling period were calculated using Pearson’s correlation coefficient (r) and p-value with the rcor.test algorithm provided in the ltm package in R (version 3.4.2, https://www.r-project.org/) [28]. The correlations between OTU pairs were also determined similarly. During the analysis process, the false discovery rate was constant kept below 5% based on the Benjamini–Hochberg procedure [29]. Finally, significant correlations (positive/negative) were visualized using network generated by the Frucherman Reingold algorithm on the Gephi package (version 0.9.1, https://gephi.org/). Within the network, relevant topological and node/edge matrices including betweenness centrality, closeness centrality, degrees and hub values were also enumerated through the network analysis plug-in [30,31].
3. Results
3.1. Physic-Chemical Parameters and Environmental Factor in QCS Reservoir
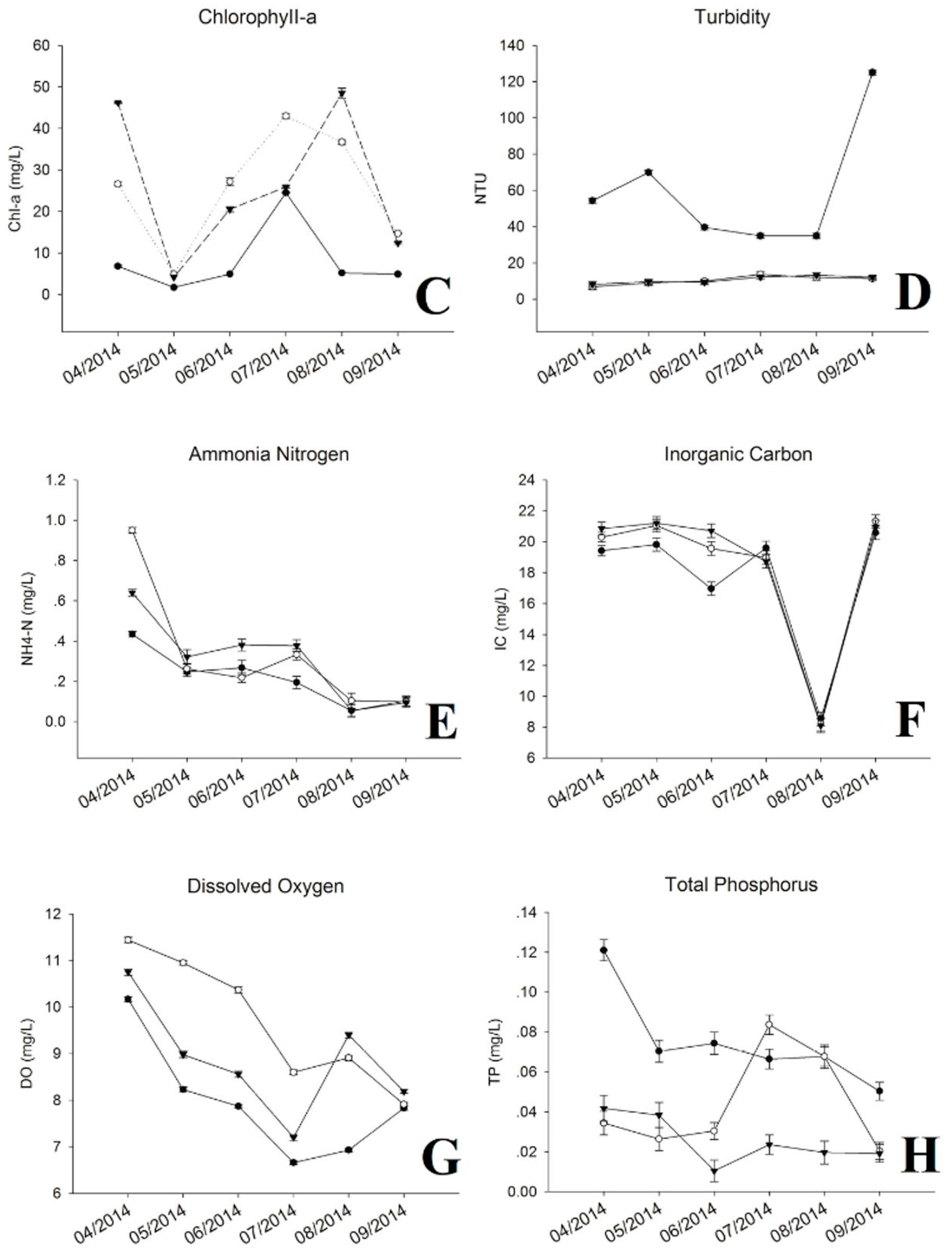
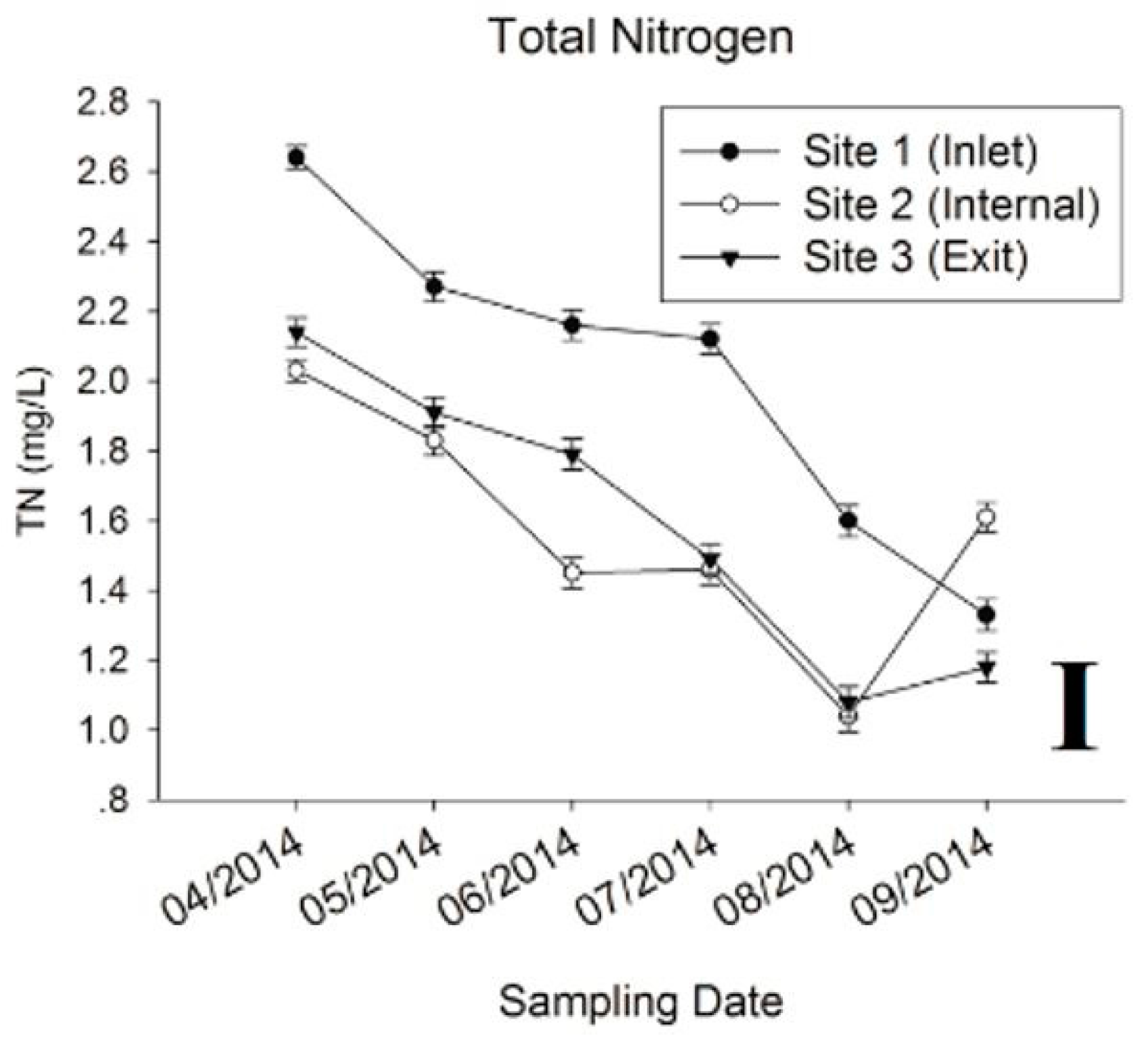
During the sampling period, water temperature varied from 15.3 to 29.1 °C, which increased rapidly from April to July, and decreased gradually from July to September (Figure 1A). The pH value showed repeated down and uptrends, where highest level was detected at the internal point from May to July, and at the exit point for other periods (Figure 1B). The concentration of chl-a presented a rapid decrease from April to May, then ascended with increased water temperature. The internal point showed the highest chl-a most of the sampling time except April and August (Figure 1C). Similar to the pH value, chl-a kept at the lowest level at the inlet site across the whole period compared with other two sites. The turbidity at internal and exit were relatively stable, ranging from 6.83 to 13.7, but higher readings were observed from August to September (34.9 to 125) at the inlet (Figure 1D). The concentrations of NH+4-N, IC, DO and TN exhibited decreased trends with increased water temperature (Figure 1E–I). Among these, levels of NH+4-N, IC, DO were higher at internal and exit sites, especially, for DO at internal point from April to July. Nutrients (TN and TP) concentrations of the inflow reduced significantly after flowing through the reservoir, except for TP, which increased and maintained at a higher level from July to August at internal point (Figure 1H,I). The variations of other environmental factors can be seen in Supplementary Materials (Table S1).
3.2. Antibiotics in QCS Reservoir
All 16 antibiotics including sulfonamides (SAs), fluoroquinolones (FQs), β-lactmas, macrolides (MLs), tetracyclines (TCs) and other common antibiotics were detected at three sampling sites throughout the sampling period (Figure S2). The concentrations of SAs exhibited an increasing trend from July to September at all three sites (Figure S2A–C), SDZ ranged from 0 to 48.90 ng/L, SMM ranged from 0 to 97.03 ng/L, and SQX ranged from 4.1 to 30.05 ng/L. The FQs revealed lower concentrations from May to July at all three sites (Figure S2D–F). Among these, NFX ranged from 48.01 to 193.84 ng/L, CFX ranged from 0 to 18.09 ng/L, and OFX ranged from 0 to 7.88 ng/L. In the group of β-lactmas, the concentration of LEX was very stable (average 14.43 ng/L) except at one time-point (July) (Figure S2G). In contrast, PEN G and PEN V revealed a large range of variation, which ranged from 0 to 133.50 ng/L and 21.7 to 157.76 ng/L, respectively (Figure S2H,I). The MLs including TYL and ETM exhibited lower concentrations across the whole sampling period, which ranged from 0 to 3.42 ng/L and 0 to 20.71 ng/L, respectively (Figure S2J,K). The concentrations of tetracyclines exhibited irregular variations in QCS Reservoir, TC and DC ranged from 4.11 to 26.69 ng/L and 20.67 to 170.97 ng/L, respectively (Figure S2L,M). The other common antibiotics such as POL ranged from 0 to 43.98 ng/L, VAN ranged from 0 to 9.50 ng/L, and LIN ranged from 0 to 9.30 ng/L (Figure S2N–P).
3.3. Dynamic Analysis of Bacterial Community Composition Based on the 16S rRNA Sequencing Data
Bacterial community composition analysis, as assessed by sequencing of V4 region of the 16S rRNA gene, identified a total of 5132 bacterial OTUs at genus level based on 97% similarity. The rarefaction curves indicated that the number of sequences were sufficient to cover the majority of species in the bacterial community within each sample (Figure S3). In QCS Reservoir, the dominant bacterial phyla identified were Proteobacteria (including α-, β- and γ-Proteobacteria, 31.3%), Actinobacteria (24.8%), Cyanobacteria (10.8%), Bacteroidetes (10.4%), Planctomycetes (8.2%), Verrucomicrobia (5.4%) and Chlorobi (2.2%) (Figure 2A). The other minor bacterial phyla (average abundance <2%) including Acidobacteria, Chloroflexi, Gemmatimonadetes, Firmicutes and Nitrospirae were also detected, in sum contributing less than six percent of total observed sequences.
Classification at level of class showed that eight major non-cyanobacterial taxa including α-, β-,γ-Proteobacteria, Acidimicrobiia, Actinobacteria, Flavobacteriia, Sphingobacteriia and OPB56 contributed more than 60% of total sequences on average in each site (Figure 2B). Thereinto, α- and β-Peoteobacteria had higher abundance at inlet site (16.5% and 16.4%, respectively) than other two sites (11.2% and 9.5% at internal site, 10.9% and 8.9% at exit site). But the γ-Proteobacteria appeared to represent an opposite trend with slightly higher abundance at internal and exit sites (5.9% and 4.8%, respectively), compared to the inlet site (3.7%). Different from Proteobacteria, the actinobacterial abundance was relatively stable at all three sites during the sampling period (average abundance ~15%). In contrast, a waving trend of Acidimicrobiia abundance was observed at internal and exit sites (internal: 4.3% to 12%; exit: 4.4% to 13%). In addition, the abundance of Flavobacteriia was obviously higher from April to June at both internal and exit sites (10.6% and 5.6%, respectively) compared to the inlet site (1.7%). The Synechococcophycideae, as the dominant taxa within cyanobacterial populations, varied from 1% to 30.5% of total sequences at internal and exit sites from July to September.
3.4. Covariance Analysis of Bacterial Community Composition and Environmental Variables
A marginal test of biotic and abiotic factors based on distance-based linear modeling (DistLM) identified 14 environmental factors (TN, NH4+-N, pH, DO, EC, turbidity, temperature, chl-a, K+, Na+, Ca2+, Mg2+, Cl− and F−) and four antibiotics (PEN G, TYL and ETM) which were significantly correlated with the variations of bacterial community composition (p < 0.05) (Tables S2 and S3). In addition, DistLM (best procedure and AICc criterion) indicated that most environmental variables had no significant effect on the concentrations of antibiotics except for the turbidity (p = 0.038) (Table S4).
Permutational MANOVA (9999 permutations) was calculated based on a reduced model across all 18 samplings in different sampling sites with time series, both significant temporal and spatial effects (p < 0.005) of environmental variables, antibiotic concentrations and bacterial community compositions were observed respectively between these three sites within QCS Reservoir (Table S5).
The richness and diversity (represented by Chao1/Ace and inverse Simpson index, respectively) of the bacterial community in each sample was calculated based normalized OTU abundance (Figure 3). Our findings showed that the inlet site had higher richness and diversity indices, while the internal and exit sites shared similar diversity but different from those found in the inlet especially in April. However, the diversity indices significantly decreased at all three sites in July, coinciding with the period of high cyanobacterial abundance. In contrast, the richness indices of samples were relatively stable over the time with a declining trend observed from inlet to exit.
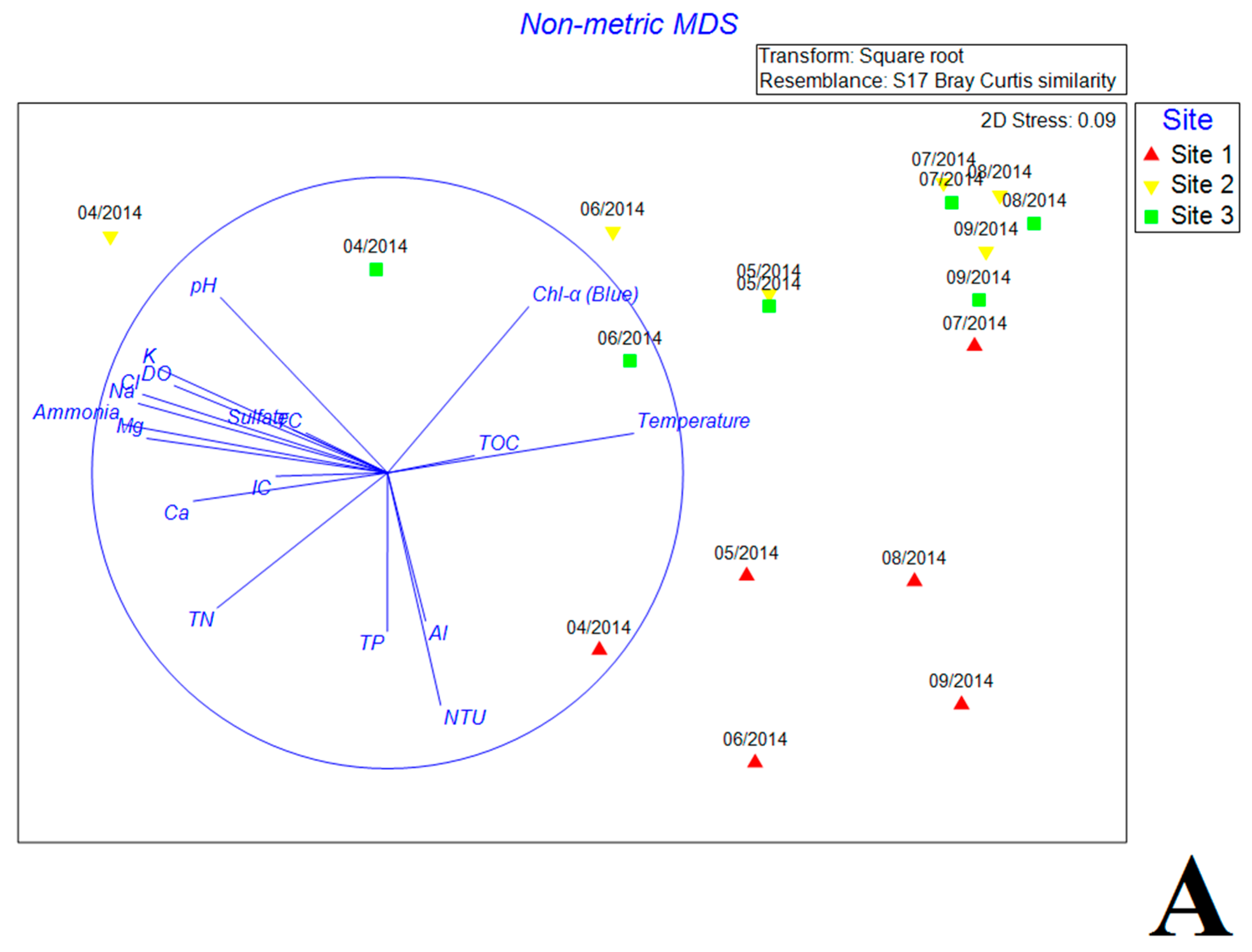
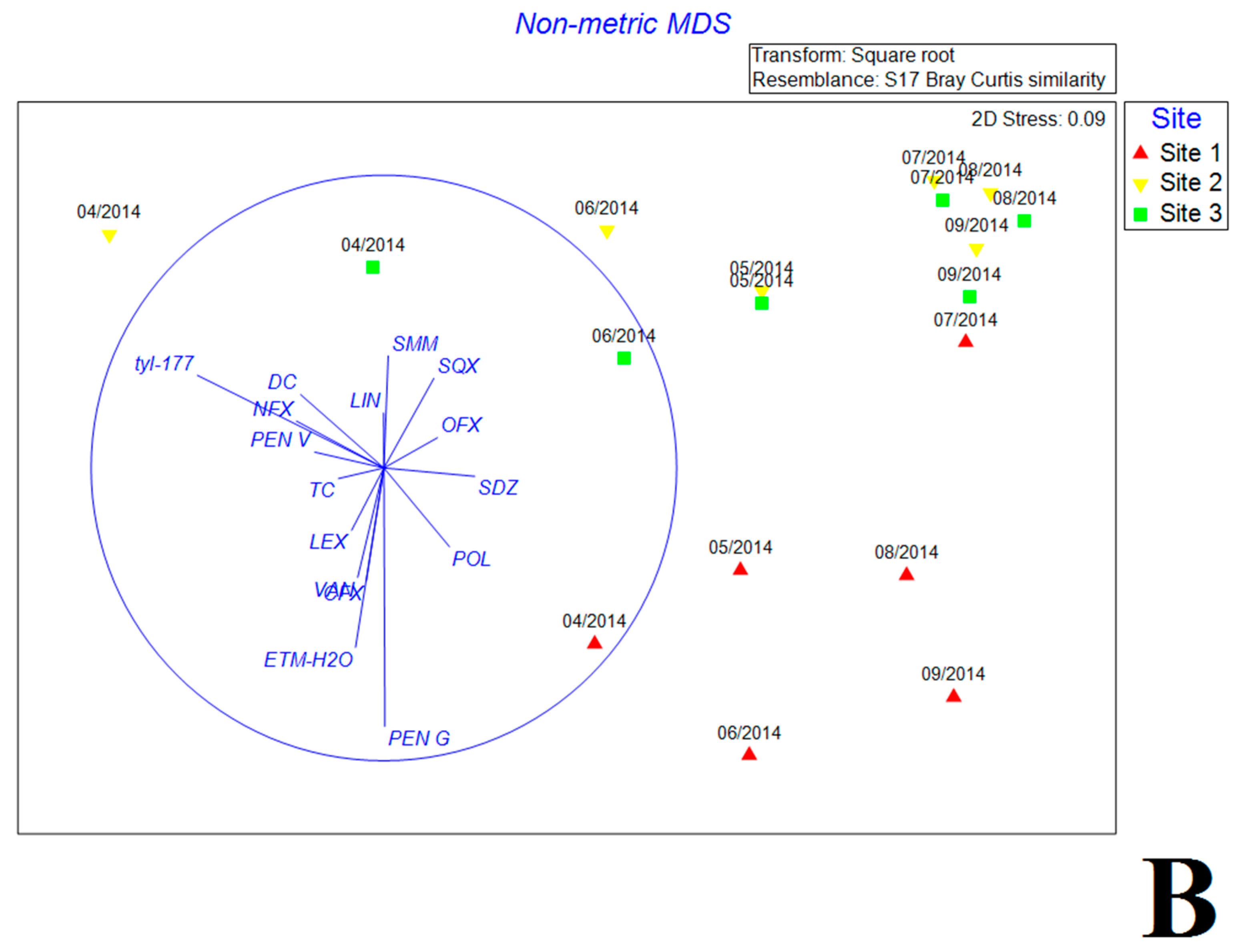
The NMDS plots based on Bray Curtis similarity showed significant distinction of bacterial community composition between inlet and the other two sites (Figure 4A,B). Most environmental factors exhibited close correlations with bacterial community variation (Figure 4A). Most samples (except samples in April at internal and exit sites) were significantly associated with high water temperature and TOC concentration. Samples of internal and exit sites collected from July to September gathered closely together and correlated with chl-a. In contrast, samples of inlet site showed scattered distribution, and associated with high turbidity, TP and Al3+. The distributions of antibiotics were different from the environmental factors, where PEN G and ETM were positively associated with the inlet samples while TYL exhibited positive correlation with internal and exit samples in April. Other antibiotics were not significantly correlated with changes of bacterial community (Figure 4B).
3.5. Multivariate Analysis of Biotic and Abiotic Factors in QCS Reservoir
A total of 99 variables, including 65 major OTUs (contributing more than one percent to any samples), 18 environmental variables and 16 antibiotics were shown in single interconnected network. A total of 4851 tested correlations were calculated by using rcor.test in the ltm package, with 474 ultimately considered significant (Table S6). Among them, 366 were positive correlations and 108 were negative. A visual correlation network was constructed with r score as the edge-weight, and values of betweenness centrality represented the size of nodes in the network (Figure 5A,B).
To illustrate the interactions among environmental variables, antibiotics and bacterial OTUs in QCS Reservoir, we explored the co-occurrence and co-exclusion patterns of these biotic/abiotic factors based on Pearson’s correlation coefficient (r). Totally, 366 pairs of positive correlations were identified from 44 genera, 15 environmental variables and 9 antibiotics. Meanwhile, 108 pairs of negative correlations were identified from 27 genera, 15 environmental variables and 2 antibiotics (Figure 5A,B). By modularity analysis, all nodes were separated into different modules in each network. Within each module, the nodes were connected closely by co-occurrence or co-exclusion patterns. Between different modules, each module was linked with other modules through the key nodes, which exhibited high betweenness centrality (>20%) in the network.
In the co-occurrence pattern network (Figure 5A), all nodes were separated into six modules including modules I (28.09%), II (21.35%), III (21.35%), IV (12.36%), V (11.24%) and VI (5.62%). Module I to III accounted for almost 71% of total network, which included 48 bacterial OTUs from 23 genera, 3 antibiotics (TYL, PEN G and ETM) and 12 environmental variables (NH4-N+, temperature, DO, TN, TP, turbidity, Chl-a, Mg2+, Na+, K+, Al3+ and Cl−). Module VI was only composed of five antibiotics (including SQX, NFX, PEN V, LIN and POL) without bacterial OTUs and other environmental variables. In module I, TYL exhibited significant and positive correlations with five environmental factors (including Mg2+, Na+, K+, Cl− and NH4-N+) and four bacterial OTUs (Comamonadaceae (β-Proteobacteria), calciphila (Bacteroidetes), Chitinophagaceae (Bacteroidetes) and Candidatus Xiphinematobacter (Verrucomicrobia)). In module II, PEN G showed positive correlations with two environmental variables (TP and turbidity), one antibiotic (ETM) and four bacterial OTUs (Holophagaceae (Acidobacteria), curvus (β-Proteobacteria), OPB56 (Chlorobi) and Nitrospira (Nitrospirae)). Additionally, ETM revealed positive correlations with PEN G and two bacterial OTUs belonged to curvus (β-Proteobacteria) and Rhodospirillaceae (α-Peoteobacteria). Within minor modules, antibiotic (TC) exhibited positive correlations with [Cerasicoccaceae] (Verrucomicrobia) and Fluviicola (Bacteroidetes) in module V, and SQX in module VI positively correlated with Cytophagaceae (Bacteroidetes) in module III.
In the co-exclusion pattern network (Figure 5B), all nodes were also separated into six modules—I (37.04%), II (35.19%), III (16.67%), IV (3.7%), V (3.7%) and VI (3.7%). Module I to III accounted for almost 89% of the total network, which included 34 bacterial OTUs from 26 genera, 1 antibiotic (PEN G) and 13 environmental variables (including temperature, DO, pH, turbidity, TN, TP, NH4-N+, Chl-a, K+, Na+, Ca2+, Mg2+ and Cl−). In module III, PEN G exhibited significant negative correlations with pH and Pirellulaceae (Planctomycetes). In addition, the antibiotic LEX revealed significant negative correlations with PHOS-HD29 (δ-Proteobacteria).
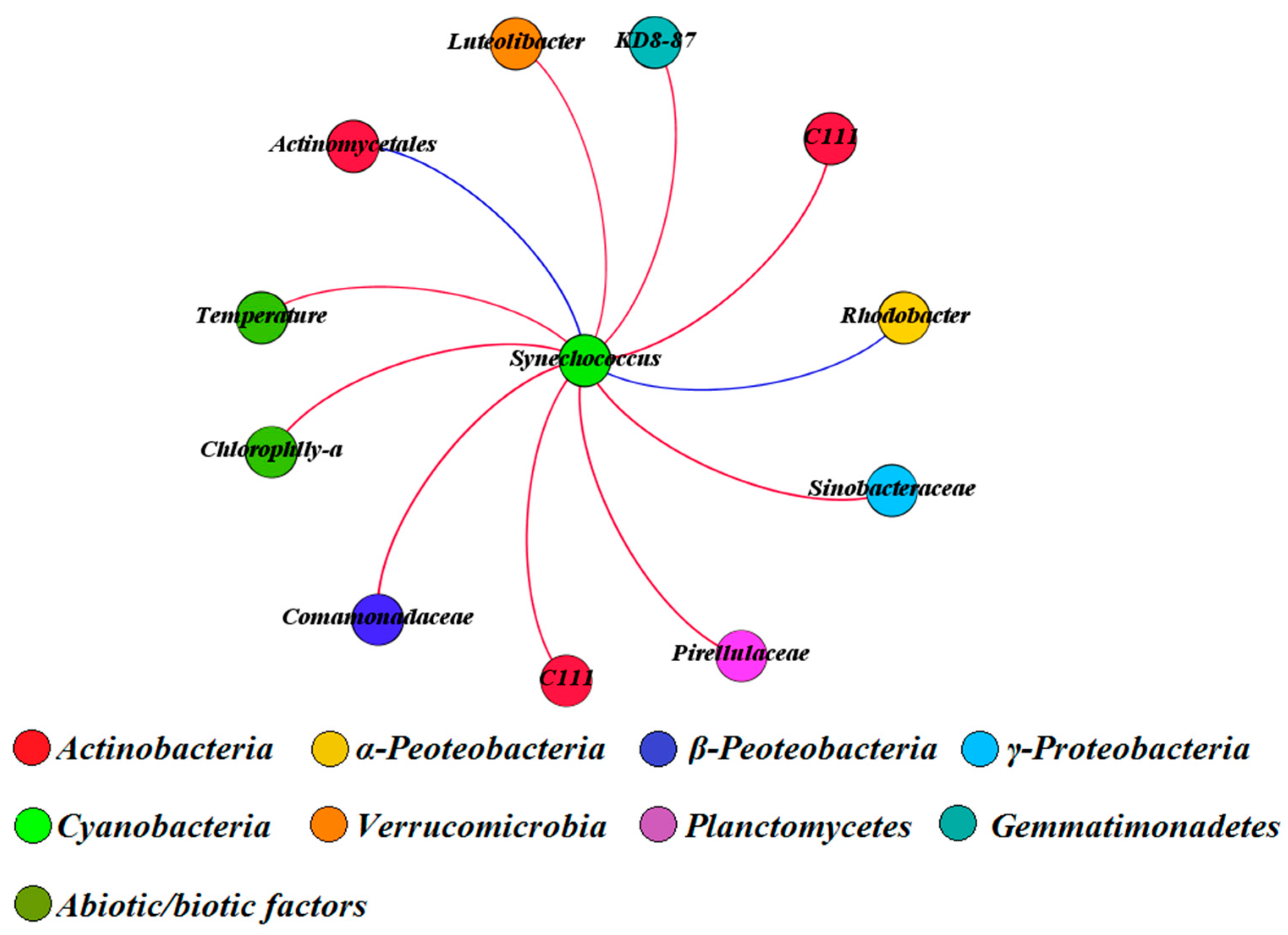
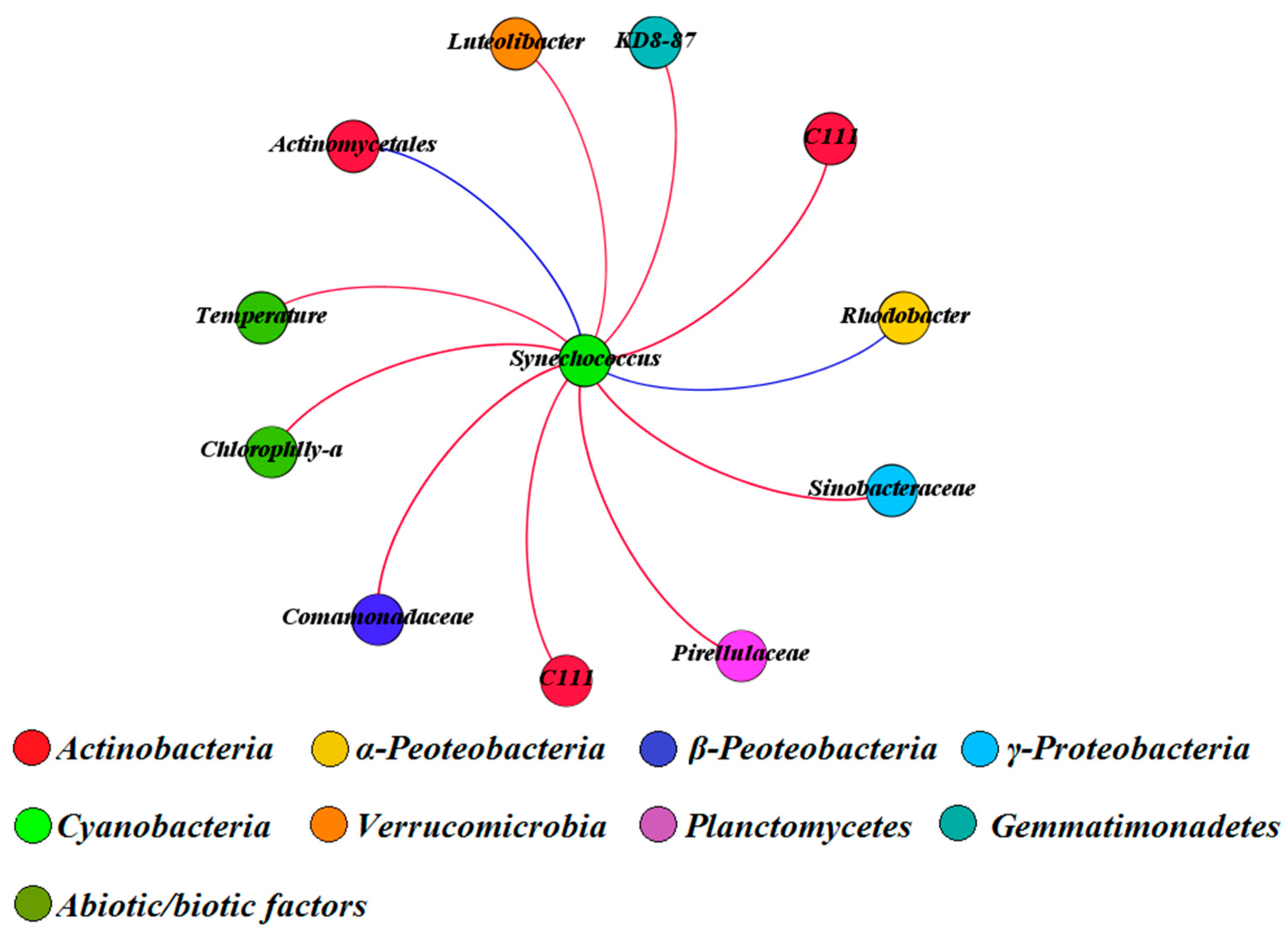
We further explored the co-occurrence and co-exclusion patterns of cyanobacterial and non-cyanobacterial OTUs within an organic correlation sub-network (Figure 6). Our results showed that temperature was the dominant environmental factor accelerating the proliferation of Synechococcus (dominant cyanobacterial taxa in QCS Reservoir). In addition, the concentration of chl-a, as well as several bacterial OTUs (such as C111 (Actinobacteria), Sinobacteraceae (γ-Proteobacteria), Comamonadaceae (β-Proteobacteria), Pirellulaceae (Planctomycetes), Luteolibacter (Verrucomicrobia) and KD8-87 (Gemmatimonadetes)) displayed significant and positive correlations with Synechococcus. In contrast, two bacterial OTUs belonged to Actinomycetales (Actinobacteria) and Rhodobacter (α-Peoteobacteria) revealed negative correlations with Synechococcus.
4. Discussion
Antibiotics as human and veterinary medicines are widely used in the prevention and treatment of diseases, and also as additives in livestock and breeding fields [17,18,32]. Research has indicated that the annual global consumption of antibiotics is about 150,000 tons, and almost 17% of the consumption is used in China [33]. With increasing use of antibiotics as medicines and animal growth promoters, plenty of these compounds have been released into aquatic environments, and pose direct/indirect threats on aquatic ecosystems [34]. In this study, we provided a systematic view on the relationships between environmental factors, antibiotics and bacterial community in an estuary aquatic ecosystem, and further explored the influences of antibiotics on co-occurrence/exclusion patterns of bacterial community.
4.1. Correlations between Environmental Parameters and Antibiotic Concentrations
In the present study, all sixteen antibiotics were detected at inlet, internal and exit of the QCS Reservoir from April to September (Figure S2). The distributions of these antibiotics varied temporally and spatially (Table S5). Apart from the LEX and TYL, all antibiotics exhibited increasing trend of different degrees from July to September (Figure S2), indicating a seasonal pattern of these compounds in the upstream area strongly polluted by agricultural and industrial activities and dense population along the Yangtze River [17]. We speculated that the inflow from the Yangtze River and WWTPs effluents were main sources of these antibiotics. This is supported by related research showing that the concentrations of antibiotics were mainly effected by upstream runoff and precipitation of Yangtze estuary [17,18,35].
Within these antibiotics, PEN G and TYL showed relative strong correlations with a variety of environmental variables (Figure 5A,B). This revealed that a huge concentration variation of PEN G and TYL were expected under different environmental conditions. Research also indicated that the process of water body self-purification further promoted the sedimentation effects of some antibiotics from water phase inside the reservoir [36], which is contributing to the decline of antibiotic concentrations in the surface water of the reservoir. Several antibiotics including SQX, NFX, PEN V, LIN and POL exhibited co-occurrence pattern and did not correlated to environmental factors (Figure 5A,B), which implied that these compounds might have similar changing patterns but were less affected by water quality and trophic status. Interestingly, although earlier research has indicated that the TOC have positively correlated with FQs concentrations in the sediment of the Yangtze estuary [18], we did not find any significant correlation between TOC and FQs concentrations in the surface water of the reservoir (Table S6) in our study. We speculate that the contradictory results could be attributed to the different metabolic transformation mechanisms of FQs between the surface water and sediment phases.
4.2. Correlations between Environmental Parameters and Bacterial Community Composition
In this study, high-throughput sequencing (HTS) technology was used to evaluate the microbial community diversity and composition in the surface water spanning from spring to summer at different sites of the reservoir. Combining information on the changes of water quality and physicochemical parameters, we further explored the co-occurrence/exclusion patterns of the bacterial community with biotic/abiotic factors, also including the influence on co-occurrence patterns between cyanobacterial and non-cyanobacterial taxa during summer.
Significant spatial and temporal effects of bacterial community composition were found in QCS Reservoir (Figure 2A,B). This is mainly because the inlet site was so close to the Yangtze River, serving as the source water into the reservoir. That means the aquatic ecological environment at inlet site was very similar with conditions in the Yangtze River characterized for higher concentrations of nutrients (N, P) and turbidity, which was affected by seriously non-point pollution and soil erosion in upstream [37,38,39]. Therefore, to some extent, the microbial community composition at inlet site was very similar to the structure of microorganisms in the Yangtze estuary. By contrast, the internal and exit sites are located in the midstream and downstream of the reservoir, respectively, with characteristics of obvious lower concentrations of TN, TP and turbidity (Figure 1). Besides, the lower water flow velocity and longer retention times might facilitate the water purification and further increased the transparency of water column in these areas [15], which might partly decrease the abundance of particle-associated bacteria (such as lineages of β-Proteobacteria) in surface water at these sites. Our study also indicated that both species richness and diversity indices obviously decreased from inlet to exit site (Figure 3). Although the water quality was improved obviously at internal and exit sites, the aquatic environment at these sites provided suitable conditions for the proliferation of cyanobacterial, and resulted in rapid increasing in abundance of Synechococcus at these sites in summer season (July to September) (Figure 2A,B). Results of sub-network further indicated that water temperature was the major environmental driving factor accelerated the Synechococcus growth, and chl-a concentration also significantly increased during this period (Figure 6). In addition, several heterotrophic bacterial OTUs belonging to Actinobacteria, Gemmatimonadetes, Planctomycetes, Verrucomicrobia, β- and γ-Proteobacteria exhibited the co-occurrence patterns with Synechococcus, which indicated that mutualism mechanisms might exist between these bacterial taxa in aspect of carbon source utilization [40,41,42,43,44]. Further research is underway to confirm whether these related bacterial taxa can be bio-indicators to predict for the proliferation of Synechococcus.
The modularity analysis on correlation network indicated that the individual bacterial taxon did not exist independently in aquatic ecosystem, but closely correlated with other bacterial taxa and responded to the changes of surrounding environment together, which was further represented in co-occurrence/exclusion patterns (Figure 5A,B). The collective behavior of bacterial community is called “quorum-sensing (QS)” [45,46,47], which can strengthen the cooperations between different bacterial taxa, and also the adaptability of bacterial community to environment changes [48]. In our research, distinct co-occurrence/exclusion patterns between bacterial community were represented by different modules within networks. The different environmental variables closely correlated with co-occurrence/exclusion patterns were also included in each module. Within each module, the bacterial taxa widely connected with others, but only few bacterial taxa with high betweeness centrality connected other bacteria in different modules (Figure 5A,B). This implied that the bacterial taxa with high betweeness centrality as bridges between different modules played critical roles in the whole network [31]. In module I of the co-occurrence network, the dominant bacterial OTUs mainly classified as Bacteroidetes and Verrucomicrobia were co-occurred with environmental variables including NH4-N+, DO, Mg2+, Na+, K+ and Cl−, which indicated that these bacterial taxa mainly existed in samples at internal and exit sites in April (Figure 4A). While in module II, major environmental variables including TN, TP, turbidity and Al3+, were co-occurred with high abundant β-Proteobacterial OTUs, which illustrated that the β-Proteobacterial taxa were mainly existed in samples from inlet site. In addition, temperature, chl-a and dominant cynaobacterial OTUs represented the composing characteristics of module III, which also implied bacterial OTUs in this module were mainly distributed at internal and exit sites in summer. Moreover, the other modules also indicated the characteristics of bacterial community under different environmental driving factors. Between different modules, ACK-M1 (Actinobacteria) in module I, Sediminibacterium (Bacteroidetes) in module II and Chitinophagaceae (Bacteroidetes) in module III as bridges linked with other modules, also exhibited high betweeness centrality. This implied the important roles of these key bacterial taxa in aquatic ecosystems, although their ecological functions were still unknown. Additionally, no connections were found between module I, II and III, but all these three modules were connected with module V, which indicated the bacterial community in module V have stronger functional heterogeneities compared with other modules. On the other hand, the co-exclusion network showed the co-exclusion patterns between environmental variables and bacterial OTUs (Figure 5B). The results indicated that the co-exclusion patterns of environmental variables and bacterial OTUs have distinct spatial (inlet and the other two sites) and temporal (spring and summer) effects. In addition, the environmental factors including temperature, NH4-N+ and pH, also bacterial OTUs including ACK-M1 (Actinobacteria), KD8-87 (Gemmatimonadetes) and Burkholderiales (β-Proteobacteria) as bridges linked with other modules, which implied the potential important roles of these biotic/abiotic factors in the network.
4.3. Correlations between Antibiotic Concentrations and Bacterial Community Composition
Most of the former correlative researches have been mainly focused on the conditions of relative higher antibiotic concentrations, and how these compounds affected the bacterial community in laboratory experiments, which strongly limited our understanding about the potential ecological impact of these antibiotics on actual aquatic ecosystems [49,50]. Hence in our study, high-throughput sequencing (HTS) combined with correlation network analysis to explore the co-occurrence/exclusion patterns and effects between antibiotics and the bacterial community in the actual estuary ecosystem, as well as the influences of these antibiotics on co-occurrence pattern of cyanobacterial and non-cyanobacterial taxa.
Total of 11 antibiotics from all 16 antibiotics, were found in the co-occurrence/exclusion pattern networks (Figure 5A,B). Among these antibiotics, SQX (module VI) exhibited a positive correlation with bacterial OTU belonging to Cytophagaceae (Bacteroidetes) in a different module (module III) (Figure 5A). Although the strains of Cytophagaceae revealed sensitivity to kinds of antibiotics [51,52], we still unknown the association between Cytophagaceae and SQX. In addition, three dominant antibiotics including TYL, PEN G and ETM were found to have significant effects (p < 0.05) on variations of bacterial community composition in QCS Reservoir (Table S3). Also, these three antibiotics distributed within different modules (including module I and II of co-occurrence patterns and module III of co-exclusion pattern) of each network (Figure 5A,B). The modular classifications of these antibiotics within the co-occurrence pattern network further reflected the different distribution characteristics between these compounds in QCS Reservoir (Figure 4B). Among these, TYL was closely associated with samples in April at internal and exit sites, while ETM and PEN G were more relevant with samples at inlet site.
Each antibiotic (TYL, ETM and PEN G) exhibited extensive co-occurrence patterns with environmental variables and bacterial OTUs within modules, and we further found that these co-occurred bacterial OTUs all belonged to gram-negative bacteria (including Acidobacteria, Bacteroidetes, Nitrospirae, Verrucomicrobia, α- and β-Proteobacteria) in QCS Reservoir. Although previous research showed that some gram-negative bacteria were effective to resist and biodegrade kinds of antibiotic including macrolides and β-Lactams in a variety of ways [53], the specific interaction mechanisms between these bacterial taxa and antibiotics were still not clear. It is noted that the PEN G exhibited a co-exclusion pattern with bacterial OTUs from Planctomycetes in our research (Figure 5B). Related research has indicated that the β-lactams mainly restrained bacterial growth by impeding the synthesis of peptidoglycan in the cell walls [54]. However, the cell structure of Planctomycetes was characterized by the absence of peptidoglycan in their cell walls [55]. These results implied that PEN G may have other potential inhibition mechanisms on bacteria such as Planctomycetes. Different to widely co-occurrence patterns of these antibiotics (TYL, ETM and PEN G) within modules, no connections of these compounds with other biotic/abiotic factors were found across different modules in the network. This indicated that the variations of these antibiotics might directly affect the bacterial taxa within co-occurrence patterns first, and then further influence the whole bacterial community structure through the changes of these co-occurred bacterial taxa in aquatic ecosystem.
In a word, no significant inhibiting effects of most detected antibiotics were found on bacterial community composition in this estuary reservoir, also no influences of antibiotics were found on the co-occurrence pattern between cyanobacteria and non-cyanobacterial taxa, which were mainly because of the lower concentrations of these antibiotics. However, there might exist a potential risk of genetic transfer of resistance to related bacteria induced by exposure to long-term exposure to low levels of antibiotics in the environment. Especially, TYL, ETM and PEN G exhibited co-occurrence patterns with multiple gram-negative bacterial taxa in the network, which indicated that these bacterial taxa played important roles during the migrating and transforming course of these antibiotics in aquatic ecosystem.
5. Conclusions
In this study, we evaluated the relationships between environmental factors, antibiotics and bacterial community composition in surface water of a large reservoir located in Yangtze estuary from spring to summer (April to September). Both significant spatial and temporal effects were found in bacterial community composition between inlet and the other two sites of the reservoir. The environmental factors showed significant influence on bacterial community composition, while having little effect on distributions of most antibiotics. No significant inhibitory effects of most antibiotics on bacterial community were found in our research. Among these antibiotics, PEN G, TYL and ETM closely correlated with variations of bacterial community composition, and exhibited co-occurrence patterns with some gram-negative bacterial taxa, which implied important functions of these bacterial taxa in the course of migration and transformation of antibiotics. Further study is required to explore the interaction mechanisms between these antibiotics and bacterial taxa. In addition, the antibiotics in low concentrations revealed no significant effect on the co-occurrence pattern between cyanobacteria and non-cyanobacterial taxa. Based on above results, continued research is necessary to evaluate the potential risk of genetic transfer of resistance to related bacteria induced by long-term exposure to low levels of antibiotics in the environment.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4441/10/2/154/s1, Figure S1: Aerial schematic of Qingcaosha (QCS) Reservoir and annotated sampling locations (Site 1 (Inlet), Site 2 (Internal), Site 3 (Exit)); Figure S2: Concentrations of antibiotics at the three sites. (A) SDZ, (B) SMM, (C) SQX, (D) NFX, (E) CFX, (F) OFX, (G) LEX, (H) PEN G, (I) PEN V, (J) TYL, (K) ETM, (L) TC, (M) DC, (N) POL, (O) VAN, (P) LIN; Figure S3: Rarefaction curves of sequences in each sample; Table S1: Supplemental water chemistry and environmental parameters; Table S2: DistLM results of abundant bacterial community data against environmental variables (999 permutations); Table S3: DistLM results of abundant bacterial community data against antibiotics (999 permutations); Table S4: DistLM results of environmental variables against antibiotics (999 permutations); Table S5: Permutational MANOVA results of environmental variables, antibiotic concentrations and bacterial community compositions (9999 permutations); Table S6: Multiple linear correlations by rcor.test in ltm package (R language).
Acknowledgments
This research grant is supported by the joint program between Shanghai Jiaotong University (SJTU) and National University of Singapore (NUS), and we are also grateful to the Campus for Research Excellence And Technological Enterprise (CREATE) programme under the joint program Energy and Environmental Sustainability Solutions for Megacities (E2S2) between Shanghai Jiaotong University (SJTU) and the National University of Singapore (NUS), also Singapore National Research Foundation (NRF) under its Environmental & Water Technologies Strategic Research Programme and administered by PUB, the Singapore’s National Water Agency (Grant number: 1102-IRIS-14-02). All sources of funding of the study should be disclosed. Please clearly indicate grants that you have received in support of your research work. Clearly state if you received funds for covering the costs to publish in open access.
Author Contributions
Zheng Xu conceived and designed the experiments, and mainly performed the HTS experiment, also analyzed the sequencing data and wrote the paper. Yue Jiang mainly performed the antibiotics detection experiment; Shu Harn Te helped polish the language. Yiliang He and Karina Yew-Hoong Gin reviewed and edited the manuscript.
Conflicts of Interest
We declare that we have no financial and personal relationships with other people or organizations that can inappropriately influence our work, there is no professional or other personal interest of any nature or kind in any product, service and/or company that could be construed as influencing the position presented in, or the review of, the manuscript entitled.
References
- Yong, H.J.; Yang, J.S.; Park, K. Changes in Water Quality After the Construction of an Estuary Dam in the Geum River Estuary Dam System, Korea. J. Coast. Res. 2014, 30, 1278–1286. [Google Scholar]
- Chen, W.; Chen, K.; Kuang, C.; Zhu, D.Z.; He, L.; Mao, X.; Liang, H.; Song, H. Influence of sea level rise on saline water intrusion in the Yangtze River Estuary, China. Appl. Ocean Res. 2016, 54, 12–25. [Google Scholar] [CrossRef]
- Cloern, J.E.; Abreu, P.C.; Carstensen, J.; Chauvaud, L.; Elmgren, R.; Grall, J.; Greening, H.; Johansson, J.O.; Kahru, M.; Sherwood, E.T. Human Activities and Climate Variability Drive Fast-Paced Change across the World’s Estuarine-Coastal Ecosystems. Glob. Chang. Biol. 2015, 22, 513–529. [Google Scholar] [CrossRef] [PubMed]
- Bound, J.P.; Voulvoulis, N. Pharmaceuticals in the aquatic environment—A comparison of risk assessment strategies. Chemosphere 2004, 56, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Kümmerer, K. Antibiotics in the aquatic environment—A review—Part II. Chemosphere 2009, 75, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for the release of antibiotics in the environment: A review. Water Res. 2013, 47, 957–995. [Google Scholar]
- Manzetti, S.; Ghisi, R. The environmental release and fate of antibiotics. Mar. Pollut. Bull. 2014, 79, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Crump, B.C.; Baross, J.A.; Simenstad, C.A. Dominance of particle-attached bacteria in the Columbia River estuary, USA. Aquat. Microb. Ecol. 1998, 14, 7–18. [Google Scholar] [CrossRef]
- Rieck, A.; Herlemann, D.P.R.; Jürgens, K.; Grossart, H.P. Particle-Associated Differ from Free-Living Bacteria in Surface Waters of the Baltic Sea. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Yung, C.M.; Ward, C.S.; Davis, K.M.; Johnson, Z.I.; Hunt, D.E. Insensitivity of Diverse and Temporally Variable Particle-Associated Microbial Communities to Bulk Seawater Environmental Parameters. Appl. Environ. Microbiol. 2016, 82, 3431–3437. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Shi, Y.; Gao, L.; Liu, J.; Cai, Y. Occurrence of antibiotics in water, sediments, aquatic plants, and animals from Baiyangdian Lake in North China. Chemosphere 2012, 89, 1307–1315. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lu, G.; Wang, Y.; Yan, Z.; Yang, X.; Ding, J.; Jiang, Z. Bioconcentration, metabolism, and biomarker responses in freshwater fish Carassius auratus exposed to roxithromycin. Chemosphere 2014, 99, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.J.; Hou, J.H.; Kuo, T.F.; Lai, H.T. Toxicity of the veterinary sulfonamide antibiotic sulfamonomethoxine to five aquatic organisms. Environ. Toxicol. Pharmacol. 2014, 38, 874–880. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; He, Y.; Kirumba, G.; Hassan, Y.; Li, J. Phosphorus fractions and phosphate sorption-release characteristics of the sediment in the Yangtze River estuary reservoir. Ecol. Eng. 2013, 55, 62–66. [Google Scholar] [CrossRef]
- Jin, X.; He, Y.; Zhang, B.; Hassan, Y.; George, K. Impact of sulfate and chloride on sediment phosphorus release in the Yangtze Estuary Reservoir, China. Water Sci. Technol. 2013, 67, 1748–1756. [Google Scholar] [CrossRef] [PubMed]
- Ou, H.S.; Wei, C.H.; Deng, Y.; Gao, N.Y. Principal component analysis to assess the composition and fate of impurities in a large river-embedded reservoir: Qingcaosha Reservoir. Environ. Sci. Process. Impacts 2013, 15, 1613–1621. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Yang, Y.; Zhou, J.; Liu, M.; Nie, M.; Shi, H.; Gu, L. Antibiotics in the surface water of the Yangtze Estuary: Occurrence, distribution and risk assessment. Environ. Pollut. 2013, 175, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Yang, Y.; Liu, M.; Yan, C.; Yue, H.; Zhou, J. Occurrence and distribution of antibiotics in the surface sediments of the Yangtze Estuary and nearby coastal areas. Mar. Pollut. Bull. 2014, 83, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zhou, J.L. Occurrence and behavior of antibiotics in water and sediments from the Huangpu River, Shanghai, China. Chemosphere 2014, 95, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Xie, B.; Yuan, Q.; Xu, W.; Lu, J. Microbial community study in newly established Qingcaosha Reservoir of Shanghai, China. Appl. Microbiol. Biotechnol. 2014, 98, 9849–9858. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Li, G.; Wang, C.; Jing, Y.; Zhu, Y.; Zhang, S.; Liu, Y. Community dynamics of prokaryotic and eukaryotic microbes in an estuary reservoir. Sci. Rep. 2014, 4, 6966. [Google Scholar] [CrossRef] [PubMed]
- Wei, F. Monitoring and Analysis Methods of Water and Wastewater; China Environmental Science Press: Beijing, China, 2002. [Google Scholar]
- Gera, A.; Alcoverro, T.; Mascaró, O.; Pérez, M.; Romero, J. Exploring the utility of Posidonia oceanica chlorophyll fluorescence as an indicator of water quality within the European Water Framework Directive. Environ. Monit. Assess. 2012, 184, 3675–3686. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berglyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Shu, H.T.; Tan, B.F.; Thompson, J.R.; Gin, Y.H. Relationship of microbiota and cyanobacterial secondary metabolites in Planktothricoides-dominated bloom. Environ. Sci. Technol. 2017, 51, 4199–4209. [Google Scholar]
- Ng, C.; Le, T.H.; Goh, S.G.; Liang, L.; Kim, Y.; Rose, J.B.; Yewhoong, K.G. A Comparison of Microbial Water Quality and Diversity for Ballast and Tropical Harbor Waters. PLoS ONE 2016, 11, e0154652. [Google Scholar] [CrossRef] [PubMed]
- Woodhouse, J.N.; Kinsela, A.S.; Collins, R.N.; Bowling, L.C.; Honeyman, G.L.; Holliday, J.K.; Neilan, B.A. Microbial communities reflect temporal changes in cyanobacterial composition in a shallow ephemeral freshwater lake. ISME J. 2016, 10, 1337–1351. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling The False Discovery Rate—A Practical And Powerful Approach To Multiple Testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. In Proceedings of the Third International Aaai Conference on Weblogs and Social Media, San Jose, CA, USA, 17–20 May 2009; Available online: https://gephi.org/publications/gephi-bastian-feb09.pdf (accessed on 23 November 2017).
- Yu, H.; Pm, K.; Sprecher, E.; Trifonov, V.; Gerstein, M. The Importance of Bottlenecks in Protein Networks: Correlation with Gene Essentiality and Expression Dynamics. PLoS Comput. Biol. 2007, 3, e59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gothwal, R.; Shashidhar, T. Antibiotic Pollution in the Environment: A Review. CLEAN Soil Air Water 2015, 43, 479–489. [Google Scholar] [CrossRef]
- Xu, W.H.; Zhang, G.; Zou, S.C.; Li, X.D.; Liu, Y.C. Determination of selected antibiotics in the Victoria Harbour and the Pearl River, South China using high-performance liquid chromatography-electrospray ionization tandem mass spectrometry. Environ. Pollut. 2007, 145, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Waksman, S.A. Microbial Antagonisms and Antibiotic Substances; The Commonwealth Fund: New York, NY, USA, 1947; pp. 1033–1034. [Google Scholar]
- Bao, L.J.; Maruya, K.A.; Snyder, S.A.; Zeng, E.Y. China’s water pollution by persistent organic pollutants. Environ. Pollut. 2012, 163, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Kümmerer, K. Antibiotics in the aquatic environment—A review—Part I. Chemosphere 2009, 75, 417–434. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liang, D.; Ren, L.; Shi, S.; Li, Z.; Zhang, T.; Huang, Y. Concentration and source identification of polycyclic aromatic hydrocarbons and phthalic acid esters in the surface water of the Yangtze River Delta, China. J. Environ. Sci. 2012, 24, 335–342. [Google Scholar] [CrossRef]
- Floehr, T.; Xiao, H.; Scholz-Starke, B.; Ottermanns, R.; Ross-Nickoll, M. Solution by dilution?—A review on the pollution status of the Yangtze River. Environ. Sci. Pollut. Res. Int. 2013, 20, 6934–6971. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.B.; Jiao, N.Z.; Feng, J.; Shu, Q.L. Research progress on Planctomycetes’ diversity and ecological function in marine environments. Microbiol. China 2014, 41, 1891–1902. [Google Scholar]
- Eiler, A.; Olsson, J.A.; Bertilsson, S. Diurnal variations in the auto- and heterotrophic activity of cyanobacterial phycospheres (Gloeotrichia echinulata) and the identity of attached bacteria. Freshw. Biol. 2010, 51, 298–311. [Google Scholar] [CrossRef]
- Li, J.; Zhang, J.; Liu, L.; Fan, Y.; Li, L.; Yang, Y.; Lu, Z.; Zhang, X. Annual periodicity in planktonic bacterial and archaeal community composition of eutrophic Lake Taihu. Sci. Rep. 2015, 5, 15488. [Google Scholar] [CrossRef] [PubMed]
- Buck, U.; Grossart, H.P.; Amann, R.; Pernthaler, J. Substrate incorporation patterns of bacterioplankton populations in stratified and mixed waters of a humic lake. Environ. Microbiol. 2010, 11, 1854–1865. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.E.; Newton, R.J.; Mcmahon, K.D. Evidence for structuring of bacterial community composition by organic carbon source in temperate lakes. Environ. Microbiol. 2010, 11, 2463–2472. [Google Scholar] [CrossRef] [PubMed]
- Agostini, V.O.; Macedo, A.J.; Muxagata, E. Evaluation of antibiotics as a methodological procedure to inhibit free-living and biofilm bacteria in marine zooplankton culture. Anais da Academia Brasileira de Ciencias 2016, 88, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Cornforth, D.M.; Popat, R.; Mcnally, L.; Gurney, J.; Scottphillips, T.C.; Ivens, A.; Diggle, S.P.; Brown, S.P. Combinatorial quorum sensing allows bacteria to resolve their social and physical environment. Proc. Natl. Acad. Sci. USA 2014, 111, 4280–4284. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, C. Exploiting Quorum Sensing Interfering Strategies in Gram-Negative Bacteria for the Enhancement of Environmental Applications. Front. Microbiol. 2016, 6, 1535. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, E.; Gothalwal, R. Effect of environmental factors on bacterial quorum sensing. Cell. Mol. Biol. 2014, 60, 46–60. [Google Scholar] [PubMed]
- Kievit, T.R.D.; Iglewski, B.H. Bacterial Quorum Sensing in Pathogenic Relationships. Infect. Immun. 2000, 68, 4839–4849. [Google Scholar] [CrossRef] [PubMed]
- Mcknight, U.S.; Rasmussen, J.J.; Kronvang, B.; Binning, P.J.; Bjerg, P.L. Sources, occurrence and predicted aquatic impact of legacy and contemporary pesticides in streams. Environ. Pollut. 2015, 200, 64–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Yang, M.; Hu, J.; Zhang, Y.; Chang, H.; Jin, F. Determination of penicillin G and its degradation products in a penicillin production wastewater treatment plant and the receiving river. Water Res. 2008, 42, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Filippini, M.; Svercel, M.; Laczko, E.; Kaech, A.; Ziegler, U.; Bagheri, H.C. Fibrella aestuarina gen. nov., sp. nov., a filamentous bacterium of the family Cytophagaceae isolated from a tidal flat, and emended description of the genus Rudanella Weon et al. 2008. Int. J. Syst. Evol. Microbiol. 2011, 61, 184–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joung, Y.; Kim, H.; Kang, H.; Lee, B.I.; Ahn, T.S.; Joh, K. Lacihabitans soyangensis gen. nov., sp. nov., a new member of the family Cytophagaceae, isolated from a freshwater reservoir. Int. J. Syst. Evol. Microbiol. 2014, 64, 3188–3194. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Bacterial resistance to antibiotics: Enzymatic degradation and modification. Adv. Drug Deliv. Rev. 2005, 57, 1451–1470. [Google Scholar] [CrossRef] [PubMed]
- Heijenoort, J.V.; Gutmann, L. Correlation between the structure of the bacterial peptidoglycan monomer unit, the specificity of transpeptidation, and susceptibility to β-lactams. Proc. Natl. Acad. Sci. USA 2000, 97, 5028–5030. [Google Scholar] [CrossRef] [PubMed]
- Fuerst, J.A.; Sagulenko, E. Beyond the bacterium: Planctomycetes challenge our concepts of microbial structure and function. Nat. Rev. Microbiol. 2011, 9, 403–413. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Water chemistry and environmental parameters. (A) Temperature; (B) pH; (C) Chlorophyll-a; (D) Turbidity; (E) Ammonia Nitrogen (NH4-N+); (F) Inorganic Carbon (IC); (G) Dissolved Oxygen (DO); (H) Total Phosphorus (TP); (I) Total Nitrogen (TN).
Figure 1.
Water chemistry and environmental parameters. (A) Temperature; (B) pH; (C) Chlorophyll-a; (D) Turbidity; (E) Ammonia Nitrogen (NH4-N+); (F) Inorganic Carbon (IC); (G) Dissolved Oxygen (DO); (H) Total Phosphorus (TP); (I) Total Nitrogen (TN).
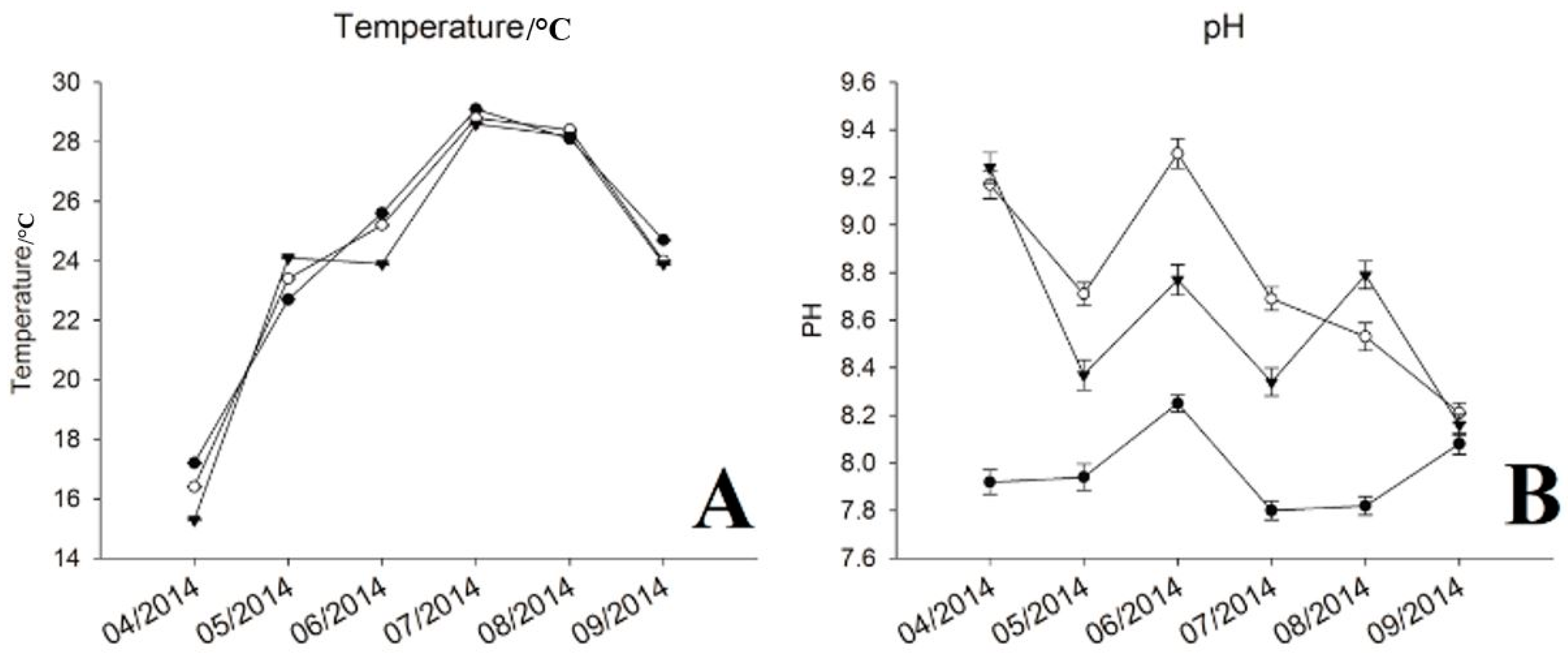
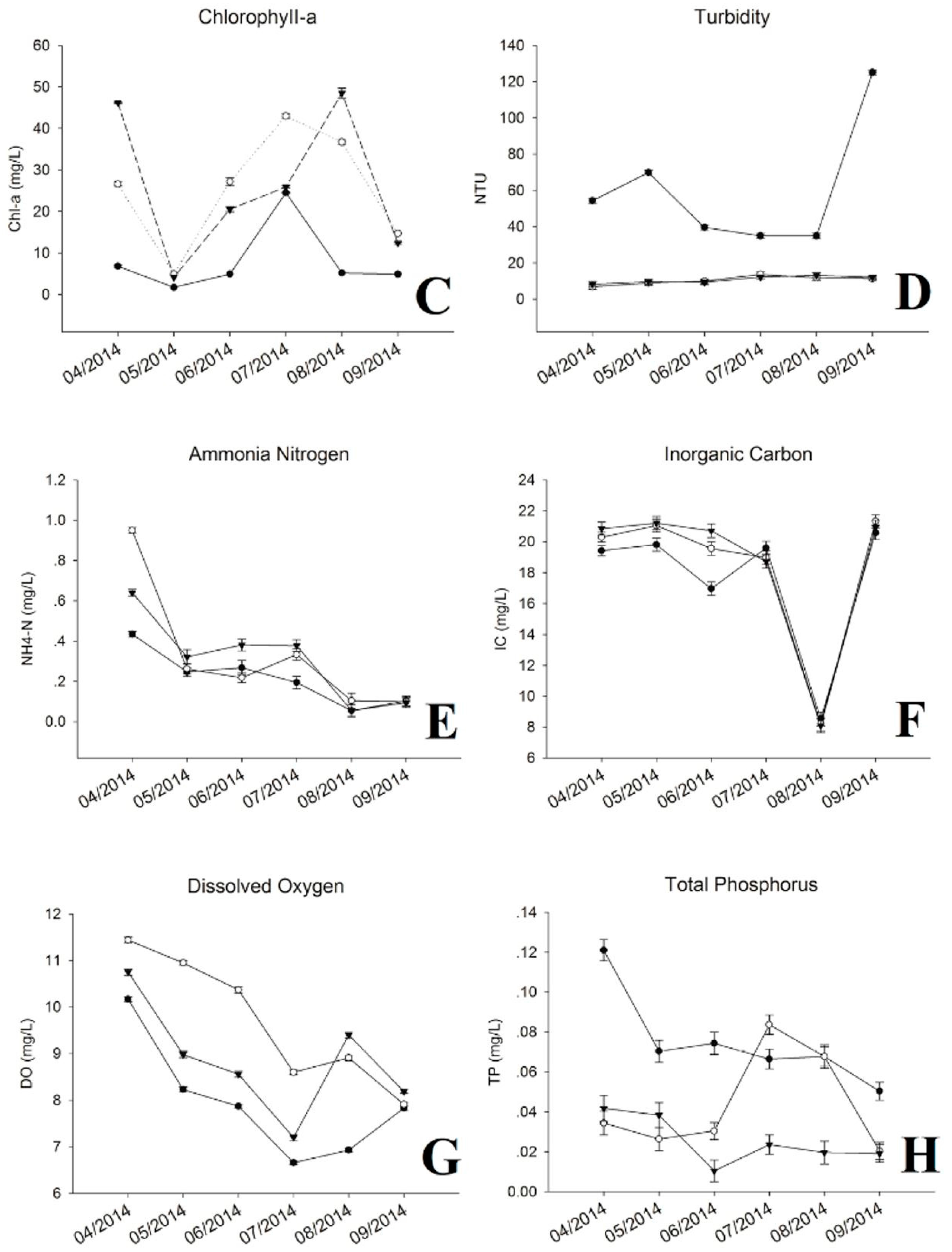

Figure 2.
Relative abundance of 16S rRNA bacterial OTUs across the whole sampling period (A) Phylum level; (B) Class level.
Figure 2.
Relative abundance of 16S rRNA bacterial OTUs across the whole sampling period (A) Phylum level; (B) Class level.

Figure 3.
Bacterial OTU Richness (Chao/Ace, Black) and diversity (Inverse Simpson Index, Red).
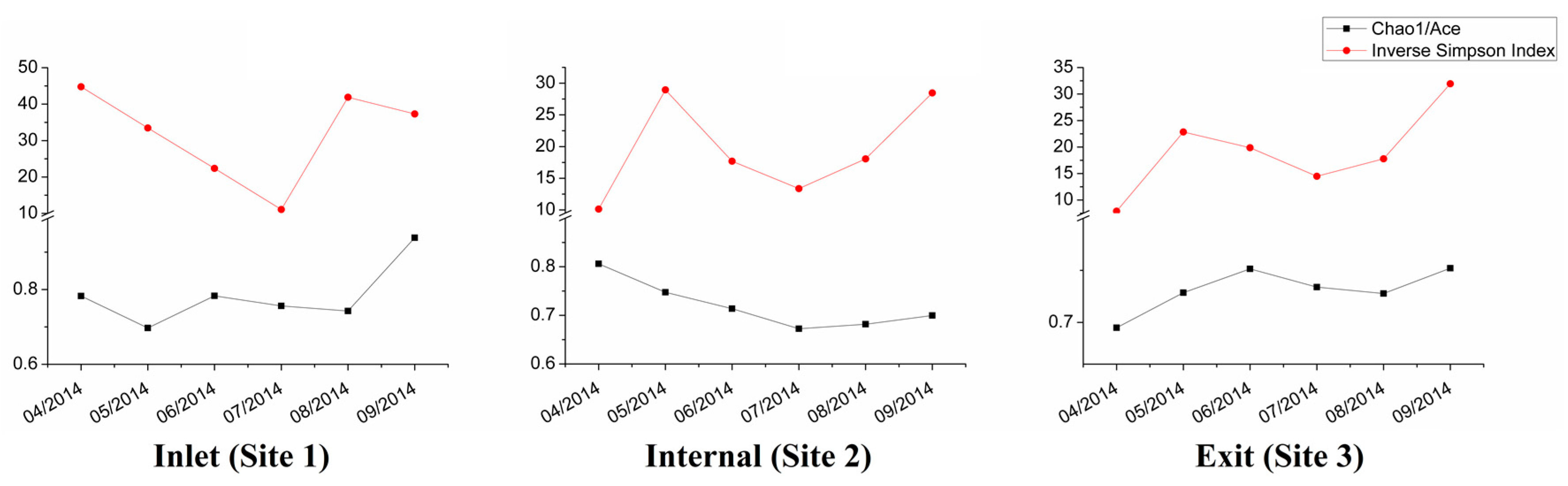
Figure 4.
The nonmetric multidimensional scaling (NMDS) reflecting the distribution of bacterial communities with environmental variables (A) and antibiotics (B) in estuary reservoir.
Figure 4.
The nonmetric multidimensional scaling (NMDS) reflecting the distribution of bacterial communities with environmental variables (A) and antibiotics (B) in estuary reservoir.
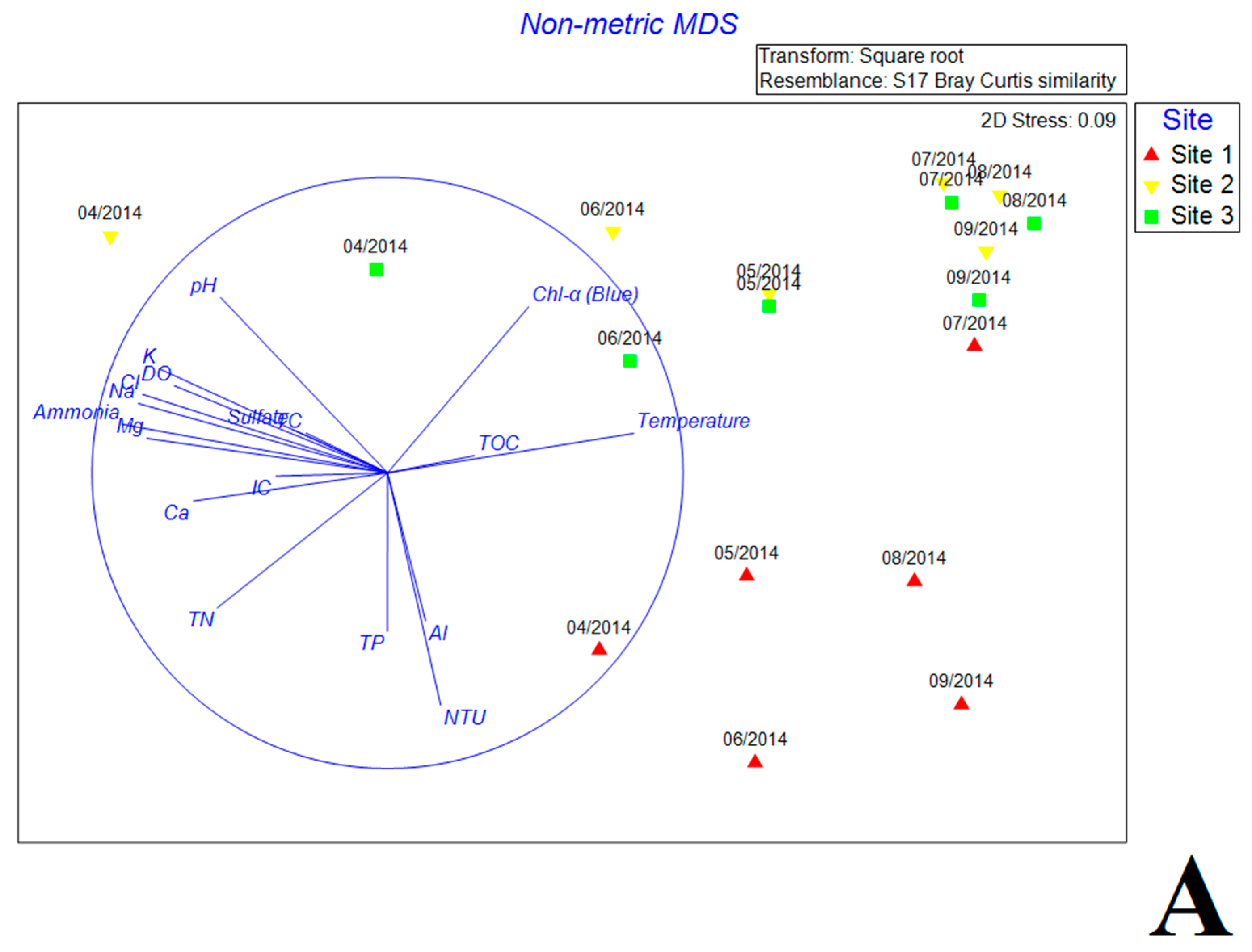
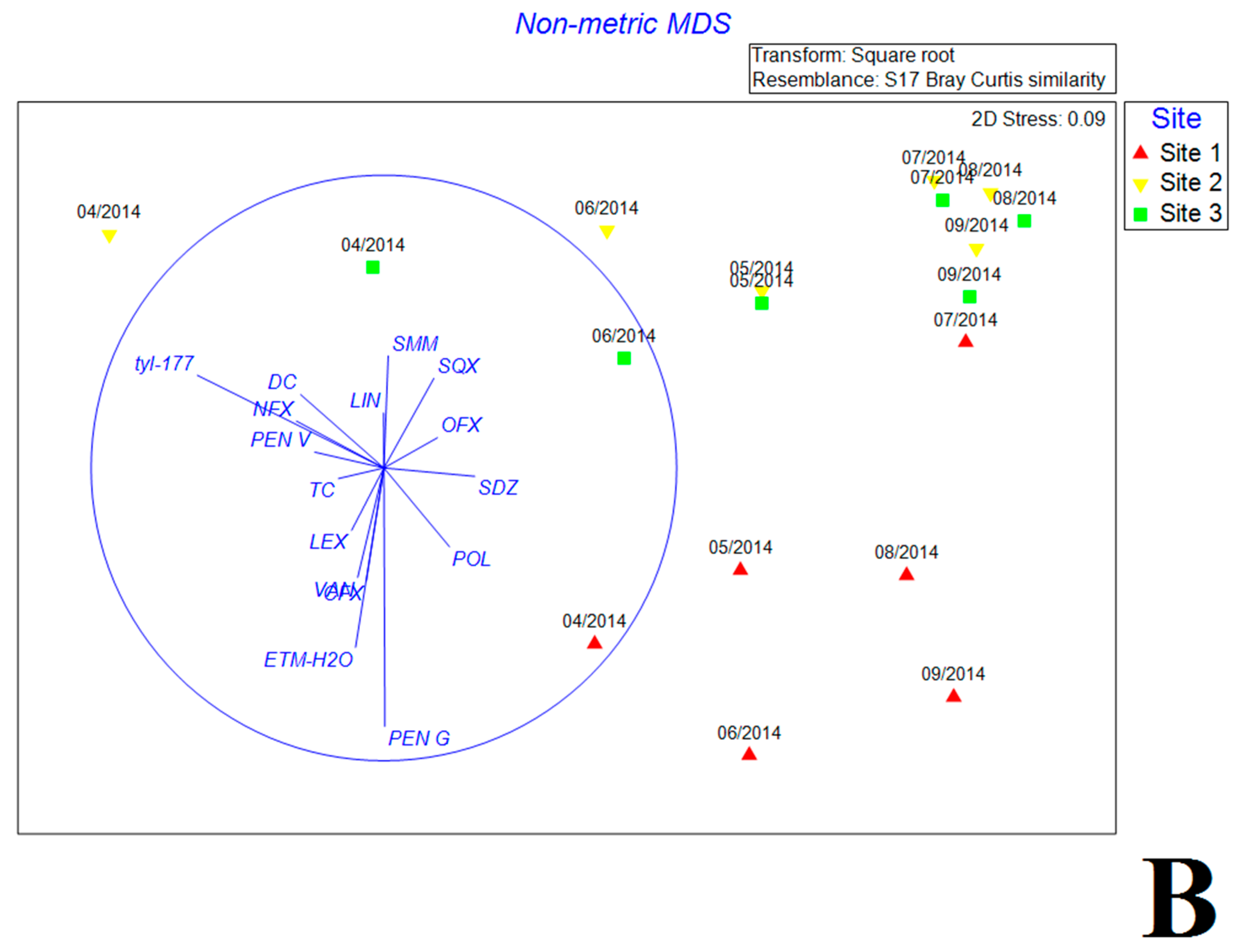
Figure 5.
The network analysis showing the co-occurrence patterns between environmental variables, antibiotics and bacterial OTUs. A connection stands for a significant (p < 0.05) and strong positive (A) or negative correlation (B). The nodes were colored according to modularity class. The size of each node represents the value of betweenness centrality.
Figure 5.
The network analysis showing the co-occurrence patterns between environmental variables, antibiotics and bacterial OTUs. A connection stands for a significant (p < 0.05) and strong positive (A) or negative correlation (B). The nodes were colored according to modularity class. The size of each node represents the value of betweenness centrality.


Figure 6.
The organic correlation sub-network reflecting the pairwise correlations between Synechococcus and other bacterial OTUs, environmental factors and antibiotics. The nodes were colored according to different bacterial phylum. A red edge represents a significant positive correlation. A blue edge represents a significant negative correlation.
Figure 6.
The organic correlation sub-network reflecting the pairwise correlations between Synechococcus and other bacterial OTUs, environmental factors and antibiotics. The nodes were colored according to different bacterial phylum. A red edge represents a significant positive correlation. A blue edge represents a significant negative correlation.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xu, Z.; Jiang, Y.; Te, S.H.; He, Y.; Gin, K.Y.-H. The Effects of Antibiotics on Microbial Community Composition in an Estuary Reservoir during Spring and Summer Seasons. Water 2018, 10, 154. https://doi.org/10.3390/w10020154
AMA Style
Xu Z, Jiang Y, Te SH, He Y, Gin KY-H. The Effects of Antibiotics on Microbial Community Composition in an Estuary Reservoir during Spring and Summer Seasons. Water. 2018; 10(2):154. https://doi.org/10.3390/w10020154
Chicago/Turabian StyleXu, Zheng, Yue Jiang, Shu Harn Te, Yiliang He, and Karina Yew-Hoong Gin. 2018. "The Effects of Antibiotics on Microbial Community Composition in an Estuary Reservoir during Spring and Summer Seasons" Water 10, no. 2: 154. https://doi.org/10.3390/w10020154
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.