An Overview of Autophagy and Yeast Pseudohyphal Growth: Integration of Signaling Pathways during Nitrogen Stress

Abstract
:1. Introduction
2. Autophagy and Filamentous Growth in Yeast
2.1. Autophagy in Saccharomyces cerevisiae and Other Filamentous Fungi
2.2. The Molecular Machinery of Autophagy
2.2.1. Autophagic Induction
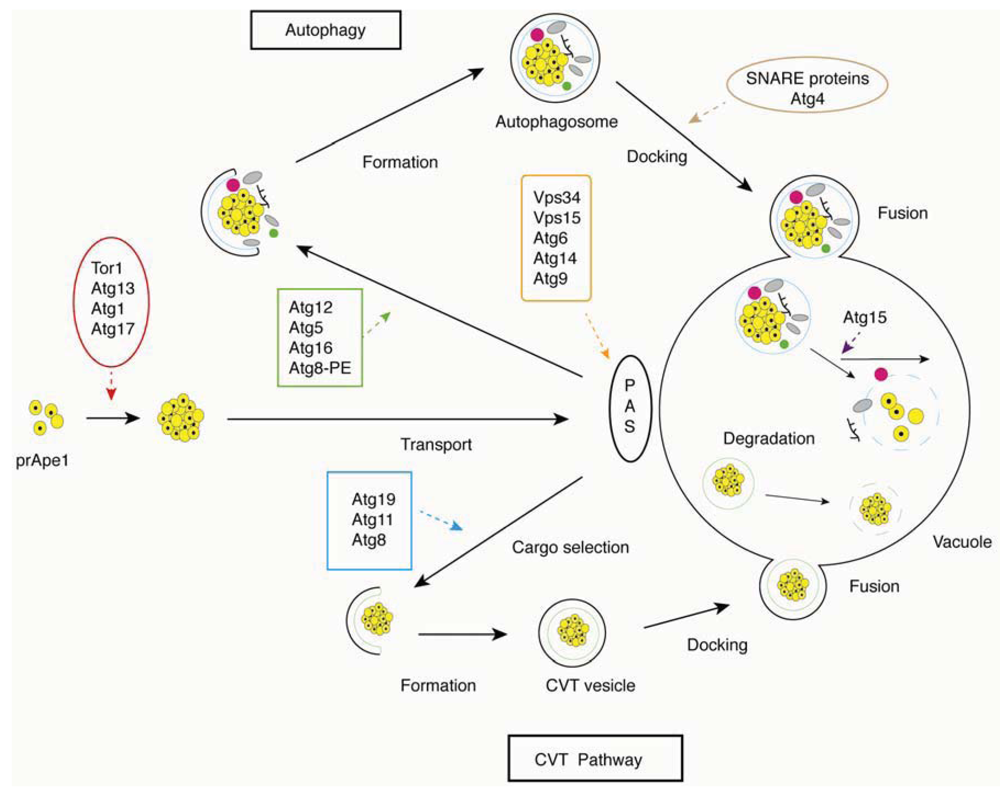
2.2.2. Cargo Selection and Packaging
2.2.3. Autophagosome Formation
2.2.4. Vesicle Docking and Fusion
2.2.5. Autophagosome Breakdown
2.3. Filamentous Growth in Saccharomyces cerevisiae and Other Yeast Species
2.3.1. The Genetic Basis of Filamentous Growth
2.3.2. Cellular Processes Contributing to Yeast Filamentation
2.3.3. Downstream Genes Mediating Increased Cell-Cell Adhesion
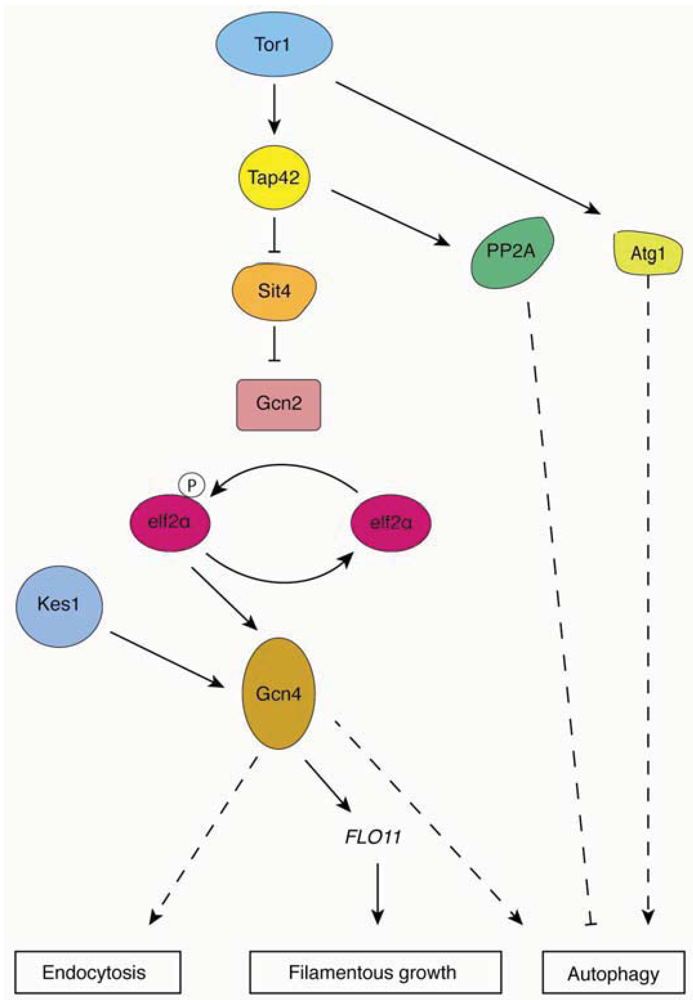
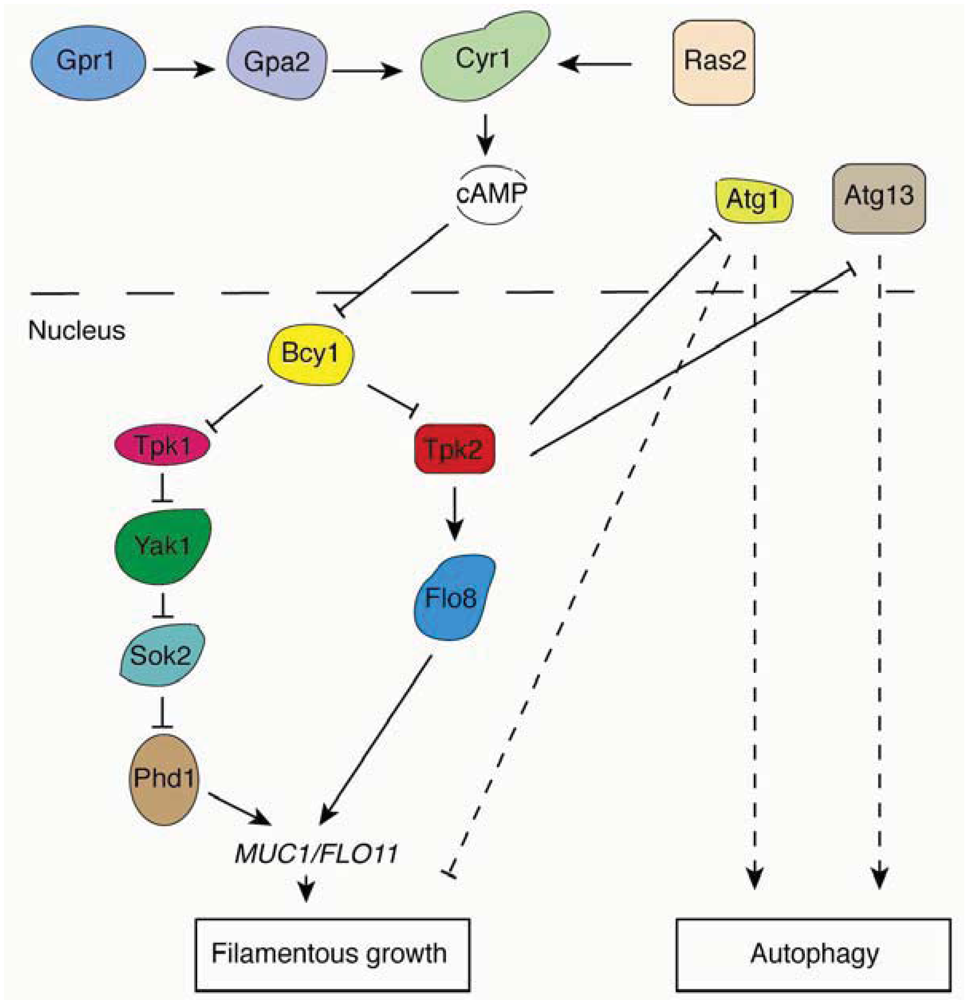
2.3.4. An Interrelationship Between Autophagy-Related Genes and Yeast Filamentation

{kind=link}
{kind=link}
{kind=link}
| Gene | Autophagy Process | Protein Function/Description | Transcript Levels in Early Fil. Growth | Deletion (Δ)/Overexpression (OE) Phenotype |
|---|---|---|---|---|
| ATG1 | Induction; retrieval | Protein kinase | Increased | Decreased fil. growth (OE); Exaggerated fil. growth (Δ) |
| ATG3 | Vesicle expansion and completion | Conjugation enzyme | Increased | Decreased fil. growth (OE) |
| ATG4 | Vesicle expansion and completion | Cysteine protease | Increased | Wild-type fil. growth (OE) |
| ATG5 | Vesicle expansion and completion | Conjugation enzyme | Increased | Untested |
| ATG6 | Vesicle nucleation | PI3P binding | Increased | Wild-type fil. growth (OE) |
| ATG7 | Vesicle expansion and completion | Activating enzyme | Increased | Decreased fil. growth (OE); Exaggerated fil. growth (Δ) |
| ATG8 | Vesicle expansion/completion | Ubiquitin-like protein | Increased | Untested |
| ATG9 | Vesicle nucleation; retrieval | Integral membrane protein | Increased | Untested |
| ATG14 | Vesicle nucleation | PI3-Kinase complex | Increased | Untested |
| ATG17 | Induction | Atg1p modulator | Increased | Decreased fil. growth (OE) |
| ATG19 | Induction | PI3P binding | Increased | Decreased fil. growth (OE) |
| ATG20 | Induction | PI3P binding | Increased | Untested |
| ATG21 | Cvt pathway | PI3P binding | Increased | Untested |
| ATG22 | Efflux from the vacuole | Vacuolar permease | Increased | Untested |
| ATG23 | Cvt pathway | PI3P binding | Unchanged | Decreased fil. growth (OE) |
| ATG24 | Autophagic body breakdown | Vacuolar membrane protein | Unchanged | Decreased fil. growth (OE) |
| ATG29 | Peroxisome sequestration | UDP-glucose | Unchanged | Decreased fil. growth (OE) |
2.4. Autophagy and Filamentous Growth Signaling Pathways
2.4.1. Tor Complex 1

2.4.2. The Ras/PKA Pathway

2.4.3. The Nutrient-Sensing Snf1p Pathway
2.5. TORC1 Regulates Endocytosis in S. cerevisiae
3. Conclusions
Acknowledgments
Conflict of Interest
References
- Magasanik, B.; Kaiser, C.A. Nitrogen regulation in Saccharomyces cerevisiae. Gene 2002, 290, 1–18. [Google Scholar] [CrossRef]
- Santangelo, G.M. Glucose Signaling in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2006, 70, 253–282. [Google Scholar] [CrossRef]
- Zaman, S.; Lippman, S.I.; Zhao, X.; Broach, J.R. How Saccharomyces Responds to Nutrients. Annu. Rev. Genet. 2008, 42, 27–81. [Google Scholar] [CrossRef]
- Hinnebusch, A.G. Translational regulation of GCN4 and the general amino acid control of yeast. Annu. Rev. Microbiol. 2005, 59, 407–450. [Google Scholar] [CrossRef]
- Klionsky, D.J. The molecular machinery of autophagy: Unanswered questions. J. Cell Sci. 2005, 118, 7–18. [Google Scholar] [CrossRef]
- Majeski, A.E.; Fred Dice, J. Mechanisms of chaperone-mediated autophagy. Int. J. Biochem. Cell Biol. 2004, 36, 2435–2444. [Google Scholar] [CrossRef]
- Wang, K.; Klionsky, D.J. Mitochondria removal by autophagy. Autophagy 2011, 7, 297–300. [Google Scholar] [CrossRef]
- Xie, Z.; Klionsky, D.J. Autophagosome formation: Core machinery and adaptations. Nat. Cell Biol. 2007, 9, 1102–1109. [Google Scholar] [CrossRef]
- Kron, S.J.; Styles, C.A.; Fink, G.R. Symmetric cell division in pseudohyphae of the yeast Saccharomyces cerevisiae. Mol. Biol. Cell 1994, 5, 1003–1022. [Google Scholar]
- Jin, R.; Dobry, C.J.; McCown, P.J.; Kumar, A. Large-Scale Analysis of Yeast Filamentous Growth by Systematic Gene Disruption and Overexpression. Mol. Biol. Cell 2008, 19, 284–296. [Google Scholar] [CrossRef]
- Piccirillo, S.; Honigberg, S.M. Sporulation patterning and invasive growth in wild and domesticated yeast colonies. Res. Microbiol. 2010, 161, 390–398. [Google Scholar] [CrossRef]
- Noda, T.; Ohsumi, Y. Tor, a Phosphatidylinositol Kinase Homologue, Controls Autophagy in Yeast. J. Biol. Chem. 1998, 273, 3963–3966. [Google Scholar] [CrossRef]
- Braus, G.H.; Grundmann, O.; Brückner, S.; Mösch, H.-U. Amino Acid Starvation and Gcn4p Regulate Adhesive Growth and FLO11 Gene Expression in Saccharomyces cerevisiae. Mol. Biol. Cell 2003, 14, 4272–4284. [Google Scholar] [CrossRef]
- Pan, X.; Heitman, J. Cyclic AMP-Dependent Protein Kinase Regulates Pseudohyphal Differentiation in Saccharomyces cerevisiae. Mol. Cell. Biol. 1999, 19, 4874–4887. [Google Scholar]
- Furuta, S.; Hidaka, E.; Ogata, A.; Yokota, S.; Kamata, T. Ras is involved in the negative control of autophagy through the class I PI3-kinase. Oncogene 2004, 23, 3898–3904. [Google Scholar] [CrossRef]
- Ma, J.; Jin, R.; Dobry, C.J.; Lawson, S.K.; Kumar, A. Overexpression of Autophagy-Related Genes Inhibits Yeast Filamentous Growth. Autophagy 2007, 3, 604–609. [Google Scholar]
- Ma, J.; Jin, R.; Jia, X.; Dobry, C.J.; Wang, L.; Reggiori, F.; Zhu, J.; Kumar, A. An Interrelationship Between Autophagy and Filamentous Growth in Budding Yeast. Genetics 2007, 177, 205–214. [Google Scholar] [CrossRef]
- Klionsky, D.J. Autophagy: From phenomenology to molecular understanding in less than a decade. Nat. Rev. Mol. Cell Biol. 2007, 8, 931–937. [Google Scholar] [CrossRef]
- Mah, L.Y.; Ryan, K.M. Autophagy and Cancer. Cold Spring Harb. Perspect. Biol. 2012, 4, a008821. [Google Scholar] [CrossRef]
- Barnett, A.; Brewer, G.J. Autophagy in Aging and Alzheimer's Disease: Pathologic or Protective? J. Alzheimers Dis. 2011, 25, 385–394. [Google Scholar]
- Pollack, J.K.; Harris, S.D.; Marten, M.R. Autophagy in filamentous fungi. Fungal Genet. Biol. 2009, 46, 1–8. [Google Scholar] [CrossRef]
- Skamnioti, P.; Gurr, S.J. Against the grain: Safeguarding rice from rice blast disease. Trends Biotechnol. 2009, 27, 141–150. [Google Scholar] [CrossRef]
- Veneault-Fourrey, C.; Talbot, N.J. Autophagic Cell Death and its Importance for Fungal Developmental Biology and Pathogenesis. Autophagy 2007, 3, 126–127. [Google Scholar]
- Hu, G.; Hacham, M.; Waterman, S.R.; Panepinto, J.; Shin, S.; Liu, X.; Gibbons, J.; Valyi-Nagy, T.; Obara, K.; Jaffe, H.A.; Ohsumi, Y.; Williamson, P.R. PI3K signaling of autophagy is required for starvation tolerance and virulenceof Cryptococcus neoformans. J. Clin. Invest. 2008, 118, 1186–1197. [Google Scholar] [CrossRef]
- Richie, D.L.; Fuller, K.K.; Fortwendel, J.; Miley, M.D.; McCarthy, J.W.; Feldmesser, M.; Rhodes, J.C.; Askew, D.S. Unexpected Link between Metal Ion Deficiency and Autophagy in Aspergillus fumigatus. Eukaryotic Cell 2007, 6, 2437–2447. [Google Scholar] [CrossRef]
- Palmer, G.E.; Kelly, M.N.; Sturtevant, J.E. Autophagy in the pathogen Candida albicans. Microbiology 2007, 153, 51–58. [Google Scholar] [CrossRef]
- Chrétien, F.; Lortholary, O.; Kansau, I.; Neuville, S.; Gray, F.; Dromer, F. Pathogenesis of Cerebral Cryptococcus neoformans Infection after Fungemia. J. Infect. Dis. 2002, 186, 522–530. [Google Scholar] [CrossRef]
- Schmelzle, T.; Hall, M.N. TOR, a Central Controller of Cell Growth. Cell 2000, 103, 253–262. [Google Scholar] [CrossRef]
- Kamada, Y.; Funakoshi, T.; Shintani, T.; Nagano, K.; Ohsumi, M.; Ohsumi, Y. Tor-Mediated Induction of Autophagy via an Apg1 Protein Kinase Complex. J. Cell Biol. 2000, 150, 1507–1513. [Google Scholar] [CrossRef]
- Kabeya, Y.; Kamada, Y.; Baba, M.; Takikawa, H.; Sasaki, M.; Ohsumi, Y. Atg17 Functions in Cooperation with Atg1 and Atg13 in Yeast Autophagy. Mol. Biol. Cell 2005, 16, 2544–2553. [Google Scholar] [CrossRef]
- Cheong, H.; Klionsky, D.J. Dual role of Atg1 in regulation of autophagy-specific PAS assembly in Saccharomyces cerevisiae. Autophagy 2008, 4, 724–726. [Google Scholar]
- Cheong, H.; Nair, U.; Geng, J.; Klionsky, D.J. The Atg1 Kinase Complex Is Involved in the Regulation of Protein Recruitment to Initiate Sequestering Vesicle Formation for Nonspecific Autophagy in Saccharomyces cerevisiae. Mol. Biol. Cell 2008, 19, 668–681. [Google Scholar]
- Kawamata, T.; Kamada, Y.; Kabeya, Y.; Sekito, T.; Ohsumi, Y. Organization of the Pre-autophagosomal Structure Responsible for Autophagosome Formation. Mol. Biol. Cell 2008, 19, 2039–2050. [Google Scholar] [CrossRef]
- Suzuki, K.; Kubota, Y.; Sekito, T.; Ohsumi, Y. Hierarchy of Atg proteins in pre-autophagosomal structure organization. Genes Cells 2007, 12, 209–218. [Google Scholar] [CrossRef]
- Talloczy, Z.; Jiang, W.; Virgin, H.W.; Leib, D.A.; Scheuner, D.; Kaufman, R.J.; Eskelinen, E.-L.; Levine, B. Regulation of starvation- and virus-induced autophagy by the eIF2α kinase signaling pathway. Proc. Natl. Acad. Sci. 2002, 99, 190–195. [Google Scholar]
- Levine, B.; Klionsky, D.J. Development by Self-Digestion: Molecular Mechanisms and Biological Functions of Autophagy. Dev. Cell 2004, 6, 463–477. [Google Scholar] [CrossRef]
- Lynch-Day, M.A.; Klionsky, D.J. The Cvt pathway as a model for selective autophagy. FEBS Lett. 2010, 584, 1359–1366. [Google Scholar] [CrossRef]
- Scott, S.V.; Guan, J.; Hutchins, M.U.; Kim, J.; Klionsky, D.J. Cvt19 Is a Receptor for the Cytoplasm-to-Vacuole Targeting Pathway. Mol. Cell 2001, 7, 1131–1141. [Google Scholar] [CrossRef]
- Shintani, T.; Huang, W.-P.; Stromhaug, P.E.; Klionsky, D.J. Mechanism of Cargo Selection in the Cytoplasm to Vacuole Targeting Pathway. Dev. Cell 2002, 3, 825–837. [Google Scholar] [CrossRef]
- Itakura, E.; Kishi, C.; Inoue, K.; Mizushima, N. Beclin 1 Forms Two Distinct Phosphatidylinositol 3-Kinase Complexes with Mammalian Atg14 and UVRAG. Mol. Biol. Cell 2008, 19, 5360–5372. [Google Scholar] [CrossRef]
- Kihara, A.; Noda, T.; Ishihara, N.; Ohsumi, Y. Two Distinct Vps34 Phosphatidylinositol 3-Kinase Complexes Function in Autophagy and Carboxypeptidase Y Sorting inSaccharomyces cerevisiae. J. Cell Biol. 2001, 152, 519–530. [Google Scholar] [CrossRef]
- Itakura, E.; Mizushima, N. Atg14 and UVRAG: Mutually exclusive subunits of mammalian Beclin 1-PI3K complexes. Autophagy 2009, 5, 534–536. [Google Scholar] [CrossRef]
- Nice, D.C.; Sato, T.K.; Stromhaug, P.E.; Emr, S.D.; Klionsky, D.J. Cooperative Binding of the Cytoplasm to Vacuole Targeting Pathway Proteins, Cvt13 and Cvt20, to Phosphatidylinositol 3-Phosphate at the Pre-autophagosomal Structure Is Required for Selective Autophagy. J. Biol. Chem. 2002, 277, 30198–30207. [Google Scholar]
- Guan, J.; Stromhaug, P.E.; George, M.D.; Habibzadegah-Tari, P.; Bevan, A.; Dunn, W.A.; Klionsky, D.J. Cvt18/Gsa12 Is Required for Cytoplasm-to-Vacuole Transport, Pexophagy, and Autophagy in Saccharomyces cerevisiae andPichia pastoris. Mol. Biol. Cell 2001, 12, 3821–3838. [Google Scholar]
- Strømhaug, P.E.; Reggiori, F.; Guan, J.; Wang, C.-W.; Klionsky, D.J. Atg21 Is a Phosphoinositide Binding Protein Required for Efficient Lipidation and Localization of Atg8 during Uptake of Aminopeptidase I by Selective Autophagy. Mol. Biol. Cell 2004, 15, 3553–3566. [Google Scholar] [CrossRef]
- Suzuki, K.; Kirisako, T.; Kamada, Y.; Mizushima, N.; Noda, T.; Ohsumi, Y. The pre-autophagosomal structure organized by concerted functions of APG genes is essential for autophagosome formation. EMBO J. 2001, 20, 5971–5981. [Google Scholar] [CrossRef]
- Kirisako, T.; Baba, M.; Ishihara, N.; Miyazawa, K.; Ohsumi, M.; Yoshimori, T.; Noda, T.; Ohsumi, Y. Formation Process of Autophagosome Is Traced with Apg8/Aut7p in Yeast. J. Cell Biol. 1999, 147, 435–446. [Google Scholar] [CrossRef]
- Mizushima, N.; Noda, T.; Ohsumi, Y. Apg16p is required for the function of the Apg12p-Apg5p conjugate in the yeast autophagy pathway. EMBO J. 1999, 18, 3888–3896. [Google Scholar] [CrossRef]
- He, C.; Song, H.; Yorimitsu, T.; Monastyrska, I.; Yen, W.-L.; Legakis, J.E.; Klionsky, D.J. Recruitment of Atg9 to the preautophagosomal structure by Atg11 is essential for selective autophagy in budding yeast. J. Cell Biol. 2006, 175, 925–935. [Google Scholar] [CrossRef]
- Yen, W.-L.; Legakis, J.E.; Nair, U.; Klionsky, D.J. Atg27 Is Required for Autophagy-dependent Cycling of Atg9. Mol. Biol. Cell 2007, 18, 581–593. [Google Scholar]
- Reggiori, F.; Shintani, T.; Chong, H.; Nair, U.; Klionsky, D.J. Atg9 Cycles Between Mitochondria and the Pre-Autophagosomal Structure in Yeasts. Autophagy 2005, 1, 101–109. [Google Scholar] [CrossRef]
- Kirisako, T.; Ichimura, Y.; Okada, H.; Kabeya, Y.; Mizushima, N.; Yoshimori, T.; Ohsumi, M.; Takao, T.; Noda, T.; Ohsumi, Y. The Reversible Modification Regulates the Membrane-Binding State of Apg8/Aut7 Essential for Autophagy and the Cytoplasm to Vacuole Targeting Pathway. J. Cell Biol. 2000, 151, 263–276. [Google Scholar] [CrossRef]
- Epple, U.D.; Suriapranata, I.; Eskelinen, E.-L.; Thumm, M. Aut5/Cvt17p, a Putative Lipase Essential for Disintegration of Autophagic Bodies inside the Vacuole. J. Bacteriol. 2001, 183, 5942–5955. [Google Scholar] [CrossRef]
- Teter, S.A.; Eggerton, K.P.; Scott, S.V.; Kim, J.; Fischer, A.M.; Klionsky, D.J. Degradation of Lipid Vesicles in the Yeast Vacuole Requires Function of Cvt17, a Putative Lipase. J. Biol. Chem. 2001, 276, 2083–2087. [Google Scholar]
- Yang, Z.; Huang, J.; Geng, J.; Nair, U.; Klionsky, D.J. Atg22 Recycles Amino Acids to Link the Degradative and Recycling Functions of Autophagy. Mol. Biol. Cell 2006, 17, 5094–5104. [Google Scholar] [CrossRef]
- Cullen, P.J.; Sprague, G.F. The Regulation of Filamentous Growth in Yeast. Genetics 2012, 190, 23–49. [Google Scholar] [CrossRef]
- Lorenz, M.C.; Cutler, N.S.; Heitman, J. Characterization of Alcohol-induced Filamentous Growth in Saccharomyces cerevisiae. Mol. Biol. Cell 2000, 11, 183–199. [Google Scholar]
- Rua, D.; Tobe, B.T.; Kron, S.J. Cell cycle control of yeast filamentous growth. Curr. Opin. Microbiol. 2001, 4, 720–727. [Google Scholar]
- Cullen, P.J.; Sprague, G.F. The Roles of Bud-Site-Selection Proteins during Haploid Invasive Growth in Yeast. Mol. Biol. Cell 2002, 13, 2990–3004. [Google Scholar] [CrossRef]
- Berman, J. Morphogenesis and cell cycle progression in Candida albicans. Curr. Opin. Microbiol. 2006, 9, 595–601. [Google Scholar]
- Kelly, M.T.; MacCallum, D.M.; Clancy, S.D.; Odds, F.C.; Brown, A.J.P.; Butler, G. The Candida albicans CaACE2 gene affects morphogenesis, adherence and virulence. Mol. Microbiol. 2004, 53, 969–983. [Google Scholar] [CrossRef]
- Gimeno, C.J.; Ljungdahl, P.O.; Styles, C.A.; Fink, G.R. Unipolar cell divisions in the yeast S. cerevisiae lead to filamentous growth: Regulation by starvation and RAS. Cell 1992, 68, 1077–1090. [Google Scholar] [CrossRef]
- Cullen, P.J.; Sprague, G.F. Glucose depletion causes haploid invasive growth in yeast. Proc. Natl. Acad. Sci. 2000, 97, 13619–13624. [Google Scholar]
- Errede, B.; Gartner, A.; Zhou, Z.; Nasmyth, K.; Ammerer, G. MAP kinase-related FUS3 from S. cerevisiae is activated by STE7 in vitro. Nature 1993, 362, 261–264. [Google Scholar]
- Liu, H.; Styles, C.; Fink, G. Elements of the yeast pheromone response pathway required for filamentous growth of diploids. Science 1993, 262, 1741–1744. [Google Scholar]
- O’Rourke, S.M.; Herskowitz, I. The Hog1 MAPK prevents cross talk between the HOG and pheromone response MAPK pathways in Saccharomyces cerevisiae. Genes Dev. 1998, 12, 2874–2886. [Google Scholar] [CrossRef]
- Vyas, V.K.; Kuchin, S.; Berkey, C.D.; Carlson, M. Snf1 Kinases with Different β-Subunit Isoforms Play Distinct Roles in Regulating Haploid Invasive Growth. Mol. Cell. Biol. 2003, 23, 1341–1348. [Google Scholar] [CrossRef]
- Prinz, S.; Avila-Campillo, I.; Aldridge, C.; Srinivasan, A.; Dimitrov, K.; Siegel, A.F.; Galitski, T. Control of Yeast Filamentous-Form Growth by Modules in an Integrated Molecular Network. Genome Res. 2004, 14, 380–390. [Google Scholar] [CrossRef] [Green Version]
- Howell, A.S.; Lew, D.J. Morphogenesis and the Cell Cycle. Genetics 2012, 190, 51–77. [Google Scholar] [CrossRef]
- Borneman, A.R.; Leigh-Bell, J.A.; Yu, H.; Bertone, P.; Gerstein, M.; Snyder, M. Target hub proteins serve as master regulators of development in yeast. Genes Dev. 2006, 20, 435–448. [Google Scholar] [CrossRef]
- Rupp, S.; Summers, E.; Lo, H.-J.; Madhani, H.; Fink, G. MAP kinase and cAMP filamentation signaling pathways converge on the unusually large promoter of the yeast FLO11 gene. EMBO J. 1999, 18, 1257–1269. [Google Scholar] [CrossRef]
- Yeh, Y.-Y.; Shah, K.H.; Herman, P.K. An Atg13 Protein-mediated Self-association of the Atg1 Protein Kinase Is Important for the Induction of Autophagy. J. Biol. Chem. 2011, 286, 28931–28939. [Google Scholar]
- Shin, C.-S.; Huh, W.-K. Bidirectional regulation between TORC1 and autophagy in Saccharomyces cerevisiae. Autophagy 2011, 7, 854–862. [Google Scholar] [CrossRef]
- De Virgilio, C.; Loewith, R. Cell growth control: Little eukaryotes make big contributions. Oncogene 2006, 25, 6392–6415. [Google Scholar] [CrossRef]
- Santhanam, A.; Hartley, A.; Düvel, K.; Broach, J.R.; Garrett, S. PP2A Phosphatase Activity Is Required for Stress and Tor Kinase Regulation of Yeast Stress Response Factor Msn2p. Eukaryotic Cell 2004, 3, 1261–1271. [Google Scholar] [CrossRef]
- Yorimitsu, T.; He, C.; Wang, K.; Klionsky, D.J. Tap42-associated protein phosphatase type 2A negatively regulates induction of autophagy. Autophagy 2009, 5, 616–624. [Google Scholar] [CrossRef]
- Natarajan, K.; Meyer, M.R.; Jackson, B.M.; Slade, D.; Roberts, C.; Hinnebusch, A.G.; Marton, M.J. Transcriptional Profiling Shows that Gcn4p Is a Master Regulator of Gene Expression during Amino Acid Starvation in Yeast. Mol. Cell. Biol. 2001, 21, 4347–4368. [Google Scholar]
- Cooper, T.G. Transmitting the signal of excess nitrogen in Saccharomyces cerevisiae from the Tor proteins to the GATA factors: Connecting the dots. FEMS Microbiol. Rev. 2002, 26, 223–238. [Google Scholar] [CrossRef]
- Bharucha, N.; Ma, J.; Dobry, C.J.; Lawson, S.K.; Yang, Z.; Kumar, A. Analysis of the Yeast Kinome Reveals a Network of Regulated Protein Localization during Filamentous Growth. Mol. Biol. Cell 2008, 19, 2708–2717. [Google Scholar] [CrossRef]
- Cutler, N.S.; Pan, X.; Heitman, J.; Cardenas, M.E. The TOR Signal Transduction Cascade Controls Cellular Differentiation in Response to Nutrients. Mol. Biol. Cell 2001, 12, 4103–4113. [Google Scholar]
- Yorimitsu, T.; Zaman, S.; Broach, J.R.; Klionsky, D.J. Protein Kinase A and Sch9 Cooperatively Regulate Induction of Autophagy in Saccharomyces cerevisiae. Mol. Biol. Cell 2007, 18, 4180–4189. [Google Scholar]
- Budovskaya, Y.V.; Stephan, J.S.; Deminoff, S.J.; Herman, P.K. An evolutionary proteomics approach identifies substrates of the cAMP-dependent protein kinase. Proc. Natl. Acad. Sci. USA 2005, 102, 13933–13938. [Google Scholar]
- Budovskaya, Y.V.; Stephan, J.S.; Reggiori, F.; Klionsky, D.J.; Herman, P.K. The Ras/cAMP-dependent Protein Kinase Signaling Pathway Regulates an Early Step of the Autophagy Process in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 20663–20671. [Google Scholar]
- Liu, H.; Styles, C.A.; Fink, G.R. Saccharomyces cerevisiae S288C Has a Mutation in FLO8, a Gene Required for Filamentous Growth. Genetics 1996, 144, 967–978. [Google Scholar]
- Wang, Z.; Wilson, W.A.; Fujino, M.A.; Roach, P.J. Antagonistic Controls of Autophagy and Glycogen Accumulation by Snf1p, the Yeast Homolog of AMP-Activated Protein Kinase, and the Cyclin-Dependent Kinase Pho85p. Mol. Cell. Biol. 2001, 21, 5742–5752. [Google Scholar]
- Zupan, J.; Raspor, P. Invasive growth of Saccharomyces cerevisiae depends on environmental triggers: A quantitative model. Yeast 2010, 27, 217–228. [Google Scholar]
- Orlova, M.; Ozcetin, H.; Barrett, L.; Kuchin, S. Roles of the Snf1-Activating Kinases during Nitrogen Limitation and Pseudohyphal Differentiation in Saccharomyces cerevisiae. Eukaryotic Cell 2010, 9, 208–214. [Google Scholar] [CrossRef]
- Shaw, B.D.; Chung, D.-W.; Wang, C.-L.; Quintanilla, L.A.; Upadhyay, S. A role for endocytic recycling in hyphal growth. Fungal Biol. 2011, 115, 541–546. [Google Scholar] [CrossRef]
- Araujo-Bazán, L.; Peñalva, M.A.; Espeso, E.A. Preferential localization of the endocytic internalization machinery to hyphal tips underlies polarization of the actin cytoskeleton in Aspergillus nidulans. Mol. Microbiol. 2008, 67, 891–905. [Google Scholar] [CrossRef]
- Upadhyay, S.; Shaw, B.D. The role of actin, fimbrin and endocytosis in growth of hyphae in Aspergillus nidulans. Mol. Microbiol. 2008, 68, 690–705. [Google Scholar] [CrossRef]
- Higuchi, Y.; Shoji, J.-Y.; Arioka, M.; Kitamoto, K. Endocytosis Is Crucial for Cell Polarity and Apical Membrane Recycling in the Filamentous Fungus Aspergillus oryzae. Eukaryotic Cell 2009, 8, 37–46. [Google Scholar] [CrossRef]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR Signaling in Growth and Metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef]
- Grossmann, G.; Opekarova, M.; Malinsky, J.; Weig-Meckl, I.; Tanner, W. Membrane potential governs lateral segregation of plasma membrane proteins and lipids in yeast. EMBO J. 2007, 26, 1–8. [Google Scholar]
- MacGurn, J.A.; Hsu, P.-C.; Smolka, M.B.; Emr, S.D. TORC1 Regulates Endocytosis via Npr1-Mediated Phosphoinhibition of a Ubiquitin Ligase Adaptor. Cell 2011, 147, 1104–1117. [Google Scholar] [CrossRef]
- Mousley, C.J.; Yuan, P.; Gaur, N.A.; Trettin, K.D.; Nile, A.H.; Deminoff, S.J.; Dewar, B.J.; Wolpert, M.; Macdonald, J.M.; Herman, P.K.; Hinnebusch, A.G.; Bankaitis, V.A. A Sterol-Binding Protein Integrates Endosomal Lipid Metabolism with TOR Signaling and Nitrogen Sensing. Cell 2012, 148, 702–715. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Song, Q.; Kumar, A. An Overview of Autophagy and Yeast Pseudohyphal Growth: Integration of Signaling Pathways during Nitrogen Stress. Cells 2012, 1, 263-283. https://doi.org/10.3390/cells1030263
Song Q, Kumar A. An Overview of Autophagy and Yeast Pseudohyphal Growth: Integration of Signaling Pathways during Nitrogen Stress. Cells. 2012; 1(3):263-283. https://doi.org/10.3390/cells1030263
Chicago/Turabian StyleSong, Qingxuan, and Anuj Kumar. 2012. "An Overview of Autophagy and Yeast Pseudohyphal Growth: Integration of Signaling Pathways during Nitrogen Stress" Cells 1, no. 3: 263-283. https://doi.org/10.3390/cells1030263
APA StyleSong, Q., & Kumar, A. (2012). An Overview of Autophagy and Yeast Pseudohyphal Growth: Integration of Signaling Pathways during Nitrogen Stress. Cells, 1(3), 263-283. https://doi.org/10.3390/cells1030263