A Review of the Applications of Chitin and Its Derivatives in Agriculture to Modify Plant-Microbial Interactions and Improve Crop Yields

Abstract
:1. Introduction
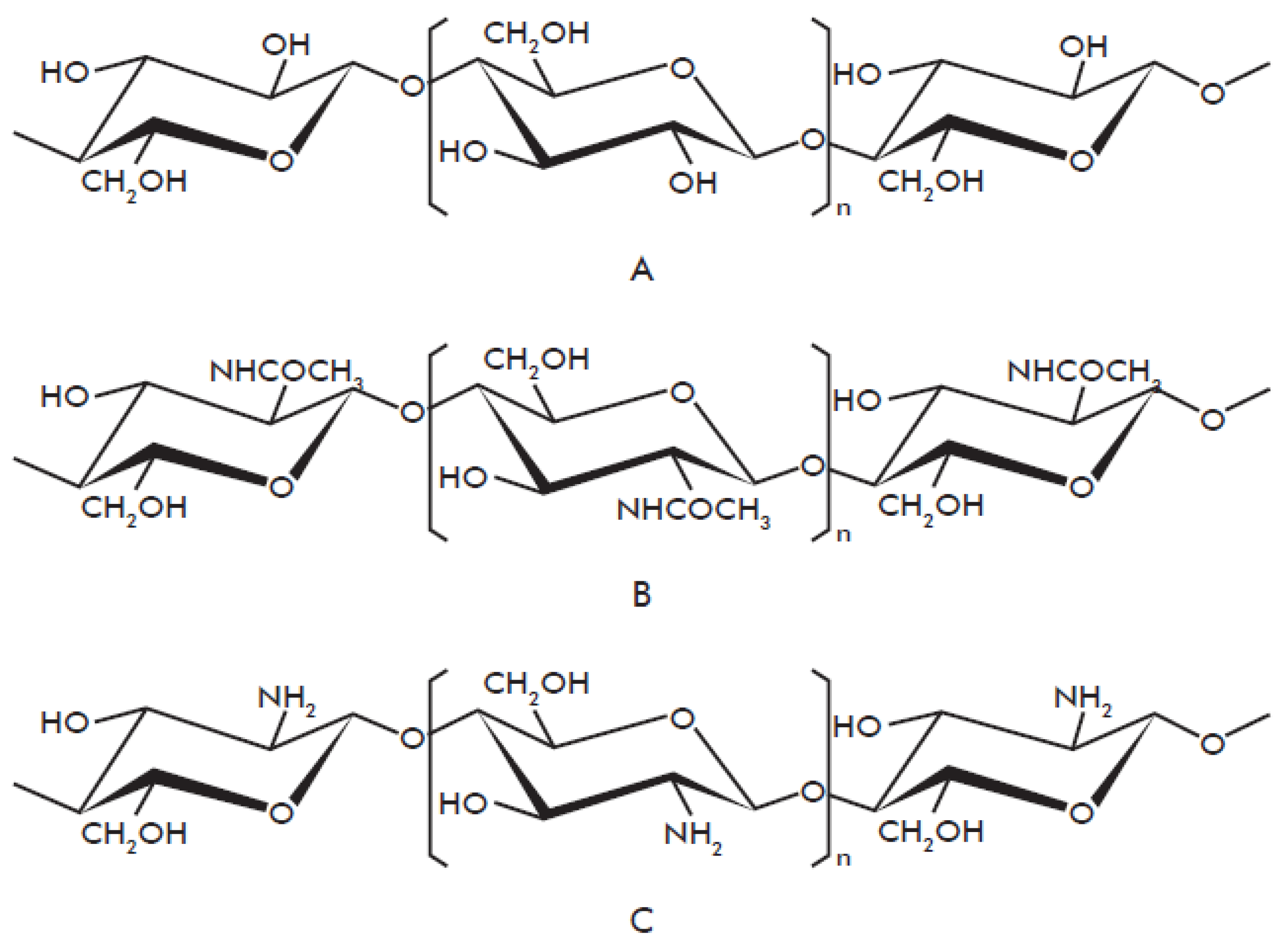
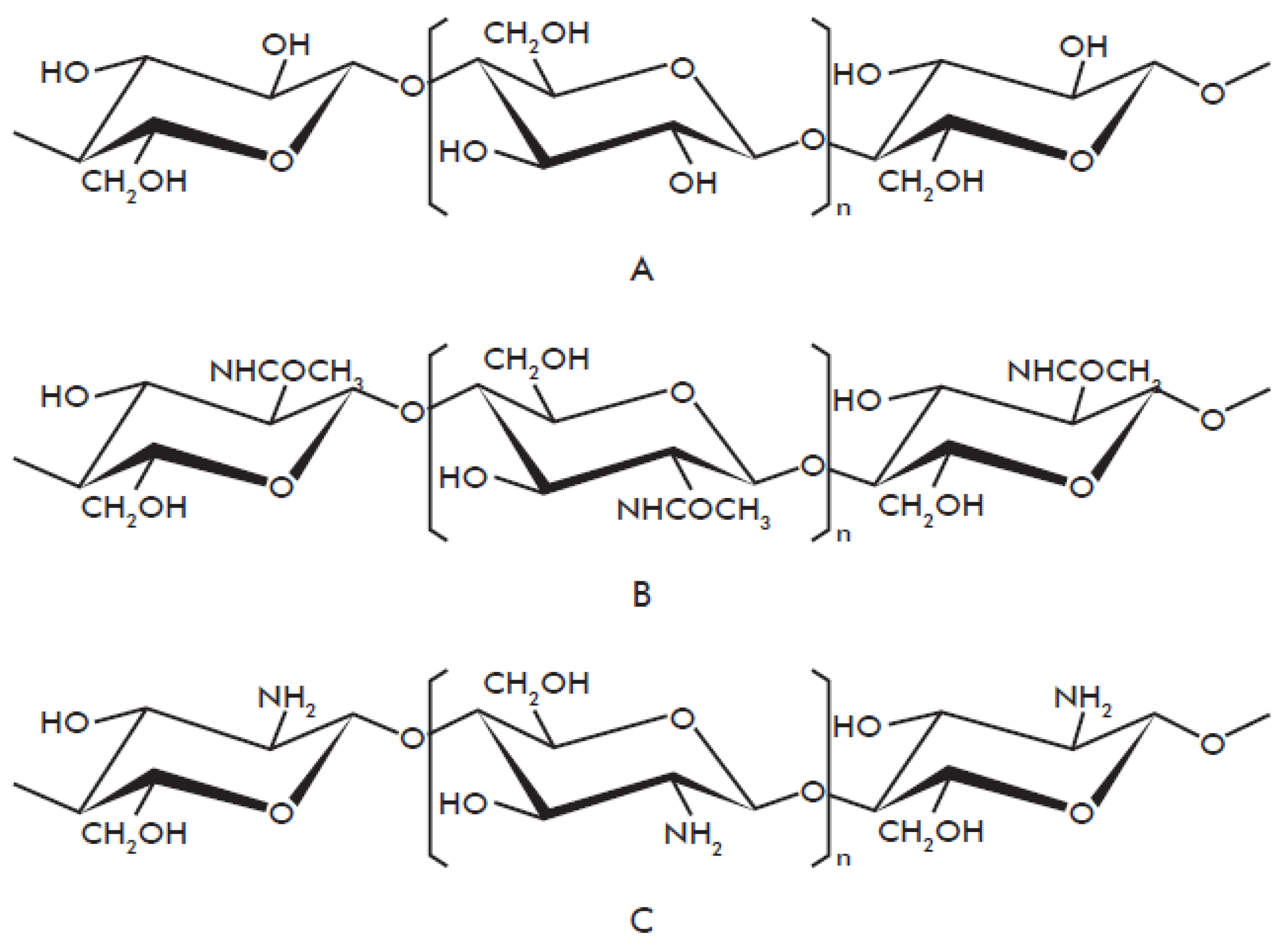
1.1. Chitin Biochemistry and Production
2. Direct Antibiosis of Chitin
2.1. Effectiveness of Chitin-Based Treatments against Fungal Pathogens
2.2. Effectiveness of Chitin-Based Treatments against Bacterial Pathogens
2.4. Effectiveness of Chitin-Based Treatments against Insect Pests
2.5. Effectiveness of Chitin-Based Treatments against Nematodes
3. Enhancement of Beneficial Microbes, both in Plant Defense and Growth
3.1. Stimulation of Antagonistic Biological Control Agents
3.2. Chitin as a Signalling Molecule for Growth-Promoting Microbes
3.3. Chitin’s Interaction with Mycorrhizal Fungi
4. Chitin-Induced Plant Defense Responses against Biotic Stress
4.1. Detection of Chitin in Plants
4.2. Chitin-Induced Defense Mechanisms
4.3. The Role of Chitinases in Plant Defense
4.4. Chitin Synthase Disruption by Pesticides
5. Regulation of Plant Growth, Development, Nutrition, and Tolerance to Abiotic Stresses
5.1. Plant Growth Promotion
5.2. Physiological Responses to Chitin Treatment
5.3. Chitin’s Effects of Plant Development
5.4. Plant Nutrition
5.5. Chitin’s Ability to Alter a Plant’S Resistance to Abiotic Stress
6. Agronomic Considerations
6.1. Efficacy and Phytotoxicity
6.2. Human/Animal Toxicity of Chitin and Chitosan
6.3. Chitin Delivery Systems
6.4. Use of Chitin-Based Treatments in Non-Field Agricultural Systems
6. Conclusions

{kind=link}
| Product class | Product type | Primary crop | Current product used | Hypothesised mode of action | Reference to evidence to chitin activity |
|---|---|---|---|---|---|
| Biocide | Fungicide | Most | Various synthetic and organic pesticides | Direct antibiosis, promotion of antagonists, induction of plant defenses. | [45,229] |
| Biocide | Oomyceticide | Most | Few of the fungicides available are effective | Disruption of endomembranes | [26] |
| Biocide | Bactericide | Most | Various synthetic and organic pesticides | Direct antibiosis, promotion of antagonists, induction of plant defenses. | [40] |
| Biocide | Viricide | Most | Few viricides currently commercially available | Induction of hypersensitivity and restriction in viral movement | [41,42,43] |
| Biocide | Nematocide | Bulbous/tuberous crops | Various synthetic and organic pesticides | Direct antibiosis, release of ammonia, promotion of antagonists | [50] |
| Biocide | Insecticide | Most | Various synthetic and organic pesticides | Direct antibiosis, promotion of antagonists, induction of plant defenses. | [45,229] |
| Biocide | Miticide | Glasshouse and orchard crops | Various synthetic and organic pesticides | Direct antibiosis, promotion of antagonists, induction of plant defenses. | No published studies identified. |
| Fertilizer | Nitrogen fertilizer | Most | Various synthetic and organic fertilizers | Utilisation of nitrogen from the chitin’s amino groups | [181,183] |
| Fertilizer | Controlled release fertilizer | Most | Resin-based fertilizer granules | Direct uptake of monomers and decomposition to ammonium by microbes | [192] |
| Growth regulator | Direct growth regulator / biostimulant/ Stress eliviator | Most | Various synthetic growth regulators and hormone inhibitors. Plus humic and fulvic acid, and seaweed extracts. | Action on specific signalling pathways; tomatal closure; direct antioxidant activity | [120,162,165,166,169,198] |
| Growth regulator | Elicitor for the production of secondary metabolites | Medicinal herbs | Jasmonates, salicylates, benzoic acid | Induction of plant biochemical defenses | [212,213,215] |
| Growth regulator | Antitranspirant | Stored fresh produce, evergreen trees | Synthetic polymers: e.g., di-1-p-Menthene | Formation of a film minimising evaporation from tissues, plus action on stomatal aperture. | [120,121] |
| Growth regulator | Wound sealant | Trees and shrubs | Wax and resin-based paints | Barrier formation, liginin and callose formation, and direct antibiosis. | [22,127,128] |
| Growth regulator | PGPR stimulator | Most | none | Supply of suitable food source and signalling compounds | [89]. No studies on hormone producing bacteria. |
| Growth regulator | Mycorrhizal stimulator | Perennial crops | none | Supply of suitable food source and signalling compounds | [65] |
| Growth regulator | Ripening retardant | Stored fresh produce | Controlled-environment storage, 1-MCP | Unknown | [225] |
| Soil conditioner | Pollution absorber | Crops growing in polluted soils | Activated charcoal, synthetic chelating agents | Cationic binding of metal ions | [199,201,202,203] |
| Soil conditioner | Water retainer | Crops in sandy/dry soils | Polyacrylamide and cellulose-based products | Gel formation | [193] |
| Biopolymer | Packaging | Stored fresh produce | Predominantly Polyethylene films | Formation of antiseptic film | [7,227,230] |
Acknowledgments
References
- Gooday, G.W. The ecology of chitin degradation. Adv. Microb. Ecol. 1990, 11, 387–419. [Google Scholar] [CrossRef]
- Gohel, V.; Singh, A.; Vimal, M.; Ashwini, P.; Chhatpar, H.S. Bioprospecting and antifungal potential of chitinolytic microorganisms. Afr. J. Biotechnol. 2006, 5, 54–72. [Google Scholar]
- Castro, S.P.M.; Paulín, E.G.L. Is chitosan a new panacea? Areas of application. the Complex World of Polysaccharides; Karunaratne, D.N., Ed.; InTech: Rijeka, Croatia, 2012. Available online: http://www.intechopen.com/books/the-complex-world-of-polysaccharides/is-chitosan-a-new-panacea-areas-of-application (accessed on 4 July 2013). [CrossRef]
- Bartnicki-Garcia, S.; Lippman, E. Fungal wall composition. In CRC Hand book of Microbiology, 2nd ed.; Laskin, A.J., Lechevalier, H.A., Eds.; CRC Press: Boca Raton, FL, USA, 1982; pp. 229–252. [Google Scholar]
- Gow, N.A.R.; Gooday, G.W. Ultrastructure of chitin in hyphae of Candida albicans and other dimorphic and mycelial fungi. Protoplasma 1983, 115, 52–58. [Google Scholar] [CrossRef]
- Bueter, C.L.; Specht, C.A.; Levitz, S.M. Innate sensing of chitin and chitosan. PLoS Pathog. 2013, 9, e1003080. [Google Scholar] [CrossRef]
- Pillai, C.K.S.; Paul, W.; Sharma, C.P. Chitin and chitosan polymers: Chemistry, solubility and fiber formation. Prog. Polym. Sci. 2009, 34, 641–678. [Google Scholar] [CrossRef]
- Muzzarelli, R.A.A. Chitin; Pergamon Press: Oxford, UK, 1977. [Google Scholar]
- Boßelmann, F.; Romano, P.; Fabritius, H.; Raabe, D.; Epple, M. The composition of the exoskeleton of two crustacea: The American lobster Homarus americanus and the edible crab Cancer pagurus. Thermochim. Acta 2007, 463, 65–68. [Google Scholar] [CrossRef]
- Ramírez, M.Á.; Rodriguez, A.T.; Alfonso, L.; Peniche, C. Chitin and its derivatives as biopolymers with potential agricultural applications. Biotecnol. Apl. 2010, 27, 270–276. [Google Scholar]
- FAO. Fishery Statistical Collections; Global Aquaculture Production. Available online: http://www.fao.org/fishery/statistics/global-aquaculture-production/en (accessed on 15 May 2013).
- Andrade, V.; Neto, B.; Fukushima, K.; Campos-Takaki, G. Effect of medium components and time of cultivation on chitin production by Mucor circinelloides (Mucor javanicus IFO 4570)—A factorial study. Revista Iberoamericana de Micología 2003, 20, 149–153. [Google Scholar]
- Law, W.M.; Lau, W.N.; Lo, K.L.; Wai, L.M.; Chiu, S.W. Removal of biocide pentachlorophenol in water system by the spent mushroom compost of Pleurotus pulmonarius. Chemosphere 2003, 52, 1531–1537. [Google Scholar] [CrossRef]
- Crestini, C.; Kovac, B.; Giovannozzi-Sermanni, G. Production and isolation of chitosan by submerged and solid-state fermentation from Lentinus edodes. Biotechnol. Bioeng. 1996, 50, 207–210. [Google Scholar] [CrossRef]
- Akin, D.E.; Condon, B.; Sohn, M.; Foulk, J.A.; Dodd, R.B.; Rigsby, L.L. Optimization for enzyme-retting of flax with pectate lyase. Ind. Crops Prod. 2007, 25, 136–146. [Google Scholar] [CrossRef]
- Yang, J.K.; Shih, I.L.; Tzeng, Y.M.; Wang, S.L. Production and purification of protease from a Bacillus subtilis that can deproteinize crustacean wastes. Enzyme Microb. Technol. 2000, 26, 406–413. [Google Scholar] [CrossRef]
- Hoell, I.A.; Vaaje-Kolstad, G.; Eijsink, V.G.H. Structure and function of enzymes acting on chitin and chitosan. Biotechnol. Genet. Eng. Rev. 2010, 27, 331–366. [Google Scholar] [CrossRef]
- Cira, L.A.; Huerta, S.; Hall, G.M.; Shirai, K. Pilot scale lactic acid fermentation of shrimp wastes for chitin recovery. Process Biochem. 2002, 37, 1359–1366. [Google Scholar] [CrossRef]
- Bowman, K.; Leong, K.W. Chitosan nanoparticles for oral drug and gene delivery. Int. J. Nanomed. 2006, 1, 117–128. [Google Scholar] [CrossRef]
- Jayakumar, R.; Prabaharan, M.; Nair, S.V.; Tamura, H. Novel chitin and chitosan nanofibers in biomedical applications. Biotechnol. Adv. 2010, 28, 142–150. [Google Scholar]
- Towheed, T.E.; Anastassiades, T.P.; Shea, B.; Houpt, J.; Welch, V.; Hochberg, M.C. Glucosamine therapy for treating osteoarthritis. Cochrane Database Syst. Reviews 2001. [Google Scholar] [CrossRef]
- Hirano, S.; Kitaura, S.; Sasaki, N.; Sakaguchi, H.; Sugiyama, M.; Hashimoto, K.; Tanatani, A. Chitin biodegradation and wound healing in tree bark tissues. J. Environ. Polym. Degrad. 1996, 4, 261–265. [Google Scholar] [CrossRef]
- Croteau, R.; Gurkewitz, S.; Johnson, M.A.; Fisk, H.J. Monoterpene and diterpene biosynthesis in lodgepole pine saplings infected with Ceratocystis clavigera or treated with carbohydrate elicitors. Plant Physiol. 1987, 85, 1123–1128. [Google Scholar] [CrossRef]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in Plant Protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef]
- Harish Prashanth, K.V.; Tharanathan, R.N. Chitin/chitosan: Modifications and their unlimited application potential—An overview. Trends Food Sci. Technol. 2007, 18, 117–131. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, X.; Han, X.; Du, Y. Antifungal activity of oligochitosan against Phytophthora capsici and other plant pathogenic fungi in vitro. Pestic. Biochem. Physiol. 2007, 87, 220–228. [Google Scholar] [CrossRef]
- Sudarshan, N.R.; Hoover, D.G.; Knorr, D. Antibacterial action of chitosan. Food Biotechnol. 1992, 6, 257–272. [Google Scholar] [CrossRef]
- Kendra, D.F.; Hadwiger, L.A. Characterization of the smallest chitosan oligomer that is maximally antifungal to Fusarium solani and elicits pisatin formation in Pisum sativum. Exp. Mycol. 1984, 8, 276–281. [Google Scholar]
- Sekiguchi, S.; Miura, Y.; Kaneko, H.; Nishimura, S.I.; Nishi, N.; Iwase, M.; Tokura, S. Molecular weight dependency of antimicrobial activity by chitosan oligomers. In Food Hydrocolloids: Structures, Properties and Functions; Nishinari, K., Doi, E., Eds.; Plenum: New York, NY, USA, 1994; pp. 71–76. [Google Scholar]
- Parra, Y.; Ramírez, M.A. Efecto de diferentes derivados de quitina sobre el crecimiento in vitro del hongo Rhizoctonia solani Kuhn. Cultivos Tropicales 2002, 23, 73–75. [Google Scholar]
- Rabea, E.I.; El Badawy, M.T.; Stevens, C.V.; Smagghe, G.; Steurbaut, W. Chitosan as antimicrobial agent: Applications and mode of action. Biomacromolecules 2003, 4, 1457–1465. [Google Scholar] [CrossRef]
- Laflamme, P.; Benhamou, N.; Bussiéres, G.; Dessureault, M. Differential effect of chitosan on root rot fungal pathogens in forest nurseries. Can. J. Bot. 1999, 77, 1460–1468. [Google Scholar] [CrossRef]
- Bell, A.A.; Hubbard, J.C.; Liu, L.; Davis, R.M.; Subbarao, K.V. Effects of chitin and chitosan on the incidence and severity of Fusarium yellows in celery. Plant Dis. 1998, 82, 322–328. [Google Scholar] [CrossRef]
- Aziz, A.; Trotel-Aziz, P.; Dhuicq, L.; Jeandet, P.; Couderchet, M.; Vernet, G. Chitosan oligomers and copper sulphate induce grapevine defense reaction and resistance to grey mould and down mildew. Phytopathology 2006, 96, 1188–1194. [Google Scholar] [CrossRef]
- Ben-shalom, N.; Ardi, R.; Pinto, R.; Aki, C.; Fallik, E. Controlling gray mould caused by Botyrtis cinerea in cucumber plants by means of chitosan. Crop Prot. 2003, 22, 285–290. [Google Scholar] [CrossRef]
- O’Herlihy, E.A.; Duffy, E.M.; Cassells, A.C. The effects of arbuscular mycorrhizal fungi band chitosan sprays on yield and late blight resistance in potato crops from microplants. Folio Geobotanica 2003, 38, 201–208. [Google Scholar]
- Muzzarelli, R.A.A.; Tarsi, R.; Filippini, O.; Giovanetti, E.; Biagini, G.; Varaldo, P.E. Antimicrobial properties of N-carboxybutyl chitosan. Antimicrob. Agents Chemother. 1990, 34, 2019–2023. [Google Scholar] [CrossRef]
- Jia, Z.; Shen, D.; Xu, W. Synthesis and antibacterial activities of quaternary ammonium salt of chitosan. Carbohydr. Res. 2001, 333, 1–6. [Google Scholar] [CrossRef]
- Ferrante, P.; Scortichini, M. Molecular and phenotypic features of Pseudomonas syringae pv. actinidiae isolated during recent epidemics of bacterial canker on yellow kiwifruit (Actinidia chinensis) in central Italy. Plant Pathol. 2010, 59, 954–962. [Google Scholar] [CrossRef]
- Vruggink, H. The effect of chitin amendment on actinomycetes in soil and on the infection of potato tubers by Streptomyces scabies. Neth. J. Plant Pathol. 1970, 76, 293–295. [Google Scholar] [CrossRef]
- Kulikov, S.N.; Chirkov, S.N.; Il’ina, A.V.; Lopatin, S.A.; Varlamov, V.P. Effect of the molecular weight of chitosan on its antiviral activity in plants. Prikl Biokhim Mikrobiol. 2006, 42, 224–228. [Google Scholar]
- Pospieszny, H.; Chirkov, S.; Atabekov, J. Induction of antiviral resistance in plants by chitosan. Plant Sci. 1991, 79, 63–68. [Google Scholar] [CrossRef]
- Faoro, F.; Sant, S.; Iriti, M.; Appiano, A. Chitosan-elicited resistance to plant viruses: A histochemical and cytochemical study. In Chitin Enzymology; Muzzarelli, R.A.A., Ed.; Atec: Grottammare, Italy, 2001; pp. 57–62. [Google Scholar]
- Chirkov, S.N. The antiviral activity of chitosan (review). Appl. Biochem. Microbiol. 2002, 38, 1–8. [Google Scholar] [CrossRef]
- Rabea, E.I.; El Badawy, M.T.; Rogge, T.M.; Stevens, C.V.; Höfte, M.; Steurbaut, W.; Smagghe, G. Insecticidal and fungicidal activity of new synthesized chitosan derivatives. Pest Manage. Sci. 2005, 61, 951–960. [Google Scholar] [CrossRef]
- Badawy, M.E.I.; El-Aswad, A.F. Insecticidal activity of chitosans of different molecular weights and chitosan-metal complexes against cotton leafworm Spodoptera littoralis and oleander aphid Aphis nerii. Plant Prot. Sci. 2012, 48, 131–141. [Google Scholar]
- Gibbs, K.E.; Morrison, F.O. The cuticle of the two-spotted spider mite, Tetranychus telarius (Linnaeus) (Acarina: Tetranychidae). Can. J. Zool. 1959, 37, 633–637. [Google Scholar] [CrossRef]
- Mothes-Wagner, U. Comparative histopathology of the chitin synthesis inhibitors nikkomycin X/Z, nikkomycin Z and polyoxin D. I: Effects on moulting, reproduction and digestion in the spider mite Tetranychus urticae. Pest Manage. Sci. 1986, 17, 607–620. [Google Scholar] [CrossRef]
- Tan, X.; Wang, S.; Li, X.; Zhang, F. Optimizing and application of microencapsulated artificial diet for Orius sauteri (Hemiptera: Anthocoridae). Acta Entomol. Sin. 2010, 53, 891–900. [Google Scholar]
- Godoy, G.; Rodriguez-Kabana, R.; Shelby, R.A.; Morgan-Jones, G. Chitin amendments for control of Meloidogyne arenaria in infested soil. II. Effects on microbial population. Nematropica 1983, 13, 63–74. [Google Scholar]
- Mian, I.H.; Godoy, G.; Shelby, R.A.; Rodriguez-Kabana, R.; Morgan-Jones, G. Chitin amendments for control of Meloidogyne arenaria in infested soil. Nematropica 1982, 12, 71–84. [Google Scholar]
- Rodriguez-Kabana, R.; Morgan-Jones, G.; Ownley-Gintis, B. Effects of chitin amendments to soil on Heterodera glycines, microbial populations, and colonization of cysts by fungi. Nematropica 1984, 14, 10–25. [Google Scholar]
- EPA. Final Registration Review Work Plan for Chitin and Chitosan 2008. Available online: http://www.epa.gov/oppsrrd1/registration_review/chitin/ (accessed on 4 July 2013).
- Westerdahl, B.B.; Carlson, H.L.; Grant, J.; Radewald, J.D.; Welch, N.; Anderson, C.A.; Darso, J.; Kirby, D.; Shibuya, F. Management of plant-parasitic nematodes with a chitin-urea soil amendment and other materials. J. Nematol. 1992, 24, 669–680. [Google Scholar]
- Belair, G.; Tremblay, N. The influence of chitin-urea amendments applied to an organic soil on a Meloidogyne hapla population and on the growth of greenhouse tomato. Phytoprotection 1995, 76, 75–80. [Google Scholar] [CrossRef]
- Rodriguez-Kabana, R.; Morgan-Jones, G.; Chet, I. Biological control of nematodes: Soil amendments and microbial antagonists. Plant Soil 1987, 100, 237–247. [Google Scholar] [CrossRef]
- Duncan, L.W. Current options for nematode management. Annu. Rev. Phytopathol. 1991, 29, 469–490. [Google Scholar] [CrossRef]
- Stirling, G.R. Biological Control of Plant-Parasitic Nematodes; CAB International: Wallingford, UK, 1991. [Google Scholar]
- Hamid, R.; Khan, M.A.; Ahmad, M.; Ahmad, M.M.; Abdin, M.Z.; Musarrat, J.; Javed, S. Chitinases: An update. J. Pharm. BioAllied Sci. 2013, 5, 21–29. [Google Scholar]
- Ayes, M.D.; Howard, S.C.; Kuzio, J.; Lopez-Ferber, M.; Possee, R.D. The complete DNA sequence of Autographa californica nuclear polyhedrosis virus. Virology 1994, 202, 586–605. [Google Scholar] [CrossRef]
- Maksimov, I.V.; Abizgil’dina, R.R.; Pusenkova, L.I. Plant growth promoting rhizobacteria as alternative to chemical crop protectors from pathogens (Review). Appl. Biochem. Microbiol. 2011, 47, 333–345. [Google Scholar] [CrossRef]
- Chandler, D.; Bailey, A.S.; Tatchell, G.M.; Davidson, G.; Greaves, J.; Grant, W.P. The development, regulation and use of biopesticides for integrated pest management. Philos. Trans. R. Soc. B. 2011, 366, 1987–1998. [Google Scholar] [CrossRef]
- Chen, F.; Wang, M.; Zheng, Y.; Luo, J.; Yang, X.; Wang, X. Quantitative changes of plant defense enzymes and phytohormone in biocontrol of cucumber Fusarium wilt by Bacillus subtilis B579. World J. Microbiol. Biotechnol. 2010, 26, 675–684. [Google Scholar] [CrossRef]
- Manjula, K.; Podile, A.R. Chitin-supplemented formulations improve biocontrol and plant growth promoting efficiency of Bacillus subtilis AF1. Can. J. Microbiol. 2001, 47, 618–625. [Google Scholar]
- Lowe, A.; Rafferty-McArdle, S.M.; Cassells, A.C. Effects of AMF- and PGPR-root inoculation and a foliar chitosan spray in single and combined treatments on powdery mildew disease in strawberry. Agric. Food Sci. 2012, 21, 28–38. [Google Scholar]
- Toyoda, H.; Matsuda, Y.; Fukamizo, T.; Nonomura, T.; Kukutani, K.; Ouchi, S. Application of chitin and chitosan degrading microbes to comprehensive biocontrol of fungal wilt pathogen, Fusarium oxysporum. In Chitin Handbook; Muzzarelli, R.A.A., Peter, M.G., Eds.; European Chitin Society, Atec: Grottammare, Italy, 1996; pp. 359–370. [Google Scholar]
- Singh, P.P.; Shin, Y.C.; Park, C.S.; Chung, Y.R. Biological control of Fusarium wilt of cucumber by chitinolytic bacteria. Phytopathology 1999, 89, 92–99. [Google Scholar] [CrossRef]
- Kishore, G.K.; Pande, S.; Podile, A.R. Chitin-supplemented foliar application of Serratia marcescens GPS 5 improves control of late leaf spot disease of groundnut by activating defense-related enzymes. J. Phytopathol. 2005, 153, 169–173. [Google Scholar] [CrossRef]
- Gooday, G.W. Physiology of microbial degradation of chitin and chitosan. Biodegradation 1990, 1, 177–190. [Google Scholar] [CrossRef]
- Sivan, A.; Chet, I. Degradation of fungal cell walls by lytic enzymes of Trichoderma haryianam. J. Gen. Microbiol. 1989, 135, 675–682. [Google Scholar]
- Lorito, M.; Harman, G.E.; Hayes, C.K.; Broadway, R.M.; Tronsmo, A.; Woo, S.L.; Di-Pietro, A. Chitinolytic enzymes produced by Trichoderma harzianum: Antifungal activity of purified endochitinase and chitobiosidase. Phytopathology 1993, 83, 302–307. [Google Scholar] [CrossRef]
- Sid Ahmed, A.; Ezziyyani, M.; Pérez Sánchez, C.; Candela, M.E. Effect of chitin on biological control activity of Bacillus spp. and Trichoderma harzianum against root rot disease in pepper (Capsicum annuum) plants. Eur. J. Plant Pathol. 2003, 109, 633–637. [Google Scholar] [CrossRef]
- López-Cervantes, J.; Rochin, K.R.F. Microbial Process and Composition for Agricultural Use. US 2012/0084886 A1, 5 April 2012. [Google Scholar]
- John, R.P.; Tyagi, R.D.; Brar, S.K.; Surampalli, R.Y.; Prévost, D. Bio-encapsulation of microbial cells for targeted agricultural delivery. Crit. Rev. Biotechnol. 2011, 31, 211–226. [Google Scholar] [CrossRef]
- El-Sayed, G.N.; Coudron, T.A.; Ignoffo, C.M. Chitinolytic activity and virulence associated with native and mutant isolates of the entomopathogenic fungus, Nomumea ileyi. J. Invertebr. Pathol. 1989, 54, 394–403. [Google Scholar] [CrossRef]
- St Leger, R.J.; Cooper, R.M.; Charnley, A.K. Characterization of chitinase and chitobiase produced by the entomopathogenic fungus Metarhizium anisopliae. J. Invertebr. Pathol. 1991, 58, 415–426. [Google Scholar] [CrossRef]
- Gill, S.S.; Cowles, E.A.; Pietrantonio, P.V. The mode of action of Bacillus thuringiensis endotoxins. Annu. Rev. Entomol. 1992, 37, 615–636. [Google Scholar] [CrossRef]
- Arora, N.; Sachdev, B.; Gupta, R.; Vimala, Y.; Bhatnagar, R.K. Characterization of a chitin-binding protein from Bacillus thuringiensis HD-1. PLoS One 2013, 8, e66603. [Google Scholar] [CrossRef]
- Ortiz-Rodríguez, T.; De La Fuente-Salcido, N.; Bideshi, D.K.; Salcedo-Hernández, R.; Barboza-Corona, J.E. Generation of chitin-derived oligosaccharides toxic to pathogenic bacteria using ChiA74, an endochitinase native to Bacillus thuringiensis. Lett. Appl. Microbiol. 2010, 51, 184–190. [Google Scholar]
- Smirnoff, W.A. Three year of aerial field experiments with Bacillus thuringiensis plus chitinase formulation against the spruce bud worm. J. Invertebr. Pathol. 1974, 24, 344–348. [Google Scholar] [CrossRef]
- Gao, Y.; Oppert, B.; Lord, J.C.; Liu, C.; Lei, Z. Bacillus thuringiensis Cry3Aa toxin increases the susceptibility of Crioceris quatuordecimpunctata to Beauveria bassiana infection. J. Invertebr. Pathol. 2012, 109, 260–263. [Google Scholar] [CrossRef]
- Wraight, S.P.; Ramos, M.E. Synergistic interaction between Beauveria bassiana- and Bacillus thuringiensis tenebrionis-based biopesticides applied against field populations of Colorado potato beetle larvae. J. Invertebr. Pathol. 2005, 90, 139–150. [Google Scholar] [CrossRef]
- Lo Scrudato, M.; Blokesch, M. A transcriptional regulator linking quorum sensing and chitin induction to render Vibrio cholerae naturally transformable. Nucleic Acids Res. 2013, 41, 3644–3658. [Google Scholar] [CrossRef]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef]
- Rudrappa, T.; Biedrzycki, M.L.; Bais, H.P. Causes and consequences of plant‐associated biofilms. FEMS Microbiol. Ecol. 2008, 64, 153–166. [Google Scholar] [CrossRef]
- Thomas, C.J.; Brown, H.L.; Hawes, C.R.; Lee, B.Y.; Min, M.K.; King, L.A.; Possee, R.D. Localization of a baculovirus-induced chitinase in the insect cell endoplasmic reticulum. J. Virol. 1998, 72, 10207–10212. [Google Scholar]
- Gopalakrishnan, B.; Muthukrishnan, S.; Kramer, K.J. Baculovirus-mediated expression of a Manduca sexta chitinase gene: Properties of the recombinant protein. Insect Biochem. Mol. Biol. 1995, 25, 255–265. [Google Scholar] [CrossRef]
- Moscardi, F.; de Souza, M.L.; de Castro, M.E.B.; Moscardi, M.L.; Szewczyk, B. Baculovirus pesticides: Present state and future perspectives. In Microbes and Microbial Technology; Springer: New York, NY, USA, 2011; pp. 415–445. [Google Scholar]
- Staehelin, C.; Schultze, M.; Tokuyasu, K.; Poinsot, V.; Promé, J.C.; Kondorosi, E.; Kondorosi, A. N-deacetylation of Sinorhizobium meliloti Nod factors increases their stability in the Medicago sativa rhizosphere and decreases their biological activity. Mol. Plant-Microbe Interact. 2000, 13, 72–79. [Google Scholar] [CrossRef]
- Berry, A.M.; McIntyre, L.; McCully, M.E. Fine structure of root hair infection leading to nodulation in the Frankia-Alnus symbiosis. Can. J. Bot. 1986, 64, 292–305. [Google Scholar] [CrossRef]
- Normand, P.; Lapierre, P.; Tisa, L.S.; Gogarten, J.P.; Alloisio, N.; Bagnarol, E.; Bassi, C.A.; Berry, A.M.; Bickhart, D.M.; Choisne, N. Genome characteristics of facultatively symbiotic Frankia sp. strains reflect host range and host plant biogeography. Genome Res. 2007, 17, 7–15. [Google Scholar]
- Cérémonie, H.; Debelle, F.; Fernandez, M.P. Structural and functional comparison of Frankia root hair deforming factor and rhizobia Nod factor. Can. J. Bot. 1999, 77, 1293–1301. [Google Scholar]
- Fitter, A.H.; Moyersoen, B. Evolutionary trends in root-microbe symbioses. Philosophical Philos. Trans. R. Soc. B. 1996, 351, 1367–1375. [Google Scholar] [CrossRef]
- Gryndler, M.; Jansa, J.; Hršelová, H.; Chvátalová, I.; Vosátka, M. Chitin stimulates development and sporulation of arbuscular mycorrhizal fungi. Appl. Soil Ecol. 2003, 22, 283–287. [Google Scholar] [CrossRef]
- Abdel-Fattah, G.M.; Mohamedin, A.H. Interactions between a vesicular-arbuscular mycorrhizal fungus (Glomus intraradices) and Streptomyces coelicolor and their effects on sorghum plants grown in soil amended with chitin of brawn scales. Biol. Fertil. Soils 2000, 32, 401–409. [Google Scholar] [CrossRef]
- El-Sayed, E.S.A.; El-Didamony, G.; El-Sayed, E.F. Effects of mycorrhizae and chitin-hydrolysing microbes on Vicia faba. World J. Microbiol. Biotechnol. 2002, 18, 505–515. [Google Scholar] [CrossRef]
- Maillet, F.; Poinsot, V.; André, O.; Puech-Pagès, V.; Haouy, A.; Gueunier, M.; Cromer, L.; Giraudet, D.; Formey, D.; Niebel, A. Fungal lipochitooligosaccharide symbiotic signals in arbuscular mycorrhizae. Nature 2011, 469, 58–63. [Google Scholar] [CrossRef]
- Oláh, B.; Brière, C.; Bécard, G.; Dénarié, J.; Gough, C. Nod factors and a diffusible factor from arbuscular mycorrhizal fungi stimulate lateral root formation in Medicago truncatula via the DMI1/DMI2 signalling pathway. Plant J. 2005, 44, 195–207. [Google Scholar] [CrossRef]
- Schultze, M.; Kondorosi, A. The role of lipochitooligosaccharides in root nodule organogenesis and plant cell growth. Curr. Opin. Genet. Dev. 1996, 6, 631–638. [Google Scholar] [CrossRef]
- Salzer, P.; Bonanomi, A.; Beyer, K.; Vögeli-Lange, R.; Aeschbacher, R.A.; Lange, J.; Wiemken, A.; Kim, D.; Cook, D.R.; Boller, T. Differential expression of eight chitinase genes in Medicago truncatula roots during mycorrhiza formation, nodulation, and pathogen infection. Mol. Plant-Microbe Interact. 2000, 13, 763–777. [Google Scholar] [CrossRef]
- Garcia-Brugger, A.; Lamotte, O.; Vandelle, E.; Bourque, S.; Lecourieux, D.; Poinssot, B.; Wendehenne, D.; Pugin, A. Early signaling events induced by elicitors of plant defenses. Mol. Plant-Microbe Interact. 2006, 19, 711–724. [Google Scholar] [CrossRef]
- Kaku, H.; Nishizawa, Y.; Minami, N.I.; Tomiyama, C.A.; Dohmae, N.; Takio, K.; Manami, E.; Shibuya, N. Plant cells recognize chitin fragments for defense signaling through a plasma membrane receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 11086–11091. [Google Scholar] [CrossRef]
- Day, R.B.; Okada, M.; Ito, Y.; Tsukada, K.; Zaghouani, H.; Shibuya, N.; Stacey, G. Binding site for chitin oligosaccharides in the soybean plasma membrane. Plant Physiol. 2001, 26, 1162–1173. [Google Scholar]
- Miya, A.; Albert, P.; Shinya, T.; Desaki, Y.; Ichimura, K.; Shirasu, K.; Narusaka, Y.; Kawakami, N.; Kaku, H.; Shibuya, N. CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsi. Proc. Natl. Acad. Sci. USA 2007, 104, 19613–19618. [Google Scholar] [CrossRef]
- Shinya, T.; Motoyama, N.; Ikeda, A.; Wada, M.; Kamiya, K.; Hayafune, M.; Kaku, H.; Shibuya, N. Functional characterization of CEBiP and CERK1 homologs in arabidopsis and rice reveals the presence of different chitin receptor systems in plants. Plant Cell Physiol. 2012, 53, 1696–1706. [Google Scholar] [CrossRef]
- Nojiri, H.; Sugimori, M.; Yamane, H.; Nishimura, Y.; Yamada, A.; Shibuya, N.; Kodama, O.; Murofushi, N.; Ohmori, T. Involvement of jasmonic acid in elicitor-induced phytoalexin production in suspension-cultured rice cells. Plant Physiol. 1996, 110, 387–392. [Google Scholar]
- Ott, P.G.; Varga, G.J.; Szatmári, A.; Bozsó, Z.; Klement, E.; Medzihradszky, K.F.; Besenyei, E.; Czelleng, A.; Klement, Z. Novel extracellular chitinases rapidly and specifically induced by general bacterial elicitors and suppressed by virulent bacteria as a marker of early basal resistance in tobacco. Mol. Plant-Microbe Interact. 2006, 19, 161–172. [Google Scholar] [CrossRef]
- Mentlak, T.A.; Kombrink, A.; Shinya, T.; Ryder, L.S.; Otomo, I.; Saitoh, H.; Terauchi, R.; Nishizawa, Y.; Shibuya, N.; Thomma, B.P.H.J.; Talbot, N.J. Effector-mediated suppression of chitin-triggered immunity by Magnaporthe oryzae is necessary for rice blast disease. The Plant Cell 2012, 24, 322–335. [Google Scholar] [CrossRef]
- Fujikawa, T.; Sakaguchim, A.; Nishizawa, Y.; Kouzai, Y.; Minami, E.; Yano, S.; Koga, H.; Meshi, T.; Nishimura, M. Surface α-1,3-glucan facilitates fungal stealth infection by interfering with innate immunity in plants. PLoS Pathog. 2012, 8, e1002882. [Google Scholar] [CrossRef]
- Minami, E.; Kuchitsu, K.; He, D.Y.; Kouchi, H.; Midoh, N.; Ohtsuki, Y.; Shibuya, N. Two novel genes rapidly and transiently activated in suspension-cultured rice cells by treatment with N-acetylchitoheptaose, a biotic elicitor for phytoalexin production. Plant Cell Physiol. 1996, 37, 563–567. [Google Scholar] [CrossRef]
- Nishizawa, Y.; Kawakami, A.; Hibi, T.; He, D.Y.; Shibuya, N.; Minami, E. Regulation of the chitinase gene expression in suspension-cultured rice cells by N-acetylchitooligosaccharides: differences in the signal transduction pathways leading to the activation of elicitor-responsive genes. Plant Mol. Biol. 1999, 39, 907–914. [Google Scholar] [CrossRef]
- Takai, R.; Hasegawa, K.; Kaku, K.; Shibuya, N.; Minami, E. Isolation and analysis of expression mechanisms of a rice gene, EL5, which shows structural similarity to ATL family from Arabidopsis, in response to N-acetylchitooligosaccharide elicitor. Plant Sci. 2001, 160, 577–583. [Google Scholar] [CrossRef]
- Rakwal, R.; Tamogami, S.; Agrawal, G.K.; Iwahashi, H. Octadecanoid signaling component “burst” in rice (Oryza sativa L.) seedling leaves upon wounding by cut and treatment with fungal elicitor chitosan. Biochem. Biophys. Res. Commun. 2002, 295, 1041–1045. [Google Scholar] [CrossRef]
- Doares, S.H.; Syrovets, T.; Weiler, E.W.; Ryan, C.A. Oligogalacturonides and chitosan activate plant defensive genes through the octadecanoid pathway. Proc. Natl. Acad. Sci. USA 1995, 92, 4095–4098. [Google Scholar] [CrossRef]
- Bohland, C.; Balkenhohl, T.; Loers, G.; Feussner, I.; Grambow, H.J. Differential induction of lipoxygenase isoforms in wheat upon treatment with rust fungus elicitor, chitin oligosaccharides, chitosan and methyl jasmonate. Plant Physiol. 1997, 114, 679–685. [Google Scholar]
- Linden, J.C.; Phisalaphong, M. Oligosaccharides potentiate methyl jasmonate-induced production of paclitaxel in Taxus Canadensis. Plant Sci. 2000, 158, 41–51. [Google Scholar] [CrossRef]
- Walker-Simmons, M.; Jin, D.; West, C.A.; Hadwiger, L.; Ryan, C.A. Comparison of proteinase inhibitor-inducing activities and phytoalexin elicitor activities of a pure fungal endopolygalacturonase, pectic fragments and chitosans. Plant Physiol. 1984, 76, 833–836. [Google Scholar] [CrossRef]
- Farmer, E.E.; Ryan, C.A. Octadecanoid precursors of jasmonic acid activate the synthesis of wound-inducible proteinase inhibitors. Plant Cell 1992, 4, 129–134. [Google Scholar]
- Iriti, M.; Faoro, F. Abscisic acid is involved in chitosan-induced resistance to tobacco necrosis virus (TNV). Plant Physiol. Biochem. 2008, 46, 1106–1111. [Google Scholar] [CrossRef]
- Bittelli, M.; Flury, M.; Campbell, G.S.; Nichols, E.J. Reduction of transpiration through foliar application of chitosan. Agr. Forest. Meteorol. 2001, 107, 167–175. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Vasyukova, N.I.; Zinoveva, L.I.; Iĺinskaya, E.A.; Perekhod, G.I.; Chalenko, N.G.; Iĺina, A.V.; Varlamov, V.P.; Ozeretskovskaya, O.L. Modulation of plant resistance to diseases by water-soluble chitosan. Appl. Biochem. Microbiol. 2001, 37, 103–109. [Google Scholar] [CrossRef]
- Kuchitsu, K.; Kikuyama, M.; Shibuya, N. N-Acetylchitooligosaccharides, biotic elicitor for phytoalexin production, induce transient membrane depolarization in suspension-cultured rice cells. Protoplasma 1993, 174, 79–81. [Google Scholar] [CrossRef]
- El Hassni, M.; El Hadrami, A.; Daayf, F.; Chérif, M.; Ait Barka, E.; El Hadrami, I. Chitosan, antifungal product against Fusarium oxysporum f. sp. albedinis and elicitor of defense reactions in date palm roots. Phytopathol. Mediterr. 2004, 43, 195–204. [Google Scholar]
- Kuchitsu, K.; Kosaka, H.; Shiga, T.; Shibuya, N. EPR evidence for generation of hydroxyl radical triggered by N-acetylchitooligosaccharide elicitor and a protein phosphatase inhibitor in suspension-cultured rice cells. Protoplasma 1995, 188, 138–142. [Google Scholar] [CrossRef]
- Amborabé, B.-E.; Bonmort, J.; Fleurat-Lessard, P.; Roblin, G. Early events induced by chitosan on plant cells. J. Exp. Bot. 2008, 59, 2317–2324. [Google Scholar] [CrossRef]
- Köhle, H.; Jeblick, W.; Poten, F.; Blaschek, W.; Kauss, H. Chitosan-elicited callose synthesis in soybean cells as a Ca2+-dependent process. Plant Physiol. 1985, 77, 544–551. [Google Scholar] [CrossRef]
- Pearce, R.B.; Ride, J.P. Chitin and related compounds as elicitors of the lignification response in wounded wheat leaves. Physiol. Plant Pathol. 1982, 20, 119–123. [Google Scholar] [CrossRef]
- Lafontaine, J.P.; Benhamou, N. Chitosan treatment: An emerging strategy for enhancing resistance of greenhouse tomato plants to infection by Fusarium oxysporum f. sp. radicislycopersici. Biocontrol Sci. Tech. 1996, 6, 111–124. [Google Scholar] [CrossRef]
- Shigo, A.L. Compartmentalization: a conceptual framework for understanding how trees grow and defend themselves. Annu. Rev. Phytopathol. 1984, 22, 189–214. [Google Scholar] [CrossRef]
- Vasil’ev, L.A.; Dzyubinskaya, E.V.; Zinovkin, R.A.; Kiselevsky, D.B.; Lobysheva, N.V.; Samuilov, V.D. Chitosan-induced programmed cell death in plants. Biochemistry (Moscow Russ. Fed.) 2009, 74, 1035–1043. [Google Scholar] [CrossRef]
- Lizama-Uc, G.; Estrada-Mota, I.A.; Caamal-Chan, M.G.; Souza-Perera, R.; Oropeza-Salín, C.; Islas-Flores, I.; Zúñiga-Aguilar, J.J. Chitosan activates a MAP-kinase pathway and modifies abundance of defense-related transcripts in calli of Cocos nucifera L. Physiol. Mol. Plant Pathol. 2007, 70, 130–141. [Google Scholar] [CrossRef]
- Schlumbaum, A.; Mauch, F.; Vogeli, U.; Boller, T. Plant chitinases are potent inhibitors of fungal growth. Nature 1986, 324, 365–367. [Google Scholar] [CrossRef]
- Herrera-Estrella, A.; Chet, I. Chitinases in biological control. Experientia. Suppl. 1999, 87, 171–184. [Google Scholar]
- Kramer, K.J.; Muthukrishnan, S.; Lowell, J.; White, F. Chitinases for insect control. In Advances in Insect Control: The Role of Transgenic Plants; Carozzi, N., Koziel, M., Eds.; Taylor and Francis: Bristol, UK, 1997; pp. 185–193. [Google Scholar]
- Huang, J.K.; Wen, L.; Swegle, M.; Tran, H.C.; Thin, T.H.; Naylor, H.M.; Muthukrishnan, S.; Reeck, G.R. Nucleotide sequence of a rice genomic clone that encodes a class I endochitinase. Plant Mol. Biol. 1991, 16, 479–480. [Google Scholar] [CrossRef]
- Ding, X.; Gopalakrishnan, B.; Johnson, L.B.; White, F.F.; Wang, X.; Morgan, T.D.; Kramer, K.J.; Muthukrishnan, S. Insect resistance of transgenic tobacco expressing an insect chitinase gene. Transgenic Res. 1998, 7, 77–84. [Google Scholar] [CrossRef]
- Someya, N.; Akutsu, K. Biocontrol of plant diseases by genetically modified microorganisms: Current status and future prospects. In PGPR: Biocontrol and Biofertilization; Siddiqui, Z.A., Ed.; Springer: Dordrecht, the Netherlands, 2006; pp. 297–312. [Google Scholar]
- Sitrit, Y.; Barak, Z.; Kapulnik, Y.; Oppenheim, A.B.; Chet, I. Expression of Serratia marcescens chitinase gene in Rhizobium meliloti during symbiosis on alfalfa roots. Mol. Plant-Microbe Interact. 1993, 6, 293–298. [Google Scholar] [CrossRef]
- Downing, K.J.; Thomson, J.A. Introduction of the Serratia marcescens chiA gene into an endophytic Pseudomonas fluorescens for the biocontrol of phytopathogenic fungi. Can. J. Microbiol. 2000, 46, 363–369. [Google Scholar]
- Lopez, O.; Fernandez-Bolanos, J.G.; Gil, M.V. New trends in pest control: the search for greener insecticides. Green Chem. 2005, 7, 431–442. [Google Scholar] [CrossRef]
- Abou-Elghar, G.E.; Fujiyoshi, P.; Matsumura, F. Significance of the sulfonylurea receptor (SUR) as the target of diflubenzuron in chitin synthesis inhibition in Drosophila melanogaster and Blattella germanica. Insect Biochem. Mol. Biol. 2004, 34, 743–752. [Google Scholar] [CrossRef]
- EPA. Green Chemistry, Designing Safer Chemicals Award 2000. Available online: http://www.epa.gov/gcc/dsca00.html (accessed on 4 July 2013).
- Bayoumi, A.E.; Pérez-Pertejo, Y.; Zidan, H.Z.; Balaña-Fouce, R.; Ordóñez, C.; Ordóñez, D. Cytotoxic effects of two antimolting insecticides in mammalian CHO-K1 cells. Ecotoxicol. Environ. Saf. 2003, 55, 19–23. [Google Scholar] [CrossRef]
- Peppuy, A.; Robert, A.; Delbecque, J.P.; Leca, J.L.; Rouland, C.; Bordereau, C. Efficacy of hexaflumuron against the fungus-growing termite Pseudacanthotermes spiniger (Sjöstedt) (isopteran, macrotermitinae). Pest Manage. Sci. 1998, 54, 22–26. [Google Scholar] [CrossRef]
- Liu, T.X.; Stansly, P.A. Lethal and sublethal effects of two insect growth regulators on adult Delphastus catalinae (Coleoptera Coccinellidae) predator whiteflies (Homoptera Aleyrodidae). Biol. Control 2004, 30, 298. [Google Scholar] [CrossRef]
- Berecibar, A.; Granjean, C.; Siriwardena, A. Synthesis and biological activity of natural aminocyclopentitol glycosidase inhibitors: Mannostatins, trehazolin, allosamidins, and their analogues. Chem. Rev. 1999, 99, 779–844. [Google Scholar] [CrossRef]
- Soderlund, D.M.; Clark, J.M.; Sheets, L.P.; Mullin, L.S.; Piccirillo, V.J.; Sargent, D.; Stevens, J.T.; Weiner, M.L. Mechanisms of pyrethroid neurotoxicity: Implications for cumulative risk assessment. Toxicology 2002, 171, 3–59. [Google Scholar] [CrossRef]
- Cohen, E. Chitin synthesis and inhibition: a revisit. Pest Manage. Sci. 2001, 57, 946–950. [Google Scholar] [CrossRef]
- Tang, B.; Wei, P.; Chen, J.; Wang, S.G.; Zhang, W.Q. Progress in gene features and functions of insect trehalases. Acta Entomol. Sin. 2012, 55, 1315–1321. [Google Scholar]
- Qian, X.; Liu, Z.; Li, Z.; Song, G. Synthesis and quantitative structure-activity relationships of fluorine-containing 4,4-dihydroxylmethyl-2-aryliminooxazo (thiazo) lidines as trehalase inhibitors. J. Agric. Food Chem. 2001, 49, 5279–5284. [Google Scholar] [CrossRef]
- Crimmins, M.T.; Tabet, E.A. Formal total synthesis of (+)-trehazolin. Application of an asymmetric Aldol-Olefin metathesis approach to the synthesis of functionalized cyclopentenes. J. Org. Chem. 2001, 66, 4012–4018. [Google Scholar] [CrossRef]
- Ando, O.; Satake, H.; Itoi, K.; Sato, A.; Nakajima, M.; Takahashi, S.; Haruyama, H.; Ohkuma, J.; Kinoshita, I.; Enokita, R. Trehazolin, a new trehalase inhibitor. J. Antibiot. 1991, 4, 1165–1168. [Google Scholar]
- Hirsch, A.M.; Valdés, M. Micromonospora: An important microbe for biomedicine and potentially for biocontrol and biofuels. Soil Biol. Biochem. 2010, 42, 536–542. [Google Scholar] [CrossRef]
- Gaughran, J.P.; Lai, M.H.; Kirsch, D.R.; Silverman, S.J. Nikkomycin Z is a specific inhibitor of Saccharomyces cerevisiae chitin synthase isozyme Chs3 in vitro and in vivo. J. Bacteriol. 1994, 176, 5857–5860. [Google Scholar]
- Li, R.K.; Rinaldi, M.G. In vitro antifungal activity of nikkomycin Z in combination with fluconazole or itraconazole. Antimicrob. Agents Chemother. 1999, 43, 1401–1405. [Google Scholar]
- EPA. Consideration of Eligibility for Registration of the New Pesticide Active Ingredient Polyoxin D Zinc Salt-DECISION MEMORANDUM 2003. Available online: http://www.epa.gov/pesticides/chem_search/reg_actions/registration/related_PC-230000_1-Jul-03.pdf (accessed on 4 July 2013).
- Bixby-Brosi, A.J.; Potter, D.A. Can a chitin-synthesis-inhibiting turfgrass fungicide enhance black cutworm susceptibility to a baculovirus? Pest Manage. Sci. 2012, 68, 324–329. [Google Scholar] [CrossRef]
- Tsugita, T.; Takahashi, K.; Muraoka, T.; Fukui, H. The application of chitin/chitosan for agriculture. (in Japanese). In Proceedings of Special Session of the 7th Symposium on Chitin and Chitosan; Japanese Society for Chitin and Chitosan: Fukui, Japan, 1993; pp. 21–22. [Google Scholar]
- Lee, Y.S.; Kim, Y.H.; Kim, S.B. Changes in the respiration, growth, and vitamin C content of soybean sprouts in response to chitosan of different molecular weights. HortScience 2005, 40, 1333–1335. [Google Scholar]
- Kim, H.J.; Chen, F.; Wang, X.; Rajapakse, N.C. Effect of chitosan on the biological properties of sweet basil (Ocimum basilicum L.). J. Agric. Food Chem. 2005, 53, 3696–3701. [Google Scholar] [CrossRef]
- Ait Barka, E.; Eullaffroy, P.; Clément, C.; Vernet, G. Chitosan improves development, and protects Vitis vinifera L. against Botrytis cinerea. Plant Cell Rep. 2004, 22, 608–614. [Google Scholar] [CrossRef]
- Wanichpongpan, P.; Suriyachan, K.; Chandrkrachang, S. Effects of Chitosan on the growth of Gerbera flower plant (Gerbera jamesonii). In Chitin and Chitosan in Life Science, Proceedings of the Eighth International Chitin and Chitosan Conference and Fourth Asia Pacific Chitin and Chitosan Symposium, Yamaguchi, Japan, 21–23 September 2000; Uragami, T., Kurita, K., Fukamizo, T., Eds.; pp. 198–201.
- Chandrkrachang, S. The applications of chitin in agriculture in Thailand. Adv. Chitin Sci. 2002, 5, 458–462. [Google Scholar]
- Pornpeanpakdee, P.; Pichyangkura, R.; Chadchawan, S.; Limpanavech, P. Chitosan effects on Dendrobium ‘Eiskul’ Protocorm-like body production. In Proceedings of the 31st Congress on Science and Technology of Thailand, Nakornrachaseema, Thailand, 18–20 October 2005; pp. 1–3.
- Nahar, S.J.; Kazuhiko, S.; Haque, S.M. Effect of Polysaccharides Including Elicitors on Organogenesis in Protocorm-like Body (PLB) of Cymbidium insigne in vitro. J. Agric. Sci. Technol. 2012, 2, 1029–1033. [Google Scholar]
- Chibu, H.; Shibayama, H.; Arima, S. Effects of chitosan application on the shoot growth of rice and soybean. Jpn. J. Crop Sci. 2002, 71, 206–211. [Google Scholar]
- Khan, W.; Prithiviraj, B.; Smith, D.L. Effect of foliar application of chitin and chitosan oligosaccharides on photosynthesis of maize and soybean. Photosynth. Res. 2002, 40, 621–624. [Google Scholar] [CrossRef]
- Sauerwein, M.; Flores, H.M.; Yamazaki, T.; Shimomura, K. Lippia dulcis shoot cultures as a source of the sweet sesquiterpene hernandulcin. Plant Cell Rep. 1991, 9, 663–666. [Google Scholar]
- Herde, O.; Peña-cortés, H.; Willmitzer, L.; Fisahn, J. Stomatal responses to jasmonic acid, linolenic acid and abscisic acid in wild-type and ABA-deficient tomato plants. Plant Cell Environ. 1997, 20, 136–141. [Google Scholar]
- Issak, M.; Okuma, E.; Munemasa, S.; Nakamura, Y.; Mori, I.C.; Murata, Y. Neither endogenous abscisic acid nor endogenous jasmonate is involved in salicylic acid-, yeast elicitor-, or chitosan-induced stomatal closure in Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2013, 77, 1111–1113. [Google Scholar] [CrossRef]
- Limpanavech, P.; Pichyangkura, R.; Khunwasi, C.; Chadchawan, S.; Lotrakul, P.; Bunjongrat, P.; Chaidee, A.; Akaraeakpanya, T. The effects of polymer type, concentration and %DD of bicatalyte modigied chitosan on flora production of Dendrobium ‘Eiskul’. In Proceedings of the National chitin-chitosan conference, Chulalongkorn University, Bangkok, Thailand, 17–18 July 2003; pp. 60–64.
- Utsunomiya, N.; Kinai, H. Effect of chitosan-oligosaccharides soil conditioner on the growth of passionfruit. J. Jpn. Soc. Hortic. Sci. 1994, 64, 176–177. [Google Scholar]
- Ohta, K.; Tanguchi, A.; Konishi, N.; Hosoki, T. Chitosan treatment affects plant growth and flower quality in Eustoma grandiflorum. HortScience 1999, 34, 233–234. [Google Scholar]
- Uddin, A.F.M.J.; Hashimoto, F.; Shimiza, K.; Sakata, Y. Monosaccharides and chitosan sensing in bud growth and petal pigmentation in Eustoma grandiflorum (Raf.) Shinn. Sci. Hortic. 2004, 100, 127–138. [Google Scholar] [CrossRef]
- Grover, A. Plant chitinases: genetic diversity and physiological roles. Crit. Rev. Plant Sci. 2012, 31, 57–73. [Google Scholar] [CrossRef]
- Guan, Y.J.; Hu, J.; Wang, X.J.; Shao, C.X. Seed priming with chitosan improves maize germination and seedling growth in relation to physiological changes under low temperature stress. J. Zhejiang Univ. Sci. B. 2009, 10, 427–433. [Google Scholar] [CrossRef]
- Bhaskara Reddy, M.V.; Arul, J.; Angers, P.; Couture, L. Chitosan treatment of wheat seeds induces resistance to Fusarium graminearum and improves seed quality. J. Agric. Food Chem. 1999, 47, 1208–1216. [Google Scholar] [CrossRef]
- Manjula, K.; Podile, A.R. Increase in seedling emergence and dry weight of pigeon pea in the field with chitin-supplemented formulations of Bacillus subtilis AF 1. World J. Microbiol. Biotechnol. 2005, 21, 1057–1062. [Google Scholar] [CrossRef]
- Malfanova, N.; Kamilova, F.; Validov, S.; Shcherbakov, A.; Chebotar, V.; Tikhonovich, I.; Lugtenberg, B. Characterization of Bacillus subtilis HC8, a novel plant-beneficial endophytic strain from giant hogweed. Microb. Biotechnol. 2011, 4, 523–532. [Google Scholar] [CrossRef]
- Yen, M.T.; Mau, J.L. Selected physical properties of chitin prepared from shiitake stipes. Food Sci. Technol. 2007, 40, 558–563. [Google Scholar]
- White, R.E. Principles and Practice of Soil Science: The Soil as a Natural Resource, 4th ed.; Blackwell: Oxford, UK, 2006. [Google Scholar]
- Spiegel, Y.; Kafkafi, U.; Pressman, E. Evaluation of a protein-chitin derivative of crustacean shells as a slow-release nitrogen fertilizer on Chinese cabbage. J. Hortic. Sci. 1988, 63, 621–628. [Google Scholar]
- Roberts, P.; Jones, D.L. Microbial and plant uptake of free amino sugars in grassland soils. Soil Biol. Biochem. 2012, 49, 139–149. [Google Scholar] [CrossRef]
- Yaroslavtsev, A.; Manucharova, N.; Stepanov, A.; Zvyagintsev, D.; Sudnitsyn, I. Microbial destruction of chitin in soils under different moisture conditions. Eurasian Soil Sci. 2009, 42, 797–806. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Antibus, R.K.; Linkins, A.E.; McClaugherty, C.A.; Rayburn, L.; Repert, D.; Weiland, T. Wood decomposition: nitrogen and phosphorus dynamics in relation to extracellular enzyme activity. Ecology 1993, 74, 1586–1593. [Google Scholar] [CrossRef]
- Rich, J.R.; Hodge, C.H. Utilization of blue crab scrap compost to suppress Meloidogyne javanica on tomato. Nematropica 1993, 23, 1–5. [Google Scholar]
- Bohn, H.L.; Myer, R.A.; O’Connor, G.A. Soil Chemistry; John Wiley & Sons: New Jersey, NJ, USA, 2002. [Google Scholar]
- Sukwattanasinitt, M.; Klaikherd, A.; Skulnee, K.; Aiba, S. Chitosan as releasing device for 2,4-D herbicide. In Chitin and Chitosan in Life Science, Proceedings of the Eighth International Chitin and Chitosan Conference and Fourth Asia Pacific Chitin and Chitosan Symposium, Yamaguchi, Japan, 21–23 September 2000; Uragami, K., kurita, K., Fukamizo, T., Eds.; pp. 198–201.
- Hadwiger, L.A.; McBride, P.O. Low-level copper plus chitosan applications provide protection against late blight of potato. Plant Health Progress. 2006. Available online: http://www.plantmanagementnetwork.org/pub/php/research/2006/chitosan (accessed on 23 August 2013).
- Tamura, H.; Nagahama, H.; Tokura, S. Preparation of chitin hydrogel under mild conditions. Cellulose 2006, 13, 357–364. [Google Scholar] [CrossRef]
- Jamnongkan, T.; Kaewpirom, S. Potassium release kinetics and water retention of controlled-release fertilizers based on chitosan hydrogels. J. Polym. Environ. 2010, 18, 413–421. [Google Scholar] [CrossRef]
- Wu, L.; Liu, M.; Liang, R. Preparation and properties of a double-coated slow-release NPK compound fertilizer with superabsorbent and water-retention. Bioresour. Technol. 2008, 99, 547–554. [Google Scholar] [CrossRef]
- Xie, W.; Xu, P.; Liu, Q. Antioxidant activity of water-soluble chitosan derivatives. Bioorg. Med. Chem. Lett. 2001, 11, 1699–1701. [Google Scholar] [CrossRef]
- Sun, T.; Zhou, D.; Xie, J.; Mao, F. Preparation of chitosan oligomers and their antioxidant activity. Eur. Food Res. Technol. 2006, 225, 451–456. [Google Scholar]
- Sun, T.; Yao, Q.; Zhou, D.; Mao, F. Antioxidant activity of N-carboxymethyl chitosan oligosaccharides. Bioorg. Med. Chem. Lett. 2008, 18, 5774–5776. [Google Scholar] [CrossRef]
- Chen, W.G.; Liu, X.; Chen, H.X. Preparation of modified chitosan with quaternary ammonium salt. Textile Bioengineering and Informatics Symposium Proceedings 2009, 1, 226–230. [Google Scholar]
- Boonlertnirun, S.; Sarobol, E.D.; Meechoui, S.; Sooksathan, I. Drought recovery and grain yield potential of rice after chitosan application. Kasetsart J. Nat. Sci. 2007, 41, 1–6. [Google Scholar]
- Maruca, R.; Suder, B.J.; Wightman, J.P. Interaction of heavy metals with chitin and chitosan. III. Chromium. J. Appl. Polym. Sci. 1982, 27, 4827–4837. [Google Scholar] [CrossRef]
- Correa-Murrieta, M.A.; López-Cervantes, J.; Sánchez-Machado, D.I.; Sánchez-Duarte, R.G.; Rodríguez-Núñez, J.R.; Núñez-Gastélum, J.A. Fe(II) and Fe(III) adsorption by chitosan-tripolyphosphate beads: Kinetic and equilibrium studies. J. Water Supply Res. Technol. AQUA 2012, 61, 331–341. [Google Scholar] [CrossRef]
- Babel, S.; Kurniawan, T.A. Low-cost adsorbents for heavy metals uptake from contaminated water: a review. J. Hazard. Mater. 2003, 97, 219–243. [Google Scholar] [CrossRef]
- Bailey, S.E.; Olin, T.J.; Bricka, R.M.; Adrian, D.D. A review of potentially low-cost sorbents for heavy metals. Water Res. 1999, 33, 2469–2479. [Google Scholar] [CrossRef]
- Sánchez-Duarte, R.G.; Sánchez-Machado, D.I.; López-Cervantes, J.; Correa-Murrieta, M.A. Adsorption of allura red dye by cross-linked chitosan from shrimp waste. Water Sci. Technol. 2012, 65, 618–623. [Google Scholar] [CrossRef]
- Wang, F.Y.; Lin, X.G.; Yin, R. Role of microbial inoculation and chitosan in phytoextraction of Cu, Zn, Pb and Cd by Elsholtzia splendens—A field case. Environ. Pollut. 2007, 147, 248–255. [Google Scholar] [CrossRef]
- Angelim, A.L.; Costa, S.P.; Farias, B.C.; Aquino, L.F.; Melo, V.M. An innovative bioremediation strategy using a bacterial consortium entrapped in chitosan beads. J. Environ. Manage. 2013, 127, 10–17. [Google Scholar] [CrossRef]
- Gentili, A.R.; Cubitto, M.A.; Ferrero, M.; Rodriguéz, M.S. Bioremediation of crude oil polluted seawater by a hydrocarbon-degrading bacterial strain immobilized on chitin and chitosan flakes. Int. Biodeterior. Biodegrad. 2006, 57, 222–228. [Google Scholar] [CrossRef]
- El Ghaouth, A.; Arul, J.; Asselin, A.; Benhamou, N. Antifungal activity of chitosan on postharvest pathogens: Induction of morphological and cytological alterations in Rhizopus stolonifer. Mycol. Res. 1992, 96, 769–779. [Google Scholar] [CrossRef]
- Palma-Guerrero, J.; Jansson, H.B.; Salinas, J.; Lopez-Llorca, L.V. Effect of chitosan on hyphal growth and spore germination of plant pathogenic and biocontrol fungi. J. Appl. Microbiol. 2008, 104, 541–553. [Google Scholar]
- Benhamou, N.; Lafontaine, P.J.; Nicole, M. Induction of systemic resistance to Fusarium crown and root rot in tomato plants by seed treatment with chitosan. Phytopathology 1994, 84, 1432–1444. [Google Scholar] [CrossRef]
- Scorza, R.; Callahan, A.; Levy, L.; Damsteegt, V.; Webb, K.; Ravelonandro, M. Post-transcriptional gene silencing in plum pox virus resistant transgenic European plum containing the plum pox potyvirus coat protein gene. Transgenic Res. 2001, 10, 201–209. [Google Scholar] [CrossRef]
- Levy, L.; Damsteegt, V.; Scorza, R.; Kolber, M. Plum pox potyvirus disease of stone fruits. American Phytopathology Society. 2000. Available online: http://www.apsnet.org/online/feature/plumpox (accessed on 10 July 2013).
- Pitta-Alavarez, S.; Giulietti, A.M. Influence of chitosan, acetic acid and citric acid on growth and tropane alkaloid production on transformed roots of Brugmansia candida Effect of medium pH and growth phase. In Vitro Cell. Dev. Biol. Plant 1999, 59, 31–38. [Google Scholar]
- Sevón, N.; Hiltunen, R.; Oksman-Caldentey, K.M. Chitosan increases hyoscyamine content in hairy root cultures of Hyoscyamus muticus. Pharm. Pharmacol. Lett. 1992, 2, 96–99. [Google Scholar]
- Merkli, A.; Christen, P.; Kapetanidis, I. Production of diosgenin by hairy root cultures of Trigonella foenum-graecum L. Plant Cell Rep. 1997, 16, 632–636. [Google Scholar] [CrossRef]
- Kim, J.H.; Shin, J.H.; Lee, H.J.; Chung, I.S.; Lee, H.J. Effect of chitosan on indirubin production from suspension culture of Polygonum tinctorium. J. Ferment. Bioeng. 1997, 83, 206–208. [Google Scholar] [CrossRef]
- Tay, L.F.; Khoh, L.K.; Loh, C.S.; Khor, E. Alginate-chitosan coacervation in production of artificial seeds. Biotechnol. Bioeng. 1993, 42, 449–454. [Google Scholar] [CrossRef]
- Kowalski, B.; Jimenez Terry, F.; Herrera, L.; Agramonte Peñalver, D. Application of soluble chitosan in vitro and in the greenhouse to increase yield and seed quality of potato minitubers. Potato Res. 2006, 49, 167–176. [Google Scholar]
- El Ghaouth, A.; Smilanick, J.L.; Wilson, C.L. Enhancement of the performance of Candida saitoana by the addition of glycolchitosan for the control of postharvest decay of apple and citrus fruit. Postharvest Biol. Technol. 2000, 19, 103–110. [Google Scholar] [CrossRef]
- Benhamou, N. Potential of the mycoparasite, Verticillium lecanii, to protect citrus fruit against Penicillium digitatum, the causal agent of green mold: A comparison with the effect of chitosan. Phytopathology 2004, 94, 693–705. [Google Scholar] [CrossRef]
- Du, J.; Gemma, H.; Iwahori, S. Effects of chitosan coating on the storage of peach Japanese pear and kiwifruit. J. Jpn. Soc. Hort. Sci. 1997, 66, 15–22. [Google Scholar] [CrossRef]
- Fornes, F.; Almela, V.; Abad, M.; Agustí, M. Low concentrations of chitosan coating reduce water spot incidence and delay peel pigmentation of clementine mandarin fruit. J. Sci. Food Agric. 2005, 85, 1105–1112. [Google Scholar] [CrossRef]
- El Ghaouth, A.; Arul, J.; Grenier, J.; Asselin, A. Antifungal activity of chitosan on two postharvest pathogens of strawberry fruits. Phytopathology 1992, 82, 398–402. [Google Scholar] [CrossRef]
- Romanazzi, G.; Nigro, F.; Ippolito, A. Short hypobaric treatments potentiate the effect of chitosan in reducing storage decay of sweet cherries. Postharvest Biol. Technol. 2003, 29, 73–80. [Google Scholar] [CrossRef]
- Romanazzi, G.; Mlikota Gabler, F.; Smilanick, J.L. Preharvest chitosan and postharvest UV-C irradiation treatments suppress gray mold of table grapes. Plant Dis. 2006, 90, 445–450. [Google Scholar] [CrossRef]
- Sivakumar, D.; Sultanbawa, Y.; Ranasingh, N.; Wijesundera, R.L.C. Effect of the combined application of chitosan and carbonate salts on the incidence of anthracnose and on the quality of papaya during storage. J. Hortic. Sci. Biotechnol. 2005, 80, 447–452. [Google Scholar]
- Azian, E.; Zaki, A.R.M.; Mohamed, M.T.M.; Kamuruzaman, S. The use of chitosan on vase life of cut chrysanthemum (Dendranthema morifolium Ramat). In Proceedings of APEC Symposium on Quality Management in Postharvest System, Bangkok, Thailand, 3–5 August 2004; p. 403.
- Aranaz, I.; Mengibar, M.; Harris, R.; Panos, I.; Miralles, B.; Acosta, N.; Galed, G.; Heras, A. Functional characterization of chitin and chitosan. Curr. Chem. Biol. 2009, 3, 203–230. [Google Scholar]
- Lonsdale, D. Available treatments for tree wounds: an assessment of their value. Arboricultural J. 1984, 8, 99–107. [Google Scholar] [CrossRef]
- Badawy, M.E.I.; Rabea, E.I.; Rogge, T.M.; Stevens, C.V.; Steurbaut, W.; Höfte, M.; Smagghe, G. Fungicidal and insecticidal activity of O-acyl chitosan derivatives. Polym. Bull. 2005, 54, 279–289. [Google Scholar] [CrossRef]
- Rodríguez-Núñez, J.R.; López-Cervantes, J.; Sánchez-Machado, D.I.; Ramírez-Wong, B.; Torres-Chavez, P.; Cortez-Rocha, M.O. Antimicrobial activity of chitosan-based films against Salmonella typhimurium and Staphylococcus aureus. Int. J. Food Sci. Technol. 2012, 47, 2127–2133. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sharp, R.G. A Review of the Applications of Chitin and Its Derivatives in Agriculture to Modify Plant-Microbial Interactions and Improve Crop Yields. Agronomy 2013, 3, 757-793. https://doi.org/10.3390/agronomy3040757
Sharp RG. A Review of the Applications of Chitin and Its Derivatives in Agriculture to Modify Plant-Microbial Interactions and Improve Crop Yields. Agronomy. 2013; 3(4):757-793. https://doi.org/10.3390/agronomy3040757
Chicago/Turabian StyleSharp, Russell G. 2013. "A Review of the Applications of Chitin and Its Derivatives in Agriculture to Modify Plant-Microbial Interactions and Improve Crop Yields" Agronomy 3, no. 4: 757-793. https://doi.org/10.3390/agronomy3040757
APA StyleSharp, R. G. (2013). A Review of the Applications of Chitin and Its Derivatives in Agriculture to Modify Plant-Microbial Interactions and Improve Crop Yields. Agronomy, 3(4), 757-793. https://doi.org/10.3390/agronomy3040757