Osteopontin and MMP9: Associations with VEGF Expression/Secretion and Angiogenesis in PC3 Prostate Cancer Cells

Abstract
:1. Introduction
2. Experimental Section
2.1. Materials
2.2. Cell Lines and Culture Conditions
2.3. Treatment of PC3 Cells with MEK Inhibitor and Curcumin
2.4. Quantification of VEGF in the Conditioned Medium
2.5. Tube Formation: An in Vitro Matrigel Angiogenesis Assay
2.6. Immunoblotting and Gelatin Zymography Analyses
2.7. Immunohistochemistry
2.8. Statistical Analysis
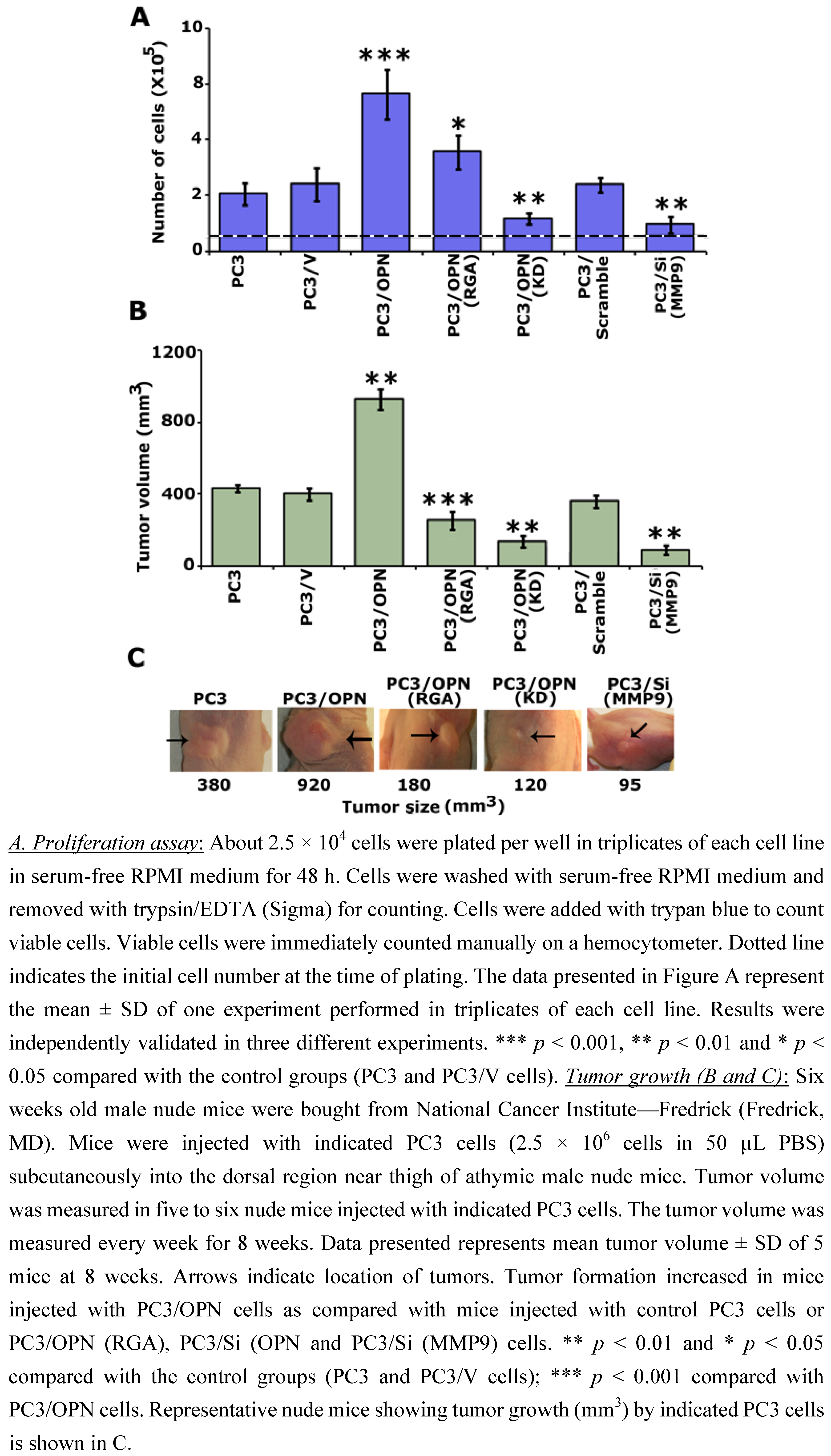
3. Results
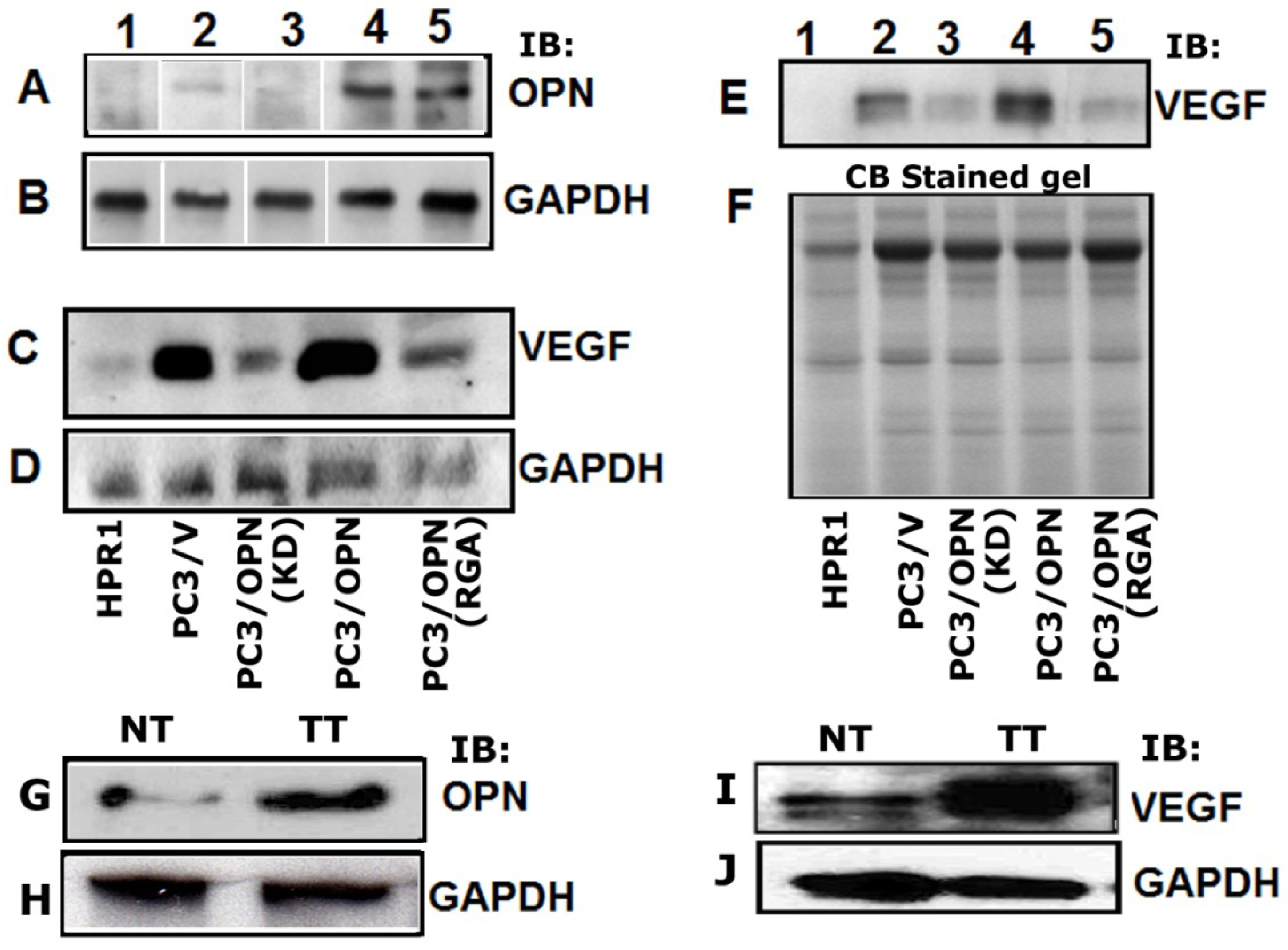
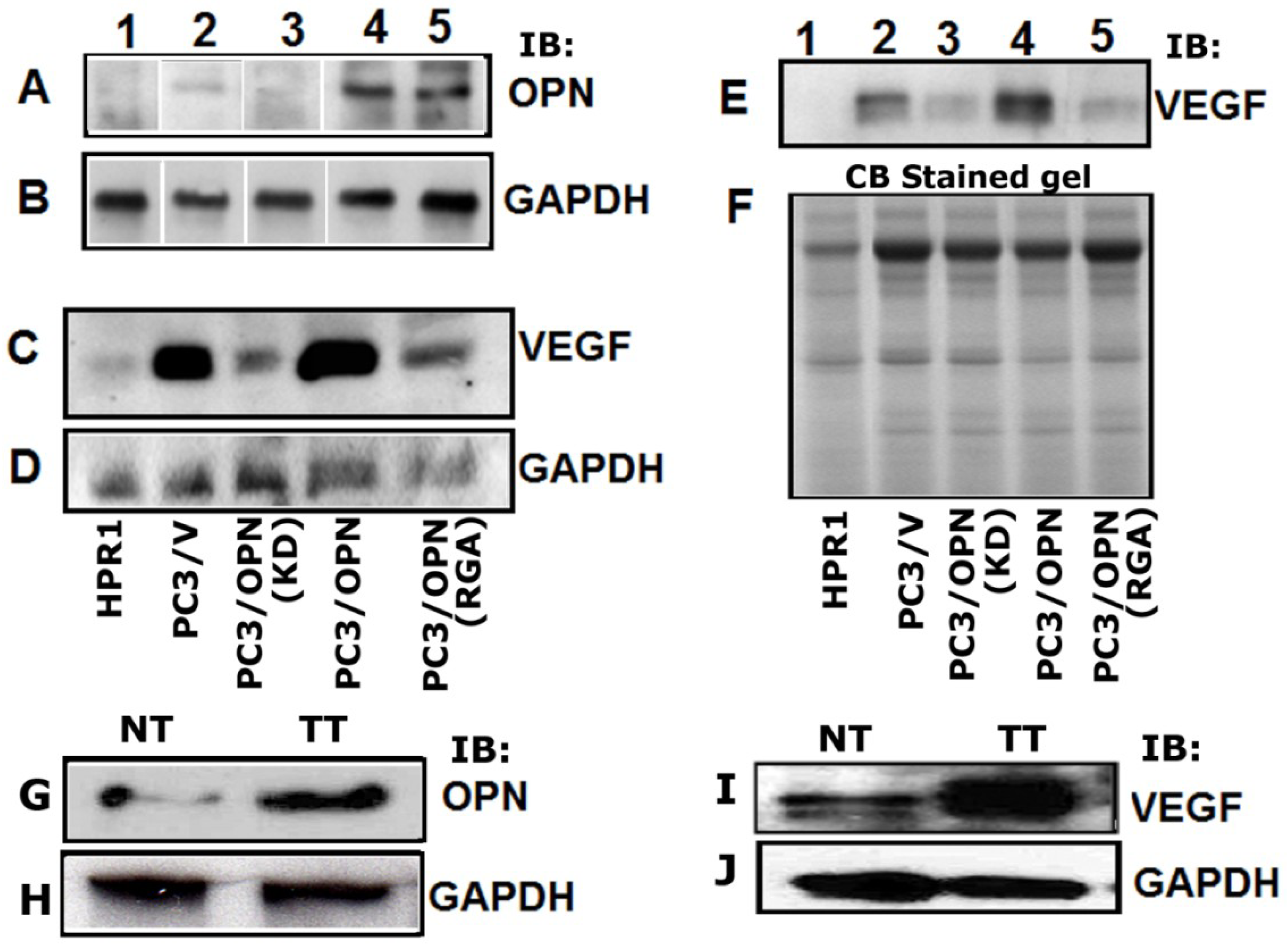
3.1. Regulation of VEGF Expression by Osteopontin
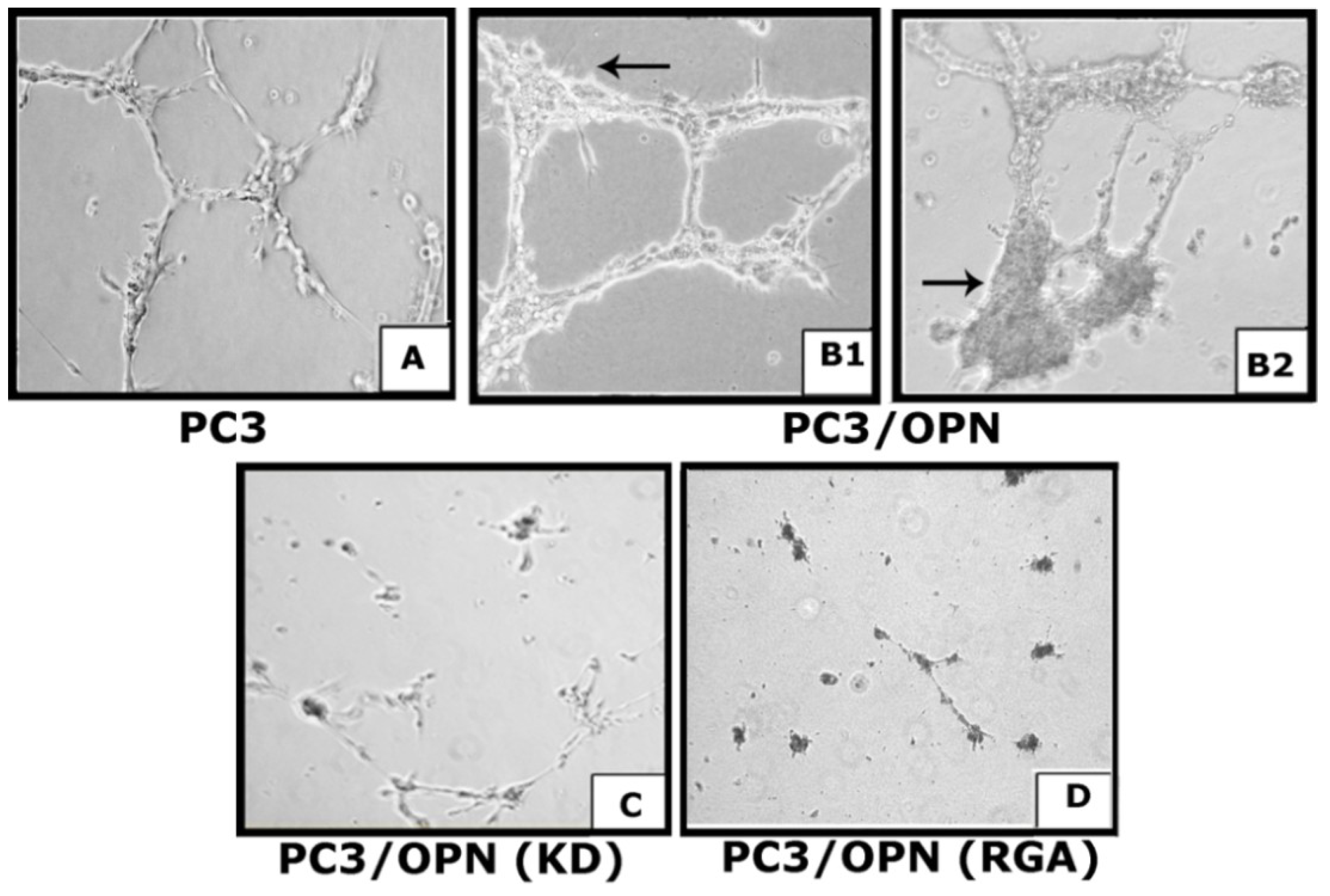
3.2. Osteopontin Deficiency Suppresses VEGF-Induced Angiogenesis in Vitro
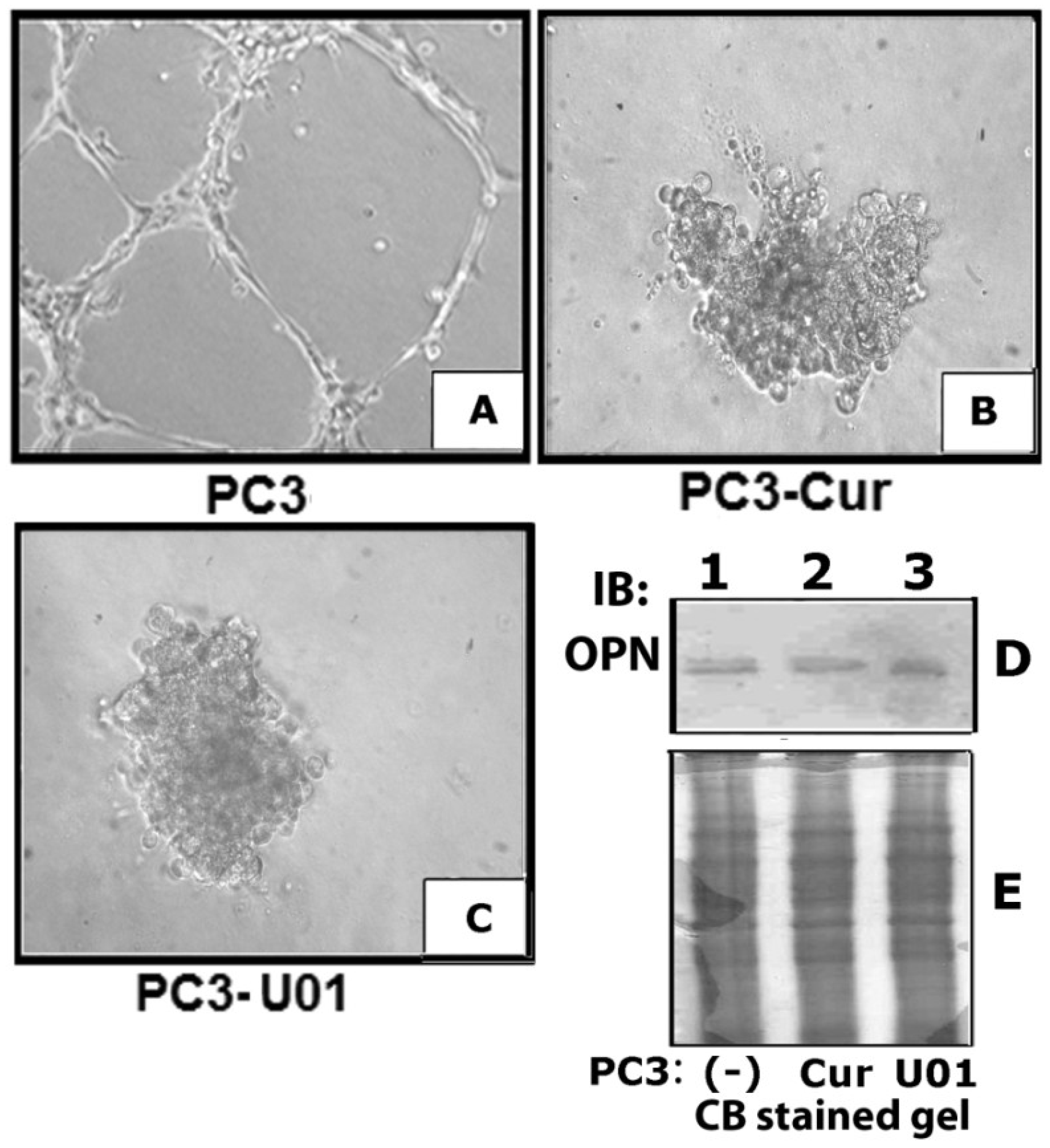
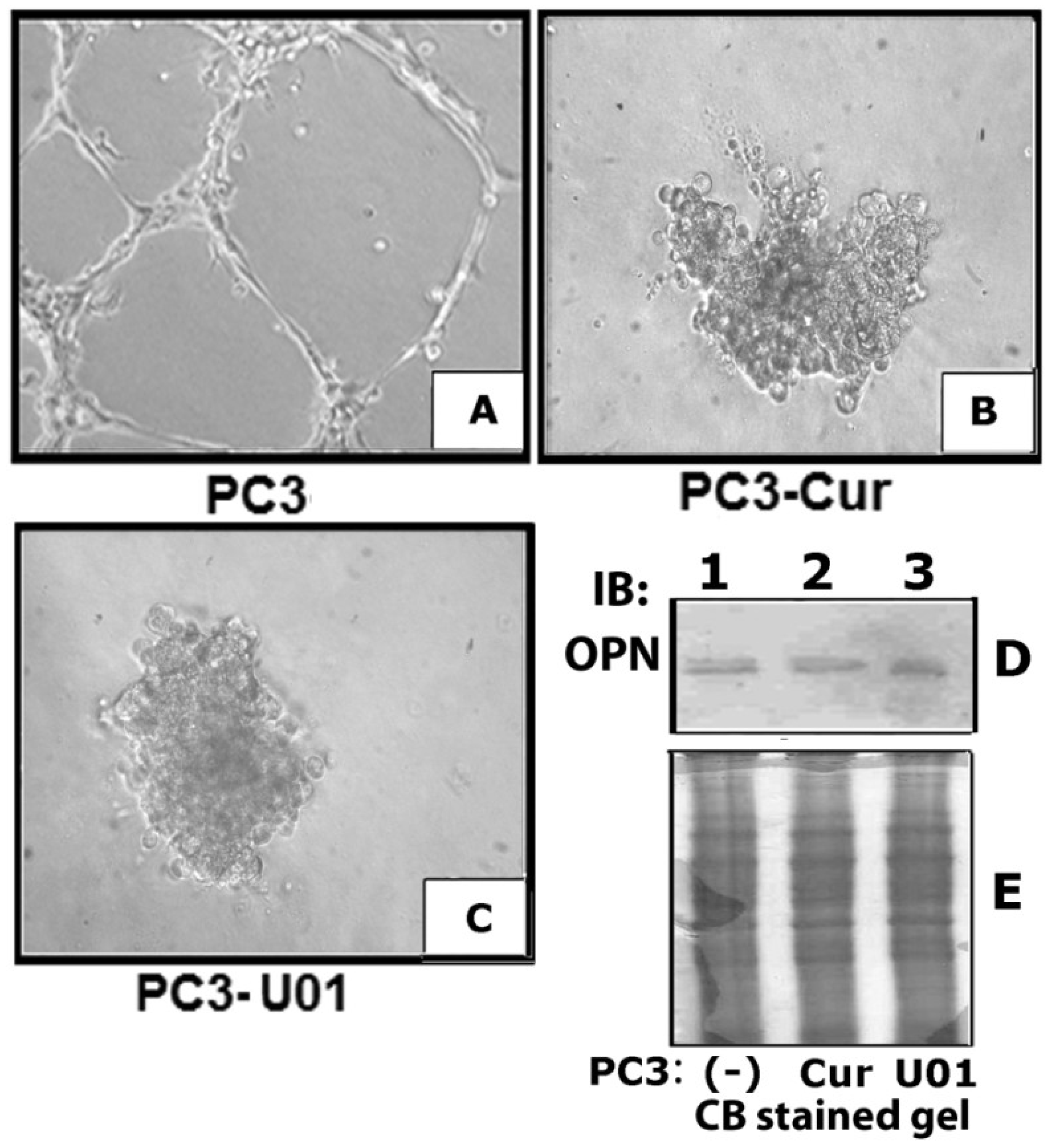
3.3. Curcumin Down-Regulates Vascular Endothelial Growth Factor (VEGF) Expression in PC3 Cells
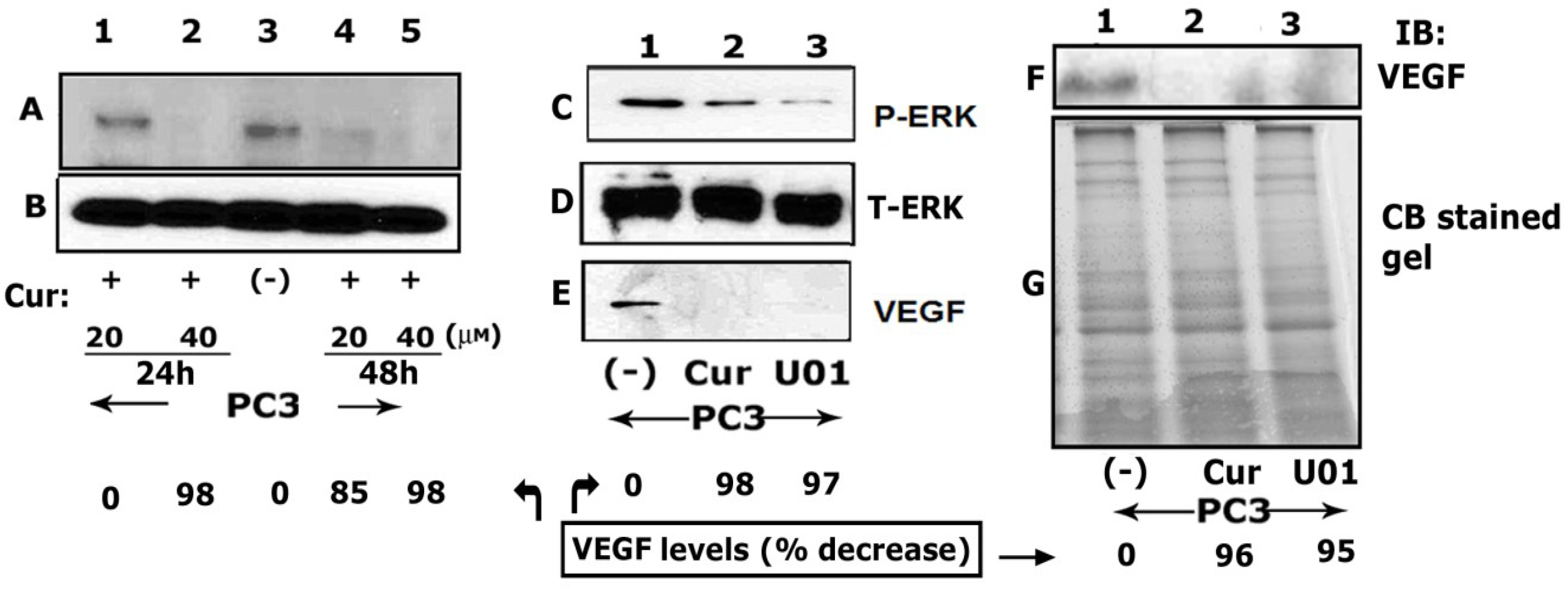
3.4. Curcumin Inhibits ERK Phosphorylation and VEGF Levels in PC3 Cells
3.5. Inhibition of ERK Phosphorylation Represses VEGF Induced Angiogenesis in Vitro
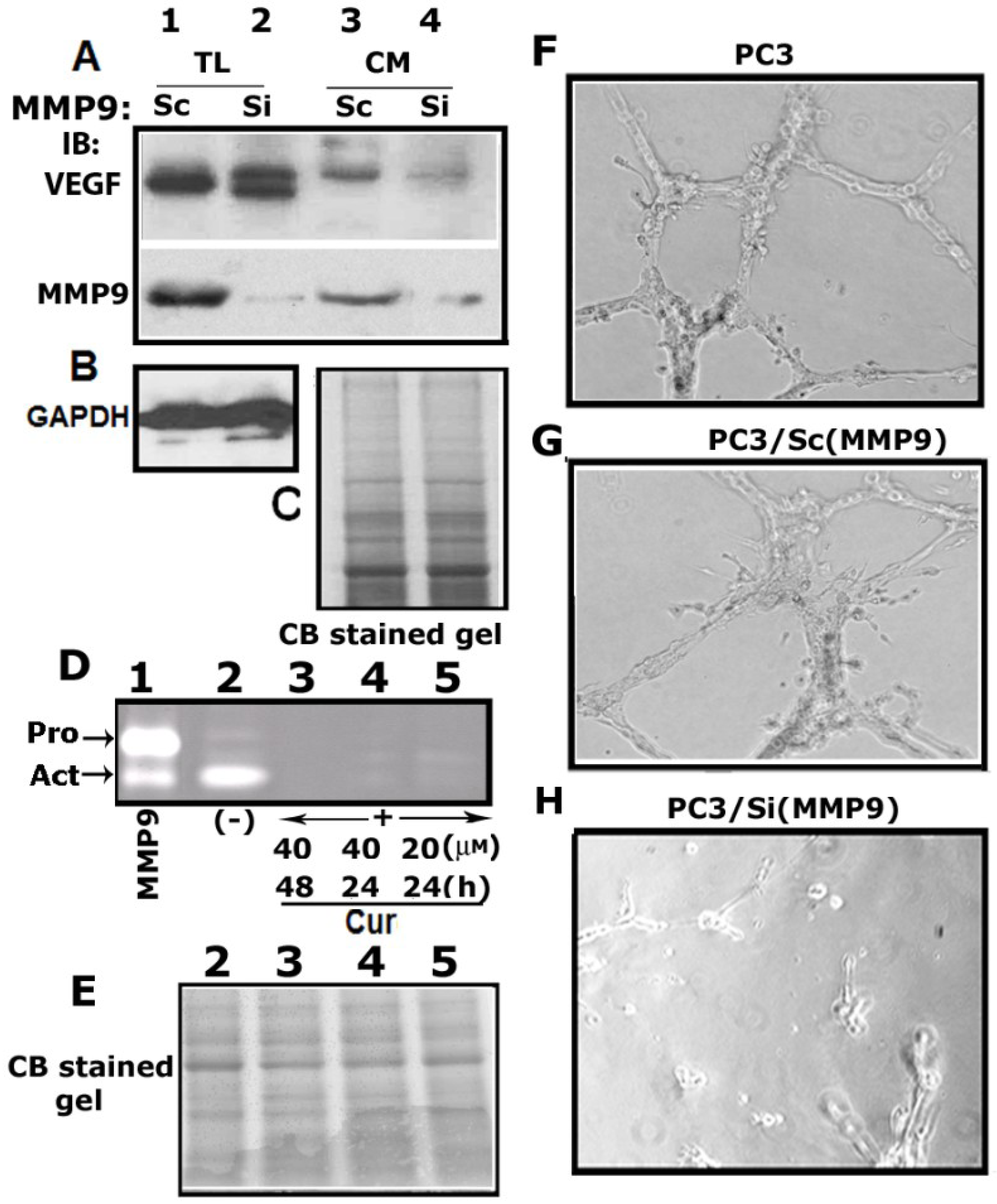
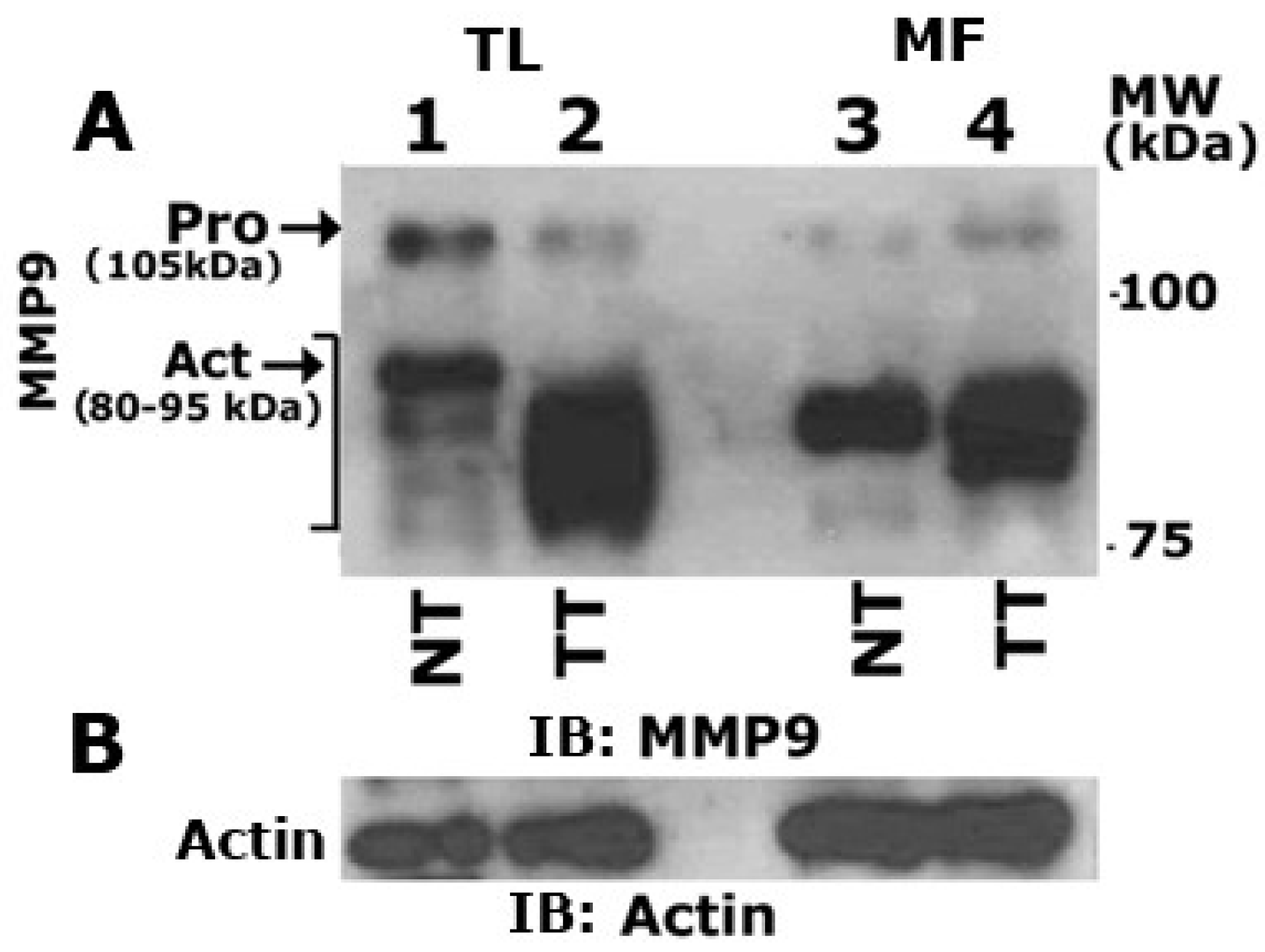
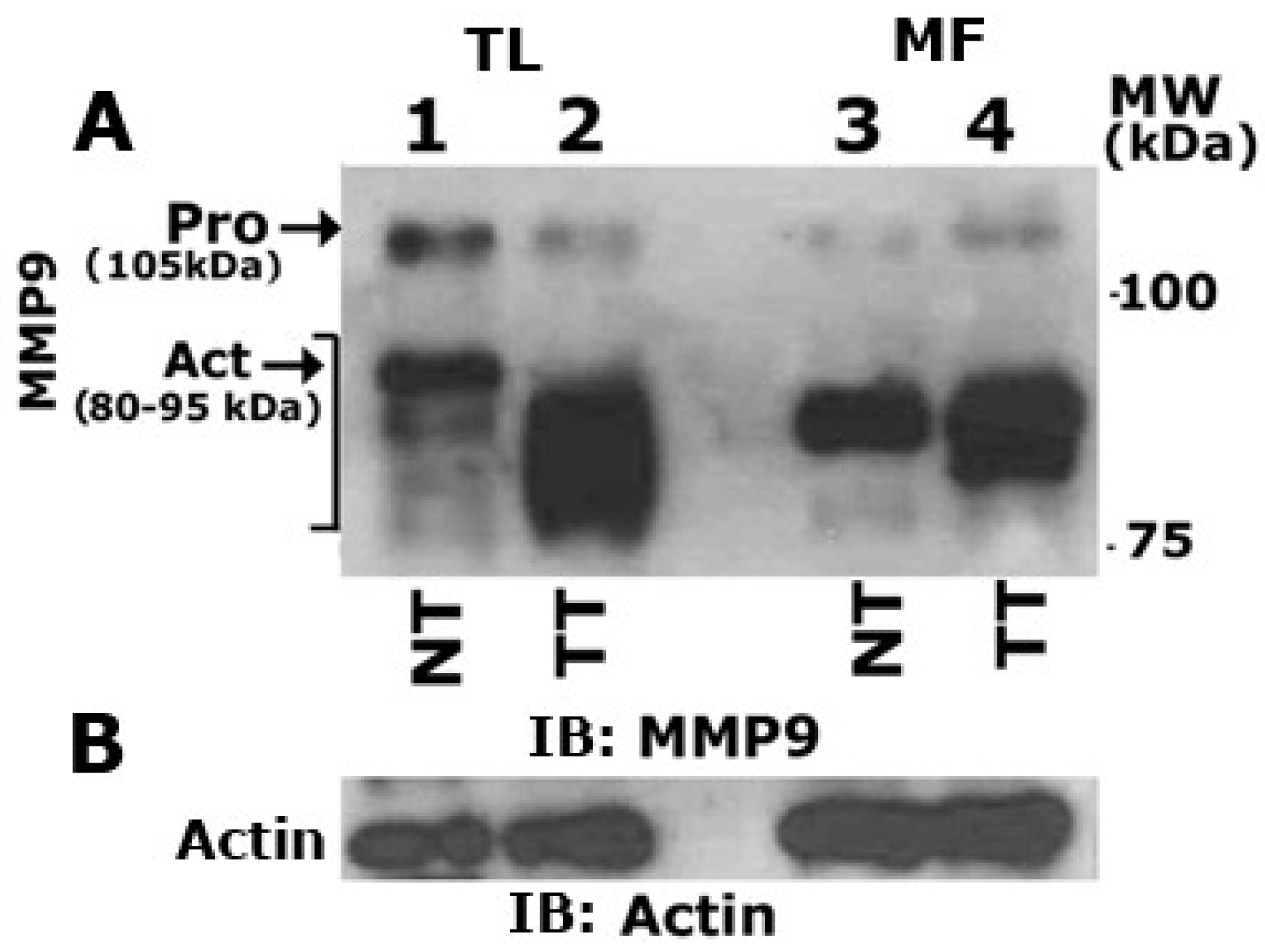
3.6. Matrix Metalloproteinase 9 (MMP9) Plays a Role in the Secretion of VEGF
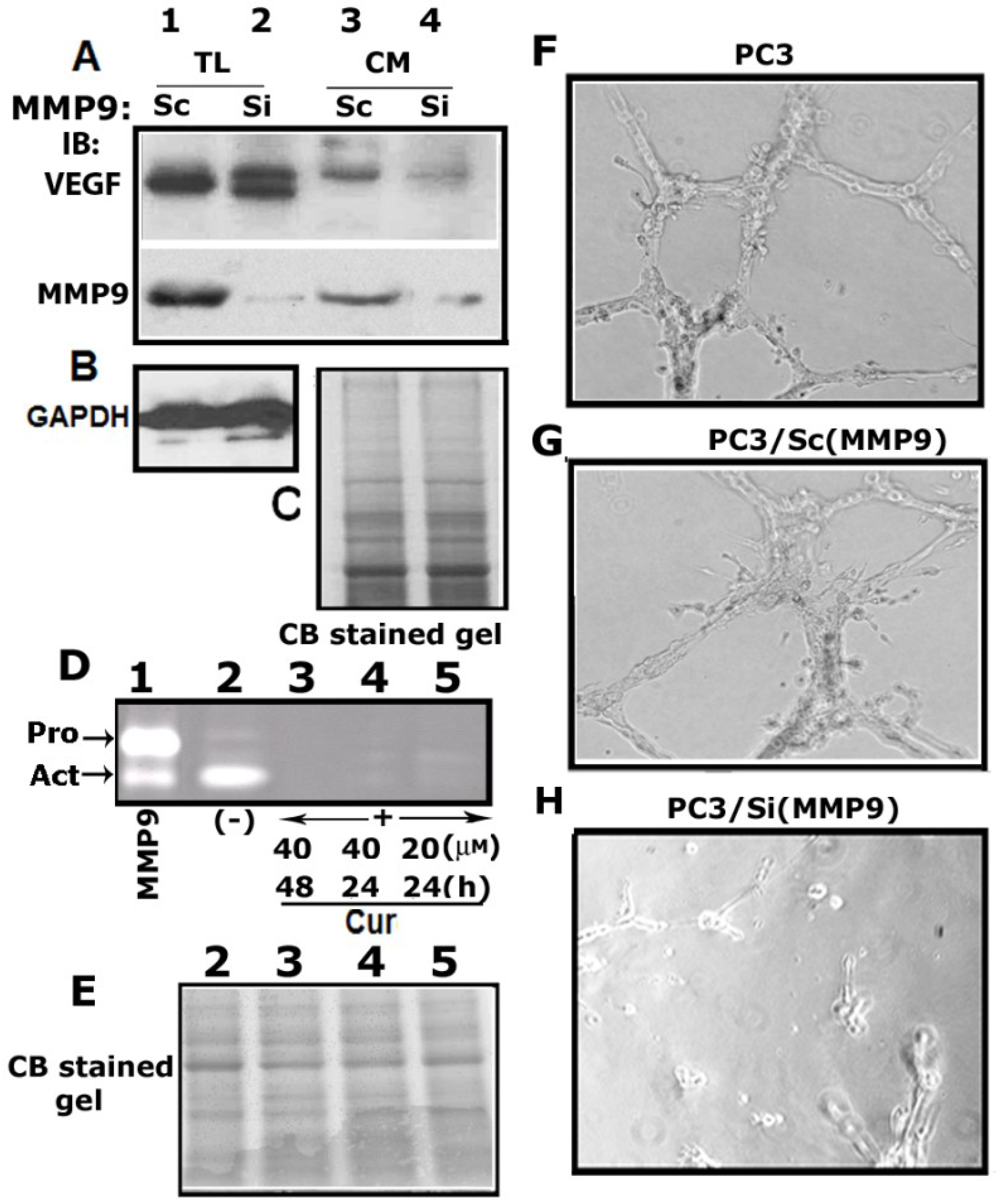
3.7. Curcumin Abrogates MMP9 Activity
3.8. MMP9 Knockdown Reduces VEGF-Induced Angiogenesis in Vitro
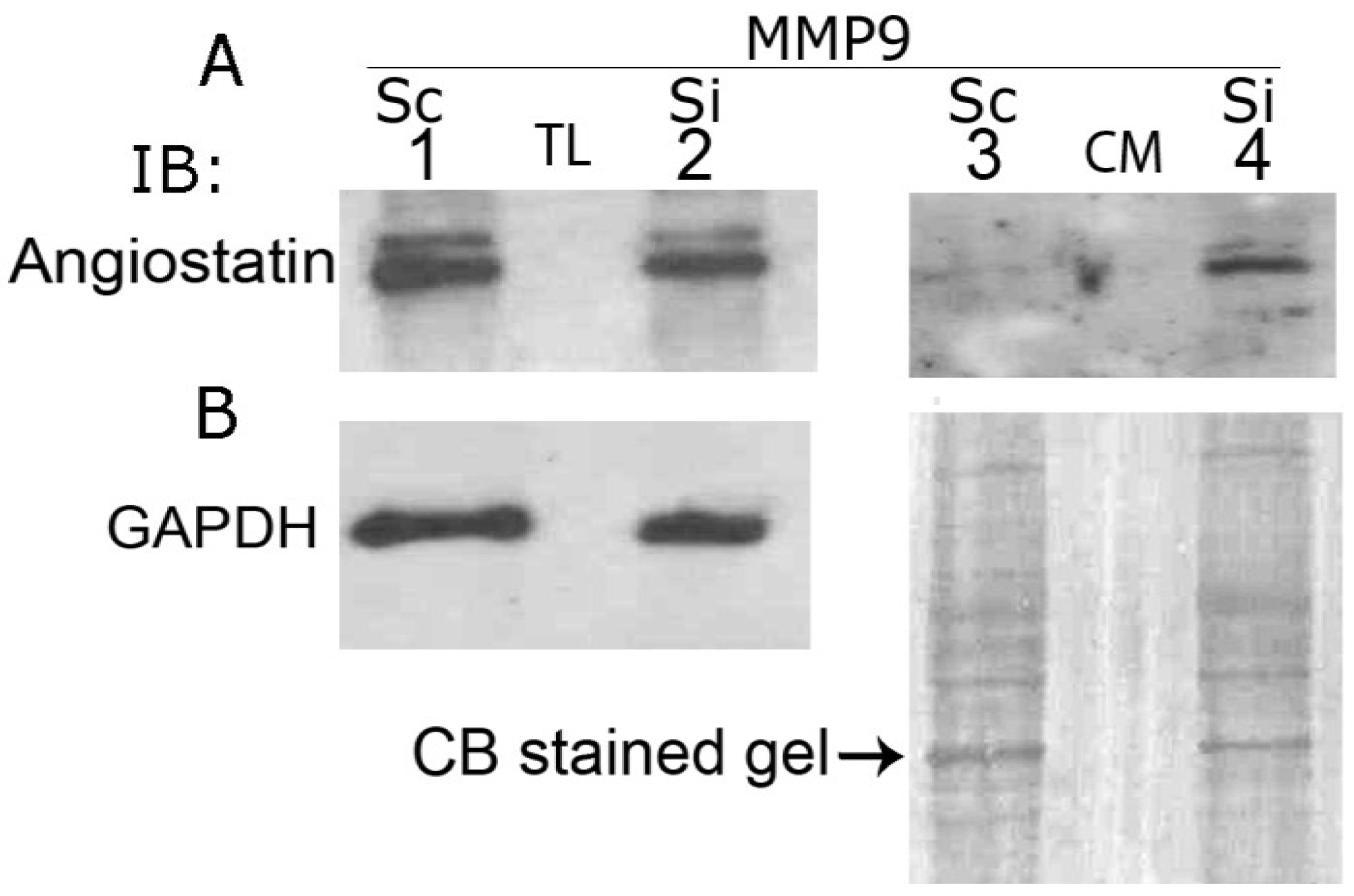
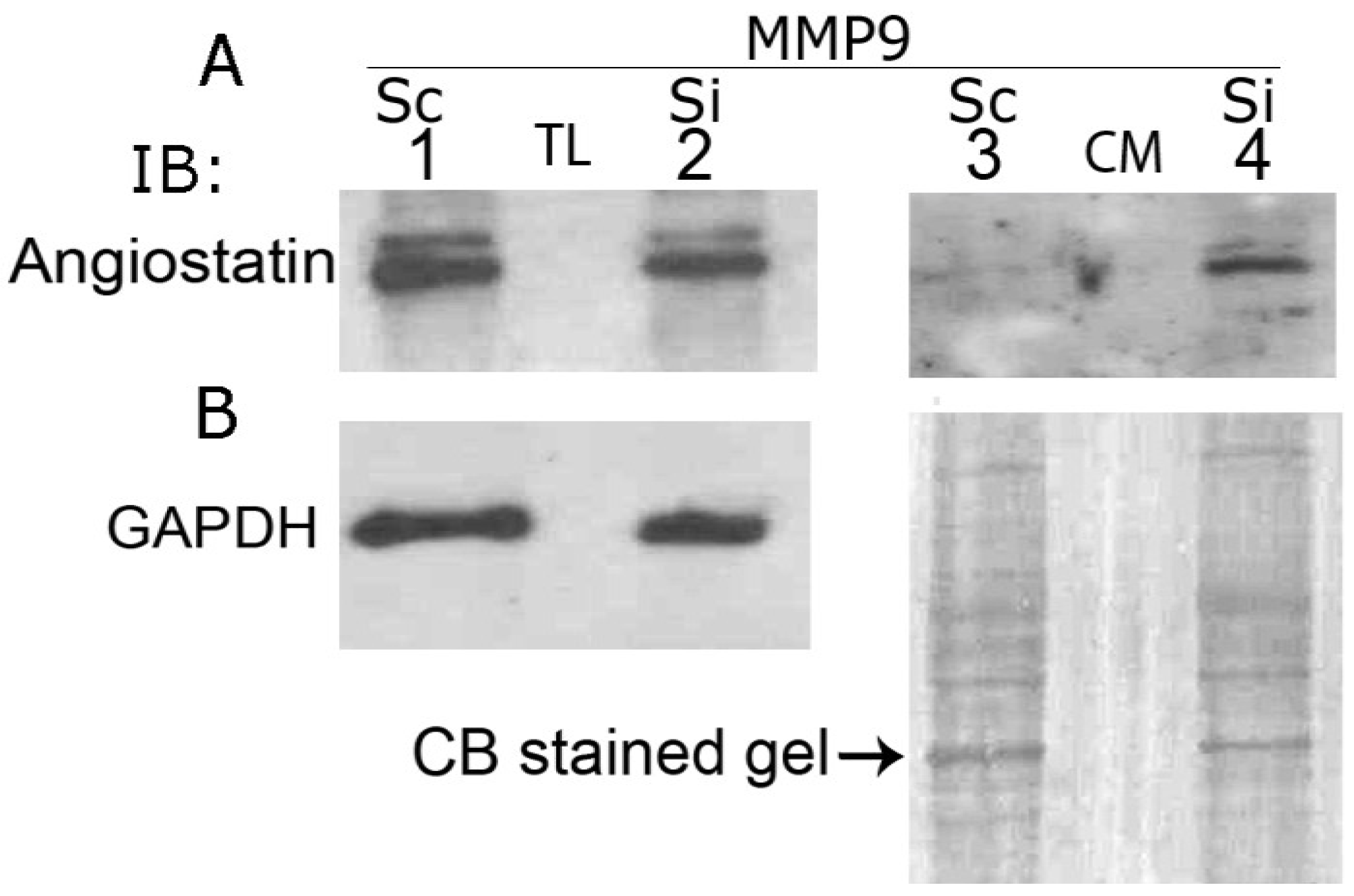
3.9. MMP9 Knockdown Increases Angiostatin Secretion by PC3 Cells
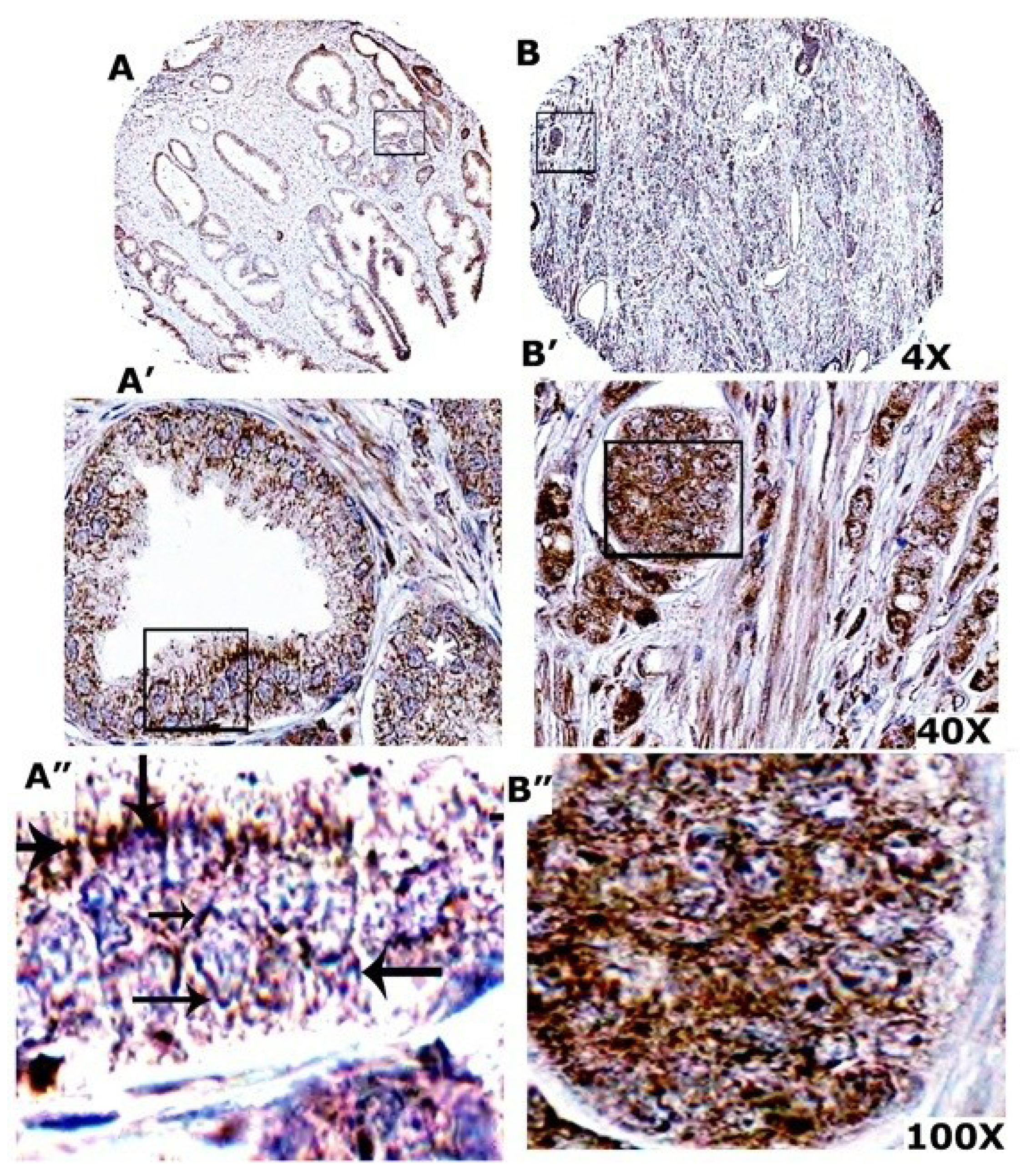
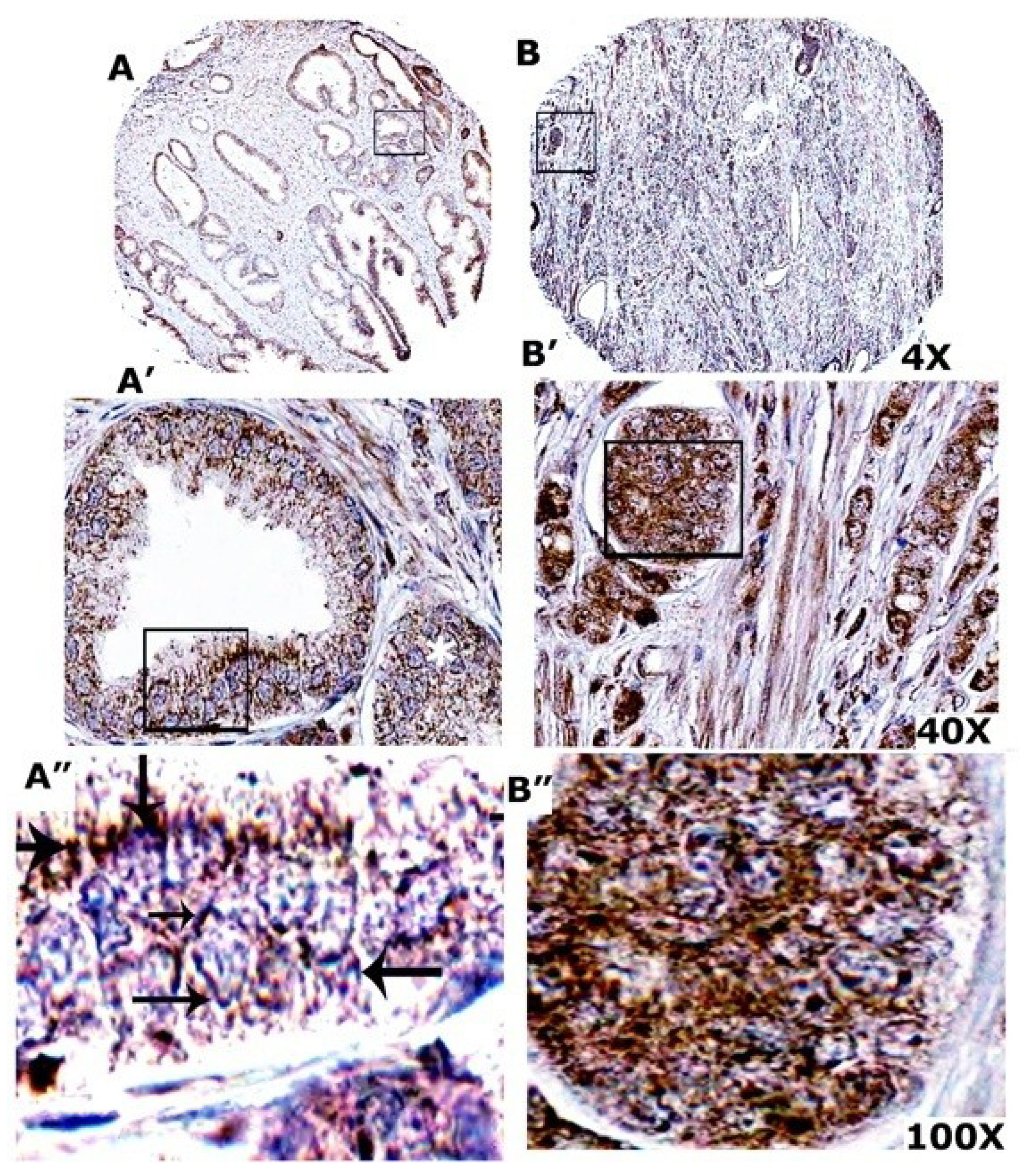
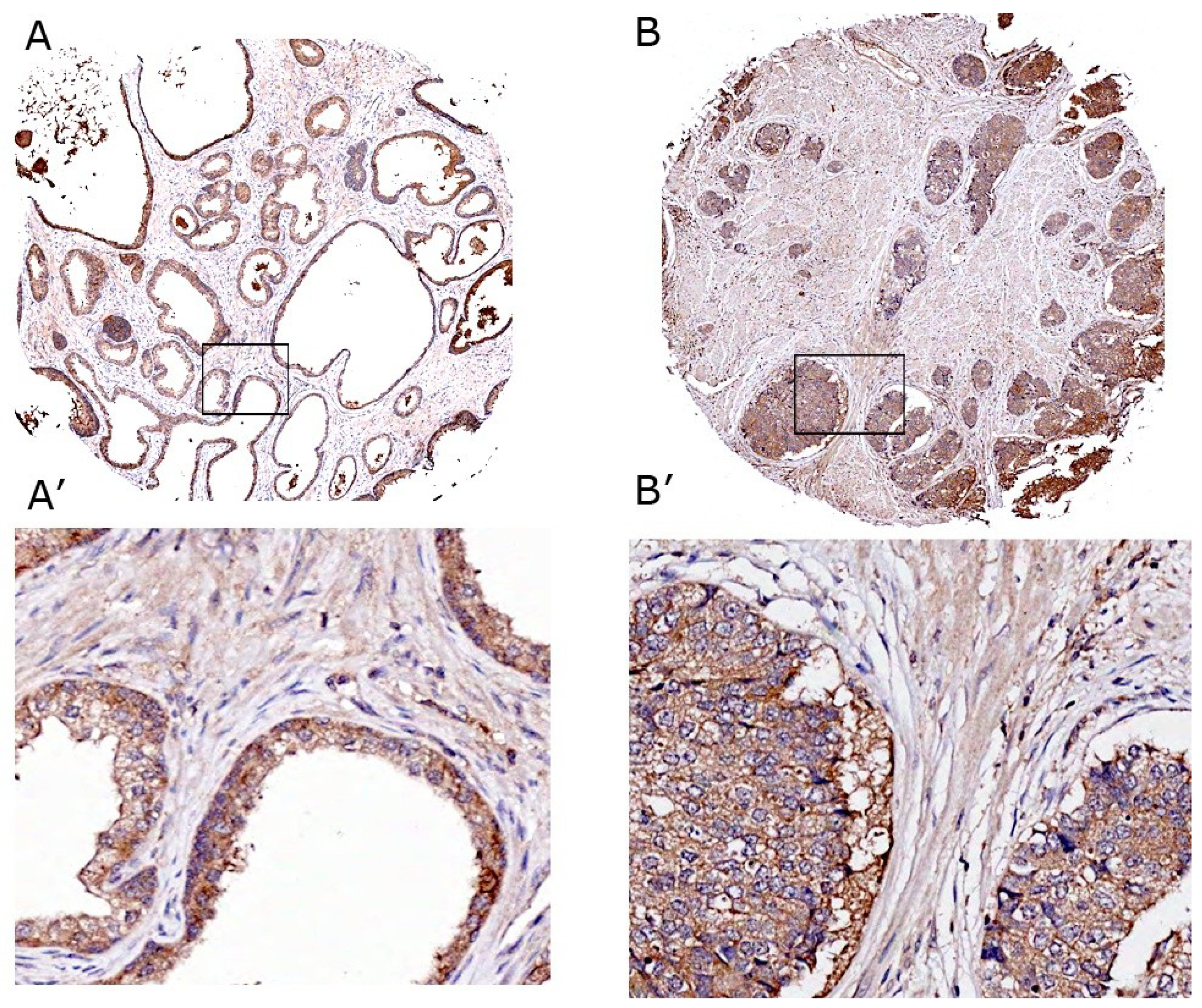
3.10. VEGF Expression Is More in Prostate Adenocarcinoma

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grade | Grade/Stage/# of cores | Cells | VEGF |
|---|---|---|---|
| Normal prostatic epithelial cells and PCa to these cells | (-)/n = 16 | Cancer cells appear normal | Normal cells = 28.0 ± 13% |
| PCa = 33 ± 12% | |||
| Stromal cells < 10% | |||
| Adenocarcinoma (Type: Malignant) | 2/IV/n = 16 | Cells appear slightly different than normal; moderately differentiated | PCa = 68.7 ± 18% ** |
| Stromal cells ~6–8% | |||
| Adenocarcinoma with necrosis (Type:Malignant) | 3/II/n = 8 | Cells appear abnormal; poorly differentiated; stroma is less | PCa = 78 ± 22% *** |
| Stromal cells ~5–8% | |||
| Adenocarcinoma (Type:Malignant) | 3/II/n = 8 | Cells appear abnormal; poorly differentiated; stroma is less. | PCa = 82 ± 28% *** |
| 3/IV/n = 16 | Stromal cells ~4–6% |
4. Discussion
5. Concluding Remarks and Future Directions
Acknowledgements
Abbreviations
| OPN | Osteopontin |
| VEGF | vascular endothelial growth factor |
| MAPK | mitogen-activated protein kinase |
| ERK1/2 | extracellular signal-regulated kinases 1 and 2 |
| MEK1 inhibitor | MAP/ERK kinase 1 inhibitor |
| MMPs | matrix metalloproteinases |
| MMP9 | matrix metalloproteinase 9 |
| integrin αvβ3 | vitronectin receptor |
| RGD | amino acid sequences such as, Arginine(R), Glycine (G) and Aspartic acid (D). It is also known as integrin binding motif |
| RGA | amino acid sequences such as Arginine (R), Glycine (G) and Alanine (A) |
| PC3/OPN | PC3 cells over expressing OPN |
| PC3/OPN (RGA) | PC3 cells over-expressing mutated OPN in integrin binding motif (RGDΔRGA) |
| PC3/OPN (KD) | PC3 cells knockdown of osteopontin |
| PI3-K | phosphatidylinositol 3-kinases |
| HMEC-1 cells | human microvascular endothelial cells-1 |
References
- Folkman, J.; Shing, Y. Angiogenesis. J. Biol. Chem. 1992, 267, 10931–10934. [Google Scholar]
- Millauer, B.; Wizigmann-Voos, S.; Schnurch, H.; Martinez, R.; Moller, N.P.; Risau, W.; Ullrich, A. High affinity VEGF binding and developmental expression suggest Flk-1 as a major regulator of vasculogenesis and angiogenesis. Cell 1993, 72, 835–846. [Google Scholar] [CrossRef]
- Ferrer, F.A.; Miller, L.J.; Andrawis, R.I.; Kurtzman, S.H.; Albertsen, P.C.; Laudone, V.P.; Kreutzer, D.L. Vascular endothelial growth factor (VEGF) expression in human prostate cancer: In situ and in vitro expression of VEGF by human prostate cancer cells. J. Urol. 1997, 157, 2329–2333. [Google Scholar] [CrossRef]
- Wu, T.T.; Wang, J.S.; Jiann, B.P.; Yu, C.C.; Tsai, J.Y.; Lin, J.T.; Huang, J.K. Expression of vascular endothelial growth factor in Taiwanese benign and malignant prostate tissues. J. Chin. Med. Assoc. 2007, 70, 380–384. [Google Scholar] [CrossRef]
- Bruni-Cardoso, A.; Johnson, L.C.; Vessella, R.L.; Peterson, T.E.; Lynch, C.C. Osteoclast-derived matrix metalloproteinase-9 directly affects angiogenesis in the prostate tumor-bone microenvironment. Mol. Cancer Res. 2010, 8, 459–470. [Google Scholar] [CrossRef]
- Ferrara, N.; Gerber, H.P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef]
- Robinson, C.J.; Stringer, S.E. The splice variants of vascular endothelial growth factor (VEGF) and their receptors. J. Cell Sci. 2001, 114, 853–865. [Google Scholar]
- Muhlhauser, J.; Merrill, M.J.; Pili, R.; Maeda, H.; Bacic, M.; Bewig, B.; Passaniti, A.; Edwards, N.A.; Crystal, R.G.; Capogrossi, M.C. VEGF165 expressed by a replication-deficient recombinant adenovirus vector induces angiogenesis in vivo. Circ. Res. 1995, 77, 1077–1086. [Google Scholar] [CrossRef]
- Rennel, E.; Waine, E.; Guan, H.; Schuler, Y.; Leenders, W.; Woolard, J.; Sugiono, M.; Gillatt, D.; Kleinerman, E.; Bates, D.; et al. The endogenous anti-angiogenic VEGF isoform, VEGF165b inhibits human tumour growth in mice. Br. J. Cancer 2008, 98, 1250–1257. [Google Scholar] [CrossRef]
- Chakraborty, G.; Jain, S.; Behera, R.; Ahmed, M.; Sharma, P.; Kumar, V.; Kundu, G.C. The multifaceted roles of osteopontin in cell signaling, tumor progression and angiogenesis. Curr. Mol. Med. 2006, 6, 819–830. [Google Scholar] [CrossRef]
- Huang, Z.; Bao, S.D. Roles of main pro- and anti-angiogenic factors in tumor angiogenesis. World J. Gastroenterol. 2004, 10, 463–470. [Google Scholar]
- Reynolds, A.; Hart, I.; Jones, D.; Salih, M.; Jones, M.; Robinson, S.; Silva, R.; Kostourou, V.; Saunders, G.; Violante, G.; et al. Stimulation of VEGF-mediated angiogenesis and tumor growth by low concentrations of anti-angiogenic integrin inhibitors. In In National Cancer Reseach Institute Conference (www.ncri.org.uk), Birmingham, UK, 5–8 October 2008. Abstract C189.
- Wang, Y.; Yan, W.; Lu, X.; Qian, C.; Zhang, J.; Li, P.; Shi, L.; Zhao, P.; Fu, Z.; Pu, P.; et al. Overexpression of osteopontin induces angiogenesis of endothelial progenitor cells via the avbeta3/PI3K/AKT/eNOS/NO signaling pathway in glioma cells. Eur. J. Cell. Biol. 2011, 90, 642–648. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Kumar, A.; Bharti, A.C. Anticancer potential of curcumin: Preclinical and clinical studies. Anticancer Res. 2003, 23, 363–398. [Google Scholar]
- Mohan, R.; Sivak, J.; Ashton, P.; Russo, L.A.; Pham, B.Q.; Kasahara, N.; Raizman, M.B.; Fini, M.E. Curcuminoids inhibit the angiogenic response stimulated by fibroblast growth factor-2, including expression of matrix metalloproteinase gelatinase B. J. Biol. Chem. 2000, 275, 10405–10412. [Google Scholar]
- Bergers, G.; Brekken, R.; McMahon, G.; Vu, T.H.; Itoh, T.; Tamaki, K.; Tanzawa, K.; Thorpe, P.; Itohara, S.; Werb, Z.; Hanahan, D. Matrix metalloproteinase-9 triggers the angiogenic switch during carcinogenesis. Nat. Cell. Biol. 2000, 2, 737–744. [Google Scholar] [CrossRef]
- Mira, E.; Lacalle, R.A.; Buesa, J.M.; de Buitrago, G.G.; Jimenez-Baranda, S.; Gomez-Mouton, C.; Martínez, A.C.; Mañes, S. Secreted MMP9 promotes angiogenesis more efficiently than constitutive active MMP9 bound to the tumor cell surface. J. Cell Sci. 2004, 117, 1847–1857. [Google Scholar] [CrossRef]
- Desai, B.; Rogers, M.J.; Chellaiah, M.A. Mechanisms of osteopontin and CD44 as metastatic principles in prostate cancer cells. Mol. Cancer 2007, 6, 18. [Google Scholar] [CrossRef]
- Desai, B.; Ma, T.; Zhu, J.; Chellaiah, M.A. Characterization of the expression of variant and standard CD44 in prostate cancer cells: Identification of the possible molecular mechanism of CD44/MMP9 complex formation on the cell surface. J. Cell Biochem. 2009, 108, 272–284. [Google Scholar] [CrossRef]
- Choo, C.K.; Ling, M.T.; Chan, K.W.; Tsao, S.W.; Zheng, Z.; Zhang, D.; Chan, L.C.; Wong, Y.C. Immortalization of human prostate epithelial cells by HPV 16 E6/E7 open reading frames. Prostate 1999, 40, 150–158. [Google Scholar] [CrossRef]
- Chellaiah, M.; Hruska, K.A. Osteopontin stimulates gelsolin associated phosphoinositide levels and PtdIns 3-hydroxyl kinase. Mol. Biol. Cell 1996, 7, 743–753. [Google Scholar]
- Pang, X.; Yi, Z.; Zhang, X.; Sung, B.; Qu, W.; Lian, X.; Aggarwal, B.B.; Liu, M. Acetyl-11-keto-beta-boswellic acid inhibits prostate tumor growth by suppressing vascular endothelial growth factor receptor 2-mediated angiogenesis. Cancer Res. 2009, 69, 5893–5900. [Google Scholar] [CrossRef]
- Gupta, A.; Cao, W.; Chellaiah, M.A. Integrin avb3 and CD44 pathways in metastatic prostate cancer cells support osteoclastogensis via RUNX2/Smad5/RANKL Signaling Axis. Mol. Cancer 2012, 11, 66:1–66:17. [Google Scholar]
- Gattorno, M.; Gregorio, A.; Ferlito, F.; Gerloni, V.; Parafioriti, A.; Felici, E.; Sala, E.; Gambini, C.; Picco, P.; Martini, A. Synovial expression of osteopontin correlates with angiogenesis in juvenile idiopathic arthritis. Rheumatology (Oxford) 2004, 43, 1091–1096. [Google Scholar]
- Desai, B.; Ma, T.; Chellaiah, M.A. Invadopodia and matrix degradation: A new property of prostate cancer cells during migration and invasion. J. Biol. Chem. 2008, 283, 13856–13866. [Google Scholar] [CrossRef]
- Thalmann, G.N.; Sikes, R.A.; Devoll, R.E.; Kiefer, J.A.; Markwalder, R.; Klima, I.; Farach-Carson, C.M.; Studer, U.E.; Chung, L.W. Osteopontin: Possible role in prostate cancer progression. Clin. Cancer Res. 1999, 5, 2271–2277. [Google Scholar]
- Khodavirdi, A.C.; Song, Z.; Yang, S.; Zhong, C.; Wang, S.; Wu, H.; Pritchard, C.; Nelson, P.S.; Roy-Burman, P. Increased expression of osteopontin contributes to the progression of prostate cancer. Cancer Res. 2006, 66, 883–888. [Google Scholar] [CrossRef]
- Woollard, D.J.; Opeskin, K.; Coso, S.; Wu, D.; Baldwin, M.E.; Williams, E.D. Differential expression of VEGF ligands and receptors in prostate cancer. Prostate 2013, 73, 563–572. [Google Scholar] [CrossRef]
- Jackson, M.W.; Bentel, J.M.; Tilley, W.D. Vascular endothelial growth factor (VEGF) expression in prostate cancer and benign prostatic hyperplasia. J. Urol. 1997, 157, 2323–2328. [Google Scholar] [CrossRef]
- Wilken, R.; Veena, M.S.; Wang, M.B.; Srivatsan, E.S. Curcumin: A review of anti-cancer properties and therapeutic activity in head and neck squamous cell carcinoma. Mol. Cancer 2011, 10, 12. [Google Scholar] [CrossRef]
- Lin, Y.G.; Kunnumakkara, A.B.; Nair, A.; Merritt, W.M.; Han, L.Y.; Rmaiz-Pena, G.N.; Kamat, A.A.; Spannuth, W.A.; Gershenson, D.M.; Lutgendorf, S.K.; et al. Curcumin inhibits tumor growth and angiogenesis in ovarian carcinoma by targeting the nuclear factor-kappaB pathway. Clin. Cancer Res. 2007, 13, 3423–3430. [Google Scholar] [CrossRef]
- Chakraborty, G.; Jain, S.; Kale, S.; Raja, R.; Kumar, S.; Mishra, R.; Kundu, G.C. Curcumin suppresses breast tumor angiogenesis by abrogating osteopontin-induced VEGF expression. Mol. Med. Report 2008, 1, 641–646. [Google Scholar]
- Plantner, J.J.; Jiang, C.; Smine, A. Increase in interphotoreceptor matrix gelatinase A (MMP-2) associated with age-related macular degeneration. Exp. Eye Res. 1998, 67, 637–645. [Google Scholar] [CrossRef]
- Kumta, S.M.; Huang, L.; Cheng, Y.Y.; Chow, L.T.; Lee, K.M.; Zheng, M.H. Expression of VEGF and MMP-9 in giant cell tumor of bone and other osteolytic lesions. Life Sci. 2003, 73, 1427–1436. [Google Scholar] [CrossRef]
- Hong, J.H.; Ahn, K.S.; Bae, E.; Jeon, S.S.; Choi, H.Y. The effects of curcumin on the invasiveness of prostate cancer in vitro and in vivo. Prostate Cancer Prostatic Dis. 2006, 9, 147–152. [Google Scholar] [CrossRef]
- Swarnakar, S.; Paul, S. Curcumin arrests endometriosis by downregulation of matrix metalloproteinase-9 activity. Indian J. Biochem. Biophys. 2009, 46, 59–65. [Google Scholar]
- Patterson, B.C.; Sang, Q.A. Angiostatin-converting enzyme activities of human matrilysin (MMP-7) and gelatinase B/type IV collagenase (MMP-9). J. Biol. Chem. 1997, 272, 28823–28825. [Google Scholar] [CrossRef]
- Yao, J.L.; Ryan, C.K.; Francis, C.W.; Kohli, M.; Taubman, M.B.; Khorana, A.A. Tissue factor and VEGF expression in prostate carcinoma: A tissue microarray study. Cancer Invest. 2009, 27, 430–434. [Google Scholar] [CrossRef]
- Yeatman, T.J.; Chambers, A.F. Osteopontin and colon cancer progression. Clin. Exp. Metastasis 2003, 20, 85–90. [Google Scholar] [CrossRef]
- Standal, T.; Borset, M.; Sundan, A. Role of osteopontin in adhesion, migration, cell survival and bone remodeling. Exp. Oncol. 2004, 26, 179–184. [Google Scholar]
- Giuliani, N.; Lunghi, P.; Morandi, F.; Colla, S.; Bonomini, S.; Hojden, M.; Rizzoli, V.; Bonati, A. Downmodulation of ERK protein kinase activity inhibits VEGF secretion by human myeloma cells and myeloma-induced angiogenesis. Leukemia 2004, 18, 628–635. [Google Scholar] [CrossRef]
- Menu, E.; Kooijman, R.; van Valckenborgh, E.; Asosingh, K.; Bakkus, M.; van Camp, B.; Vanderkerken, K. Specific roles for the PI3K and the MEK-ERK pathway in IGF-1-stimulated chemotaxis, VEGF secretion and proliferation of multiple myeloma cells: Study in the 5T33MM model. Br. J. Cancer 2004, 90, 1076–1083. [Google Scholar] [CrossRef]
- Anand, P.; Sundaram, C.; Jhurani, S.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin and cancer: An “old-age” disease with an “age-old” solution. Cancer Lett. 2008, 267, 133–164. [Google Scholar] [CrossRef]
- Ayala, G.; Liu, C.; Nicosia, R.; Horowitz, S.; Lackman, R. Microvasculature and VEGF expression in cartilaginous tumors. Hum. Pathol. 2000, 31, 341–346. [Google Scholar] [CrossRef]
- Verheul, H.M.; Pinedo, H.M. Tumor Growth: A Putative Role for Platelets? Oncologist 1998, 3, 0–0i. [Google Scholar]
- Dai, J.; Peng, L.; Fan, K.; Wang, H.; Wei, R.; Ji, G.; Cai, J.; Lu, B.; Li, B.; Zhang, D.; et al. Osteopontin induces angiogenesis through activation of PI3K/AKT and ERK1/2 in endothelial cells. Oncogene 2009, 28, 3412–3422. [Google Scholar] [CrossRef]
- Isowa, S.; Shimo, T.; Ibaragi, S.; Kurio, N.; Okui, T.; Matsubara, K.; Hassan, N.M.; Kishimoto, K.; Sasaki, A. PTHrP regulates angiogenesis and bone resorption via VEGF expression. Anticancer Res. 2010, 30, 2755–2767. [Google Scholar]
- Robertson, B.W.; Bonsal, L.; Chellaiah, M.A. Regulation of Erk1/2 activation by osteopontin in PC3 human prostate cancer cells. Mol. Cancer 2010, 9, 260. [Google Scholar] [CrossRef]
- Binion, D.G.; Otterson, M.F.; Rafiee, P. Curcumin inhibits VEGF-mediated angiogenesis in human intestinal microvascular endothelial cells through COX-2 and MAPK inhibition. Gut 2008, 57, 1509–1517. [Google Scholar] [CrossRef]
- Swarnakar, S.; Ganguly, K.; Kundu, P.; Banerjee, A.; Maity, P.; Sharma, A.V. Curcumin regulates expression and activity of matrix metalloproteinases 9 and 2 during prevention and healing of indomethacin-induced gastric ulcer. J. Biol. Chem. 2005, 280, 9409–9415. [Google Scholar]
- Philip, S.; Bulbule, A.; Kundu, G.C. Matrix metalloproteinase-2: Mechanism and regulation of NF-kappaB-mediated activation and its role in cell motility and ECM-invasion. Glycoconj. J. 2004, 21, 429–441. [Google Scholar] [CrossRef]
- Pozzi, A.; Moberg, P.E.; Miles, L.A.; Wagner, S.; Soloway, P.; Gardner, H.A. Elevated matrix metalloprotease and angiostatin levels in integrin alpha 1 knockout mice cause reduced tumor vascularization. Proc. Natl. Acad. Sci. USA 2000, 97, 2202–2207. [Google Scholar]
- Binda, M.M.; Matar, P.; Gonzalez, A.D.; Rozados, V.R.; Gervasoni, S.I.; Scharovsky, O.G.; Bonfil, R.D. Differential production of angiostatin by concomitant antitumoral resistance-inducing cancer cells. Int. J. Cancer 2002, 100, 14–21. [Google Scholar] [CrossRef]
- Zetter, B.R. Angiogenesis and tumor metastasis. Annu. Rev. Med. 1998, 49, 407–424. [Google Scholar] [CrossRef]
- Pozzi, A.; LeVine, W.F.; Gardner, H.A. Low plasma levels of matrix metalloproteinase 9 permit increased tumor angiogenesis. Oncogene 2002, 21, 272–281. [Google Scholar] [CrossRef]
Supplemental Materials


© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gupta, A.; Zhou, C.Q.; Chellaiah, M.A. Osteopontin and MMP9: Associations with VEGF Expression/Secretion and Angiogenesis in PC3 Prostate Cancer Cells. Cancers 2013, 5, 617-638. https://doi.org/10.3390/cancers5020617
Gupta A, Zhou CQ, Chellaiah MA. Osteopontin and MMP9: Associations with VEGF Expression/Secretion and Angiogenesis in PC3 Prostate Cancer Cells. Cancers. 2013; 5(2):617-638. https://doi.org/10.3390/cancers5020617
Chicago/Turabian StyleGupta, Aditi, Cindy Q. Zhou, and Meenakshi A. Chellaiah. 2013. "Osteopontin and MMP9: Associations with VEGF Expression/Secretion and Angiogenesis in PC3 Prostate Cancer Cells" Cancers 5, no. 2: 617-638. https://doi.org/10.3390/cancers5020617