Responsiveness of Urinary and Plasma Alkylresorcinol Metabolites to Rye Intake in Finnish Women

Abstract
:1. Introduction
2. Results and Discussion

{kind=link}
| Variable | Low rye intake (n = 20) | Medium rye intake (n = 20) | High rye intake (n = 20) | p-values |
|---|---|---|---|---|
| Age (years) | 47 ± 13 | 45 ± 15 | 49 ± 15 | 0.598 |
| Weight (kg) | 60 ± 11 | 61 ± 8 | 63 ± 6 | 0.648 |
| Body mass index (kg/m²) | 22 ± 3 | 23 ± 3 | 23 ± 3 | 0.549 |
| Age at menopause (years) | 50 ± 3 | 50 ± 1 | 49 ± 4 | 0.768 |
| Age at menarche (years) | 13 ± 1 | 13 ± 1 | 13 ± 1 | 0.576 |
| Smoker (%) | 25 | 5 | 10 | 0.153* |
| Physically active (%) | 55 | 50 | 50 | 0.935* |
| Age at first pregnancy (years) | 26 ± 3 | 27 ± 5 | 25 ± 5 | 0.521 |
| Number of children | 1.9 ± 1.5 | 1.1 ± 1.3 | 1.5 ± 1.5 | 0.183 |
| P-DHBA (nmol/L)P-DHPPA (nmol/L) | 85 ± 5676 ± 37 | 97 ± 6989 ± 53 | 102 ± 45110 ± 43∆† | 0.333**0.043** |
| U-DHBA (μmol/24h) | 21 ± 8 | 26 ± 12 | 32 ± 9∆† | 0.005** |
| U-DHPPA (μmol/24h) | 32 ± 15 | 40 ± 26 | 48 ± 13∆† | 0.001** |
| Sum P-AR metabolites (nmol/L) | 157 ± 87 | 187 ± 121 | 210 ± 87 | 0.185 |
| Sum U-AR metabolites (μmol/24h) | 52 ± 23 | 66 ± 38 | 79 ± 22∆† | 0.003** |
| Rye intake (g/d) | 23 ± 9† | 44 ± 4∆ | 68 ± 18∆ | 0.000 |
| Wheat intake (g/d) | 79 ± 30 | 79 ± 20 | 71 ± 36 | 0.615 |
| Cereal fiber intake (g/d) | 7.8 ± 2.3† | 9.4 ± 1.6∆ | 11.9 ± 2.6∆† | 0.000 |
| Total fiber intake (g/d) | 16 ± 7 | 19 ± 5 | 20 ± 5 | 0.156 |
| Total kilocalorie intake (kcal/d) | 1774 ± 281 | 1866 ± 285 | 1832 ± 410 | 0.676 |
| Total fat intake (g/d) | 76 ± 15 | 76 ± 15 | 72 ± 18 | 0.682 |
| SFA (g/d) | 36 ± 11 | 39 ± 9 | 38 ± 11 | 0.715 |
| MUFA (g/d) | 25 ± 5 | 24 ± 6 | 22 ± 6 | 0.293 |
| PUFA (g/d) | 10 ± 3 | 10 ± 4 | 9 ± 4 | 0.825 |
| Rye intake (g/d) with covariables | Rye intake (g/d) without covariables | |
|---|---|---|
| P-DHBA (nmol/L) | 0.299* | 0.321* |
| P-DHPPA (nmol/L) | 0.397** | 0.385** |
| P-DHBA+DHPPA (nmol/L) | 0.215 | 0.334* |
| U-DHBA (μmol/24h) | 0.524*** | 0.516*** |
| U-DHPPA (μmol/24h) | 0.438*** | 0.444*** |
| U-DHBA+DHPPA (μmol/24h) | 0.365** | 0.476*** |
| Rye intake groups | P-DHBA (nmol/L) | P-DHPPA (nmol/L) | P-DHBA/DHPPA (ratio) | U-DHBA (μmol/24h) | U-DHPPA (μmol/24h) | U-DHBA/DHPPA (ratio) |
|---|---|---|---|---|---|---|
| Low (reference) | 1 | 1 | 1 | 1 | 1 | 1 |
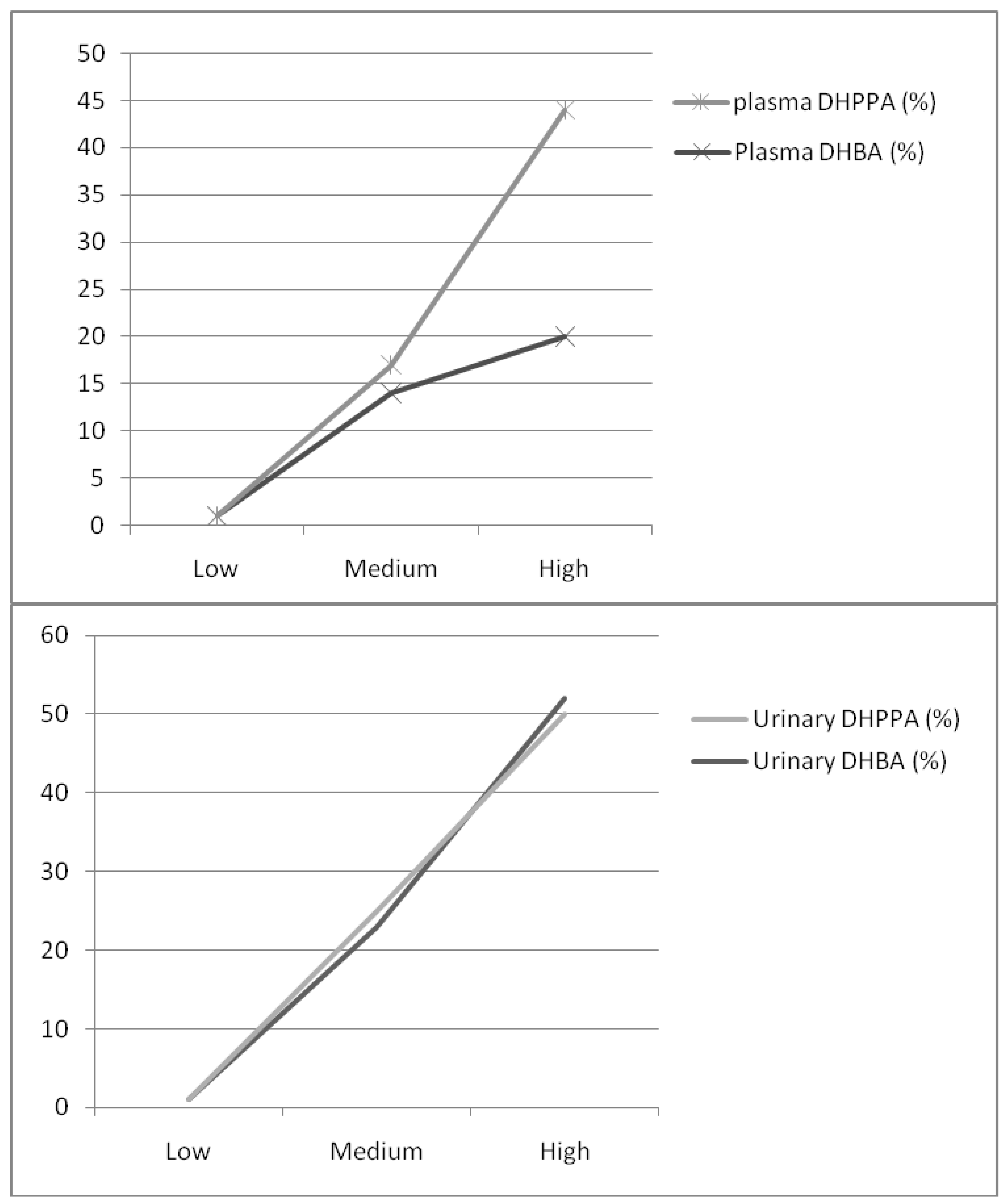
| Medium (+21 g/d) | +14% | +17% | +18% | 23% | 25% | +1% |
| High group (+45 g/d) | +20% | +44%∆† | +25% | 52%∆† | 50%∆† | −6% |

3. Experimental
3.1. Subjects
3.2. Data Collection
3.3. Dietary Intake
3.4. Urinary Alkylresorcinol Metabolites
3.5. Plasma Alkylresorcinol Metabolites
3.6. Statistical Analysis
4. Conclusions
Acknowledgements
References
- de Munter, J.S.; Hu, F.B.; Spiegelman, D.; Franz, M.; van Dam, R.M. Whole grain, bran, and germ intake and risk of type 2 diabetes: a prospective cohort study and systematic review. PLoS Med. 2007, 4, 261. [Google Scholar] [CrossRef]
- Jensen, M.K.; Koh-Banerjee, P.; Hu, F.B.; Franz, M.; Sampson, L.; Gronbaek, M.; Rimm, E.B. Intakes of whole grains, bran, and germ and the risk of coronary heart disease in men. Am. J. Clin. Nutr. 2004, 80, 1492–1499. [Google Scholar]
- Slavin, J. Why whole grains are protective: biological mechanisms. Proc. Nutr. Soc. 2003, 62, 129–134. [Google Scholar] [CrossRef]
- Kaaks, R.; Ferrari, P.; Ciampi, A.; Plummer, M.; Riboli, E. Uses and limitations of statistical accounting for random error correlations, in the validation of dietary questionnaire assessments. Public Health Nutr. 2002, 5, 969–976. [Google Scholar] [CrossRef]
- Wolever, T.M.; Jenkins, D.J. What is a high fiber diet? Adv. Exp. Med. Biol. 1997, 427, 35–42. [Google Scholar] [CrossRef]
- Kaaks, R.J. Biochemical markers as additional measurements in studies of the accuracy of dietary questionnaire measurements: conceptual issues. Am. J. Clin. Nutr. 1997, 65, 1232S–1239S. [Google Scholar]
- Ross, A.B; Kamal-Eldin, A.; Aman, P. Gas chromatographic analysis of alkylresorcinols in rye (secale cereale L) grains. J. Sci. Food Agric. 2001, 81, 1405–1411. [Google Scholar] [CrossRef]
- Ross, A.B.; Kamal-Eldin, A.; Aman, P. Dietary alkylresorcinols: absorption, bioactivities, and possible use as biomarkers of whole-grain wheat- and rye-rich foods. Nutr. Rev. 2004, 62, 81–95. [Google Scholar] [CrossRef]
- Koskela, A.; Linko-Parvinen, A.M.; Hiisivuori, P.; Samaletdin, A.; Kamal-Eldin, A.; Tikkanen, M.J.; Adlercreutz, H. Quantification of alkylresorcinol metabolites in urine by HPLC with coulometric electrode array detection. Clin. Chem. 2007, 53, 1380–1383. [Google Scholar] [CrossRef]
- Ross, A.B.; Aman, P.; Kamal-Eldin, A. Identification of cereal alkylresorcinol metabolites in human urine-potential biomarkers of wholegrain wheat and rye intake. J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 2004, 809, 125–130. [Google Scholar] [CrossRef]
- Guyman, L.A.; Adlercreutz, H.; Koskela, A.; Li, L.; Beresford, S.A.; Lampe, J.W. Urinary 3-[3, 5-dihydroxyphenyl]-1-propanoic acid, an alkylresorcinol metabolite, is a potential biomarker of whole-grain intake in a U.S. population. J. Nutr. 2008, 138, 1957–1962. [Google Scholar]
- Ross, A.B.; Shepherd, M.J.; Bach Knudsen, K.E.; Glitso, L.V.; Bowey, E.; Phillips, J.; Rowland, I.; Guo, Z.X.; Massy, D.J. Absorption of dietary alkylresorcinols in ileal-cannulated pigs and rats. Br. J. Nutr. 2003, 90, 787–794. [Google Scholar] [CrossRef]
- Linko, A.M.; Parikka, K.; Wahala, K.; Adlercreutz, H. Gas chromatographic-mass spectrometric method for the determination of alkylresorcinols in human plasma. Anal. Biochem. 2002, 308, 307–313. [Google Scholar] [CrossRef]
- Aubertin-Leheudre, M.; Koskela, A.; Marjamaa, A.; Adlercreutz, H. Plasma alkylresorcinols and urinary alkylresorcinol metabolites as biomarkers of cereal fiber intake in Finnish women. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 2244–2248. [Google Scholar] [CrossRef]
- Landberg, R.; Kamal-Eldin, A.; Andersson, A.; Vessby, B.; Aman, P. Alkylresorcinols as biomarkers of whole-grain wheat and rye intake: plasma concentration and intake estimated from dietary records. Am. J. Clin. Nutr. 2008, 87, 832–838. [Google Scholar]
- Linko, A.M.; Juntunen, K.S.; Mykkanen, H.M.; Adlercreutz, H. Whole-grain rye bread consumption by women correlates with plasma alkylresorcinols and increases their concentration compared with low-fiber wheat bread. J. Nutr. 2005, 135, 580–583. [Google Scholar]
- Landberg, R.; Linko, A.M.; Kamal-Eldin, A.; Vessby, B.; Adlercreutz, H.; Aman, P. Human plasma kinetics and relative bioavailability of alkylresorcinols after intake of rye bran. J. Nutr. 2006, 136, 2760–2765. [Google Scholar]
- Landberg, R.; Aman, P.; Friberg, L.E.; Vessby, B.; Adlercreutz, H.; Kamal-Eldin, A. Dose response of whole-grain biomarkers: alkylresorcinols in human plasma and their metabolites in urine in relation to intake. Am. J. Clin. Nutr. 2009, 89, 290–296. [Google Scholar]
- Aubertin-Leheudre, M.; Koskela, A.; Samaletdin, A.; Adlercreutz, H. Plasma Alkylresorcinol metabolites as potential biomarkers of whole-grain wheat and rye cereal fiber intake in women. Br. J. Nutr. 2009, 103, 339–343. [Google Scholar] [CrossRef]
- Luhrmann, P.M.; Herbert, B.M.; Gaster, C.; Neuhauser-Berthold, M. Validation of a self-administered 3-day estimated dietary record for use in the elderly. Eur. J. Nutr. 1999, 38, 235–240. [Google Scholar] [CrossRef]
- Paul, A.A.; Southgate, D.A. McCance and Widdowson's 'The composition of foods': dietary fibre in egg, meat and fish dishes. J. Hum. Nutr. 1979, 33, 335–336. [Google Scholar]
- Koskela, A.; Samaletdin, A.; Aubertin-Leheudre, M.; Adlercreutz, H. Quantification of alkylresorcinol metabolites in plasma by high-performance liquid chromatography with coulometric electrode array detection. J. Agric. Food Chem. 2008, 56, 7678–7681. [Google Scholar] [CrossRef]
- Borel, P. Factors affecting intestinal absorption of highly lipophilic food microconstituents (fat-soluble vitamins, carotenoids and phytosterols). Clin. Chem. Lab. Med. 2003, 41, 979–994. [Google Scholar] [CrossRef]
- van't Veer, P.; Kardinaal, A.F.; Bausch-Goldbohm, R.A.; Kok, F.J. Biomarkers for validation. Eur. J. Clin. Nutr. 1993, 47, S58–S63. [Google Scholar]
- Slotnick, M.J.; Nriagu, J.O. Validity of human nails as a biomarker of arsenic and selenium exposure: A review. Environ. Res. 2006, 102, 125–139. [Google Scholar] [CrossRef]
- Maruvada, P.; Srivastava, S. Biomarkers for cancer diagnosis: implications for nutritional research. J. Nutr. 2004, 134, 1640S–645S. [Google Scholar]
- Linko-Parvinen, A.M.; Landberg, R.; Tikkanen, M.J.; Adlercreutz, H.; Penalvo, J.L. Alkylresorcinols from whole-grain wheat and rye are transported in human plasma lipoproteins. J. Nutr. 2007, 137, 1137–1142. [Google Scholar]
- Ross, A.B.; Chen, Y.; Frank, J.; Swanson, J.E.; Parker, R.S.; Kozubek, A.; Lundh, T.; Vessby, B.; Aman, P.; Kamal-Eldin, A. Cereal alkylresorcinols elevate gamma-tocopherol levels in rats and inhibit gamma-tocopherol metabolism in vitro. J. Nutr. 2004, 134, 506–510. [Google Scholar]
- Linko, A.M.; Adlercreutz, H. Whole-grain rye and wheat alkylresorcinols are incorporated into human erythrocyte membranes. Br. J. Nutr. 2005, 93, 11–13. [Google Scholar] [CrossRef]
- Birringer, M.; Drogan, D.; Brigelius-Flohe, R. Tocopherols are metabolized in HepG2 cells by side chain omega-oxidation and consecutive beta-oxidation. Free Radic. Biol. Med. 2001, 31, 226–232. [Google Scholar] [CrossRef]
- Ross, A.B.; Becker, W.; Chen, Y.; Kamal-Eldin, A.; Aman, P. Intake of alkylresorcinols from wheat and rye in the United Kingdom and Sweden. Br. J. Nutr. 2005, 94, 496–499. [Google Scholar] [CrossRef]
- Landberg, R.; Kamal-Eldin, A.; Andersson, S.O.; Johansson, J.E.; Zhang, J.X.; Hallmans, G.; Aman, P. Reproducibility of plasma alkylresorcinols during a 6-week rye intervention study in men with prostate cancer. J. Nutr. 2009, 139, 975–980. [Google Scholar] [CrossRef]
- Linko, A.M.; Ross, A.B.; Kamal-Eldin, A.; Serena, A.; Kjaer, A.K.; Jorgensen, H.; Penalvo, J.L.; Adlercreutz, H.; Aman, P.; Knudsen, K.E. Kinetics of the appearance of cereal alkylresorcinols in pig plasma. Br. J. Nutr. 2006, 95, 282–287. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Aubertin-Leheudre, M.; Koskela, A.; Samaletdin, A.; Adlercreutz, H. Responsiveness of Urinary and Plasma Alkylresorcinol Metabolites to Rye Intake in Finnish Women. Cancers 2010, 2, 513-522. https://doi.org/10.3390/cancers2020513
Aubertin-Leheudre M, Koskela A, Samaletdin A, Adlercreutz H. Responsiveness of Urinary and Plasma Alkylresorcinol Metabolites to Rye Intake in Finnish Women. Cancers. 2010; 2(2):513-522. https://doi.org/10.3390/cancers2020513
Chicago/Turabian StyleAubertin-Leheudre, Mylène, Anja Koskela, Adile Samaletdin, and Herman Adlercreutz. 2010. "Responsiveness of Urinary and Plasma Alkylresorcinol Metabolites to Rye Intake in Finnish Women" Cancers 2, no. 2: 513-522. https://doi.org/10.3390/cancers2020513
APA StyleAubertin-Leheudre, M., Koskela, A., Samaletdin, A., & Adlercreutz, H. (2010). Responsiveness of Urinary and Plasma Alkylresorcinol Metabolites to Rye Intake in Finnish Women. Cancers, 2(2), 513-522. https://doi.org/10.3390/cancers2020513