1. Introduction
Cnidarian envenomation represents a significant medical burden worldwide. Envenomation involves the triggering and discharge of hundreds to thousands of specialized cells, called cnidocytes or nematocytes. Certain cnidocytes contain penetrant cnidae; these microscopic venom-injecting capsules are called nematocysts. Upon contact, nematocysts can deliver a potently toxic chemical cocktail into unsuspecting prey or human victims [
1]. Pelagic cnidarian species that possess long tentacles armed with cnidocytes are colloquially known as “jellies” or “jellyfish”, and represent the most dangerous members of the phylum. Jellyfish envenomation can range from mild to life threatening, depending on the species involved, and the amount of venom delivered [
2,
3,
4].
The marine siphonophores
Physalia utriculus (Indo-Pacific; commonly called bluebottle) and
P. physalis (Atlantic; referred to as Portuguese man o’ war) are dangerous marine organisms commonly found in tropical oceans. Each consists of an above-water float (pneumatophore), various specialized feeding and reproductive structures and long, stinging tentacles used for capturing prey. These marine stingers are often found in large groups or “armadas” that can be blown onshore by strong winds. While deaths from
Physalia stings are rare [
5,
6], stings can be excruciating and lead to systemic complications, including headache, vomiting, abdominal pain and diarrhea [
1,
2,
3,
4,
7,
8,
9]. Their penetrant cnidae (nematocysts) have long tubules, which can deliver venom nearly 1 mm into tissues and remain embedded, likely causing secondary immunological reactions [
10]. A recent study has described a marked increase in
Physalia stings along the French Atlantic coasts [
11], further stressing the urgency of standardized, evidence-based care for
Physalia stings.
While vinegar dousing has been recommended historically [
12], there is no universally accepted first aid for
Physalia stings, and first-aid approaches have recently become increasingly controversial [
13]. Because studies have shown that physiological responses to all cnidarian envenomation are dose-dependent, and only a small portion of available cnidae discharge upon initial contact [
14], one of the key first steps in first aid is to ensure the safe removal of adherent tentacles and undischarged cnidae (which are capable of firing for at least two weeks after their separation from the tentacle [
15]). Thus, a critical initial first-aid goal is to remove stinging material without increasing the amount of venom injected or the number of cnidae discharged into the skin, and thus rinse solutions that irreversibly inhibit cnidae discharge are preferred. Rinses that trigger functional cnidae discharge to effect further impalement of the epidermis with venom deposition cause more harm than good, potentially turning a mild or moderate sting into a severe one. In a somewhat counter-intuitive manner, however, certain rinses such as sea water which may appear at first consideration to be “inert” and represent a “do no harm” type of approach, simply allow the tentacles, as well as dissociated intact cnidae, to be moved further on the surface of the skin at which point the tentacle cnidae or isolated intact cnidae sting, thus “spreading” the sting area and increasing the “venom load”. Further, another potential area of unexpected contradiction is the finding that while some rinses induce discharge of cnidae along an isolated tentacle on a microscope slide, the discharged cnidae are not functionally capable of impaling skin and/or the venom is immediately inactivated. These two findings caused us to carefully re-examine the extant literature. The prevailing premise has been that microscope slide assays of tentacle responses to potential rinse solutions directly correlate to venom load and thus utility in sting management. The finding that this is not the case [
16] is pivotal in efforts to develop optimal sting management protocols.
The few investigations on potential rinse solutions for
Physalia stings have been fraught with inconsistent and contradictory results. Initial studies reported that alcohols cause massive cnidae discharge, while discharge is absent or inhibited in the presence of weak acetic acid solutions (~5%, in distilled water), or household vinegar [
17], a finding confirmed in another hydromedusan species,
Olindias sambaquiensis [
18]. Some reduction in pain was also noted with vinegar application on
Physalia stings in a prospective controlled clinical trial (
N = 20) [
19]. However, microscopic examination in another study [
20] found moderate cnidae discharge in the presence of vinegar (an average of “2” out of “5”, with 5 being “maximal” discharge), and a more recent paper [
21] observed what they described as a relatively high degree of discharge among the data observed (nematocysts per 1 mm of linear tentacle; roughly double the discharge observed after exposure to 1:10 dilution of 70% ethanol) from
Physalia physalis tentacles with the application of dilute (1:10) 5% acetic acid. Because of the variability in laboratory-based results, it was suggested in the 1980s that vinegar use for
Physalia stings be discontinued [
20,
22]. This suggestion has become standard; medical doctors warn of vinegar’s danger, even calling its use “forbidden” in the case of
Physalia stings [
23] (p. 86), and national and international organizations often question or warn against the use of vinegar if
Physalia is implicated (e.g., [
24,
25,
26]). The recommendation has stuck despite the fact that the combined evidence from the highest-quality studies supports vinegar as the best course of first aid [
27].
In this study, we sought to investigate the degree of cnidae discharge during a sting event. We then sought to re-examine potential rinse solutions previously tested by other investigators, as well as popular folk remedies (such as urine), to rigorously determine which solutions inhibit Physalia cnidae discharge to determine the best choices for first-aid use. We compared the results for Atlantic and Pacific Physalia species to look for species-specific responses, as well as determine whether there is likely to be a universally applicable solution for Physalia species. Lastly, we examined whether the results of solution-based methods that evaluate cnidae discharge correlate to functional measures of venom load (in a blood agarose-based model), to further evaluate whether solution-based studies are clinically relevant.
3. Discussion
The integrated structural and functional approach taken in this study demonstrates a potential explanation for the level of controversy in the literature related to sting first aid management. Based upon our findings in cubozoa [
15] and other species including the current study of the siphonophore
Physalia, it appears that TSA-based physical data and the presence or absence of discharged cnidae, is not a useful predictor of sting severity, i.e., a functional metric of venom induced tissue damage. In summary, this study further supports the use of cnidae-inactivating rinse solutions as a primary step in the cnidarian envenomation first aid. We found that roughly 0.5% of tentacle cnidae discharged within the first 10 min of tentacle contact to the blood agar blood cell tissue model; in previous work, stinging of this proxy was indistinguishable from an authentic human sting [
14]. A maximum of about 50% discharge was elicited when tentacles were stimulated chemically or mechanically (
Table 1 and
Table 2), indicating that there may be a biological constraint on the total amount of cnidae available to discharge. Given that cnidae are spent and replenished during feeding, this observation makes sense, as its very likely many cnidae may be immature. If we assume that our maximal inducible discharge represents the maximum available cnidae during a sting, then we would estimate that about 1% of available cnidae discharge during the initial sting event. Many of the undischarged yet available cnidae could be shed during tentacle contact, as we found large numbers of undischarged cnidae on the agarose surface. Many of these cnidae were able to fire when gently pressed, and thus should be considered active and dangerous. Between the tentacle itself and the contact area, nearly 100 times the number of stinging cnidae may be available to discharge into an unfortunate sting victim, and thus safe and effective removal of adherent tentacles and undischarged cnidae should be considered a first-aid priority.
Overall, we found strong similarities in the responses of both species to various solutions. In two cases, however, we saw slight differences in vitro: the Pacific P. utriculus was less responsive to both urine and tap water than the Atlantic P. physalis. These discrepancies may be due to differences in collection method rather than representing species-specific responses. All Pacific Physalia used in the TSAs were collected after they washed ashore and were beached for some time (<2 h beached, but exactly how long for each individual was unknown). The Atlantic specimens used for the TSAs, on the other hand, were collected live from the Gulf Stream using nets. As such, they would not have experienced any period of time out of water or dehydration or depletion due to rolling contact with sand and may thus have been more sensitive to certain solutions, especially aqueous ones.
To our knowledge, this study represents the first laboratory test of shaving cream on
Physalia tentacles, which is often suggested by authoritative web sources (e.g., [
28,
29]) including the United Kingdom National Health Service [
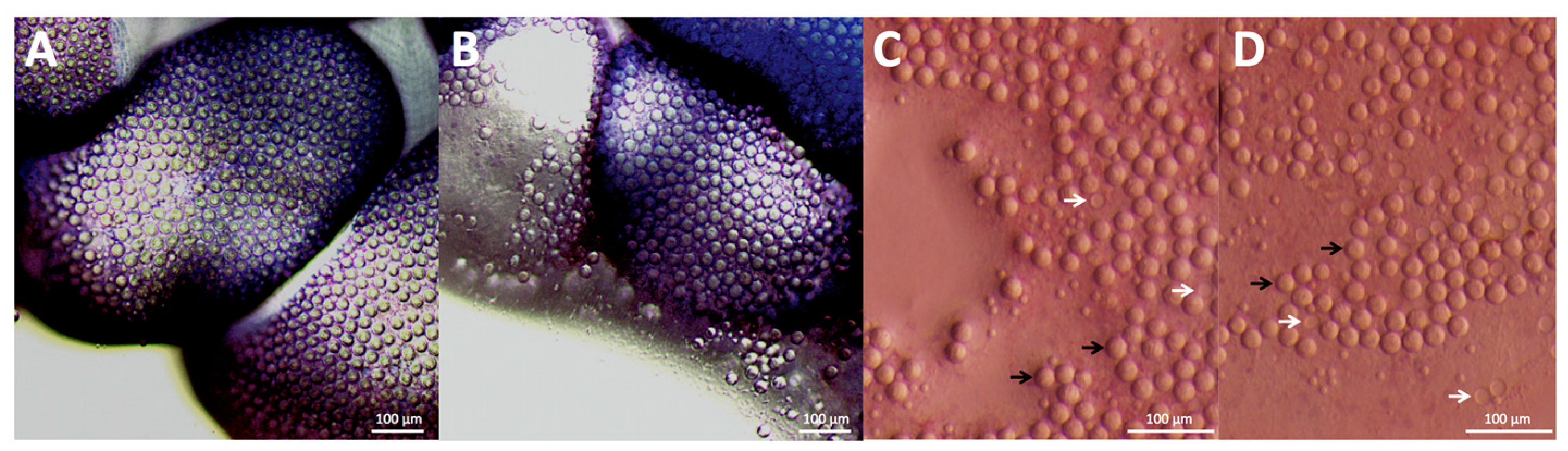
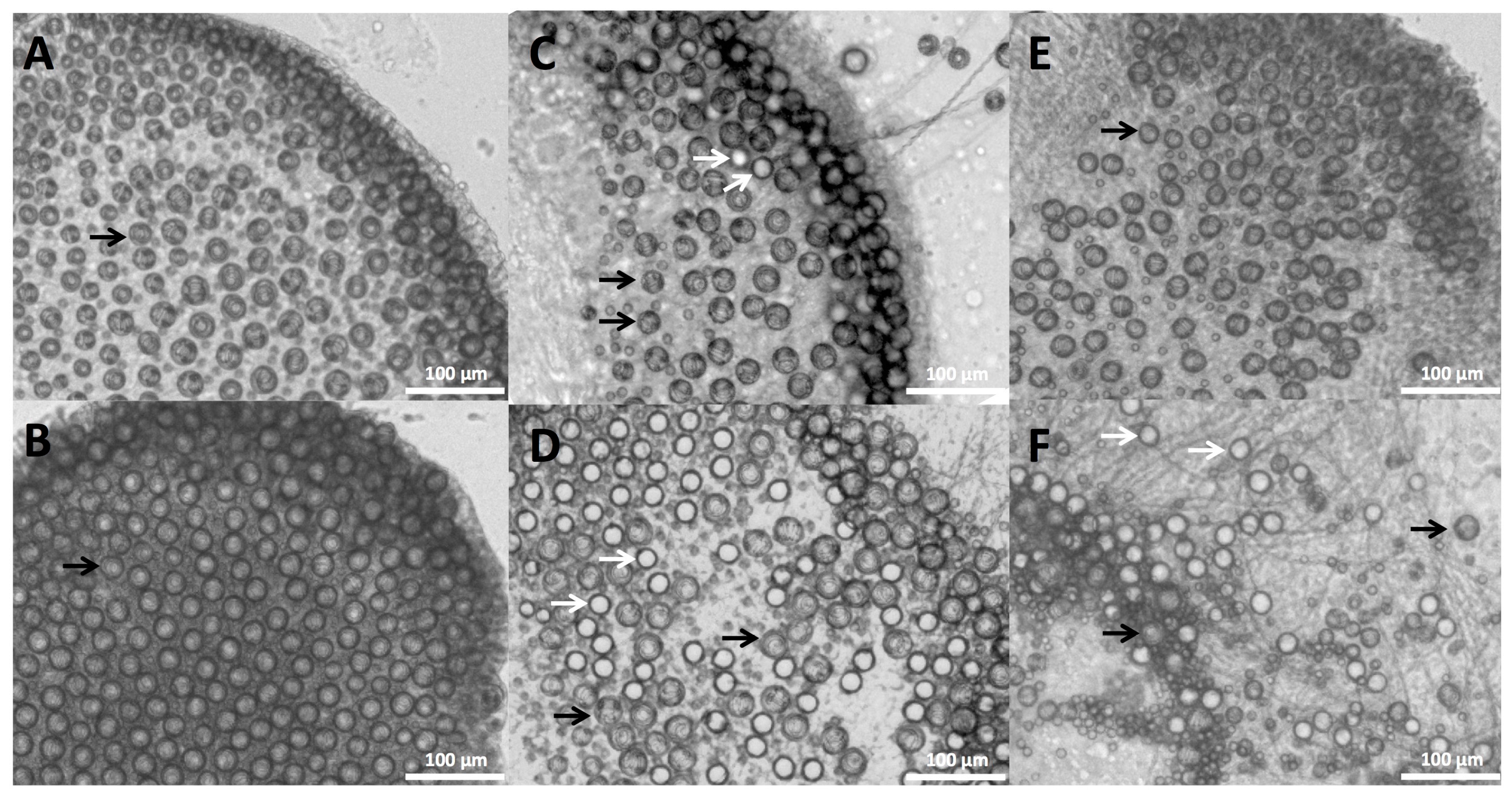
25] to be used in combination with a razor or credit card to “shave” away adherent cnidae. As scraping the skin would undoubtedly act to apply pressure to any adherent cnidae and therefore increase discharge [
15] (in our test, slight site pressure increased adherent discharge by three fold; see
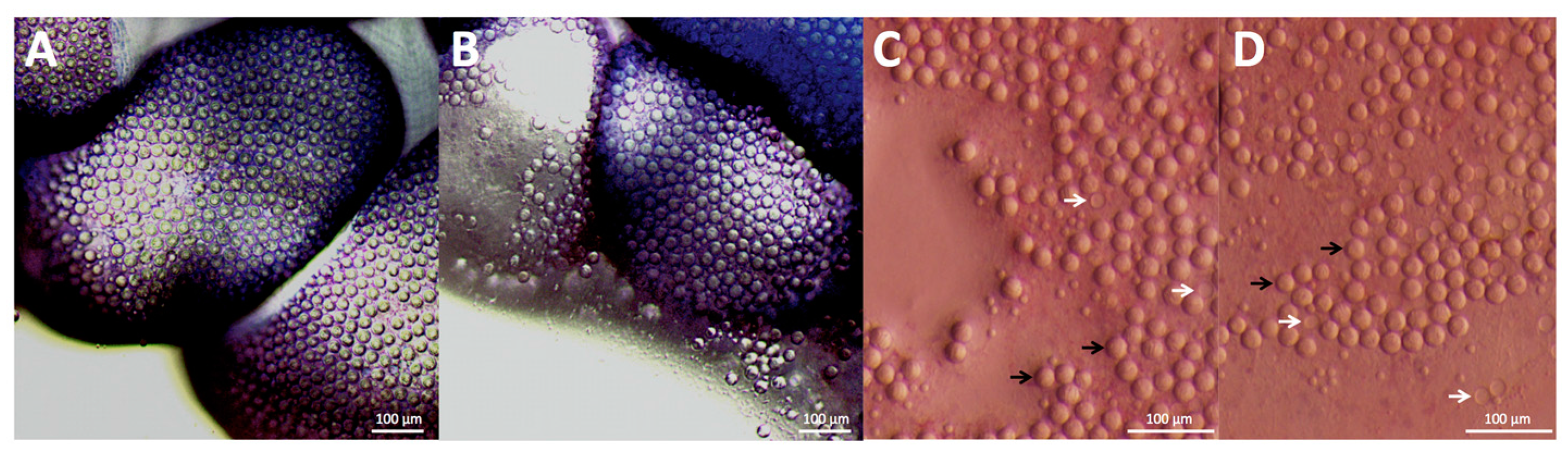
Figure 1C,D), we tested the ability of shaving cream and other compounds to prevent cnidae discharge regardless of whether they induce discharge. No readily available solutions outside of vinegars fully inhibited pressure-induced discharge, and many showed large increases in discharge after pressure application (
Table 2), thus our data would suggest scraping the site should not occur unless the area is doused with vinegar first.
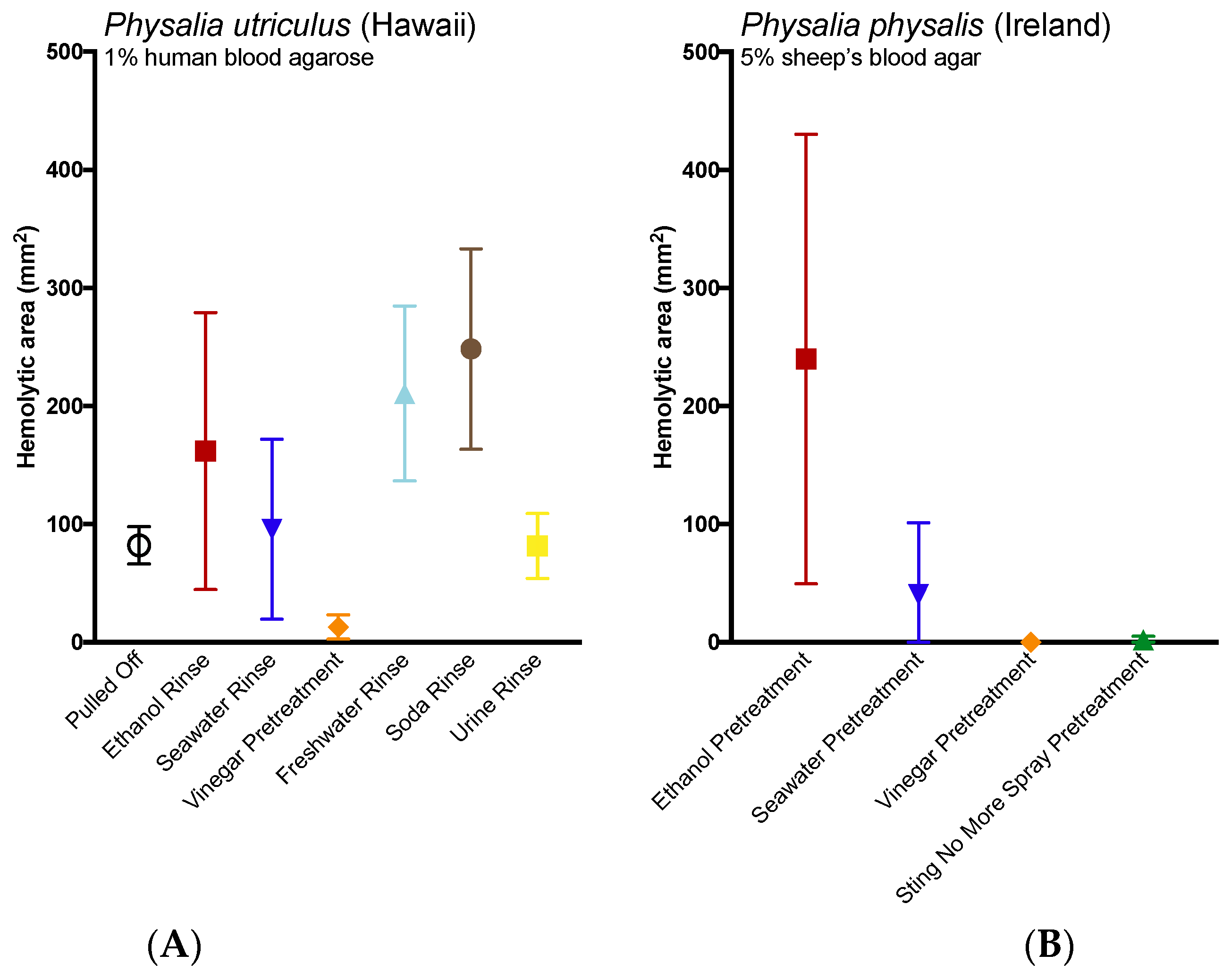
These results strongly supported the use of vinegar as a rinse solution; vinegars acted as potent inhibitors of both chemically-induced and pressure-induced cnidae discharge (
Table 1 and
Table 2). We found that several varieties of vinegar as well as laboratory-made dilute acetic acid solutions inhibited cnidae discharge in both species no matter the stimulus (
P. physalis results can be found in
Table A1). Further, the pre-treatment of tentacles for 30 s with vinegar significantly reduced their ability to sting (
Figure 3). These results are concurrent with Burnett et al., 1983 [
17], but contrast three other studies [
20,
21,
22]. However, a closer examination of the methods used in the contrasting studies may explain the apparent discrepancies in results. Exton [
20] rated discharge from vinegar, methylated spirits and water relative to the “maximal” (a score of 5) and “minimal” (a score of 0) discharge seen; vinegar scored 2/5, spirits scored 5/5, and water scored 0/5. It is difficult to compare these results with our own as the scale is unclear and there was no indication of variability or error; at best, we can say that in their study vinegar was substantially better than ethanol, which would be consistent with our results. Birsa et al. [
21] found that “5% acetic acid” induced discharge in
P. physalis; the addition of acetic acid led to nearly double the amount of discharge of cnidae as “70% ethanol” (100 and 53 ± 26 nematocysts per mm of tentacle, respectively). Their alcohol results contrast Exton’s as well as the results presented here, as we found 70% ethanol induced dramatically more discharge in both species tested (
Table 1). However, Birsa et al. [
20] noted that their discharge metric—the number of nematocysts discharged per millimeter of tentacle length—was flawed due to the variability of nematocyst density on tentacles, and thus they ”suggest that at best it is a semi-quantitative” (p. 427). It is unclear why they did not count undischarged cnidae to obtain a standardized value, but since there is no indication of how variable the cnidae density was between tentacles or what the average density was, it is unclear whether the count for 5% acetic acid is comparable to the other counts in their table, or what degree of discharge their counts represent (if our tentacle counts of cnidae density were used, then 100 cnidae per mm would translate to 1.8% discharge, as compared with our findings of 0.5% for 5% acetic acid and 1.7% for 0.5% acetic acid). However, perhaps most confounding is that Birsa et al. [
20] also reported in their methods that they added 100 µL of their test solution to a well containing 1 mL of seawater and the tentacle, which would have resulted in a final concentration of 0.5% acetic acid rather than 5% and 7% ethanol rather than 70%. Further complicating comparisons is that acetic acid appears to have been only counted once (the value for acetic acid does not have error margins while values for their other solutions do), thus it is impossible to determine if that count of discharged cnidae is representative of the animals’ response. We found in our studies that the dilution of acetic acid solutions completely abolished their protective effects, and that diluted solutions could even induce slight discharge (
Table 2); meanwhile, dilute alcohol solutions induced markedly lower discharge than concentrated alcohols (
Table 1). Based on microscope images alone, it appears that very little discharge was induced in
P. physalis overall, as only a few isolated tubules can be seen (p. 428,
Figure 1A for acetic acid,
Figure 1D for ethanol)—which would be consistent with the results presented in
Table 2 for diluted solutions. The only other paper to report discharge from vinegar in
Physalia had methods too vague to compare [
22]. Fenner et al. stated that in their experiments, discharge occurred in “up to 80% of nematocysts”, but that this only occurred in “some, not all of the specimens” and that “conflicting results were obtained if other parts of the tentacles were tested, even from the same jellyfish”, and that “approximately 30%...showed some discharge” [
22] (p. 499). Studies on other hydrozoans have found limited or no discharge induced by vinegar and strong discharge from alcohols, similar to our results [
18,
30].
The inhibition of discharge by vinegar was not due to its acidic nature alone, as other solutions with similar pH were unable to prevent discharge. Vinegar and dilute acetic acid solutions have long been used to preserve food and fix tissues [
31]. Vinegar or ~5% acetic acid causes marked swelling of collagen by increasing the absorption of water [
31]. It has long been documented that vinegar exposure “fixes” nematocysts rendering them incapable of functional discharge. This is likely due to the fact that nematocyst capsule walls are comprised of collagen and that acetic acid-induced swelling irreversibly alters the structural features required for functional firing [
10]. We found strong evidence against the use of ethanol and freshwater, as these solutions significantly increased cnidae discharge (
Table 2) and worsened stings in the TBAA model (
Figure 3). In addition, our results do not support the use of the most infamous sting treatment: urine. It is unclear exactly when the use of urine for jellyfish stings became popular (certainly, a scene featuring the treatment method in the sitcom “Friends” aided its spread [
32]), but it has become one of the most persistent myths in toxinology. Urine induced significant discharge in both species of
Physalia tested, with discharge on par with ethanol application for
P. physalis, the larger and more dangerous species. While urine did not cause significant increases in hemolysis in our functional assay, we did not have the opportunity to test
P. physalis using the TBAA. Furthermore, even with the less-responsive species, we did detect an increase in the hemolytic zone size after 22 h, which was not statistically significant perhaps due to the small sample sizes (
N = 5). Further investigations into the effects of urine in both solution-based and functional assays should be conducted, and, in the meantime, its use for treating stings discouraged.
Although we did not see a significant increase in discharge from the application of soda in the TSA (
Table 2), it most strongly increased hemolysis in the TBAA (
Figure 4). Why soda caused such damage in the ex vivo model is unclear; soda alone did not cause hemolysis. It is possible that sugars or other compounds present or carbonation acted to enhance cnidae discharge in the presence of perceived prey (blood cells). Further tests are required to examine the basis of the increase in hemolysis triggered by the addition of soda. However, perhaps more importantly, this discrepancy highlights the need for investigating potential first aid approaches using multiple methods, as a lack of induction of discharge in the TSA does not consistently correlate with the functional metric of venom activity, hemolysis, and thus is of questionable value as a determinant of efficacy of first aids. Live tentacle cnidocytes are known to be associated with sensory hair cells [
33], and cnidae triggering involves both chemo- and mechano-sensory input. From a methodological standpoint, the effects of solutions on a tentacle laying on an inert glass surface differ from tentacle contact to the epidermis of live prey. Thus, the TSA does not necessarily predict potential changes in cnidae triggering in the presence of human cells, nor can it account for the physical movement of the tentacle or cnidae that may be induced by the application of a first-aid. For example, rinsing with an “inert” solution (defined as not triggering discharge in the TSA) may have deleterious effects in functional models, as the act of rinsing could cause tentacles to shift or roll. Such movements could place more cnidae in contact with the skin locally, increasing stinging, as well as move the stinging cells to previously untouched patches of skin, increasing the size of the sting site. Experimentally, this has been demonstrated to occur with cubozoans; seawater rinsing was found to worsen stings even though it does not directly induce discharge [
15]. While the TSA can be used as a preliminary screen to look for potent cnidae triggers, which can be dangerous to apply to stings (such as ethanol), or effective inhibitors of discharge (such as vinegar), it should not be considered a conclusive test for efficacy and safety, especially for solutions which neither trigger nor inhibit cnidae discharge in vitro.
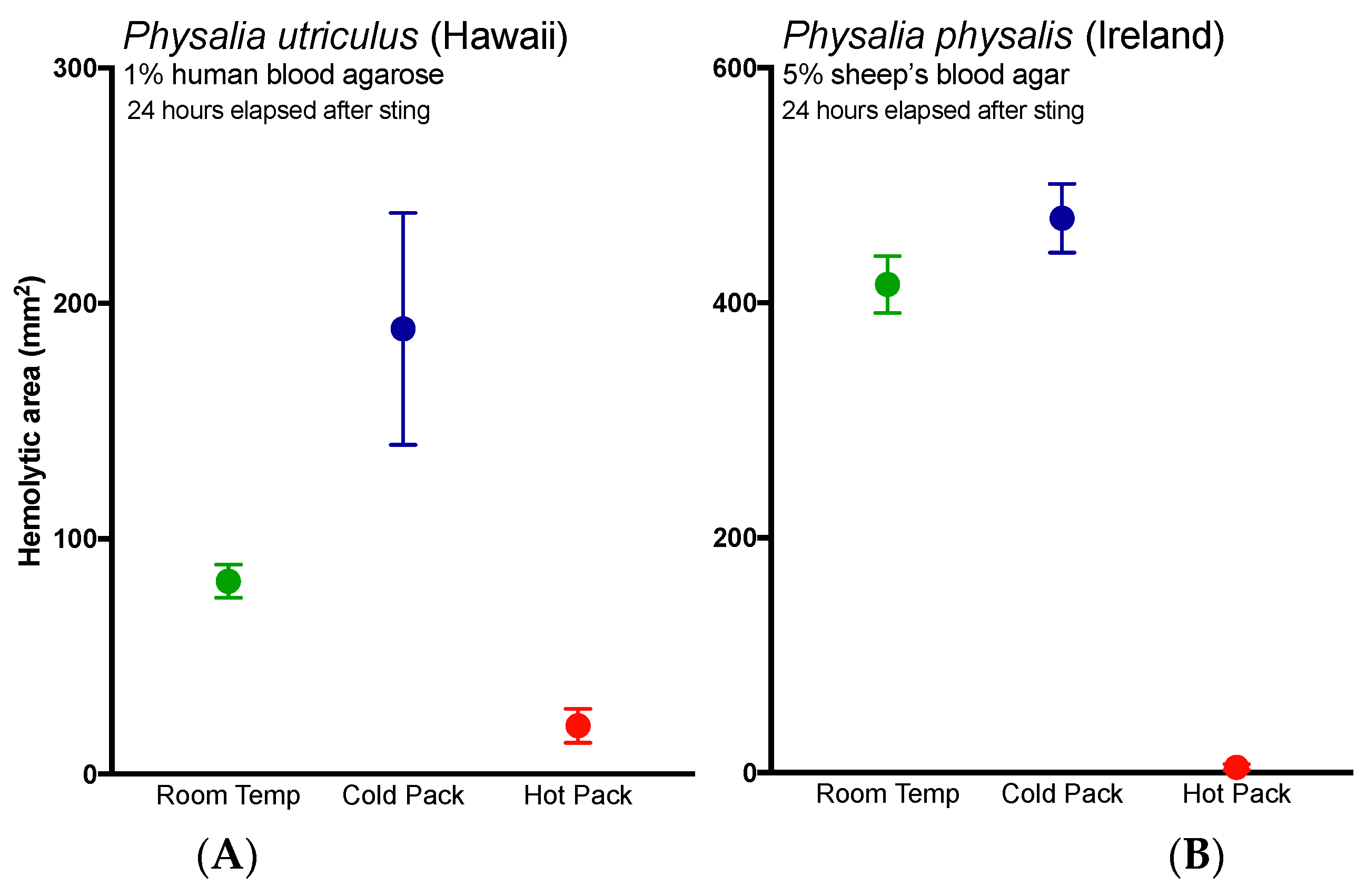
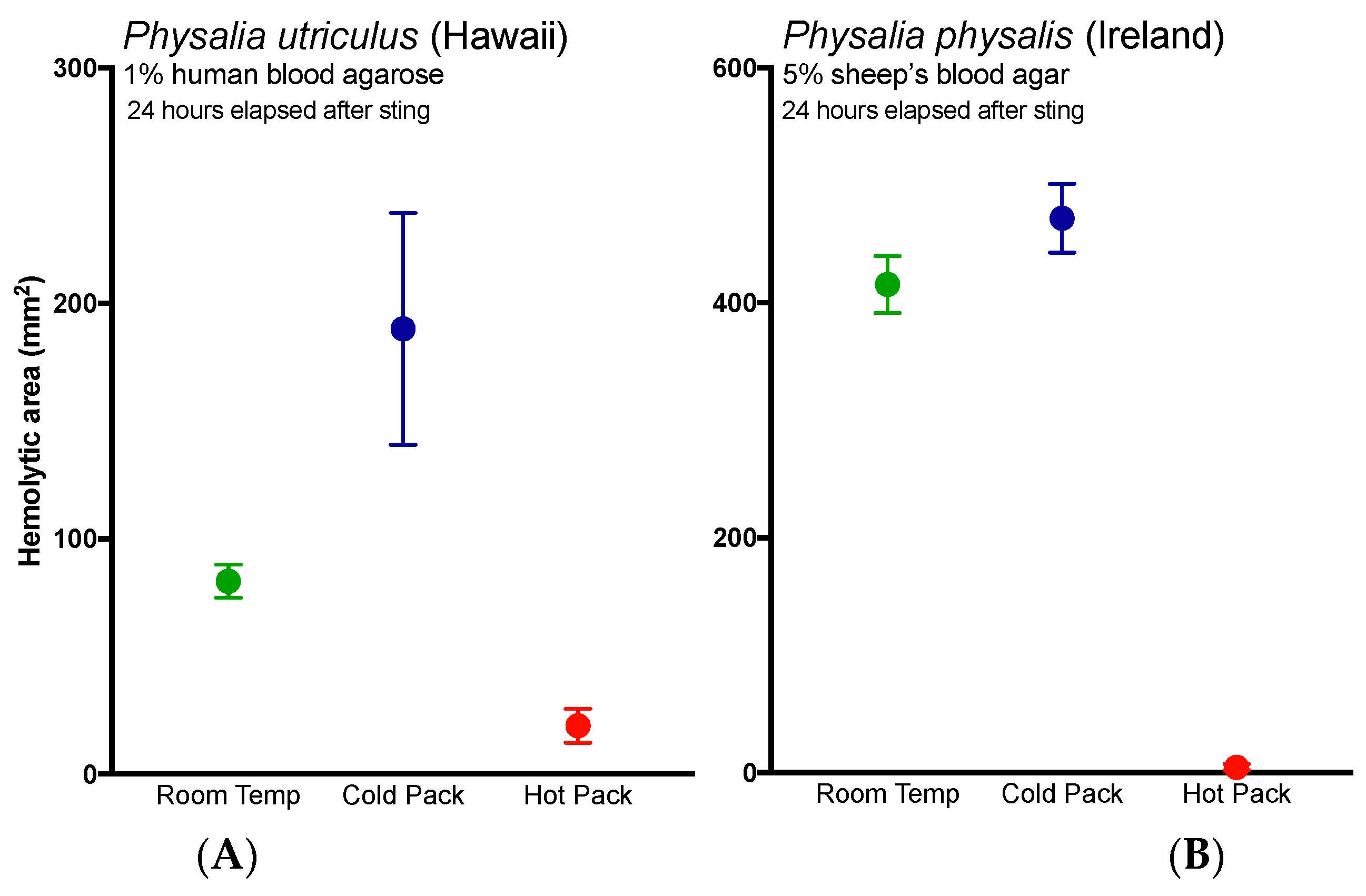
Lastly, these results bolster previous studies that support the use of heat in the treatment of
Physalia stings [
34,
35], as the application of heat significantly reduced hemolysis in the TBAA (
Figure 3A, red line versus black line). The application of cold not only failed to reduce hemolysis, it worsened stings (
Figure 3, blue lines). While it is possible that the application of cold increased discharge of shed cnidae, thus directly increasing venom load, we posit an alternative possibility. These results suggest that while the physical trauma of a sting and initial pain are acute events, venom pathogenic mechanisms may have a protracted kinetic course. The enzyme kinetics have yet to be carefully elucidated but time course studies (data not shown) reveal that the lipase reaction kinetics exhibit substrate to product conversion over 12 h comparable to other cnidarian lipases [
36]. The effects of initial temperature treatments on such kinetics are not known; however, it is reasonable to believe that cold treatment could enhance venom activity, as previous studies have found that activity is preserved at cold temperatures and abolished by hot ones (for a review, see [
37]). Further, anecdotal accounts have noted increases in pain or “reawakening” (rebound) of the sting upon rewarming after the application of cold [
38,
39]. To our knowledge, this is the first quantitative report of both acute and protracted harm from the use of cold packs, and warrants immediate reevaluation of the use of cold packs in the treatment of cnidarian envenomations. Studies evaluating the possible injurious effects of cold pack application in more lethal species should be conducted expediently.
5. Materials and Methods
The chemicals for the solutions used in all assays are as follows: seawater (locally collected, 0.45 μm filtered), white vinegar (Bakers and Chefs CJ314, SAM’s West Inc., Bentonville, AR, USA), cider vinegar (Heinz Brand, H.J. Heinz Corp., Pittsburgh, PA, USA), malt vinegar (Heinz Brand, H.J. Heinz Corp., Pittsburgh, PA, USA), balsamic vinegar (Safeway, Pleasanton, CA, USA), shaving cream (Gillette Regular Foam, Procter and Gamble, Boston, MA, USA), baking soda (mixed 3:1 with filtered seawater; Arm and Hammer, Church and Dwight Co., Princeton, NJ, USA), dish soap (Dawn brand, Procter and Gamble, Cincinnati, OH, USA), acetic acid (Fisher Scientific, Fair Lawn, NJ, USA), lidocaine (4% in 150 mM saline; MP Biomedicals LLC, Solon, OH, USA), ethanol (Pharmco-Aaper, Brookfield, CT, USA), isopropanol (Fisher Scientific), copper gluconate (30 mM in 150 mM saline; Strem Chemicals, Newburyport, MA, USA), magnesium sulfate (50 mM in filtered 150 mM saline; Fisher Scientific), sodium chloride (Fisher Scientific), hydrochloric acid (Fisher Scientific), lemon juice (Mott’s LLP, Pano, TX, USA), regular cola (Pepsi; Pepsico, Purchase, NY, USA), Sting No More® products (contents include copper gluconate, urea, and magnesium sulfate; Alatalab Solutions™ LLC, Honolulu, HI, USA), copper gluconate (30 mM in 150 mM saline; Strem Chemicals, Newburyport, MA, USA). Urine was freshly collected from a willing volunteer prior to use and 0.45 μm filtered. The vinegar used in Ireland was Tesco distilled vinegar (produced in the UK for Tesco Stores Ltd., Chestnut, UK).
5.1. Animal Collection
Live P. utriculus were collected after sustained strong winds from Kailua Beach on the eastern side of Oahu (21°23′47.7″ N 157°43′29.2″ W). Animals were placed in individual containers with ample seawater and kept cool until use. Live P. physalis for solution-based assays were collected from gulfstream waters off Miami’s South Beach (25°50′48.3″ N 79°48′57.7″ W). Animals were netted and bagged in gallon Ziploc bags with ample seawater and kept cool until use. Animals were warmed to room temperature in 0.45 μm-filtered seawater before their tentacles were used. All experiments were conducted within 72 h of collection. Live P. physalis for blood agar assays were also collected from several beaches (Derrynane, Co. Kerry, Youghal, Co. Cork and Ardmore, Co. Waterford, all in Ireland) during periods of strong southwesterly winds. The majority of specimens were collected within a few hours of stranding and several were collected from the incoming tide. The specimens were typically small, with pneumatophore lengths of 60 mm to 150 mm. They were kept in ample amounts of ambient seawater for up to 24 h, before having their fishing tentacles excised. Tentacles were excised close to the pneumatophore using a pair of dissecting scissors and were handled with a fine forceps before being immediately used in the experiments.
5.2. Envenomation Modeling
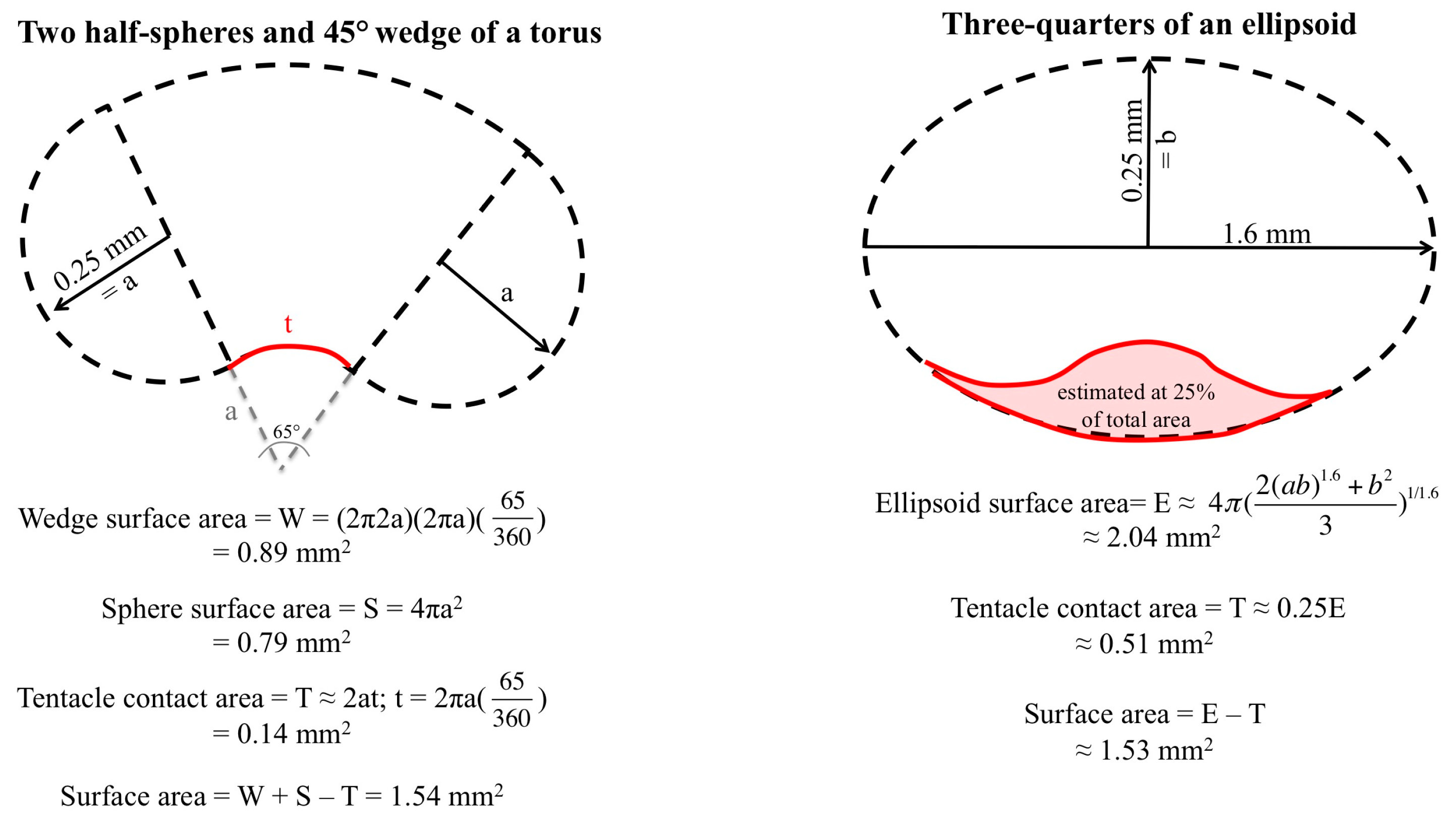
The
Physalia cnidome consists of several cnidae types, including two nematocysts: large and small heterotrichous anisorhizas [
10,
40]. Given their size and the length of their tubules (up to 1 mm long), the most important nematocysts for venom delivery are the bigger anisorhizas; we thus sought to quantify the discharge of these large, penetrant cnidae during a sting event. We utilized live human RBC from normal donors (approved protocol CHS#12561, University of Hawaii Committee on Human Studies) and low melting point agarose (Nusieve GTG Agarose, Lonza, Rockland, ME, USA) to constitute a live red blood cell agarose, a proxy for human tissue described in [
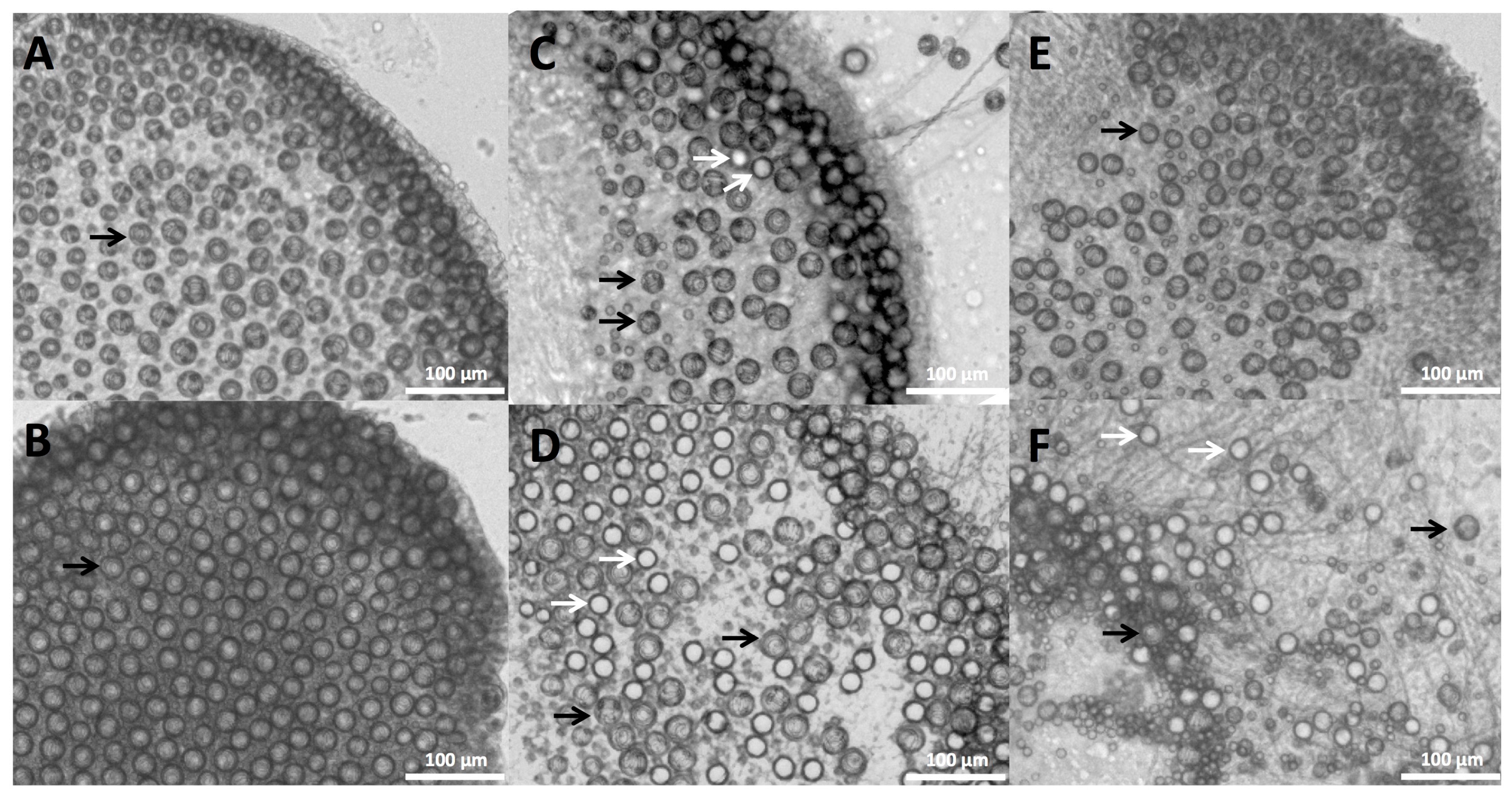
14]. To calculate tentacle cnidae and cnidae discharge, approximately 1 cm of freshly cut tentacle was weighed, placed on a cover slip, and imaged for total length and width as well as bead length and width at 0.7× magnification (Olympus model SZX16, Olympus Corporation, Tokyo, Japan). Tentacle cnidae before stinging were imaged from below using an inverted scope at 10× magnification (Olympus model CKX41SF, Olympus Corporation, Tokyo, Japan). Tentacles were then placed on blood agarose and allowed to sting spontaneously for 10 min, with images for stinging length taken at 5 min into the sting duration. Images for length and width measurements as well as cnidae density were taken again post-sting. Adherent cnidae were photographed at 11.5× magnification (Olympus model SZX16, Olympus Corporation, Tokyo, Japan). All tentacle measurements and cnidae counts for density and discharge were determined from microscope images using ImageJ (U.S. National Institutes of Health, Bethesda, MD, USA).
5.3. Tentacle Solution Assay (TSA)
Method 1: Test for induction of discharge: Freshly cut tentacles (length 1–2 cm) were placed on clean, dry microscope slides and examined quickly for discharge; any lengths with notable discharge were discarded. 60 µL of the test solution was then added to the tentacle. After one minute of incubation, a cover slip was gently placed over the tentacle. Preliminary tests with seawater confirmed that coverslip addition did not induce significant discharge.
Method 2: Test for inhibition of chemically induced discharge: tentacles were treated as Method 1, but after 1 min of incubation, the test solution was carefully removed, and 60 μL of 100% ethanol was added. A coverslip was carefully placed after one minute of incubation with ethanol.
Method 3: Test for inhibition of pressure-induced discharge: tentacles were treated as Method 1, but after the 1-minute incubation and addition of the cover slip, the slide-tentacle-slip sandwich was pressed upon by hand for 30 s.
All photos were taken of the tentacles ten minutes after the various treatments through the coverslip using a dissecting microscope at 10× magnification (Olympus model CKX41SF, Olympus Corporation, Tokyo, Japan). Counts of discharged and undischarged cnidae were performed in ImageJ, and statistical analyses were performed using Graphpad Prism ver. 6.0 (GraphPad Software, La Jolla, CA, USA).
5.4. Tentacle Blood Agarose Assay
Effect of solutions and temperature treatments on sting severity was evaluated using a variation of the Tentacle Blood Agarose Assay (TBAA) ex vivo envenomation model outlined in [
14]. Briefly, for
P. utriculus studies, red blood cells were collected from normal human donors (approved protocol CHS#12561, University of Hawaii Committee on Human Studies) and suspended in a low melting point agarose to create an envenomation model where the area of hemolysis serves as a proxy for venom load. The agarose was comprised of 1% RBC and 1.5% Nusieve GTG Agarose (Lonza, Rockland, ME, USA) in modified RPMI (“YRPMI”: 23.81 mM NaHCO
3, Fisher; 102.67 mM NaCl, BDH; 5.37 mM KCl, Fisher; 0.41 mM MgSO
4·7H
2O, Fisher; 25 mM HEPES, Fisher; 6.67 mM NaH
2PO
4, Fisher; 0.42 mM Ca(NO
3)
2·4H
2O, Fisher). For
P. physalis studies, premade sheep’s blood agar plates were used instead (Remel™, Lenexa, KS, USA). The two different media types used reflect the availability of appropriate laboratory space for work with human blood products.
For P. utriculus, blood agarose lengths (75 mm × 25 mm × 2 mm) were placed on glass slides and stung with whole fishing tentacles (~70 mm length) from freshly caught specimens. A 5-slide weight was then added to increase stinging. For rinse solution tests, the weight was removed after 5 min of stinging and 200 µL of the test solution was added to the tentacle; the weight was then replaced and the tentacle was allowed to sting for another 5 min. For temperature tests, the tentacles were allowed to sting for 10 min, with the weight briefly lifted at the 5-minute mark to mimic the rinse solution variant. After a total of 10 min of stinging, the weights and tentacles were removed, and saran-wrapped temperature packs were placed carefully over the slides and a mercury thermometer (Hot packs: Heat Wave XT reusable hand warmer, Bent Grass Concepts LLC, Greenwood Village, CO, USA; Cold packs: CVS Instant Cold Pack, CVS Pharmacy Inc., Woonsocket, RI, USA). For the room temperature replicates, a non-initiated hot pack was placed on the slides. Temperatures under the packs were recorded at 1, 2, 3, 4, 5, 10, 15 and 20 min. All slides were incubated at room temperature overnight, with photos taken at 5 and 24 h after the sting.
For P. physalis, standard 5% sheep’s blood agar plates were stung with 2- to 3-inch sections of fishing tentacles from freshly caught specimens. A glass slide was added as weight to increase stinging. For pretreament tests, tentacles were first incubated in test solutions for 2 min and rinsed twice for 2 min in seawater prior to 5 min of stinging. For temperature tests, the tentacles were allowed to sting for 5 min. After the tentacles were removed, and hot water packs (~45 °C) or ice packs were placed carefully over the plates. All plates were incubated at room temperature overnight, with photos taken at 3, 18 and 24 h after the sting.
Images were recorded using a dissecting microscope (Olympus model SZX16, Olympus Corporation, Tokyo, Japan) or a digital SLR camera (Fujifilm FinePix S4800, Fujifilm, Tokyo, Japan). The area of the zone of hemolysis was calculated using ImageJ. Briefly, scale was set using the known slide width and subsections (50 mm × 20 mm or 50 mm × 15 mm) were taken from each replicate for analysis to remove edge effects. Controls were used to set the color threshold for no hemolysis. The total area of the hemolytic zone was taken directly from the “analyze particles” function. Hemolytic zone was evaluated as the area exhibiting >80% hemolysis. Outliers were defined using the median absolute deviation (MAD) method detailed in [
41], with the level of decision set conservatively at 3; any replicates which were outliers at all time points were removed. Shapiro–Wilk normality tests were conducted on all data sets from the 24-hour timepoint; if the data from 1/3 or more of the treatments was not normally distributed, then Kruskal–Wallis tests were used to compare means. Otherwise, one-way ANOVAs were used. All statistical analyses and post-hoc multiple comparisons were conducted in GraphPad Prism vers. 6.0 (GraphPad Software, Inc., La Jolla, CA, USA).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}