
Detection of a Planktothrix agardhii Bloom in Portuguese Marine Coastal Waters





Abstract
:1. Introduction
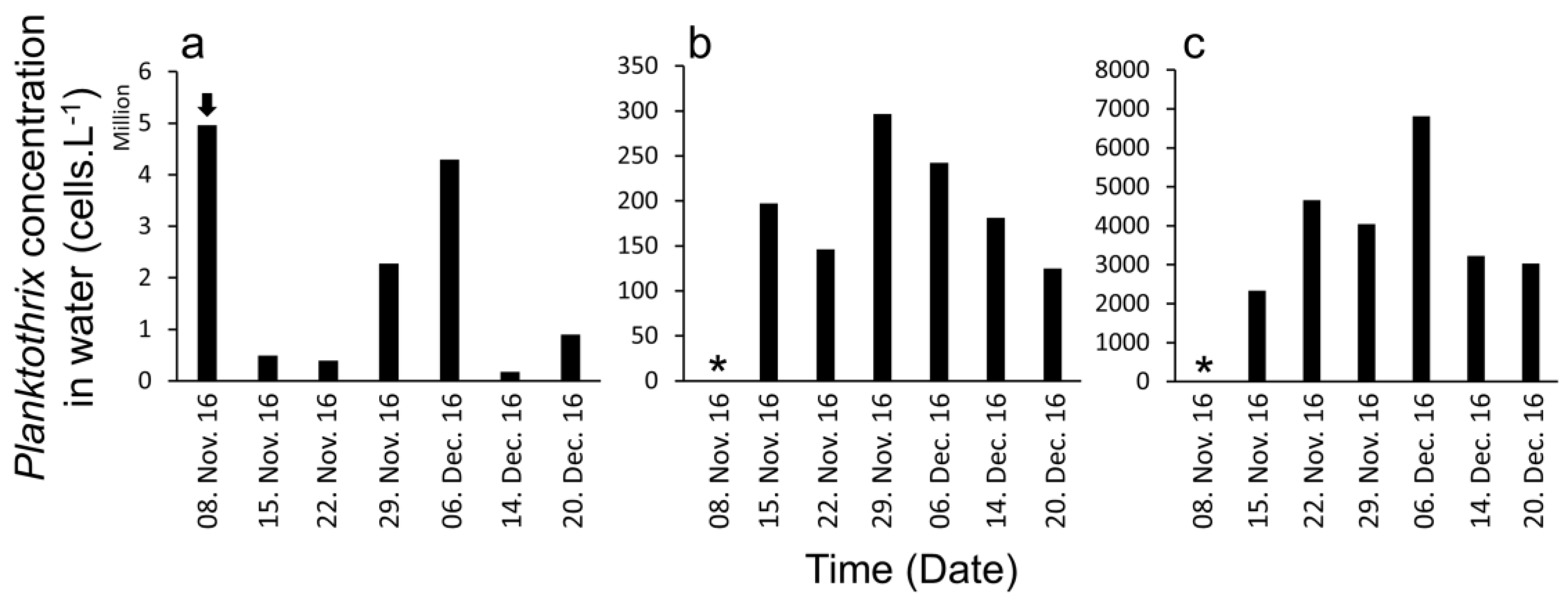
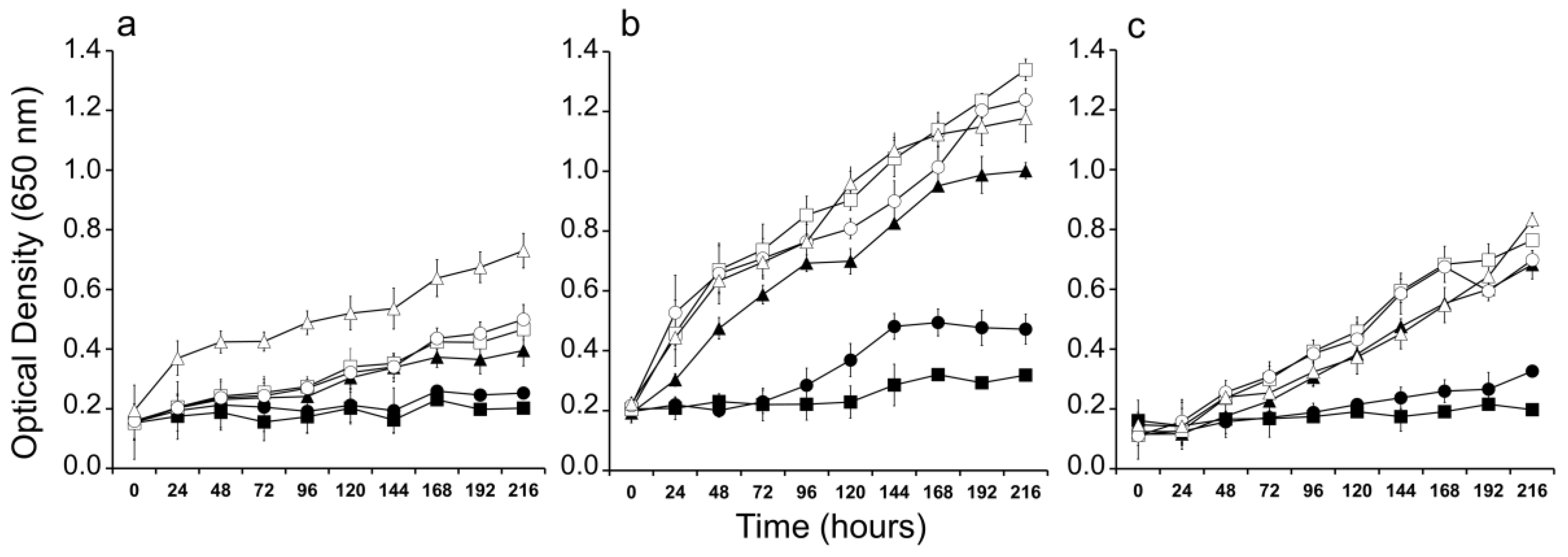
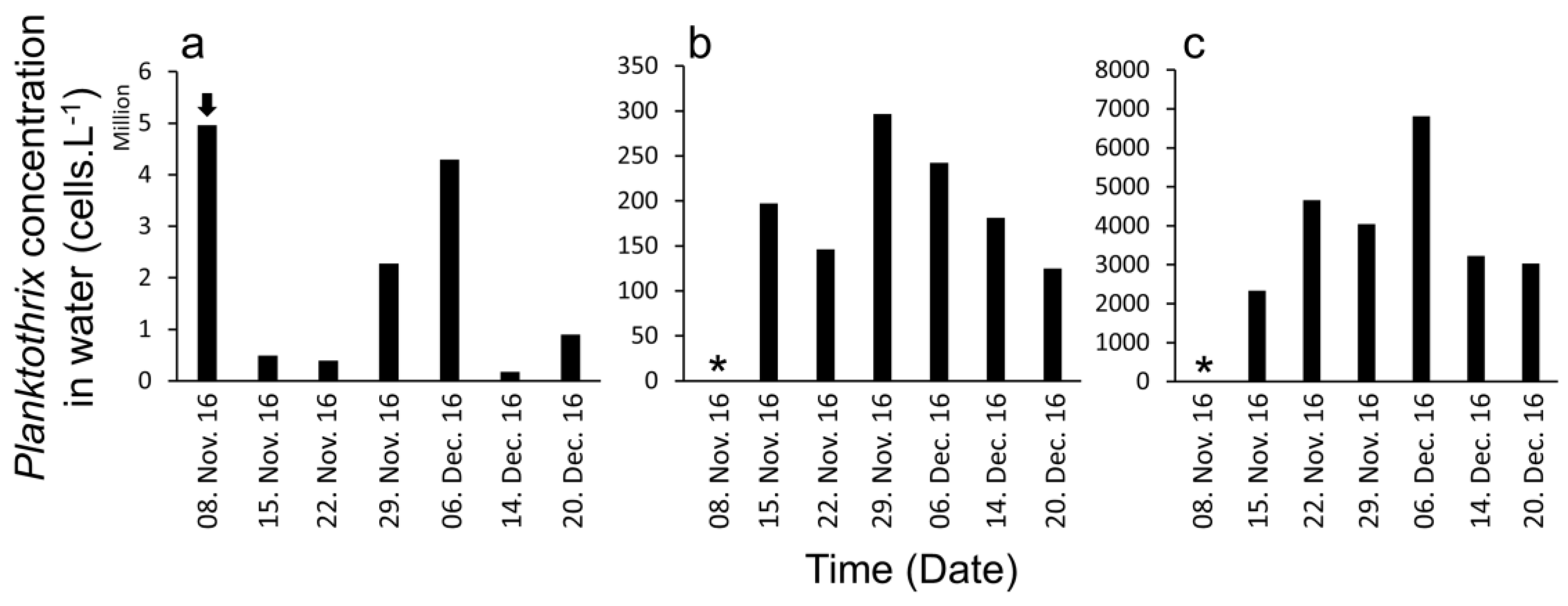
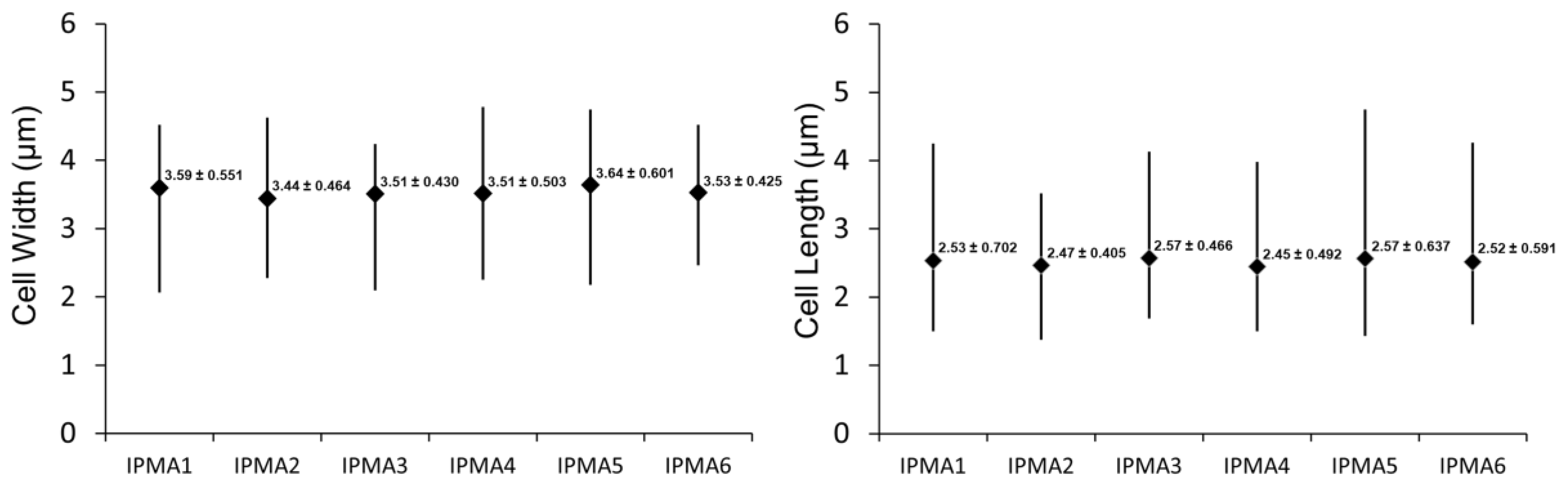
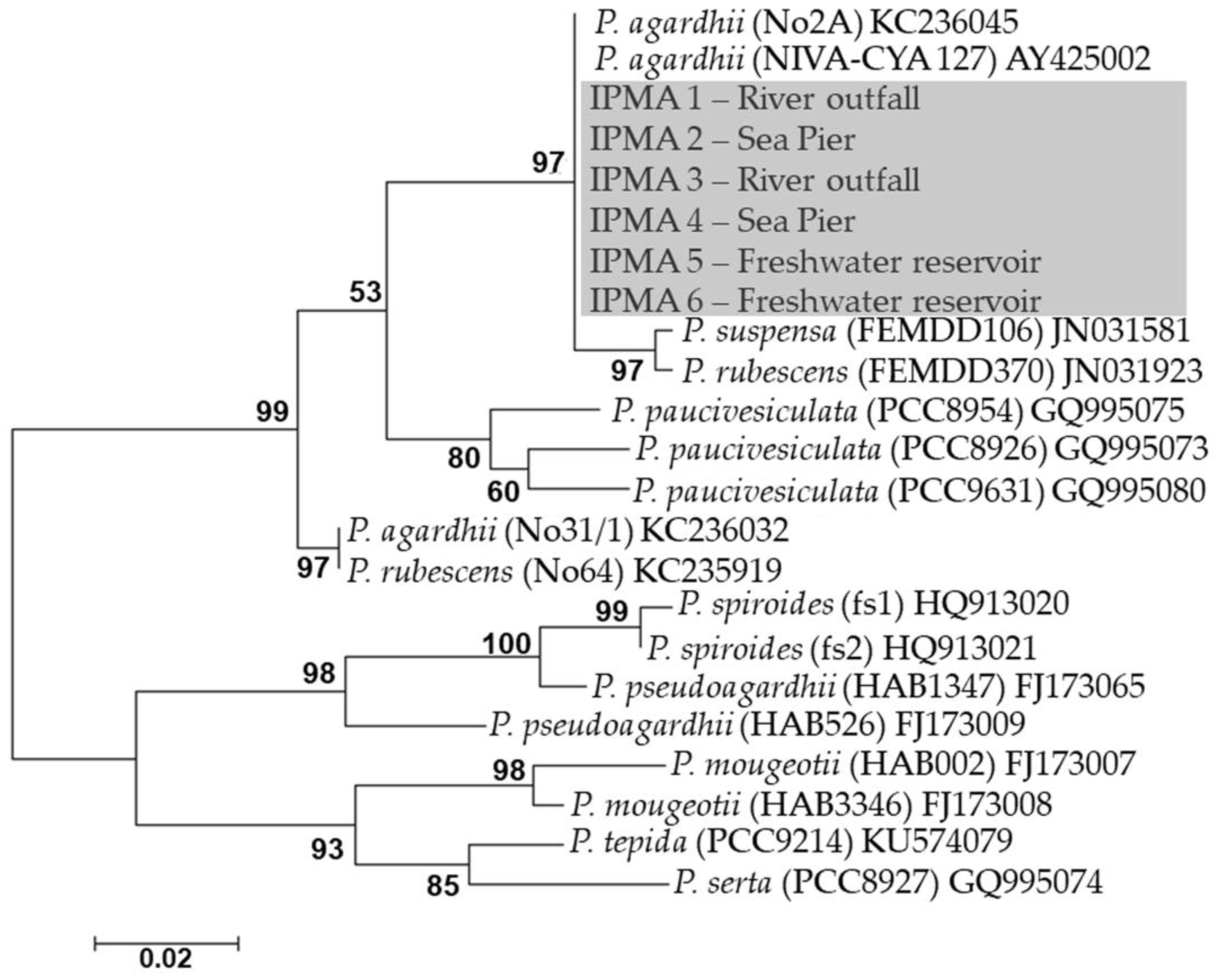
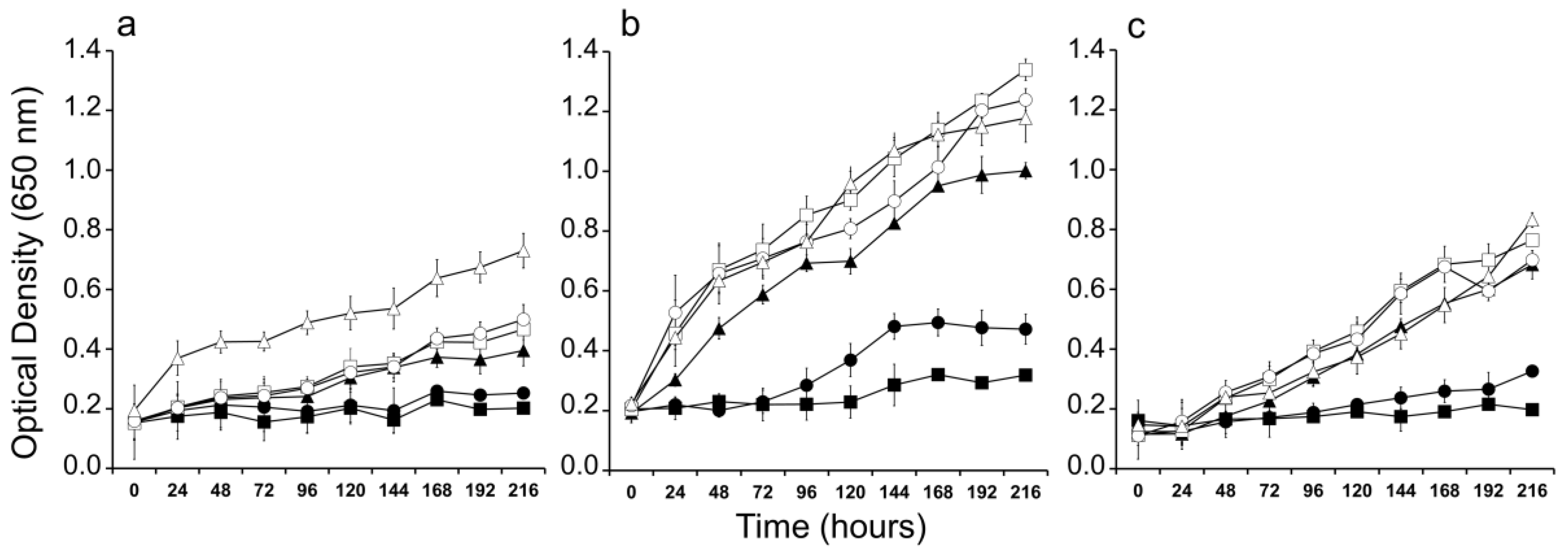
2. Results
3. Discussion
4. Materials and Methods
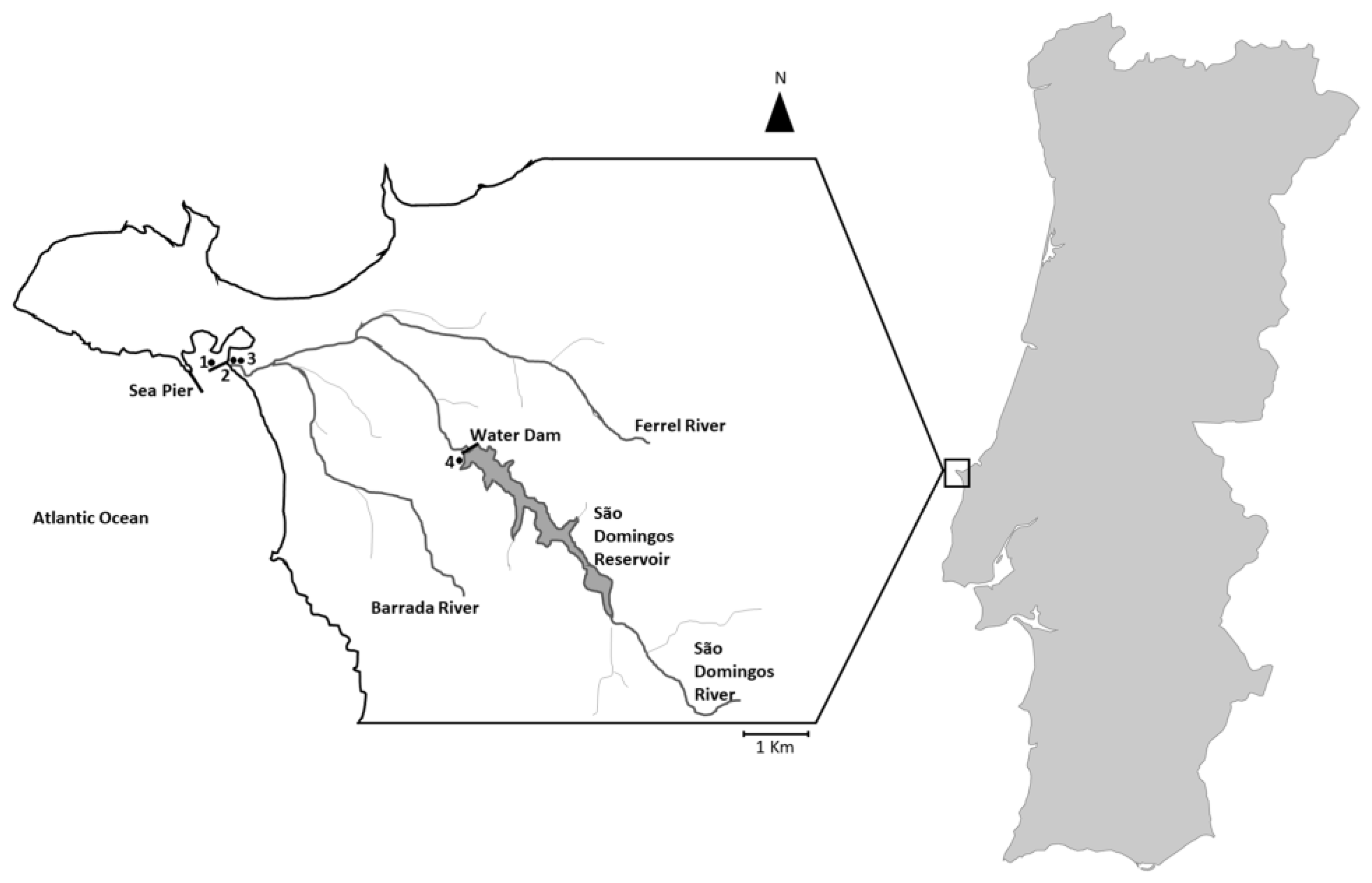
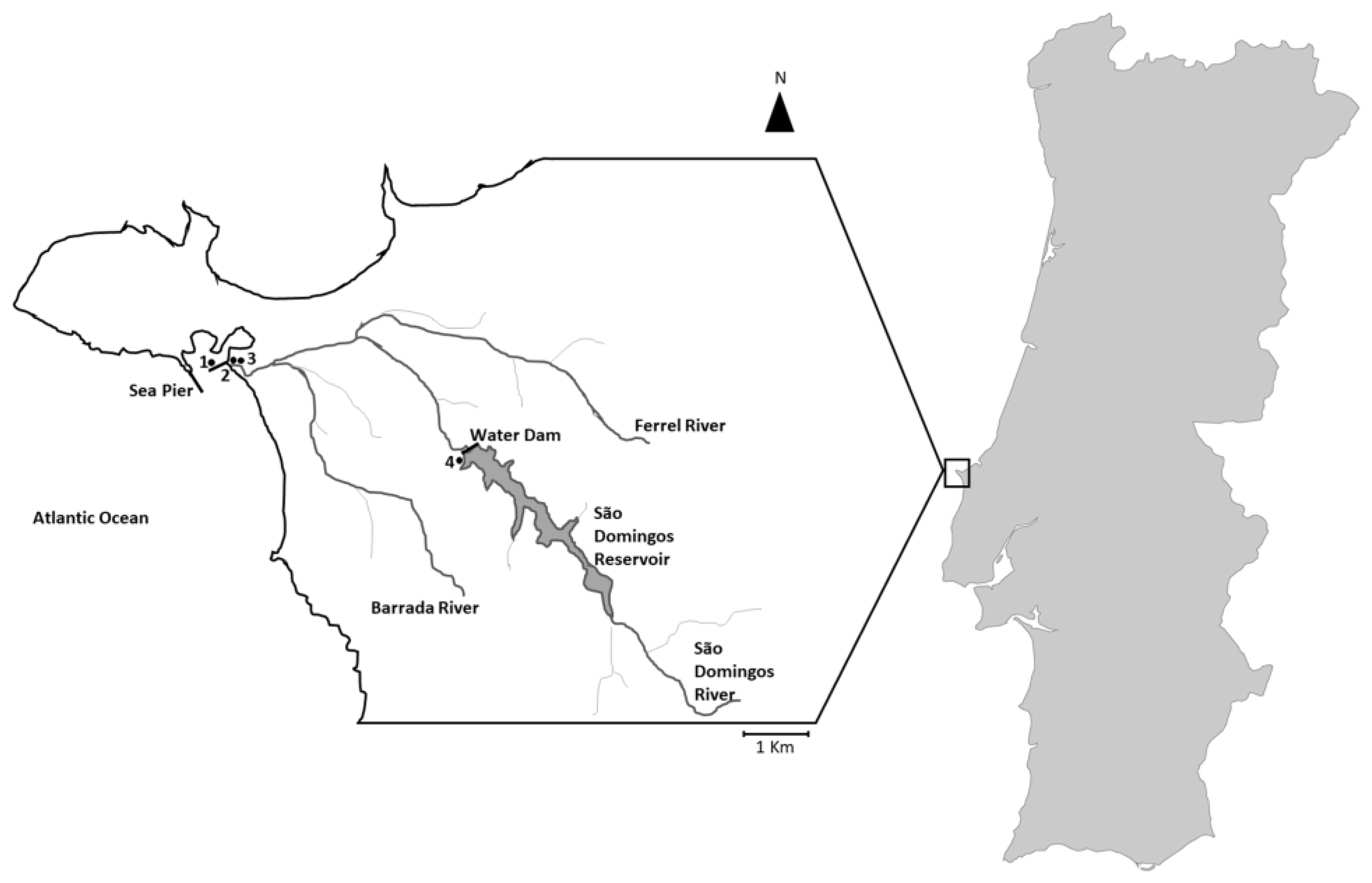
4.1. Location and Sampling
4.2. Filament Isolation and Culture Establishment
4.3. Cyanobacteria Identification and Quantification
4.4. Toxin Analysis
4.5. Salinity Tolerance Experiments
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pouria, S.; Andrade, A.; Barbosa, J.; Cavalcanti, R.; Barreto, V.; Ward, C.; Preiser, W.; Poon, G.; Neild, G.; Codd, G. Fatal Microcystin intoxication in haemodialysis unit in Caruaru, Brazil. Lancet 1998, 325, 21–26. [Google Scholar] [CrossRef]
- Jochimsen, E.; Carmichael, W.; An, J.; Cardo, D.; Cookson, S.; Holmes, C.; Antunes, M.; Melo Filho, D.; Lyra, T.; Barreto, V.; et al. Liver failure and death after exposure to Microcystins at a hemodialysis center in Brazil. N. Engl. J. Med. 1998, 338, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; (World Health Organization) WHO: Geneva, Switzerland, 1999; ISBN 0-419-23930-8. [Google Scholar]
- Buratti, F.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.; Heath, M.; Holland, P.; Munday, R.; McGregor, G.; Ryan, K. Identification of a benthic Microcystin-producing filamentous cyanobacterium (Oscillatoriales) associated with a dog poisoning in New Zealand. Toxicon 2010, 55, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Backer, L.; Manassaram-Baptiste, D.; LePrell, R.; Bolton, B. Cyanobacteria and Algae Blooms: Review of Health and Environmental Data from the Harmful Algal Bloom-Related Illness Surveillance System (HABISS) 2007–2011. Toxins 2015, 7, 1048–1064. [Google Scholar] [CrossRef] [PubMed]
- Vidal, F.; Sedan, D.; D’Agostino, D.; Cavalieri, M.; Mullen, E.; Varela, M.; Flores, C.; Caixach, J.; Andrinolo, D. Recreational Exposure during Algal Bloom in Carrasco Beach, Uruguay: A Liver Failure Case Report. Toxins 2017, 9, 267. [Google Scholar] [CrossRef] [PubMed]
- Pick, F. Blooming algae: A Canadian perspective on the rise of toxic cyanobacteria. Can. J. Fish. Aquat. Sci. 2016, 73, 1149–1158. [Google Scholar] [CrossRef]
- Naselli-Flores, L.; Barone, R.; Chorus, I.; Kurmayer, R. Toxic cyanobacterial blooms in reservoirs under a semiarid Mediterranean climate: The magnification of a problem. Environ. Toxicol. 2007, 22, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Gkelis, S.; Papadimitriou, T.; Zaoutsos, N.; Leonardos, I. Anthropogenic and climate-induced change favors toxic cyanobacteria blooms: Evidence from monitoring a highly eutrophic, urban Mediterranean lake. Harmful Algae 2014, 39, 322–333. [Google Scholar] [CrossRef]
- Pearl, H.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, J.; Davis, T.; Burford, M.; Gobler, C. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Pearl, H.; Paul, V. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2011, 46, 1349–1363. [Google Scholar] [CrossRef] [PubMed]
- Posch, T.; Köster, O.; Salcher, M.; Pernthaler, J. Harmful filamentous cyanobacteria favoured by reduced water turnover with lake warming. Nat. Clim. Chang. 2012, 2, 809–813. [Google Scholar] [CrossRef]
- Sandrini, G.; Ji, X.; Verspagen, J.; Tann, R.; Slot, P.; Luimstra, M.; Schuurmans, M.; Matthijs, H.; Huisman, J. Rapid adaptation of harmful cyanobacteria to rising CO2. Proc. Natl. Acad. Sci. USA 2016, 113, 9315–9320. [Google Scholar] [CrossRef] [PubMed]
- Visser, P.; Verspagen, J.; Sandrini, G.; Stal, L.; Matthijs, H.; Davis, T.; Paerl, H.; Huisman, J. How rising CO2 and global warming may stimulate harmful cyanobacterial blooms. Harmful Algae 2016, 54, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Park, S.B. Algal blooms hit South Korean rivers. Nat. News 2012. [Google Scholar] [CrossRef]
- Ryu, H.; Shin, R.; Lee, J. Morphology and taxonomy of the Aphanizomenon spp. (Cyanophyceae) and related species in the Nakdong River. South Korea J. Ecol. Environ. 2016, 41, 6. [Google Scholar] [CrossRef]
- Gibble, C.; Kudela, R. Detection of persistent Microcystin toxins at the land-sea interface in Monterey Bay, California. Harmful Algae 2014, 39, 146–153. [Google Scholar] [CrossRef]
- Preece, E.; Moore, B.; Hardy, J. Transfer of Microcystin from freshwater lakes to Puget Sound, WA and toxin accumulation in marine mussels (Mytilus trossulus). Ecotoxicol. Environ. Saf. 2015, 122, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Gibble, C.; Peacock, M.; Kudela, R. Evidence of freshwater algal toxins in marine shellfish: Implications for human and aquatic health. Harmful Algae 2016, 59, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Preece, E.; Hardy, J.; Moore, B.; Bryan, M. A review of Microcystin detections in Estuarine and Marine waters: Environmental implications and human health risk. Harmful Algae 2017, 61, 31–45. [Google Scholar] [CrossRef]
- Ibelings, B.; Backer, L.; Kardinaal, E.; Chorus, I. Current approaches to cyanotoxin risk assessment and risk management around the globe. Harmful Algae 2015, 49, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.; Sinha, R.; Incharoensakdi, A. The cyanotoxin-Microcystins: Current overview. Rev. Environ. Sci. Biotechnol. 2014, 13, 215–249. [Google Scholar] [CrossRef]
- Takahashi, T.; Umehara, A.; Tsutsumi, H. Diffusion of Microcystins (cyanobacteria hepatotoxins) from the reservoir of Isahaya Bay, Japan, into the marine and surrounding ecosystems as a result of large-scale drainage. Mar. Pollut. Bull. 2014, 89, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Testai, E.; Buratti, F.; Funari, E.; Manganelli, M.; Vichi, S.; Arnich, N.; Biré, R.; Fessard, V.; Sialehaamoa, A. Review and analysis of occurrence, exposure and toxicity of cyanobacteria toxins in food. Eur. Food Saf. Auth. Support. Publ. 2016, 13. [Google Scholar] [CrossRef]
- Chorus, I. Current Approaches to Cyanotoxin Risk Assessment, Risk Management and Regulations in Different Countries; Federal Environment Agency (Umweltbundesamt): Dessau-Roßlau, Germany, 2012; ISSN 1862-4804. [Google Scholar]
- Decreto-Lei n.o 216/1998 de 18 de Setembro, Diário da República, Série I-A, n.o 216. Available online: http://data.dre.pt/eli/dec-lei/293/1998/09/18/p/dre/pt/html (accessed on 30 November 2017).
- Council Directive 91/493/CEE of 22 of July 1991, Adopting the Health Rules for the Production and the Placing on the Market of Fishery Products. Available online: http://data.europa.eu/eli/dir/1991/493/oj (accessed on 30 November 2017).
- Decreto-Lei n.o 306/2007 de 27 de Agosto, Diário da República, Série I, n.o 164. Available online: https://dre.pt/application/dir/pdf1sdip/2007/08/16400/0574705765.pdf (accessed on 30 November 2017).
- Council Directive 98/83/EC of 3 of November 1998 on the Quality of Water Intended for Human Consumption. Available online: http://eur-lex.europa.eu/legalcontent/EN/TXT/PDF/?uri=CELEX:31998L0083&from=PT (accessed on 30 November 2017).
- Decreto-Lei n.o 135/2009 de 3 de Junho, Diário da República, Série I, n.o 107. Available online: http://snirh.pt/snirh/_divulgacao/legislacao/site/verlei.php?id=20646 (accessed on 30 November 2017).
- Decreto-Lei n.o 113/2012 de 23 de Maio, Diário da República, Série I, n.o 100. Available online: http://publicos.pt/documento/id177865/decreto-lei-113/2012 (accessed on 30 November 2017).
- Council Directive 2006/7/EC of 15 February 2006 Concerning the Management of Bathing Water Quality and Repealing Directive 76/160/EEC. Available online: http://eurlex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2006:064:0037:0051:EN:PDF (accessed on 30 November 2017).
- Suda, S.; Watanabe, M.; Otsuka, S.; Mahakahant, A.; Yongmanitchai, W.; Nopartnaraporn, N.; Liu, Y.; Day, J. Taxonomic revision of water-bloom-forming species of oscillatorioid cyanobacteria. Int. J. Syst. Evol. Microbiol. 2002, 52, 1577–1595. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Kudela, R.; Mekebri, A.; Crane, D.; Oates, S.; Tinker, M.; Staedler, W.; Miller, W.; Toy-Choutka, W.; Dominik, S.; et al. Evidence for a Novel Marine Harmful Algal Bloom: Cyanotoxin (Microcystin) Transfer from Land to Sea Otters. PLoS ONE 2010, 5, e12576. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, V.; Cerqueira, M. Phytoplankton community of river Minho (international section). Limnetica 2001, 20, 135–141. [Google Scholar]
- Sobrino, C.; Matthiensen, A.; Vidal, S.; Galvão, H. Occurrence of Microcystins along the Guadiana estuary. Limnetica 2004, 23, 133–144. [Google Scholar]
- Rocha, C.; Galvão, H.; Barbosa, A. Role of transient silicon limitation in the development of cyanobacteria blooms in the Guadiana estuary, south-western Iberia. Mar. Ecol. Prog. Ser. 2002, 228, 35–45. [Google Scholar] [CrossRef]
- Mulvenna, V.; Dale, K.; Priestly, B.; Mueller, U.; Humpage, A.; Shaw, G.; Allinson, G.; Falconer, I. Health Risk Assessment for Cyanobacterial Toxins in Seafood. Int. J. Environ. Res. Public Health 2012, 9, 807–820. [Google Scholar] [CrossRef] [PubMed]
- Rücker, J.; Wiedner, C.; Zippel, P. Factors controlling the dominance of Planktothrix agardhii and Limnothrix redekei in eutrophic shallow lakes. Hydrobiologia 1997, 342–343, 107–115. [Google Scholar] [CrossRef]
- Hasler, P.; Poulícková, A. Diurnal changes in vertical distribution and morphology of a natural population of Planktothrix agardhii (Gom.) Anagnostidis et Komárek (Cyanobacteria). Hydrobiologia 2003, 506–509, 195–201. [Google Scholar] [CrossRef]
- Halstvedt, C.; Rohrlack, T.; Andersen, T.; Skulberg, O.; Edvardsen, B. Seasonal dynamics and depth distribution of Planktothrix spp. in Lake Steinsfjorden (Norway) related to environmental factors. J. Plankton Res. 2007, 29, 471–482. [Google Scholar] [CrossRef]
- Pawlik-Skowroμska, B.; Pirszel, J.; Kornijów, R. Spatial and temporal variation in Microcystin concentrations during perennial bloom of Planktothrix agardhii in a hypertrophic lake. Ann. Limnol. Int. J. Limnol. 2008, 44, 145–150. [Google Scholar] [CrossRef]
- Bonilla, S.; Aubriot, L.; Soares, M.; Gonzales-Piana, M.; Fabre, A.; Huszar, V.; Lürling, M.; Antoniades, D.; Padisák, J.; Kruk, C. What drives the distribution of the bloom forming cyanobacteria Planktothrix agardhii and Cylindrospermopsis raciborskii? FEMS Microbiol. Ecol. 2012, 79, 594–607. [Google Scholar] [CrossRef] [PubMed]
- Gaget, V.; Welker, M.; Rippka, R.; de Marsac, N. A polyphasic approach leading to the revision of the genus Planktothrix (Cyanobacteria) and its type species, P. agardhii, and proposal for integrating the emended valid botanical taxa, as well as three new species, Planktothrix paucivesiculata sp. nov. ICNP, Planktothrix tepida sp. nov. ICNP, and Planktothrix serta sp. nov. ICNP, as genus and species names with nomenclatural standing under the ICNP. Syst. Appl. Microbiol. 2015, 38, 141–158. [Google Scholar] [CrossRef] [PubMed]
- Vergalli, J.; Bertrand, C.; Borla, A.; Franquet, E.; Fayolle, S. Salt tolerance of Planktothrix agardhii (Gomont) Anagnostidis & Komárek (cyanoprokaryota) between freshwater and brackish strains in batch cultures. Algol. Stud. 2016, 150, 3–19. [Google Scholar] [CrossRef]
- Chomérat, N.; Garnier, R.; Bertrand, C.; Cazaubon, A. Seasonal succession of cyanoprokaryotes in a hypereutrophic oligomesohaline lagoon from the South of France, Estuarine. Coast. Shelf Sci. 2007, 72, 591–602. [Google Scholar] [CrossRef]
- Romo, S.; Fernández, F.; Ouahid, Y.; Barón-Sola, A. Assessment of Microcystins in lake water and fish (Mugilidae, Liza sp.) in the largest Spanish coastal lake. Environ. Monit. Assess. 2012, 184, 939–949. [Google Scholar] [CrossRef] [PubMed]
- De Pace, R.; Valeria, V.; Bucci, M.; Pasquale, G.; Milena, B. Microcystin Contamination in Sea Mussel Farms from the Italian Southern Adriatic Coast following Cyanobacterial Blooms in an Artificial Reservoir. J. Ecosyst. 2014. [Google Scholar] [CrossRef]
- IPMA. Bivalves—Biotoxinas. Available online: https://www.ipma.pt/en/pescas/bivalves/index.jsp (accessed on 23 September 2017).
- Staub, R. Ernährungsphysiologisch-autökologische Untersuchungen an Oscillatoria rubescens D.C. Schweiz. Z. Hydrol. 1961, 23, 82–198. [Google Scholar]
- EN15204. Water Quality. Guidance Standard on the Enumeration of Phytoplankton Using Inverted Microscopy (Utermöhl Technique); British Standards Institution: London, UK, 2006. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota-2. Teil: Oscillatoriales; Elsevier GmbH: Heidelberg, Germany, 2005; pp. 354–363. ISBN 978-3-8274-1914-9. [Google Scholar]
- Komárek, J.; Komárková, J. Taxonomic review of the cyanoprokaryotic genera Planktothrix and Planktothricoides. Czech Phycol. 2004, Olomouc 4, 1–18. [Google Scholar]
- Lin, S.; Wua, Z.; Yu, G.; Zhu, M.; Yu, B.; Li, R. Genetic diversity and molecular phylogeny of Planktothrix (Oscillatoriales, cyanobacteria) strains from China. Harmful Algae 2010, 9, 87–97. [Google Scholar] [CrossRef]
- Edgar, R. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, D36–D42. [Google Scholar] [CrossRef] [PubMed]
- Briand, E.; Gugger, M.; Francois, J.; Bernard, C.; Humbert, J.; Quiblier, C. Temporal variations in the dynamics of potentially Microcystin-producing strains in a bloom-forming Planktothrix agardhii (cyanobacterium) population. Appl. Environ. Microbiol. 2008, 74, 3839–3848. [Google Scholar] [CrossRef] [PubMed]
- Stepánkováa, T.; Ambrozováa, L.; Bláhaa, L.; Giesy, J.; Hilscherováa, K. In vitro modulation of intracellular receptor signalling and cytotoxicity induced by extracts of cyanobacteria, complex water blooms and their fractions. Aquat. Toxicol. 2011, 105, 497–507. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Number | Origin | Date of Isolation |
|---|---|---|
| IPMA1 | River outfall | 29 November 2016 |
| IPMA2 | Sea Pier | 8 November 2016 |
| IPMA3 | River outfall | 29 November 2016 |
| IPMA4 | Sea Pier | 8 November 2016 |
| IPMA5 | S. Domingos reservoir | 15 November 2016 |
| IPMA6 | S. Domingos reservoir | 15 November 2016 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Churro, C.; Azevedo, J.; Vasconcelos, V.; Silva, A. Detection of a Planktothrix agardhii Bloom in Portuguese Marine Coastal Waters. Toxins 2017, 9, 391. https://doi.org/10.3390/toxins9120391
Churro C, Azevedo J, Vasconcelos V, Silva A. Detection of a Planktothrix agardhii Bloom in Portuguese Marine Coastal Waters. Toxins. 2017; 9(12):391. https://doi.org/10.3390/toxins9120391
Chicago/Turabian StyleChurro, Catarina, Joana Azevedo, Vitor Vasconcelos, and Alexandra Silva. 2017. "Detection of a Planktothrix agardhii Bloom in Portuguese Marine Coastal Waters" Toxins 9, no. 12: 391. https://doi.org/10.3390/toxins9120391