Highly Toxic Ribbon Worm Cephalothrix simula Containing Tetrodotoxin in Hiroshima Bay, Hiroshima Prefecture, Japan

Abstract
:1. Introduction
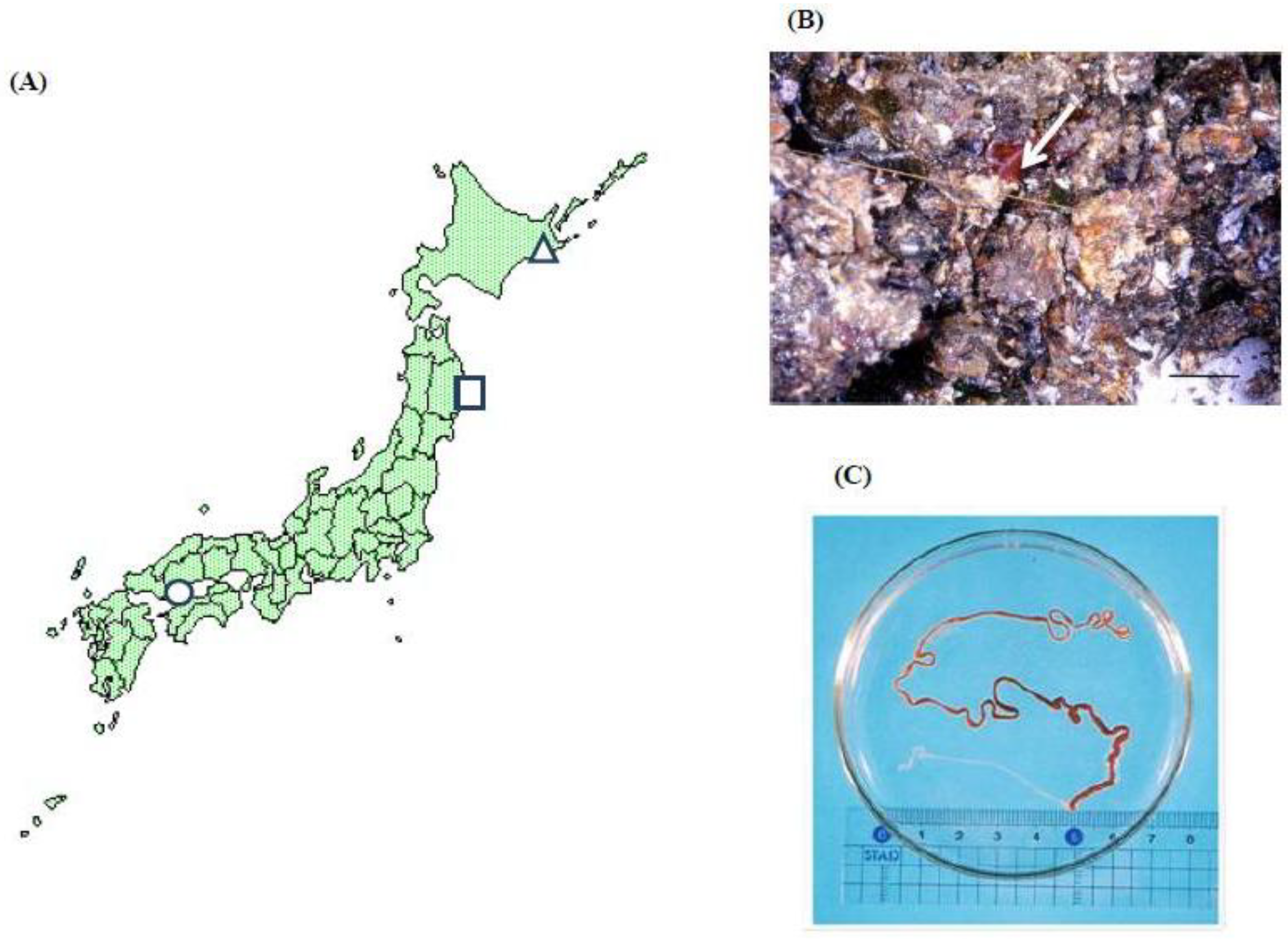
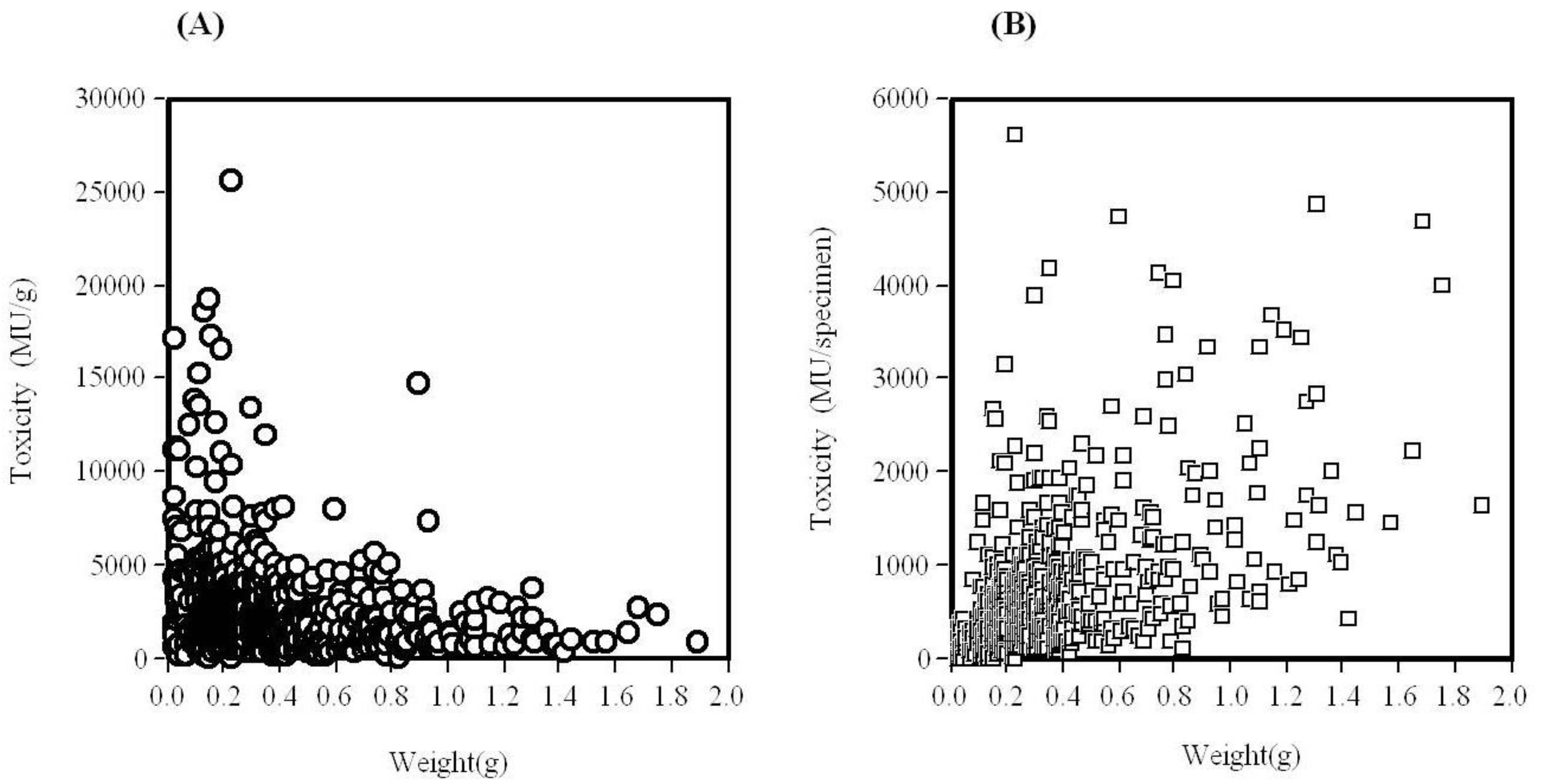
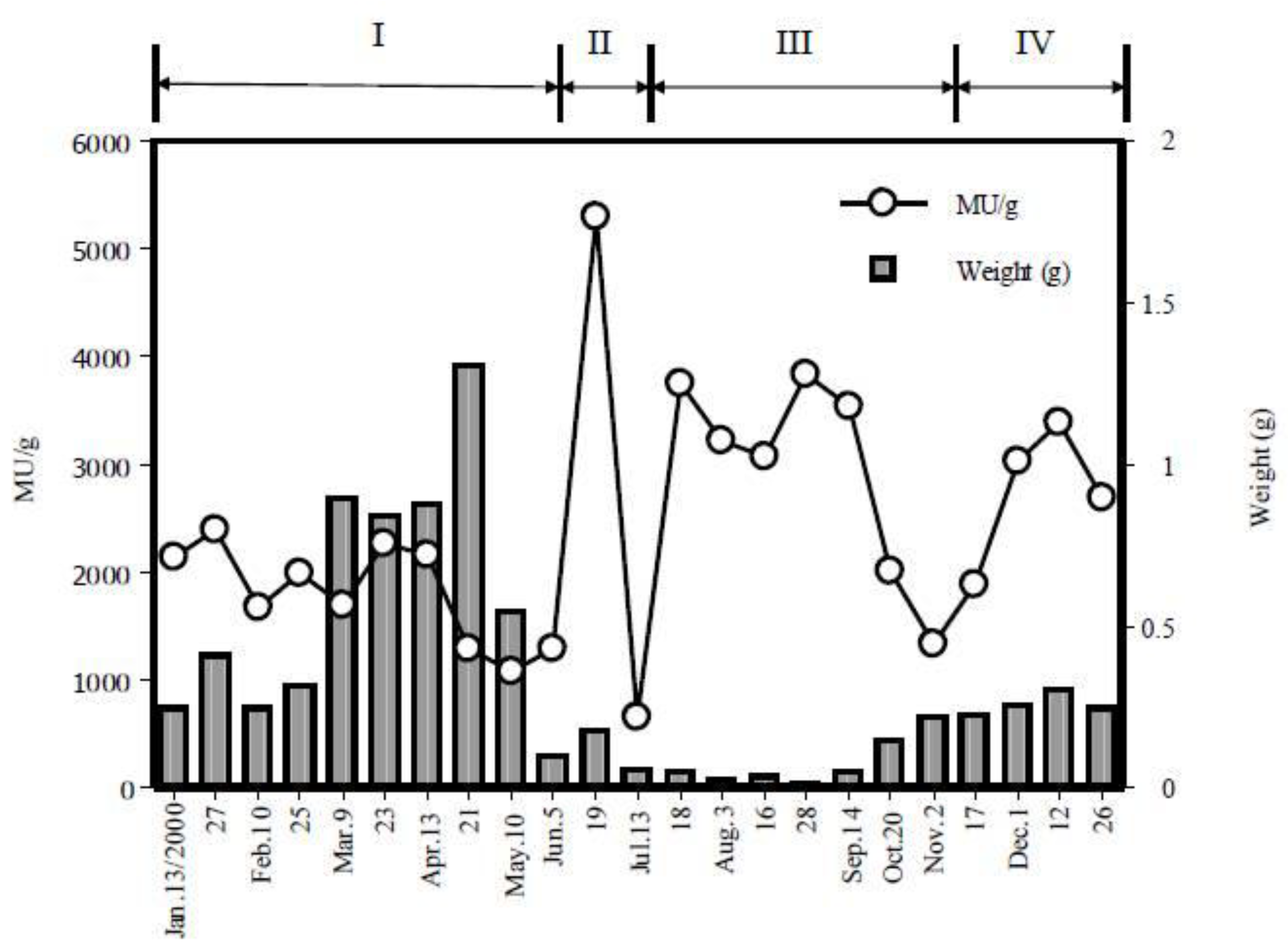
2. Toxicological Surveillance of Ribbon Worms in Hiroshima Bay

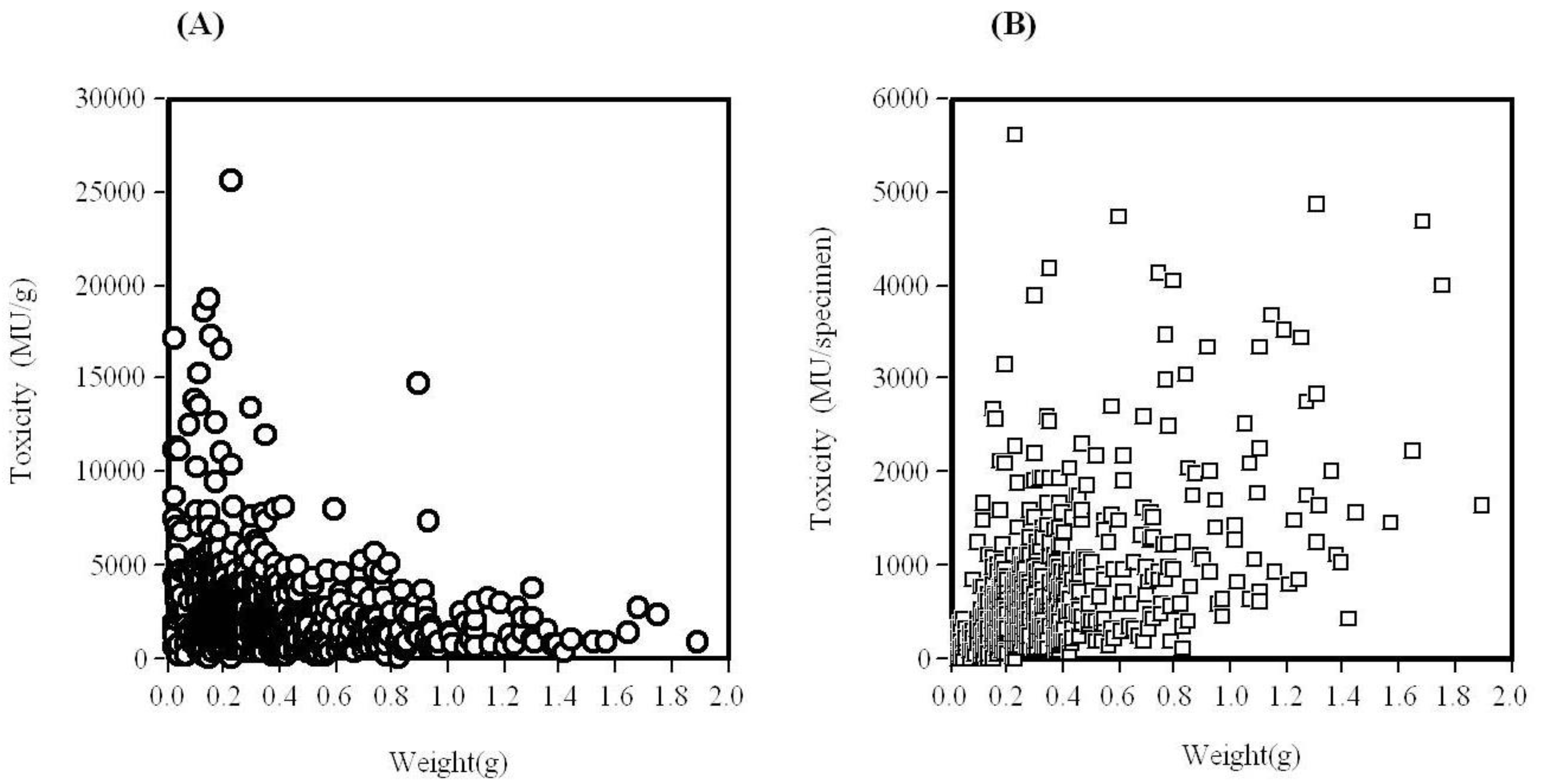
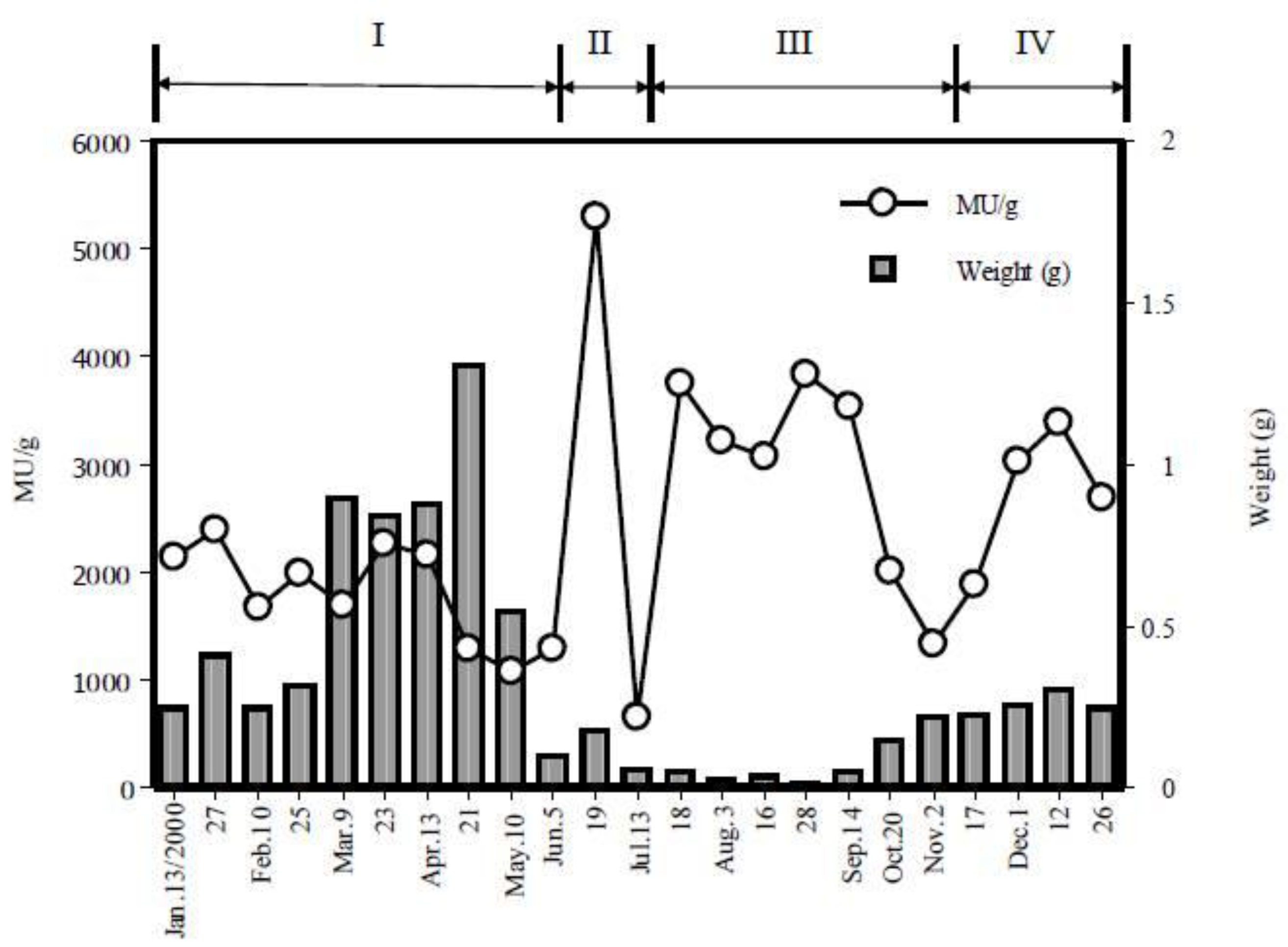
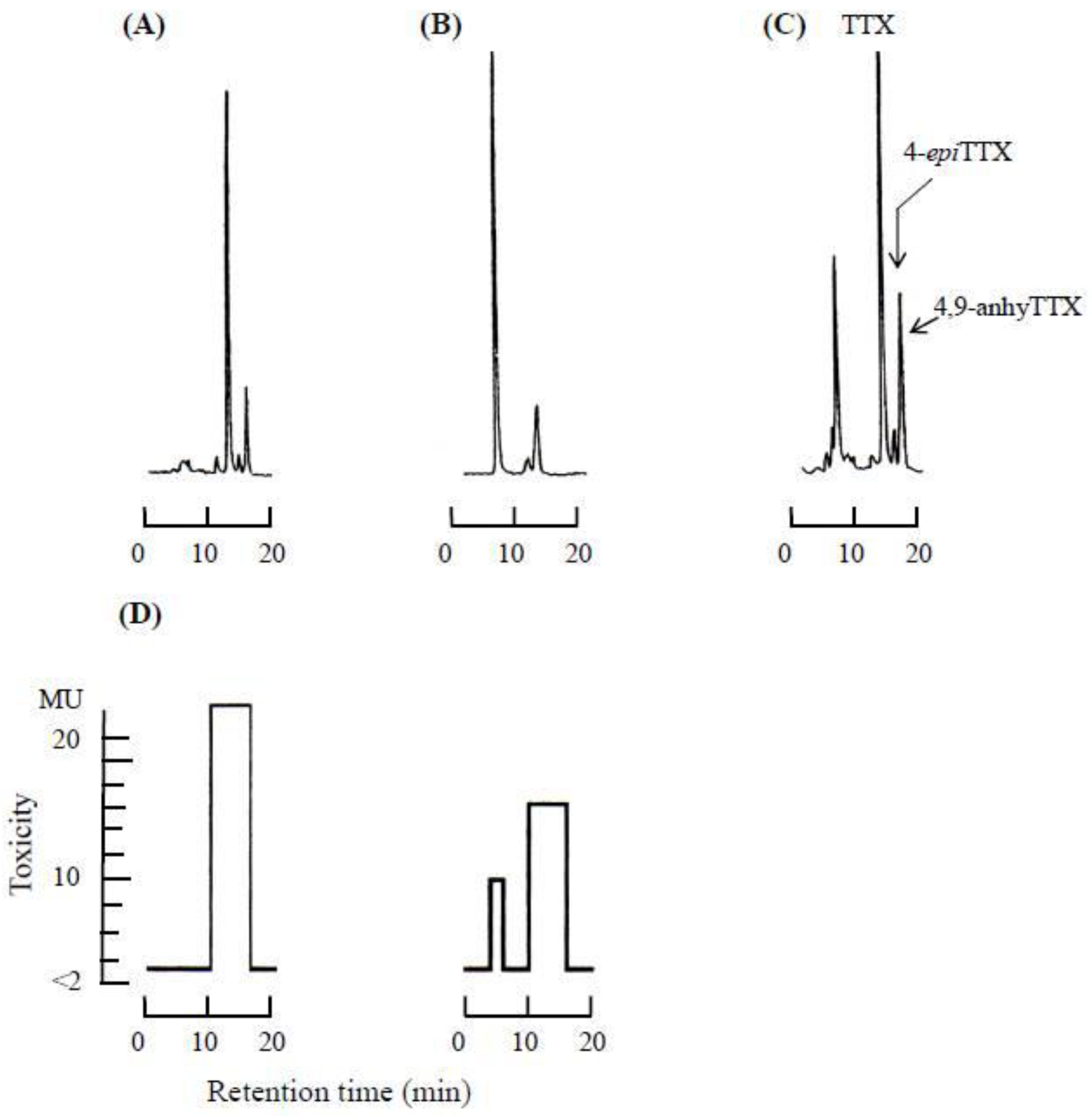
3. Isolation of Main Toxic Component
3.1. Mouse Bioassay for Lethal Potency
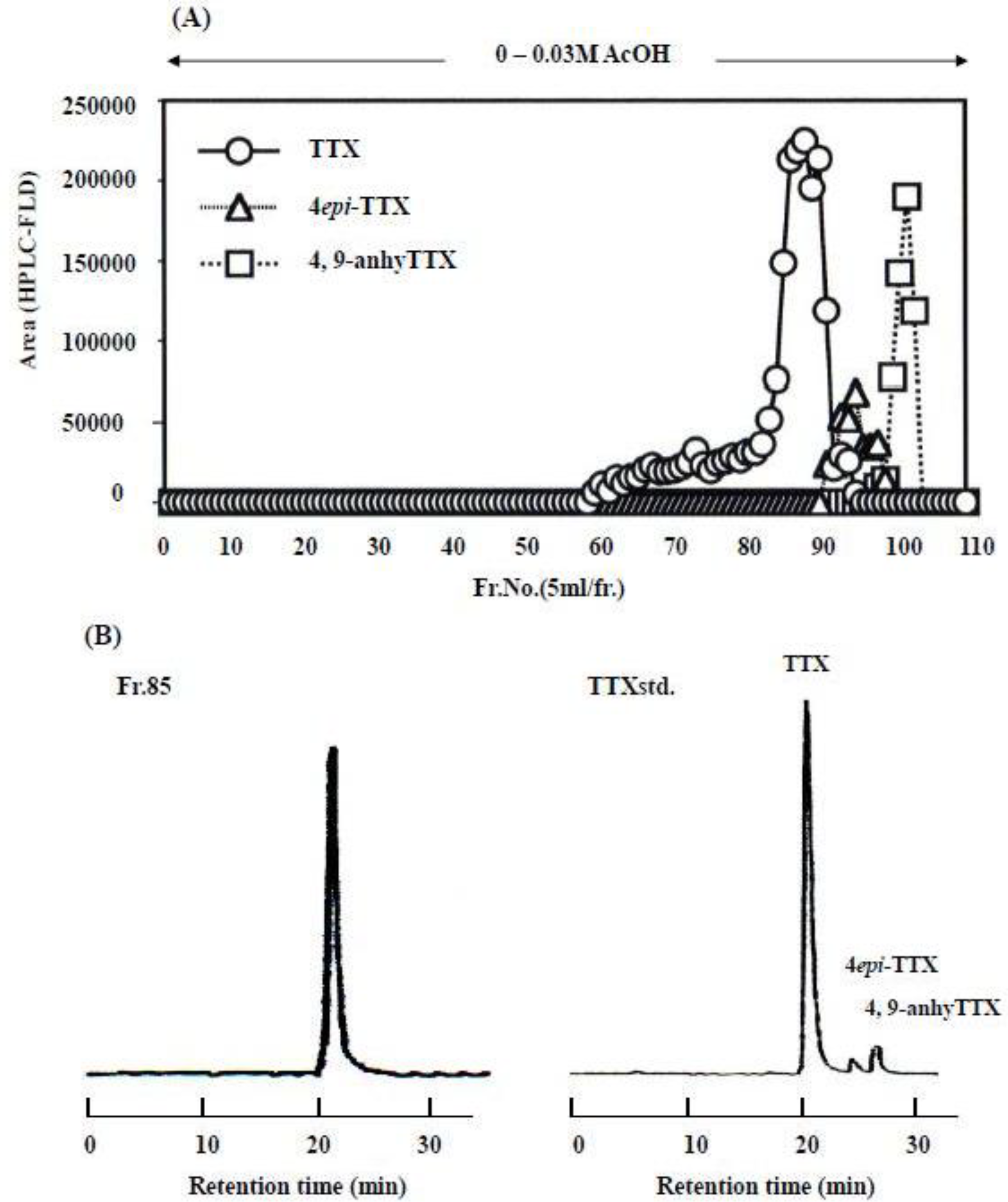
3.2. Column Chromatography


3.3. Instrumental Analysis
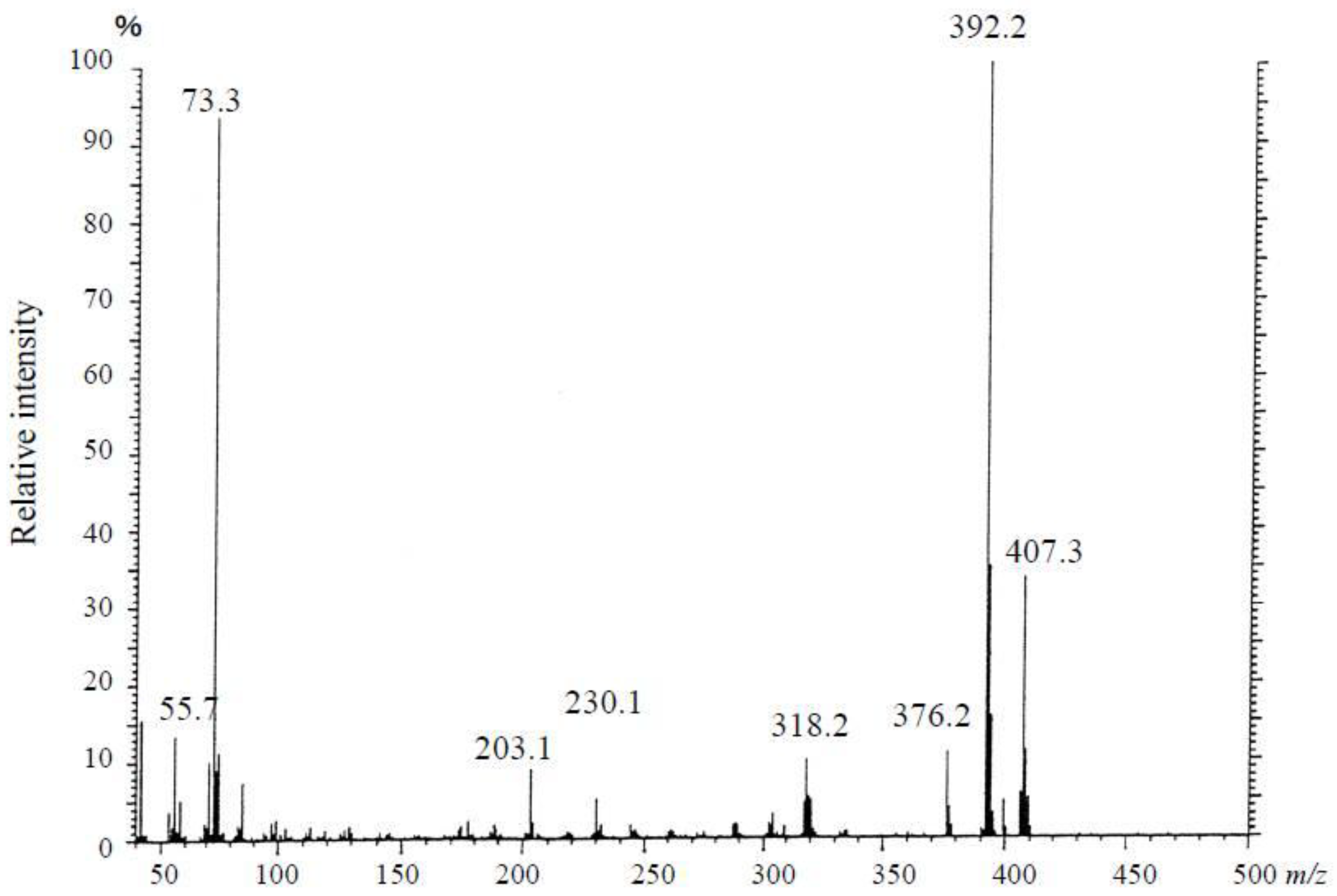
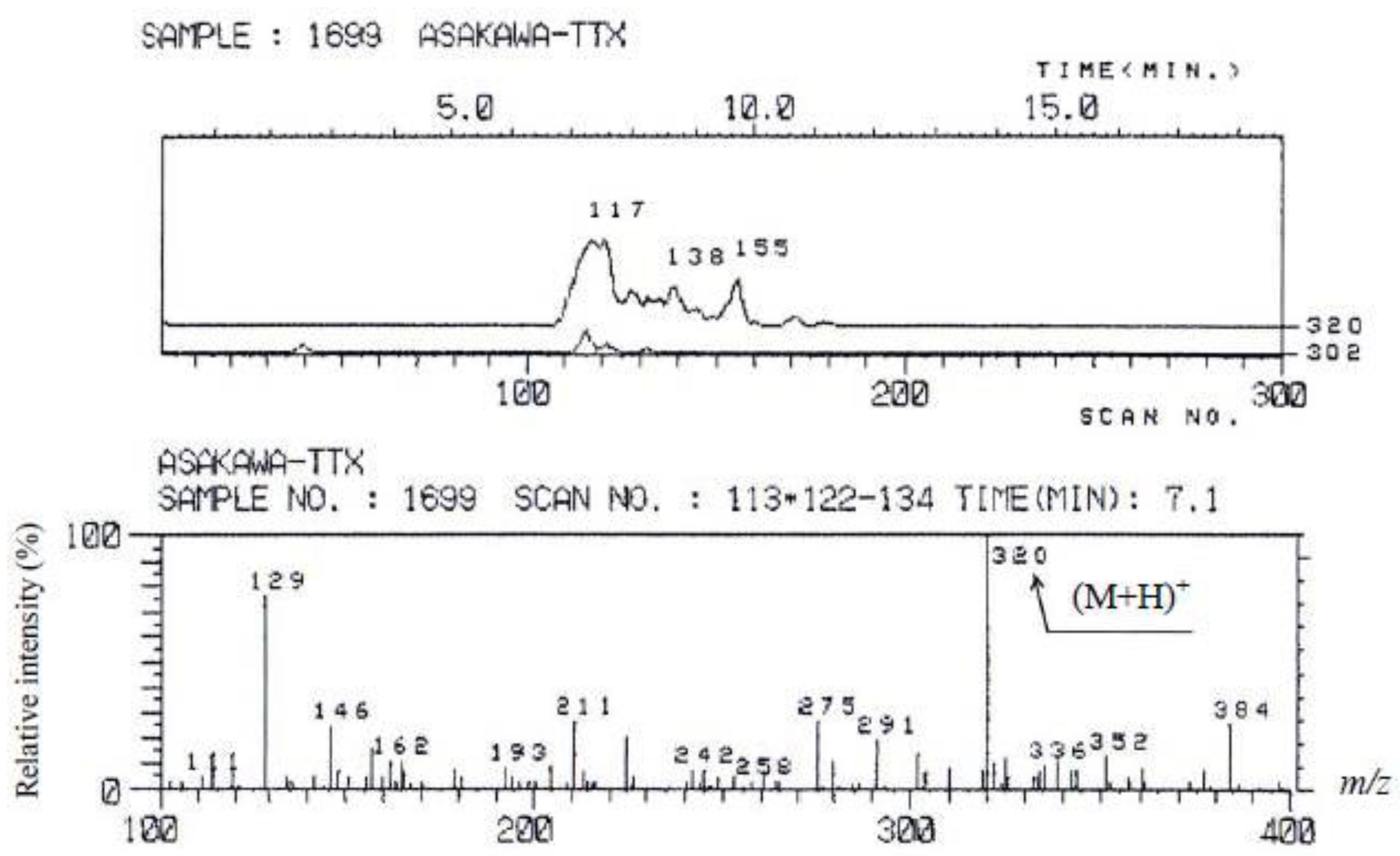
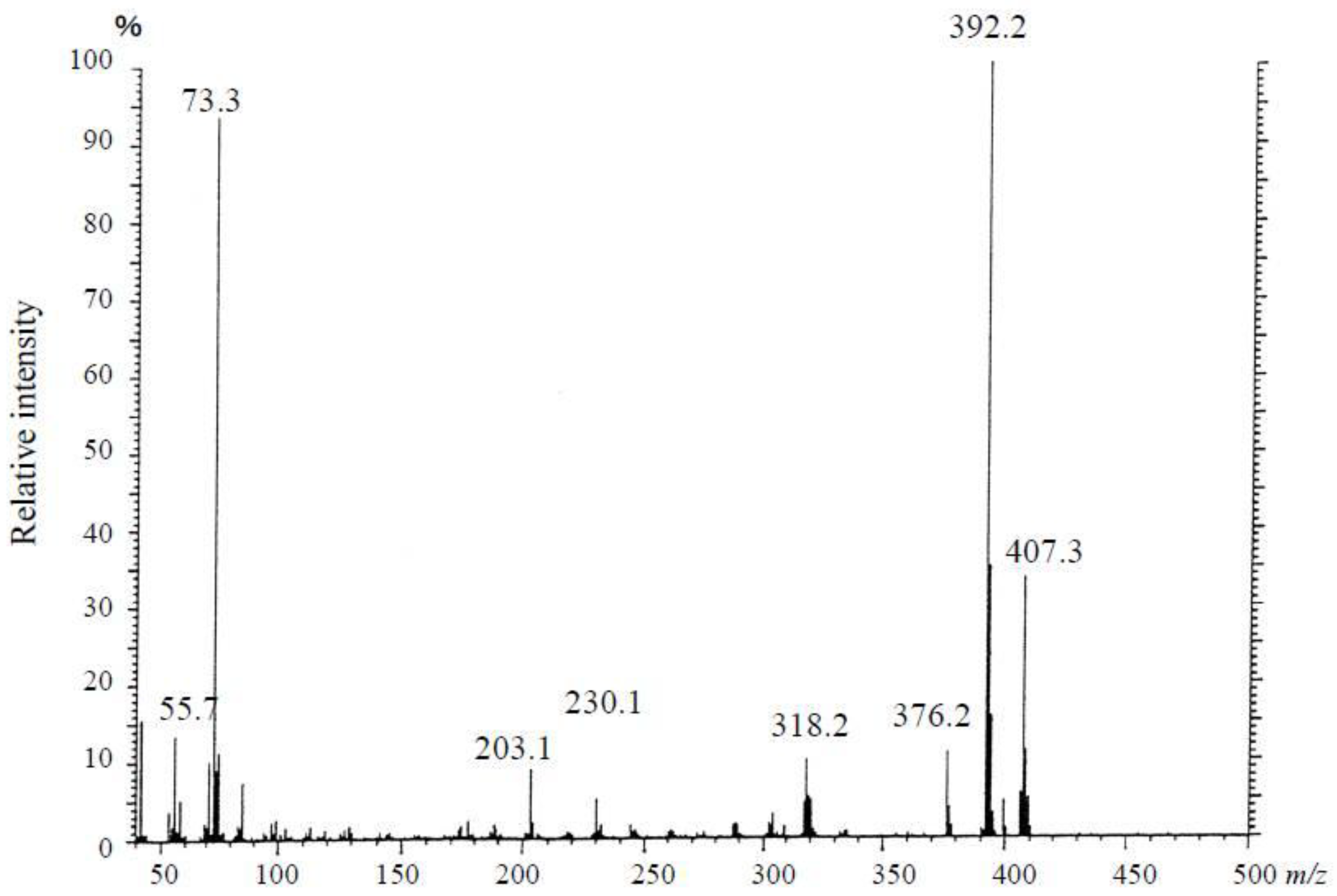
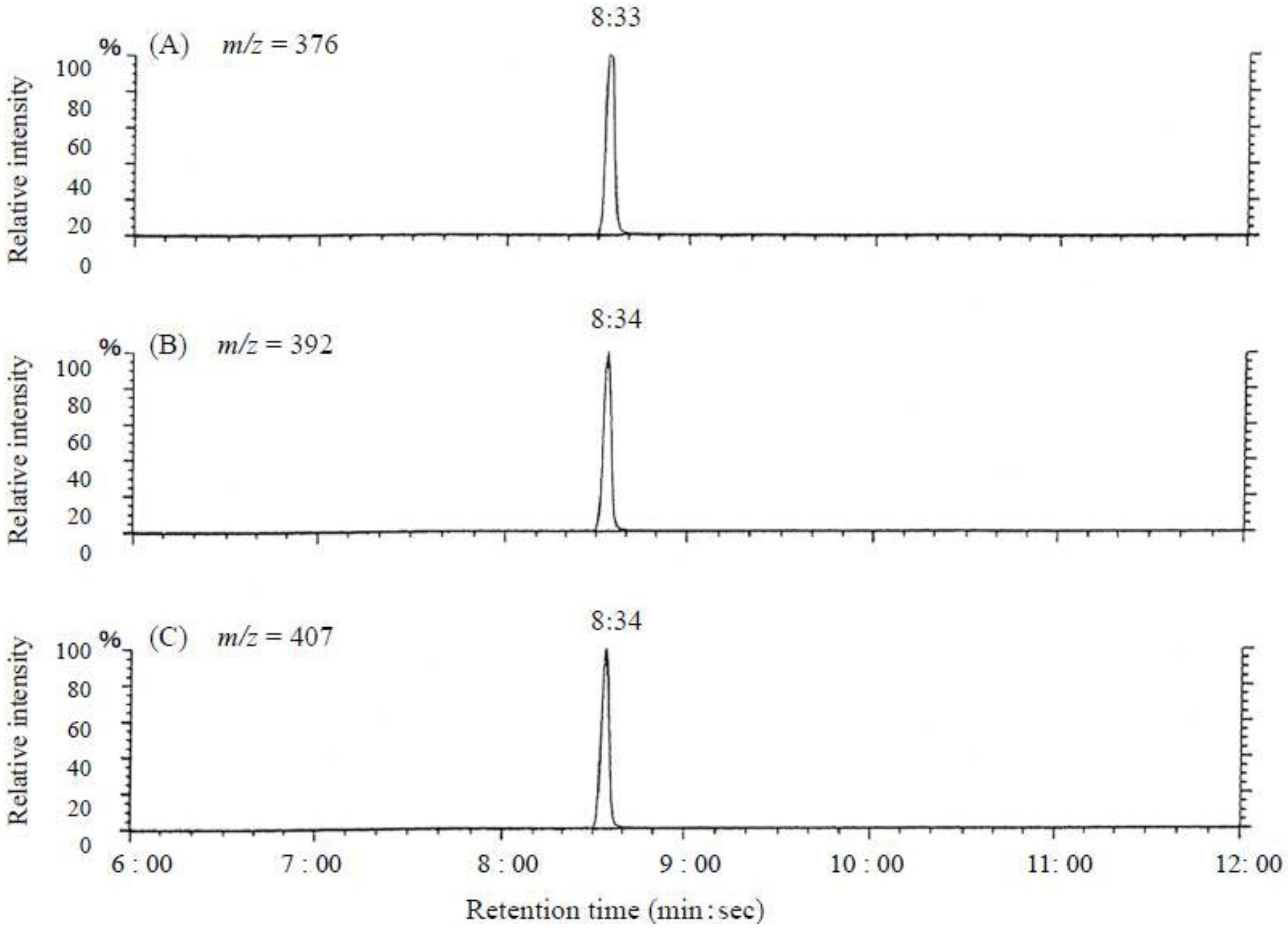
3.3.1. Mass Spectrometry
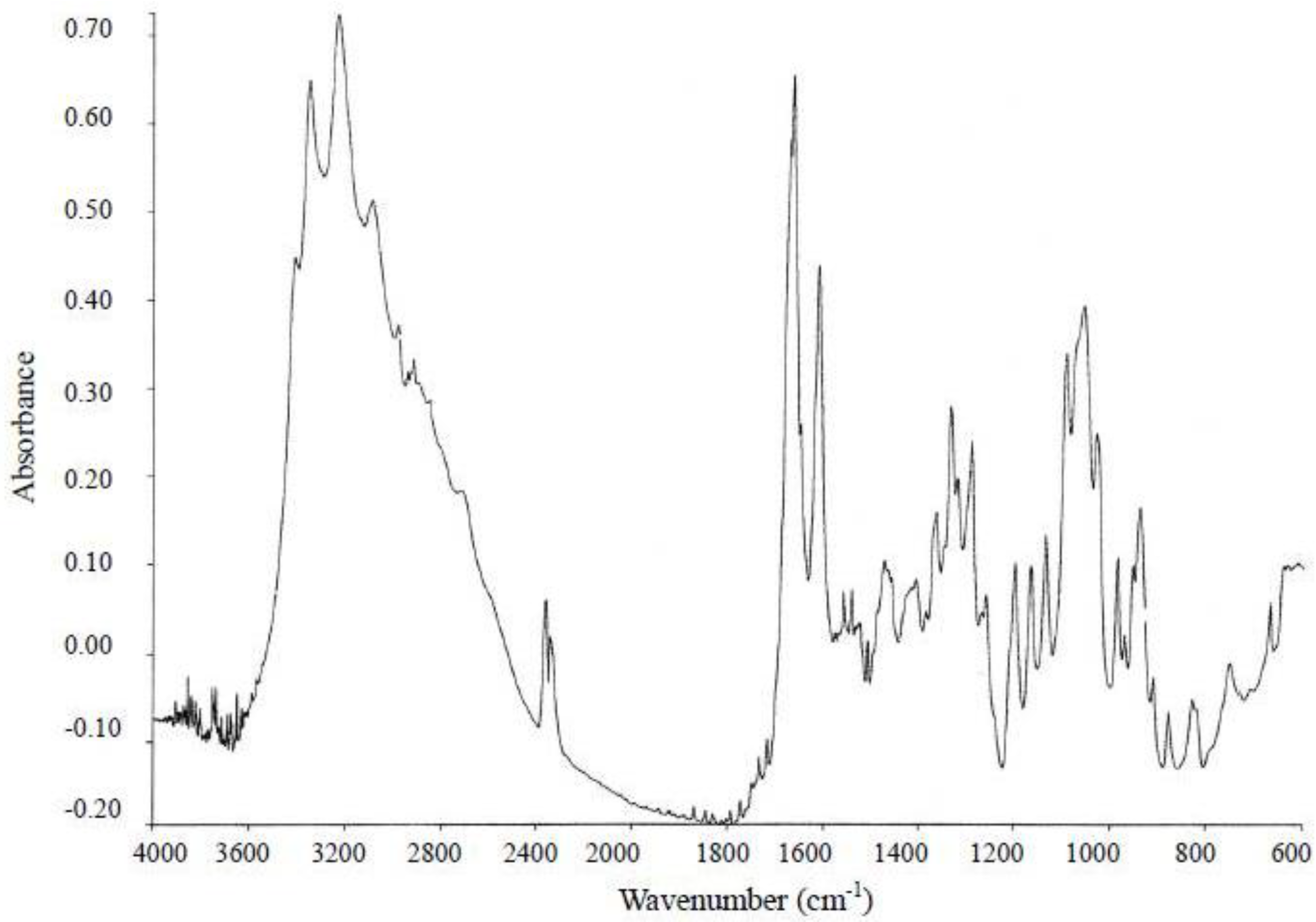
3.3.2. IR Spectrometry
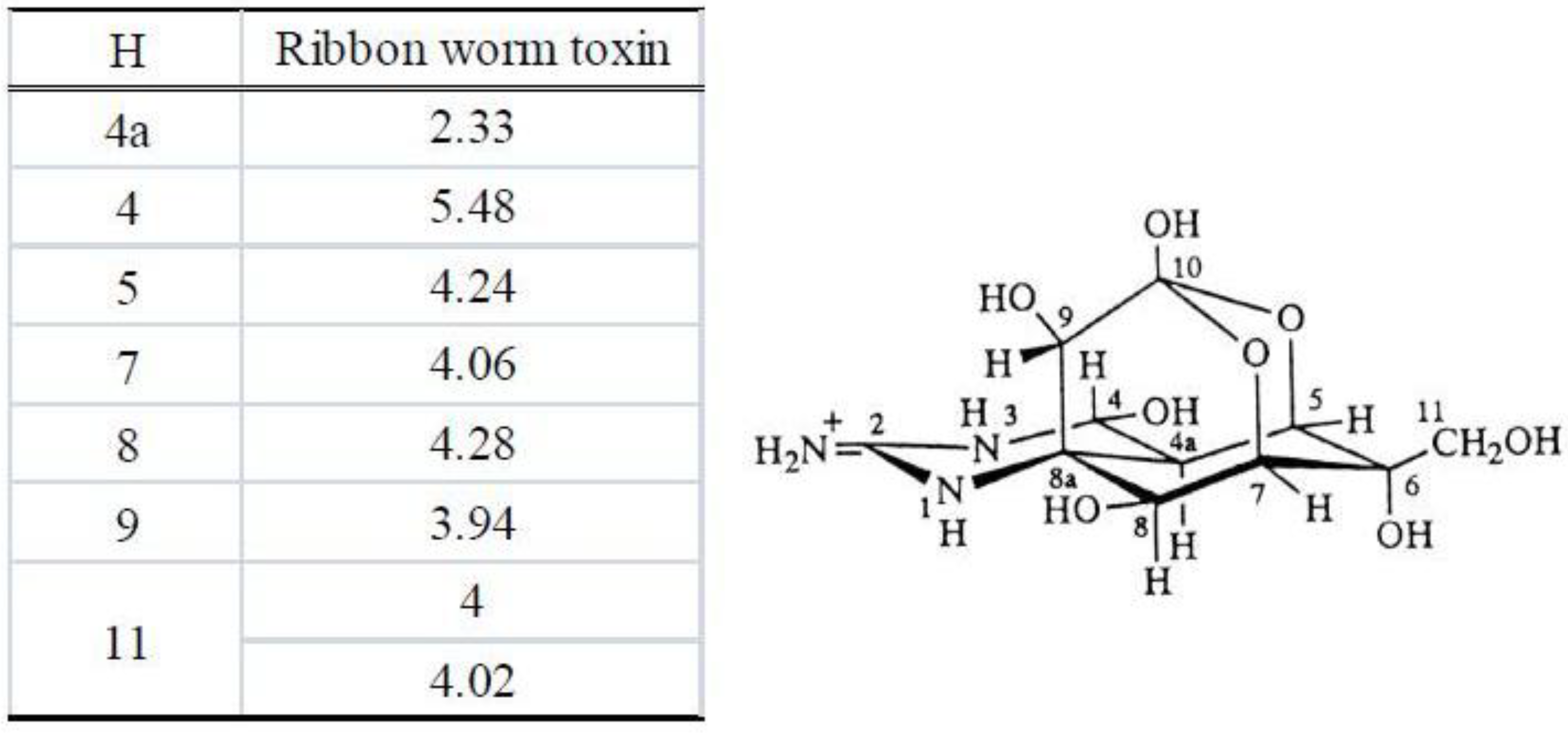
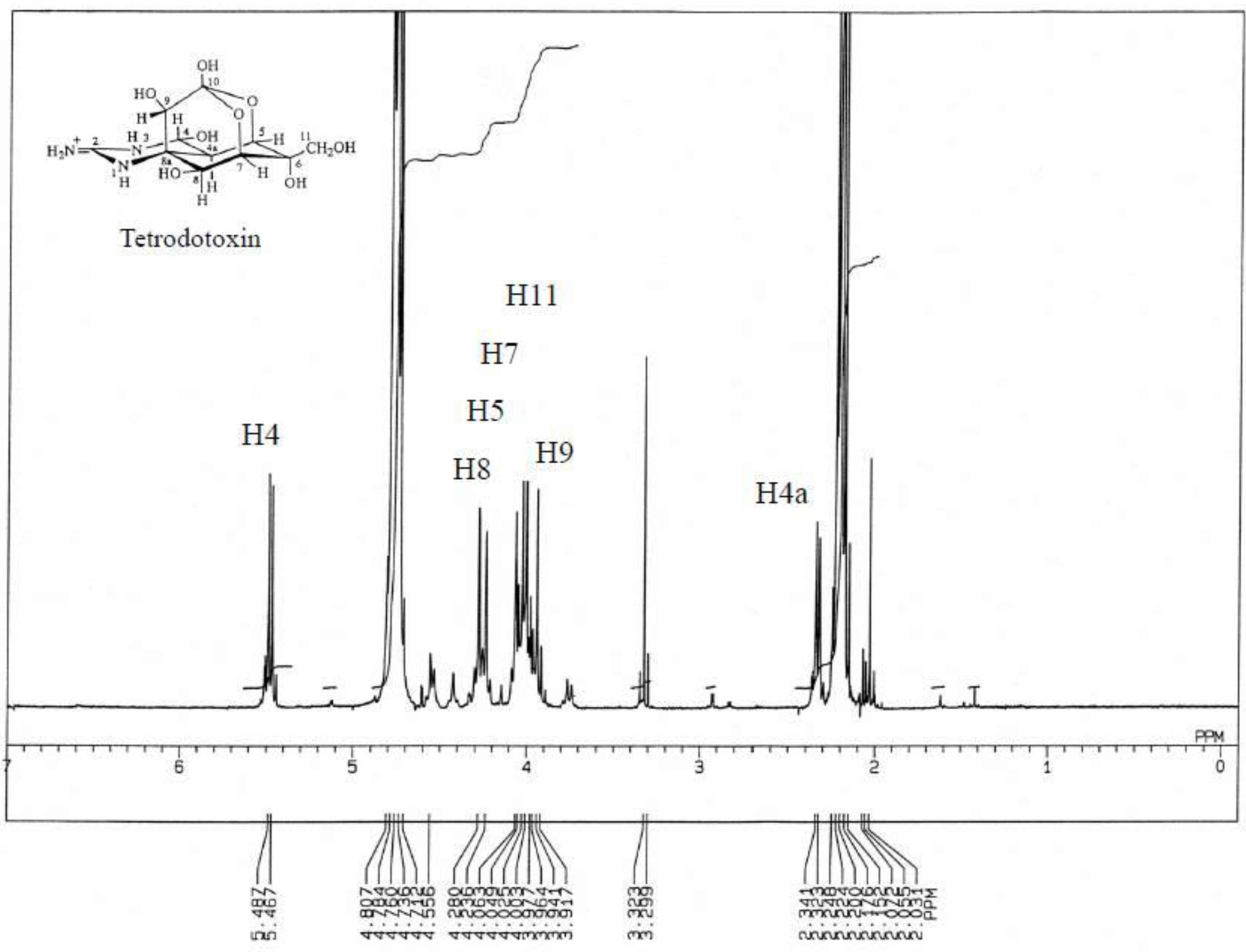
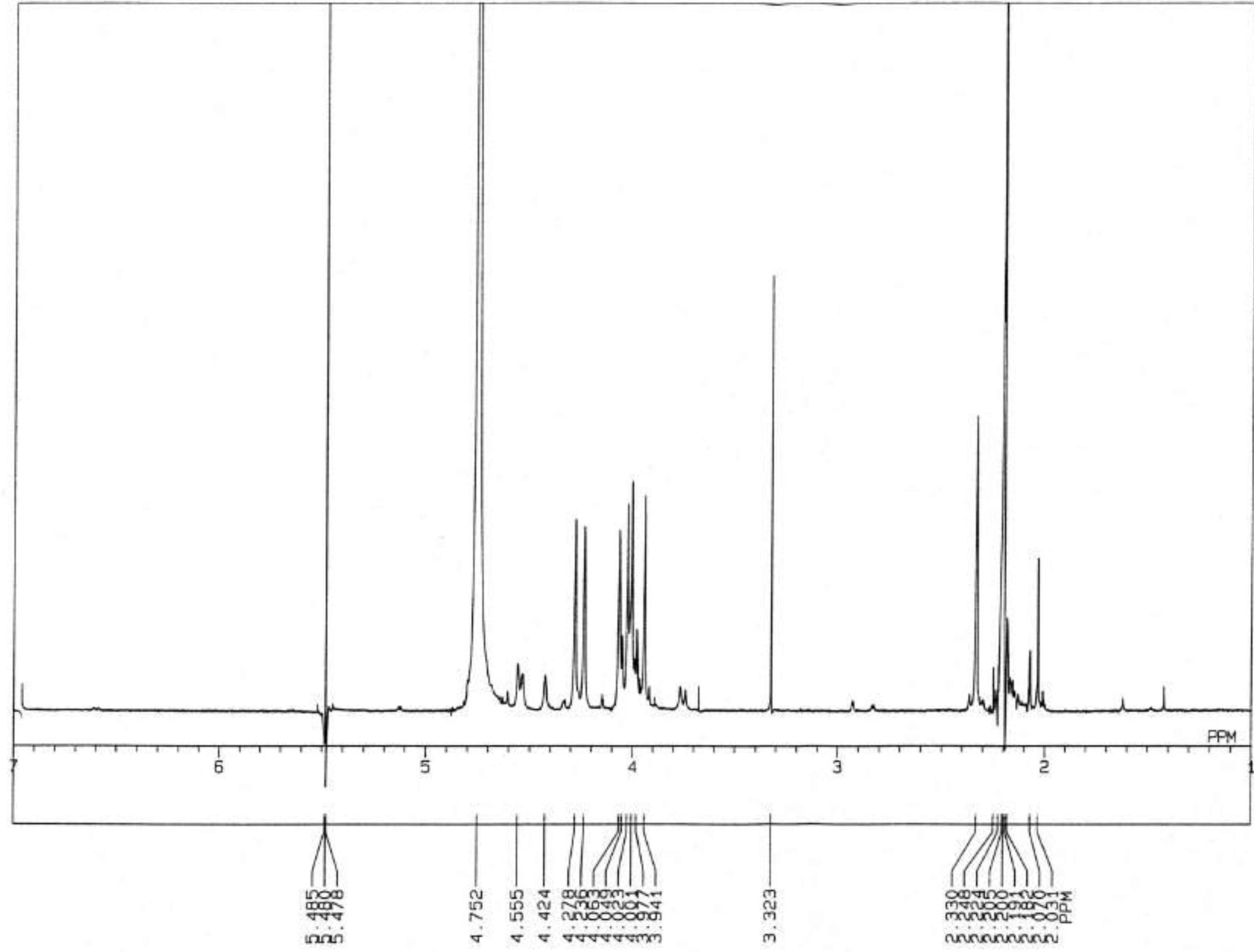
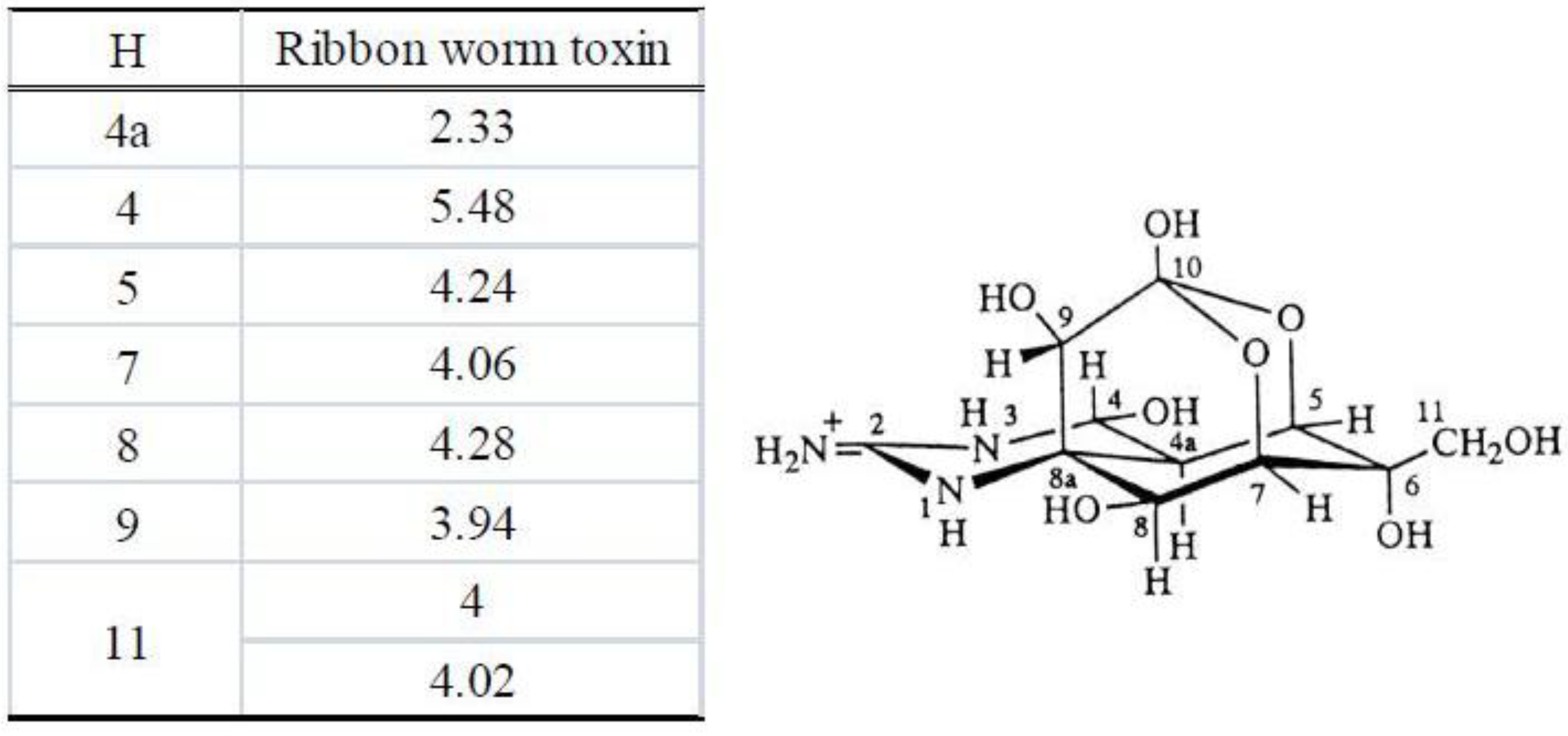
3.3.3. Nuclear Magnetic Resonance Spectrometry
3.3.4. Gas Chromatography-Mass Spectrometry
3.4. Results of the Instrumental Analysis





4. Distribution of TTX in C. simula from Hiroshima Bay
5. Toxicological Surveillance of the Nemertean Species from Other Locations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ribbon worms | Date of | No. of | Weight (g) | Toxicity (MU/g ) | HPLC-FLD | GC-MS | ||
|---|---|---|---|---|---|---|---|---|
| collection | specimens | (Mean ± S.D.) | (Mean ± S.D.) | TTX | 4epi-TTX | 4,9-anhyTTX | ||
| Nemertellina yamaokai | 30 July 2000 | 16 | 0.04 *1 | ND | − | − | − | − |
| Micrura akkeshiensis | 30 July 2000 | 3 | 0.02 *1 | ND | *2 | *2 | *2 | − |
| Malacobdella japonica | 30 July 2000 | 3 | 0.03 ± 0.01 | ND | − | − | ± | − |
| Tetrastemma nigrifrons | 30 July 2000 | 3 | 0.03 ± 0.02 | ND | − | − | − | − |
| Tetrastemma stimpsoni | 30 July 2000 | 5 | 0.02 *1 | ND | *2 | *2 | *2 | *2 |
| Amphiporus sp. | 30 July 2000 | 2 | 0.03 ± 0.01 | ND | + | − | − | − |
| Lineus bilineatus | 30 July 2000 | 12 | 0.03 *1 | ND | − | − | − | − |
| Cephalothrix simula | 30 July 2000 | 3 | 0.12 ± 0.06 | 1223 ± 97 | + | ± | + | + |
| Lineus torquatus | 30 July 2000 | 1 | 4.23 | ND | + | ± | + | + |
| Lineus alborostratus | 30 July 2000 | 1 | 1.81 | ND | + | + | + | + |
| Cephalothrix simula | 9 April 2008 | 6 | 0.17 ± 0.11 | 2958 ± 2483 | + | + | + | + |
| 1 July 2008 | 11 | 0.42 ± 0.22 | 1226 ± 614 | + | + | + | + | |
| 18 October 2008 | 8 | 0.29 ± 0.21 | 1683 ± 2220 | + | + | + | + | |
| 11 January 2009 | 11 | 0.21 ± 0.11 | 579 ± 343 | + | + | + | + | |
| Ribbon worms | Date of collection | No. of specimens | Weight (g) | Toxicity (MU/g ) | HPLC-FLD | GC-MS | ||
|---|---|---|---|---|---|---|---|---|
| (Mean ± S.D.) | (Mean ± S.D.) | TTX | 4epi-TTX | 4,9-anhyTTX | ||||
| Cephalothrix simula | 26 September 2000 | 2 | 0.25 * | 2021 ± 247 | + | ± | + | + |
| Nipponnemertes punctatula | 28 September 2000 | 2 | 0.03 ± 0.01 | ND | + | ± | ± | + |
| Cephalothrix simula | 28 September 2000 | 2 | 0.07 * | 1708 ± 279 | + | ± | ± | + |
| unknown specimen | 28 September 2000 | 12 | 0.22 * | ND | − | − | + | + |
6. Origin of TTX Compounds in the Ribbon Worm
7. General Discussion
Acknowledgements
Conflict of Interest
References
- Gibson, R. Nemerteans; Hutchinson: London, UK, 1972; p. 224. [Google Scholar]
- Kajihara, H.; Chernyshev, A.V.; Sun, S.-C.; Sundberg, P.; Crandall, F.B. Checklist of nemertean genera and species published between 1995 and 2007. Species Divers. 2008, 13, 245–274. [Google Scholar]
- Thollesson, M.; Norenburg, J.L. Ribbon worm relationships: A phylogeny of the phylum Nemertea. Proc. Biol. Sci. 2003, 270, 407–414. [Google Scholar] [CrossRef]
- Andrade, S.C.S.; Strand, M.; Schwartz, M.; Chen, H.-X.; Kajihara, H.; von Döhren, J.; Sun, S.-C.; Junoy, J.; Thiel, M.; Norenburg, J.L.; et al. Disentangling ribbon worm relationships: Multi-locus analysis supports traditional classification of the phylum Nemertea. Cladistics 2012, 28, 141–159. [Google Scholar] [CrossRef]
- Kajihara, H. A taxonomic catalogue of Japanese nemerteans (phylum Nemertea). Zoolog. Sci. 2007, 24, 287–326. [Google Scholar] [CrossRef]
- McDermott, J.J.; Roe, P. Food, feeding behavior and feeding ecology of nemerteans. Amer. Zool. 1985, 25, 113–125. [Google Scholar]
- Christy, J.H.; Goshima, S.; Backwell, P.R.Y.; Kreuter, T.J. Nemertean predation on the tropical fiddler crab Uca musica. Hydrobiologia 1998, 365, 233–239. [Google Scholar]
- Thiel, M.; Kruse, I. Status of the Nemertea as predators in marine ecosystems. Hydrobiologia 2001, 456, 21–32. [Google Scholar] [CrossRef]
- Thiel, M.; Ullrich, N.; Vásquez, N. Predation rates of nemertean predators: The case of a rocky shore hoplonemertean feeding on amphipods. Hydrobiologia 2001, 456, 45–57. [Google Scholar] [CrossRef]
- Bourque, D.; Miron, G.; Landry, T. Predation on soft-shell clams (Mya arenaria) by the nemertean Cerebratulus lacteus in Atlantic Canada: Implications for control measures. Hydrobiologia 2001, 456, 33–44. [Google Scholar] [CrossRef]
- Bourque, G.; Miron, G.; Landry, T. Predator-prey relationship between the nemertean Cerebratulus lacteus and the soft-shell clam, Mya arenaria: Surface-exploration activity and qualitative observations on feeding behaviour. Can. J. Zool. 2002, 80, 1204–1211. [Google Scholar]
- Heine, J.N.; McClintock, J.B.; Slattery, M.; Weston, J. Energetic composition, biomass, and chemical defense in the common antarctic nemertean Parborlasia corrugatus McIntos. J. Exp. Mar. Biol. Ecol. 1991, 153, 15–25. [Google Scholar] [CrossRef]
- Bacq, Z.M. Les poisons des Nemertiens. Bull. Cl. Sci. Acad. R. Belg. 1936, 22, 1072–1079. [Google Scholar]
- Bacq, Z.M. L“amphiporine’ et la ‘némertine’, poisons des vers némertiens. Arch. Int. Physiol. 1937, 44, 190–204. [Google Scholar] [CrossRef]
- Kem, W.R. A study of occurrence of anabaseine in Paranemertes and other nemertines. Toxicon 1971, 9, 23–32. [Google Scholar] [CrossRef]
- Kem, W.R. Biochemistry of nemertines toxins. In Marine Pharmacognosy. Action of Marine Biotoxins at the Cellular Level; Martin, D.F., Padilla, G.M., Eds.; Academic Press: New York, NY, USA, 1973; pp. 37–84. [Google Scholar]
- Kem, W.R. Structure and action of nemertine toxins. Amer. Zool. 1985, 25, 99–111. [Google Scholar]
- Kem, W.R.; Scott, K.N.; Duncan, J.H. Hoplonemertine worms—A new source of pyridine neurotoxins. Experientia 1976, 32, 684–686. [Google Scholar] [CrossRef]
- Kem, W.R. Pyridine alkaloid distribution in the hoplonemertines. Hydrobiologia 1988, 156, 145–151. [Google Scholar] [CrossRef]
- Turbeville, J.M.; Ruppert, E.E. Comparative ultrastructure and the evolution of nemertines. Amer. Zool. 1985, 25, 53–71. [Google Scholar]
- Turbeville, J.M. An ultrastructural analysis of coelomogenesis in the hoplonemertine Prosorhochmus americanus and the polychaete Magelona sp. J. Morphol. 1986, 187, 51–60. [Google Scholar] [CrossRef]
- Gontcharoff, M. Note sur l’alimentation de quelques némertes. Ann. Sci. Nat. 1948, 10, 75–78. [Google Scholar]
- Jennings, J.B.; Gibson, R. Observations on the nutrition of seven species of rhynchocoelan worms. Biol. Bull. 1969, 136, 405–433. [Google Scholar] [CrossRef]
- McDermott, J.J. The feeding biology of Nipponnemerte pulcher (Johnston) (Hoplonemertea), with some ecological implications. Ophelia 1984, 23, 1–21. [Google Scholar] [CrossRef]
- Wang, H.-Y.; Sun, S.-C.; Li, Q.-L. Laboratory observations on the feeding behavior and feeding rate of the nemertean Procephalothrix simulus. Biol. Bull. 2008, 214, 166–175. [Google Scholar] [CrossRef]
- Asakawa, M.; Toyoshima, T.; Shida, Y.; Noguchi, T.; Miyazawa, K. Paralytic toxins in a ribbon worm Cephalothrix species (Nemertean) adherent to cultured oysters in Hiroshima Bay, Hiroshima Prefecture, Japan. Toxicon 2000, 38, 763–773. [Google Scholar] [CrossRef]
- Asakawa, M.; Toyoshima, T.; Ito, K.; Bessho, K.; Yamaguchi, C.; Tsunetsugu, S.; Shida, Y.; Kajihara, H.; Mawatari, S.F.; Noguchi, T.; Miyazawa, K. Paralytic toxicity in the ribbon worm Cephalothrix species (Nemertea) in Hiroshima Bay, Hiroshima Prefecture, Japan and the isolation of tetrodotoxin as a main component of its toxins. Toxicon 2003, 41, 747–733. [Google Scholar] [CrossRef]
- Kawabata, T. Assay Method for Tetrodotoxin. In Food Hygiene Examination Manual; Kawabata, T., Ed.; Japanese Food Hygiene Association: Tokyo, Japan, 1978; Volume 2, pp. 232–239. [Google Scholar]
- Miyzawa, K.; Higashiyama, M.; Ito, K.; Noguchi, T.; Arakawa, O.; Shida, Y.; Hashimoto, K. Tetrodotoxin in two species of ribbon worm (Nemertini), Lineus fuscoviridis and Tubulanus punctatus. Toxicon 1988, 26, 897–864. [Google Scholar]
- Ali, A.E.; Arakawa, O.; Noguchi, T.; Miyazawa, K.; Shida, Y.; Hashimoto, K. Tetrodotoxin and related substances in a ribbon worm Cephalothrix linearis (Nemertean). Toxincon 1990, 28, 1083–1093. [Google Scholar]
- Asakawa, M.; Gomez-Delan, G.; Tsuruda, S.; Shimomura, M.; Shida, Y.; Taniyama, T.; Barte-Quilantang, M.; Shindo, J. Toxicity assessment of the xanthid crab Demania cultripes from Cebu Island, Philippines. J. Toxicol. 2010. [Google Scholar] [CrossRef]
- Noguchi, T.; Mahmud, Y. Current methodologies for detection of tetrodotoxin. Toxin Rev. 2001, 20, 35–50. [Google Scholar] [CrossRef]
- Goto, T.; Kishi, Y.; Takahashi, S.; Hirata, Y. Tetrodotoxin. Tetrahedron 1965, 21, 2059–2088. [Google Scholar] [CrossRef]
- Narita, H.; Noguchi, T.; Maruyama, J.; Ueda, Y.; Hashimoto, K.; Watanabe, Y.; Hida, K. Occurrence of tetrodo toxin in a trumpet shell, ‘boshubora’ Charonia sauliae. Bull. Japan. Soc. Sci. Fish. 1981, 47, 935–941. [Google Scholar] [CrossRef]
- Maruyama, J.; Noguchi, T.; Matsunaga, S.; Hashimoto, K. Fast atom bombardment- and secondary ion-mass spectrometry of paralytic shellfish poisons and tetrodotoxin. Agric. Biol. Chem. 1984, 48, 2783–2788. [Google Scholar] [CrossRef]
- Nagashima, Y.; Nishio, S.; Noguchi, T.; Arakawa, O.; Kanoh, S.; Hashimoto, K. Detection of tetrodotoxin by thin-layer chromatography/fast atom bombardment mass spectrometry. Anal. Biochem. 1988, 175, 258–262. [Google Scholar] [CrossRef]
- Tsuda, K.; Ikuma, S.; Kawamura, M.; Tachikawa, R.; Sakai, K.; Tamura, C.; Amakasu, O. Tetrodotoxin. VII. On the structures of tetrodotoxin and its derivatives. Chem. Pharm. Bull. 1964, 12, 1357–1374. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Brown, G.B.; Mosher, H.S.; Furman, F.A. Tetrodotoxin: Occurrence in atelopid frogs of Costa Rica. Science 1975, 189, 151–152. [Google Scholar]
- Sheumack, D.D.; Howden, M.E.; Spence, H.I.; Quinn, R.J. Maculotoxin: A neurotoxin from the venom glands of the octopus Hapalochlaena maculosa identified as tetrodotoxin. Science 1978, 199, 188–189. [Google Scholar]
- Hashimoto, Y. Nemertine Toxins. In Marine Toxins and Other Bioactive Metabolites; Hashimoto, Y., Ed.; Japan Scientific Societies Press: Tokyo, Japan, 1979; pp. 298–302. [Google Scholar]
- Kawatsu, K.; Hamano, Y.; Yoda, T.; Terano, Y.; Shibata, T. Rapid and high sensitive enzyme immunoassay for quantitative determination of tetrodotoxin. Jpn. J. Med. Sci. Biol. 1997, 50, 133–150. [Google Scholar]
- Tanu, M.B.; Mahmud, Y.; Arakawa, O.; Takatani, T.; Kajihara, H.; Kawatsu, K.; Hamano, Y.; Asakawa, M.; Miyazawa, K.; Noguchi, Y. Immunoenzymatic visualization of tetrodotoxin (TTX) in Cephalothrix species (Nemertea: Anopla: Palaeonemertea: Cephalotrichidae) and Planocera reticulata (Platyhelminthes: Turbellaria: Polycladida: Planoceridae). Toxicon 2004, 44, 515–520. [Google Scholar] [CrossRef]
- Asakawa, M.; Kajihara, H. Toxicity of Several Species of Ribbon Worms in the Coastal Area of Japan. In Proceedings of Program and Abstracts in the Japanese Society of Fisheries Science, Autumn Meeting, Kyoto, Japan, 22–25 September 2010; p. 91.
- Noguchi, T.; Jeon, J.-K.; Arakawa, O.; Sugita, H.; Deguchi, Y.; Hashimoto, K. Occurrence of tetrodootxin and anhydrotetrodotoxin in Vibrio sp. isolated from the intestines of a xanthid crab, Atergatis floridus. J. Biochem. 1986, 99, 311–314. [Google Scholar]
- Noguchi, T.; Hwang, D.F.; Arakawa, O.; Sugita, H.; Deguchi, Y.; Hashimoto, K. Vibrio alginolyticus, a tetrodotoxin-producing bacterium, in the intestines of the puffer Fugu vermicularis vermicularis. Mar. Biol. 1987, 94, 625–630. [Google Scholar] [CrossRef]
- Asakawa, M.; Tsunetsugu, S.; Ito, K.; Shida, Y.; Miyazawa, K. Tetrodotoxin-Producing Ability of Bacteria Isolated from the Ribbon Worm Cephalothrix sp. (Nemertea) in Hiroshima Bay, Hiroshima Prefecture, Japan. In Proceedings of 14th World Congress on AnimalPlant & Microbial Toxins, Aderade, Australia, 2003; p. 110.
- McEvoy, E.G.; Rogers, A.; Gibson, R. Preliminary investigation of Vibrio alginolyticus-like bacteria associated with marine nemerteans. Hydrobiologia 1998, 365, 287–290. [Google Scholar] [CrossRef]
- Carrol, S.; McEvoy, E.G.; Gibson, R. The production of tetrodotoxin-like substances by nemertean worms in conjunction with bacteris. J. Exp. Mar. Biol. Ecol. 2003, 288, 51–63. [Google Scholar] [CrossRef]
- Mosher, H.S.; Fuhrman, F.A.; Buchwald, H.D.; Fisher, H.G. Tarichatoxin-tetrodotoxin: A potent neurotoxin. Science 1964, 144, 1100. [Google Scholar]
- Noguchi, T.; Hashimoto, Y. Isolation of tetrodotoxin from a goby Gobius criniger. Toxicon 1973, 11, 305–307. [Google Scholar] [CrossRef]
- Pires, O.R., Jr.; Sebben, A.; Schwartz, E.F.; Morales, C.; Bloch, C., Jr.; Schwartz, C.A. Further report of the occurrence of tetrodotoxin and new analogues in the anuran family Brachycephalida. Toxicon 2005, 45, 73–79. [Google Scholar] [CrossRef]
- Williams, B.L.; Caldwell, R.L. Intra-organismal distribution of tetrodotoxin in two species of blue-ringed octopuses (Hapalochlaena fasciata and H. lunulata). Toxicon 2009, 54, 345–353. [Google Scholar] [CrossRef]
- Noguchi, T.; Maruyama, J.; Ueda, Y.; Hashimoto, K.; Harada, T. Occurrence of tetrodotoxin in the Japanese ivory shell Babylonia japonica. Bull. Jpn. Soc. Sci. Fish. 1981, 47, 909–913. [Google Scholar] [CrossRef]
- Noguchi, T.; Maruyama, J.; Narita, H.; Hashimoto, K. Occurrence of tetrodotoxin in the gastropod mollusk Tutufa lissostoma (frog shell). Toxicon 1984, 22, 219–226. [Google Scholar] [CrossRef]
- McNabb, P.; Selwood, A.I.; Munday, R.; Wood, S.A.; Taylor, D.I.; Mackenzie, L.A.; Ginkel, R.V.; Rhodes, L.L.; Cornelisen, C.; Heasman, K.; et al. Detection of tetrodotoxin from the grey side-gilled sea slug—Pleurobranchaea maculate, and associated dog neurotoxicosis on beaches adjacent to the Hauraki Gulf, Auckland, New Zealan. Toxicon 2010, 56, 466–473. [Google Scholar] [CrossRef]
- Noguchi, T.; Narita, H.; Maruyama, J.; Hashimoto, K. Tetrodotoxin in the starfish Astropecten polyacanthus, in association with toxification of a trumpet shell, “boshubora” Charonia saulia. Nippon Suisann Gakkaishi 1982, 48, 1173–1177. [Google Scholar] [CrossRef]
- Noguchi, T.; Uzu, A.; Koyama, K.; Hashimoto, K. Occurrence of tetrodotoxin as the major toxin in a xanthid crab Atergatis floridus. Nippon Suisann Gakkaishi 1983, 49, 1887–1892. [Google Scholar] [CrossRef]
- Yasumura, D.; Oshima, Y.; Yasumoto, T.; Alcala, A.C. Tetrodotoxin and paralytic shellfish toxins in Philippine crabs. Agric. Biol. Chem. 1986, 50, 593–598. [Google Scholar]
- Jeon, J.K.; Miyazawa, K.; Noguchi, T.; Narita, H.; Ito, K.; Hashimoto, K. Occurrence of paralytic toxicity in marine flatworm. Nippon Suisann Gakkaishi 1986, 52, 1065–1069. [Google Scholar] [CrossRef]
- Miyazawa, K.; Jeon, J.K.; Maruyama, J.; Noguchi, T.; Ito, K.; Hashimoto, K. Occurrence of tetrodotoxin in the flatworm Planocera multitentaculata. Toxicon 1986, 24, 645–650. [Google Scholar] [CrossRef]
- Miyazawa, K.; Jeon, J.K.; Maruyama, J.; Noguchi, T.; Ito, K.; Hashimoto, K. Distribution of tetrodotoxin in the tissues of the flatworm Planocera multitentaculata (Platyhelminthes). Toxicon 1987, 25, 975–980. [Google Scholar] [CrossRef]
- Kungswan, A.; Nagashima, Y.; Noguchi, T.; Shida, Y.; Suvapeepan, S.; Suwansakornkul, P.; Hashimoto, K. Tetrodotoxin in the horseshoe crab Carcinoscorpius rotundicauda inhabiting Thailand. Nippon Suisann Gakkaishi 1987, 53, 261–266. [Google Scholar] [CrossRef]
- Thuesen, E.V.; Kogure, K.; Hashimoto, K.; Nemoto, T. Poisonous arrowworms: A tetrodotoxin venom in the marine phylum Chaetognatha. J. Exp. Mar. Biol. Ecol. 1988, 116, 249–256. [Google Scholar] [CrossRef]
- Kems, W.R. Structure and membrane actions of a marine worm protein cytolysin, Cerebratulus toxin A-III. Toxicology 1994, 87, 189–203. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Asakawa, M.; Ito, K.; Kajihara, H. Highly Toxic Ribbon Worm Cephalothrix simula Containing Tetrodotoxin in Hiroshima Bay, Hiroshima Prefecture, Japan. Toxins 2013, 5, 376-395. https://doi.org/10.3390/toxins5020376
Asakawa M, Ito K, Kajihara H. Highly Toxic Ribbon Worm Cephalothrix simula Containing Tetrodotoxin in Hiroshima Bay, Hiroshima Prefecture, Japan. Toxins. 2013; 5(2):376-395. https://doi.org/10.3390/toxins5020376
Chicago/Turabian StyleAsakawa, Manabu, Katsutoshi Ito, and Hiroshi Kajihara. 2013. "Highly Toxic Ribbon Worm Cephalothrix simula Containing Tetrodotoxin in Hiroshima Bay, Hiroshima Prefecture, Japan" Toxins 5, no. 2: 376-395. https://doi.org/10.3390/toxins5020376