Isolation and Molecular Characterization of Two Lectins from Dwarf Elder (Sambucus ebulus L.) Blossoms Related to the Sam n1 Allergen

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
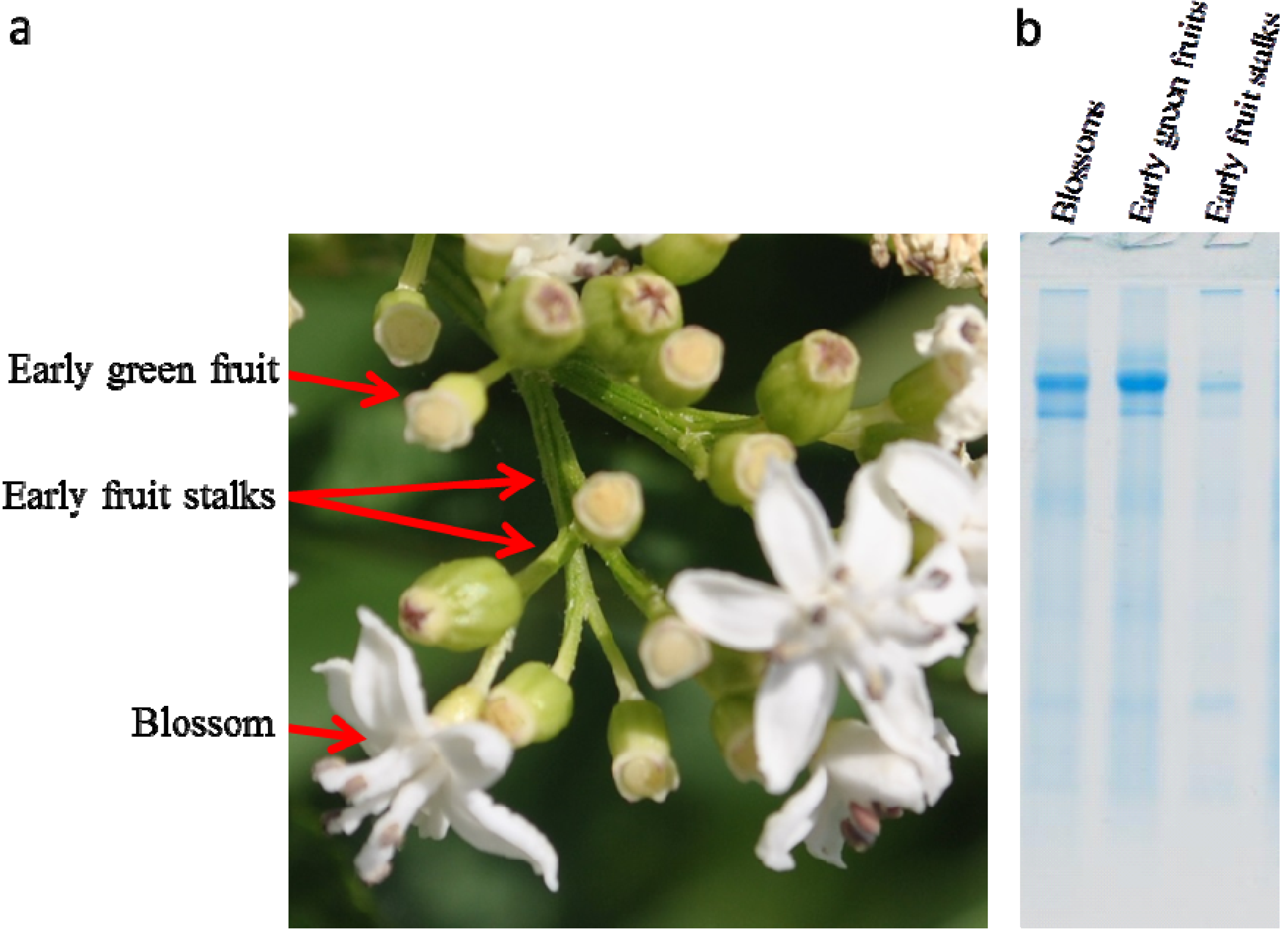
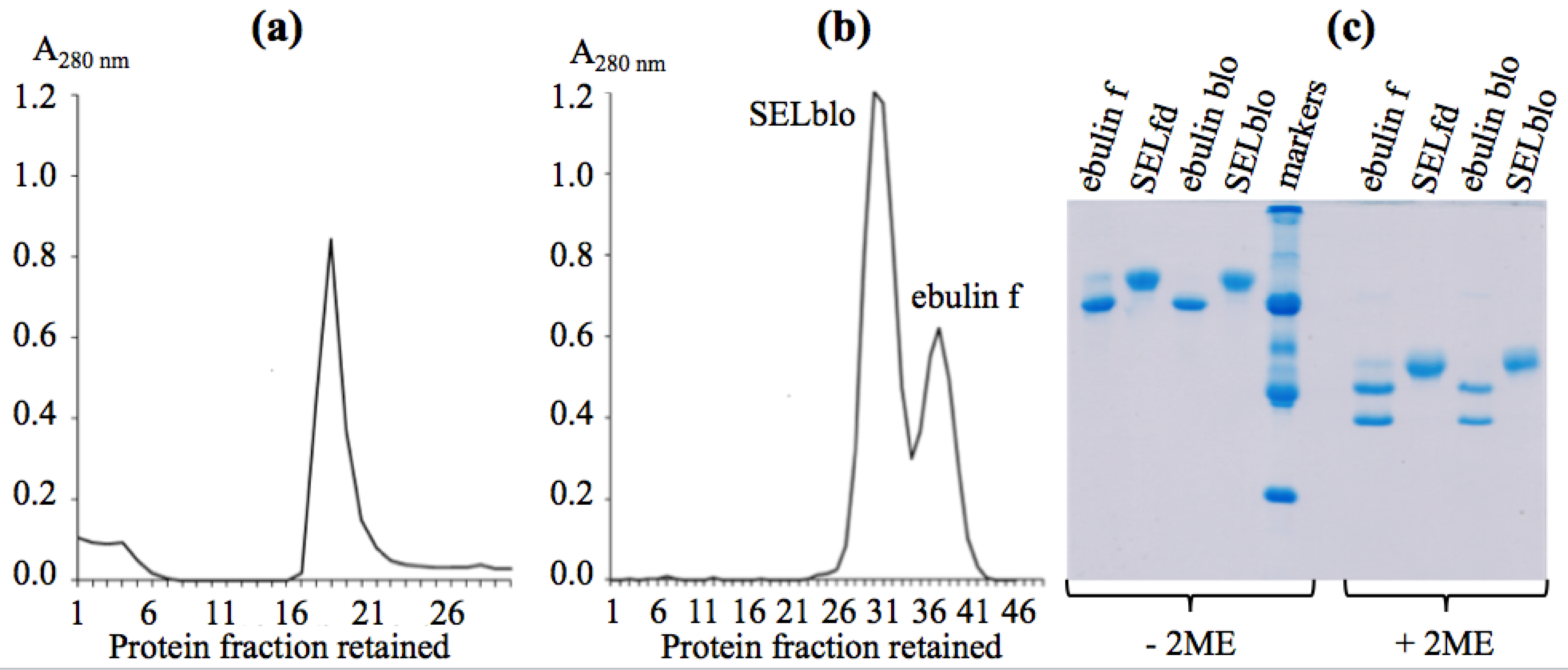
2.1. Isolation of Dwarf Elder Blossom d-Galactose-Specific Lectins
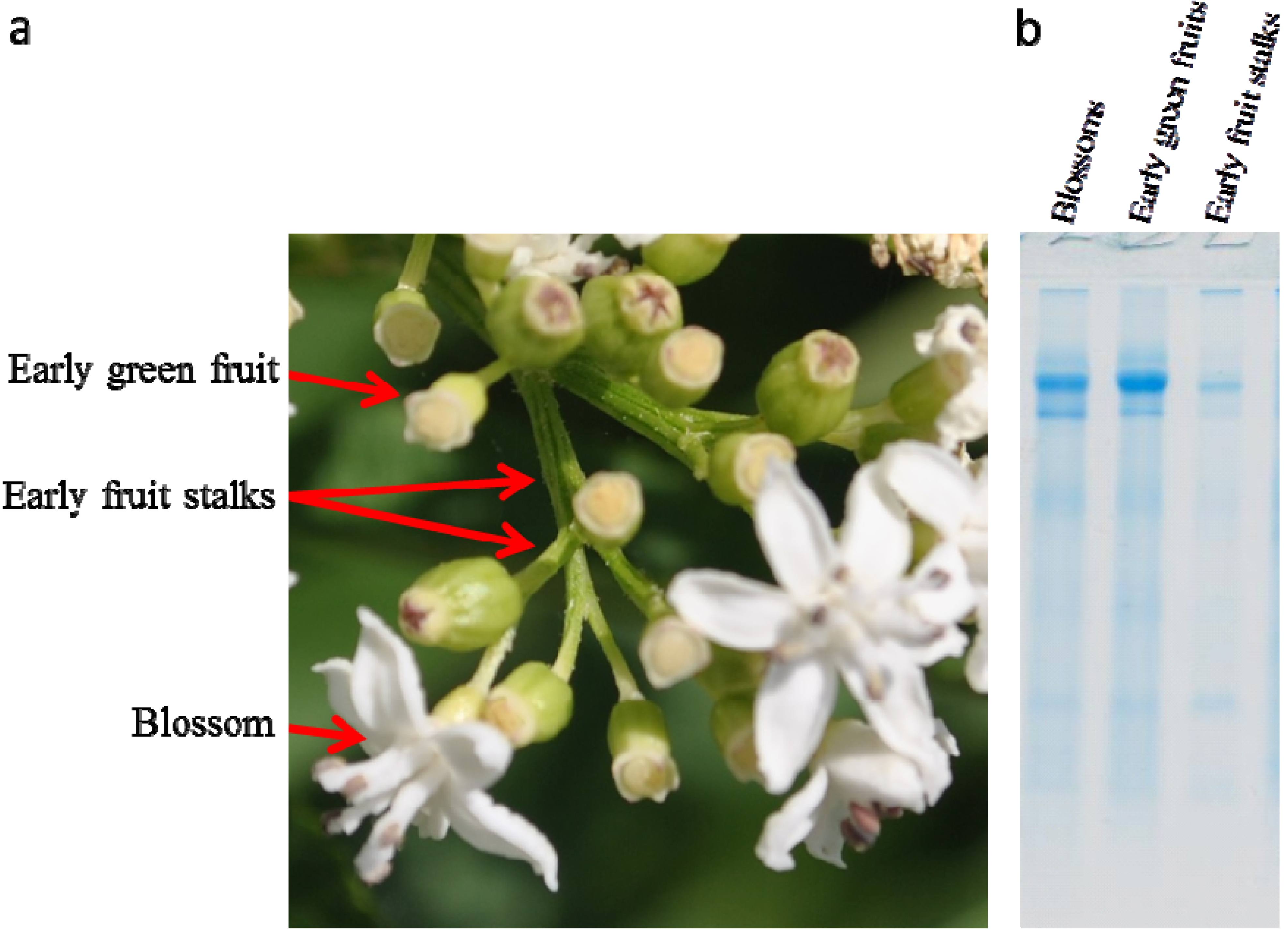
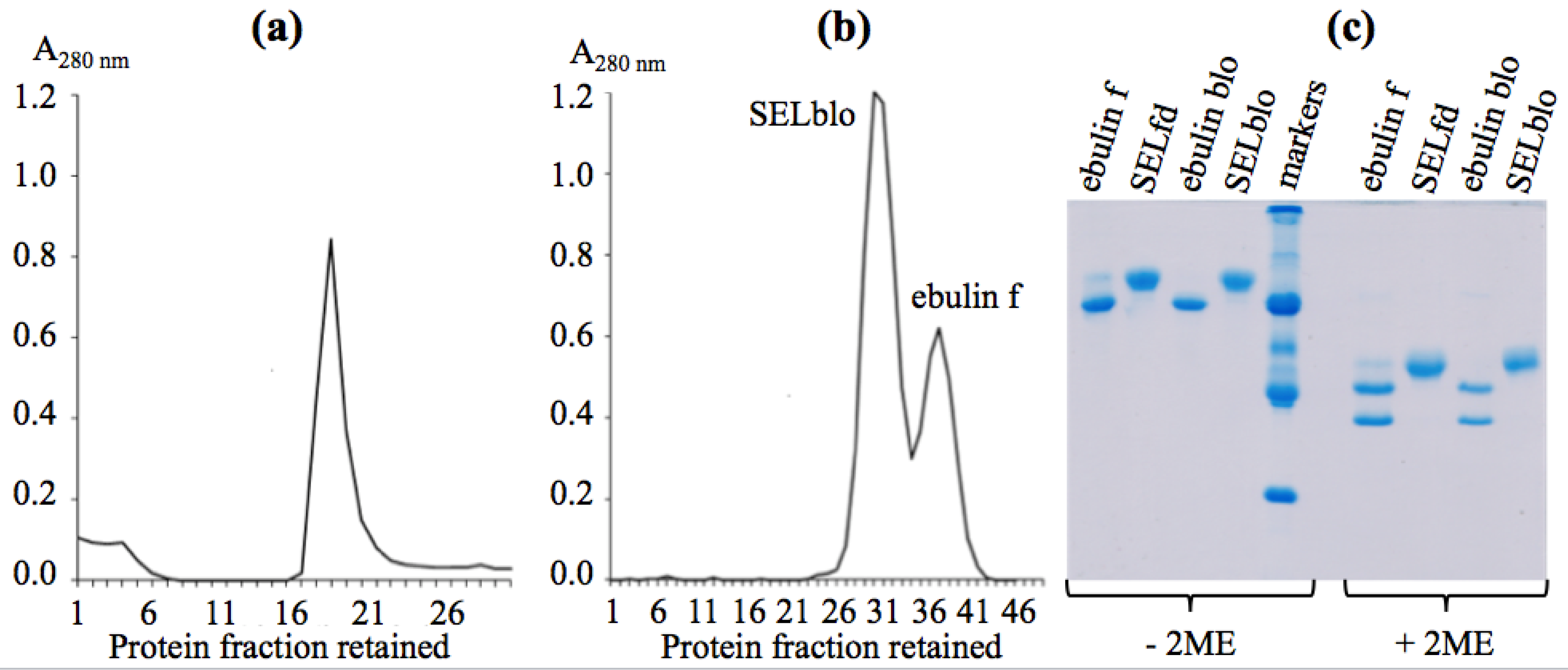
2.2. Molecular Characterization by SDS-PAGE
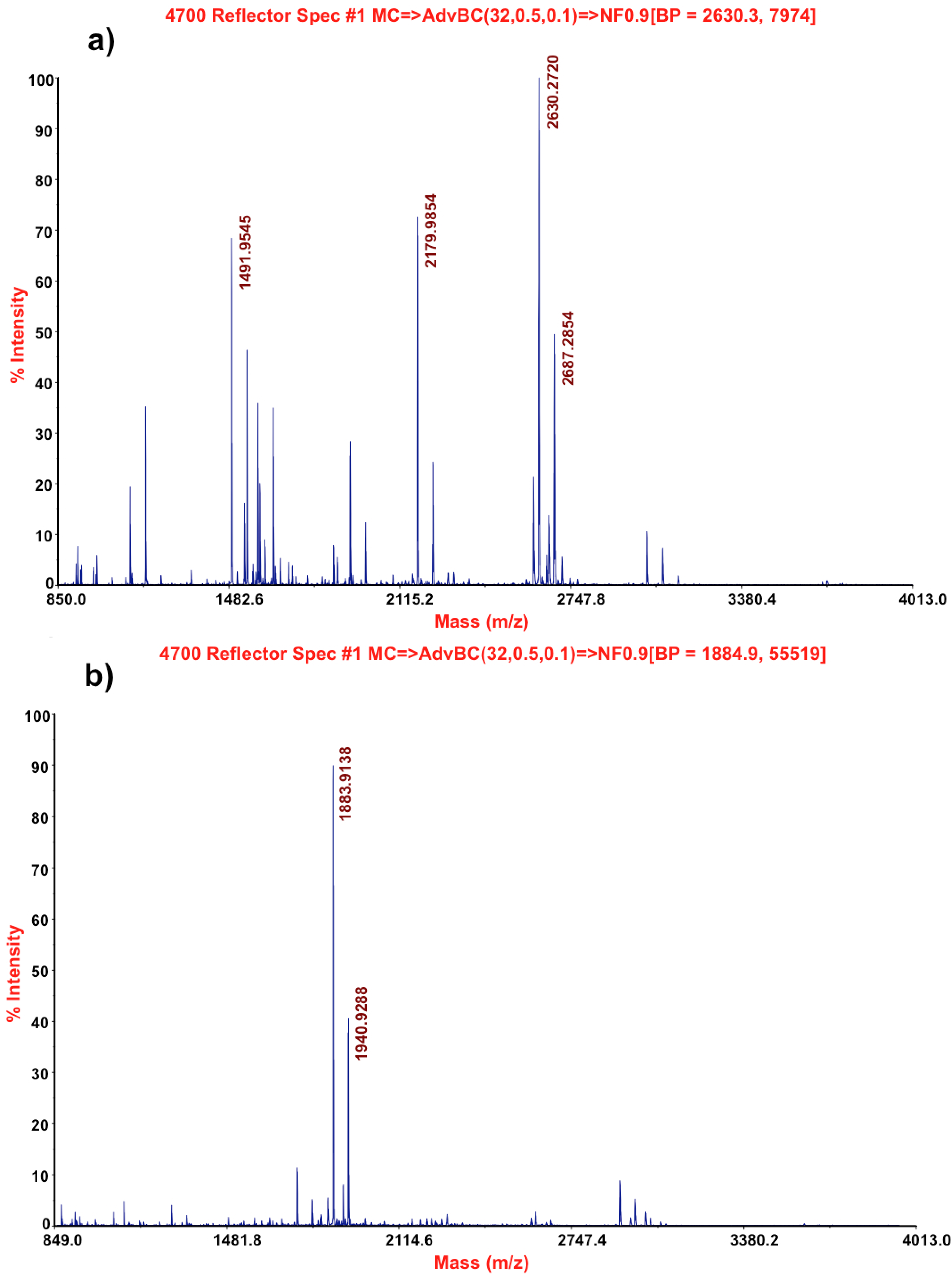
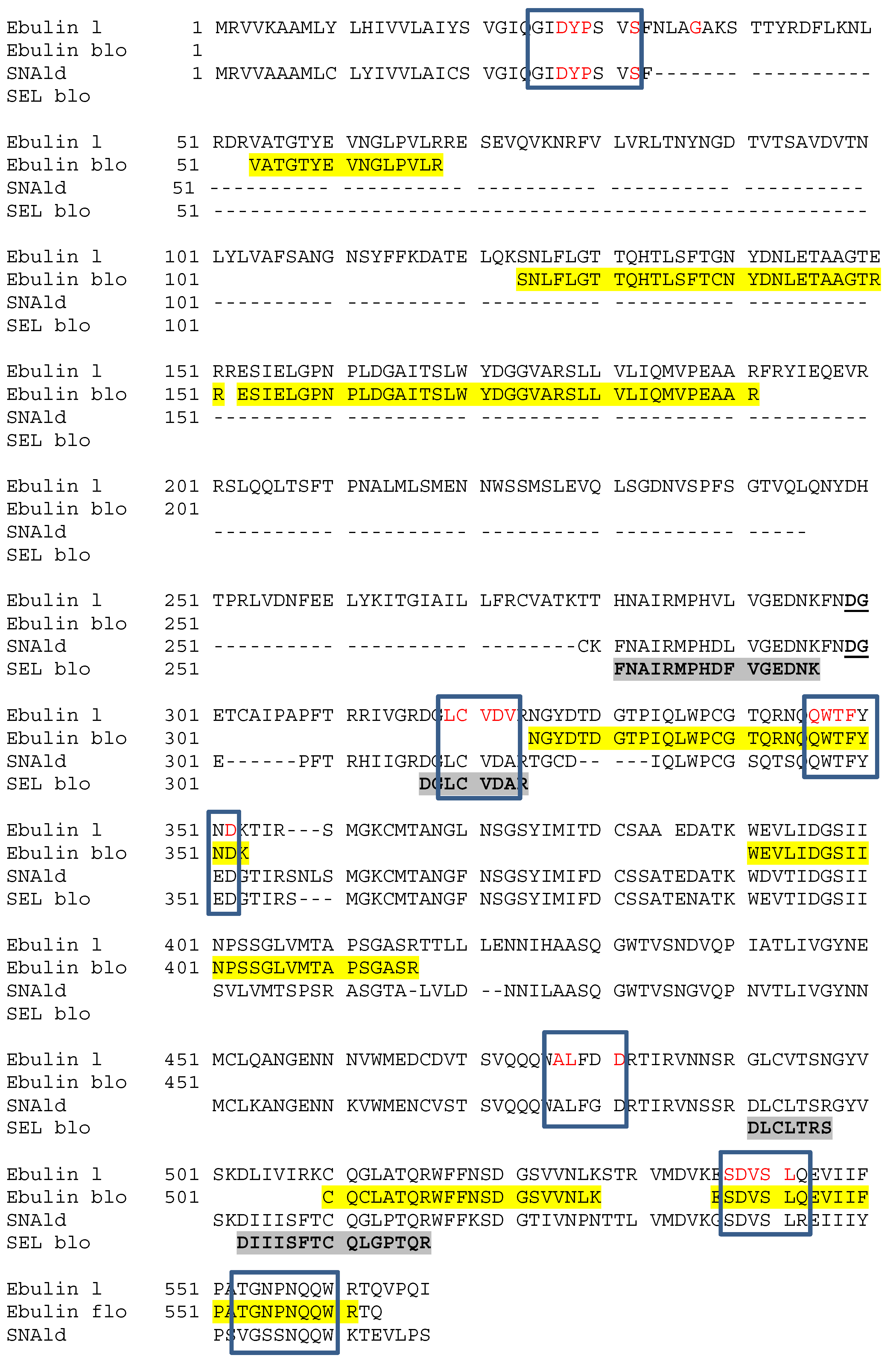
2.3. Molecular Characterization by Mass Spectrometry


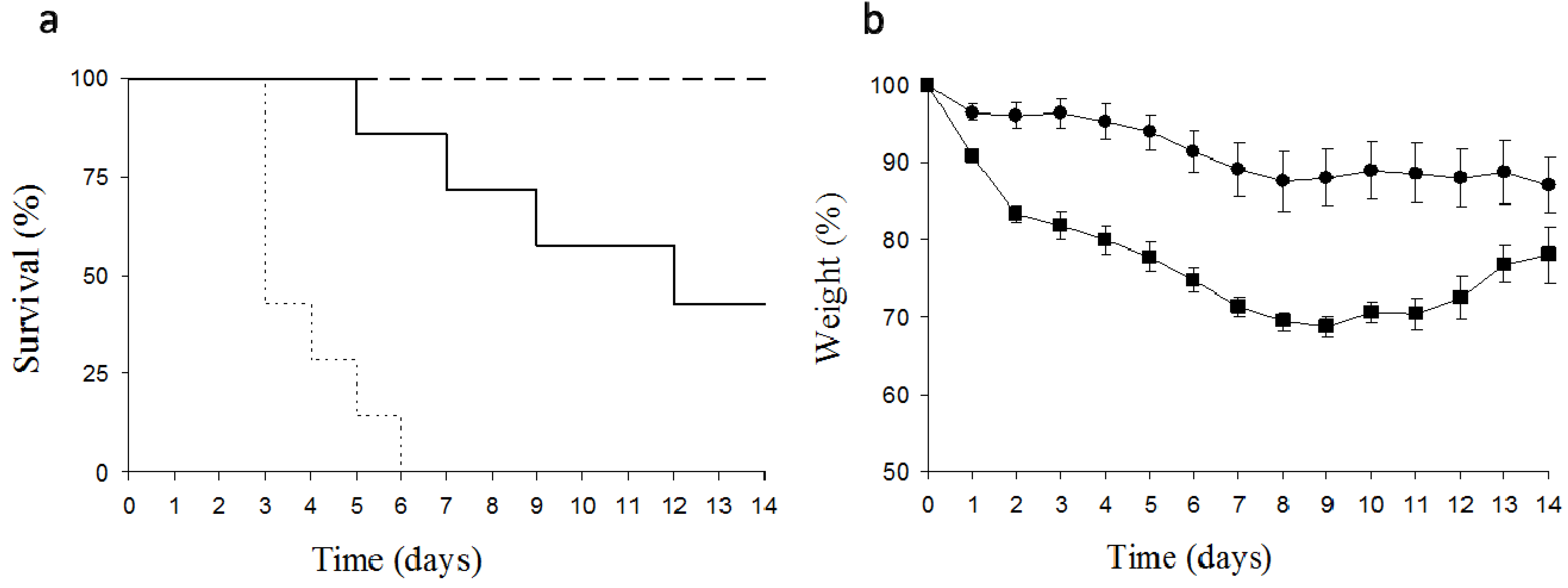
2.4. Toxicity of Dwarf Elder Blossom d-Galactose-Specific Lectins to Swiss Mice

3. Discussion
4. Experimental Section
4.1. Isolation of d-Galactose-Binding Lectins from S. ebulus Blossoms
4.2. Isolation of Crude Extracts
4.3. SDS-Polyacrylamide Gel Electrophoresis of d-Galactose-Binding Lectins from S. ebulus Blossoms
4.4. Mass-Spectrometry Analysis of the Lectins
4.5. Tryptic Fingerprinting and Database Search
4.6. Toxicity of Dwarf Elder Blossom D-Galactose-Specific Lectins
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Mariotti-Lippi, L.; Bellini, C.; Mori, S. Palaeovegetational reconstruction based on pollen and seeds/fruits from a bronze age archaeological site in Tuscany (Italy). Plant Biosyst. 2010, 144, 902–908. [Google Scholar] [CrossRef]
- Martin, L.; Jacomet, S.; Thiebault, S. Plant economy during the Neolithic in a mountain context: The case of “Le Chenet des Pierres” in the French Alps (Bozel-Savoie, France). Veg. Hist. Archaeobot. 2008, 17, s113–s122. [Google Scholar] [CrossRef]
- Veberic, R.; Jakopic, J.; Stampar, F.; Schmitzer, V. European elderberry (Sambucus nigra L.) rich in sugars, organic acids, anthocyanins and selected polyphenols. Food Chem. 2009, 114, 511–515. [Google Scholar] [CrossRef]
- Kaack, K.; Austed, T. Interaction of vitamin C and flavonoids in elderberry (Sambucus nigra L.) during juice processing. Plant Foods Hum. Nutr. 1998, 52, 187–198. [Google Scholar] [CrossRef]
- Girbes, T.; Ferreras, J.M.; Arias, F.J.; Stirpe, F. Description, distribution, activity and phylogenetic relationship of ribosome-inactivating proteins in plants, fungi and bacteria. Mini Rev. Med. Chem. 2004, 4, 461–476. [Google Scholar] [CrossRef]
- Kiselova, Y.; Ivanova, D.; Chervenkov, T.; Gerova, D.; Galunska, B.; Yankova, T. Correlation between the in vitro antioxidant activity and polyphenol content of aqueous extracts from bulgarian herbs. Phytother. Res. 2006, 20, 961–965. [Google Scholar] [CrossRef]
- Vlachojannis, J.E.; Cameron, M.; Chrubasik, S. A systematic review on the sambuci fructus effect and efficacy profiles. Phytother. Res. 2010, 24, 1–8. [Google Scholar] [CrossRef]
- Roschek, B.; Fink, R.C.; McMichael, M.D.; Li, D.; Alberte, R.S. Elderberry flavonoids bind to and prevent H1N1 infection in vitro. Phytochemistry 2009, 70, 1255–1261. [Google Scholar] [CrossRef]
- Suntar, I.P.; Akkol, E.K.; Yalcin, F.N.; Koca, U.; Keles, H.; Yesilada, E. Wound healing potential of Sambucus ebulus L. leaves and isolation of an active component, quercetin 3-O-glucoside. J. Ethnopharmacol. 2010, 129, 106–114. [Google Scholar] [CrossRef]
- Ferreras, J.M.; Citores, L.; Iglesias, R.; Jiménez, P.; Girbés, T. Use of Ribosome-inactivating proteins from Sambucus for the construction of immunotoxins and conjugates for cancer therapy. Toxins 2011, 3, 420–441. [Google Scholar]
- Sharon, N. Lectins: Past, present and future. Biochem. Soc. Trans. 2008, 36, 1457–1460. [Google Scholar] [CrossRef]
- Jiménez, P.; Cordoba-Diaz, D.; Cabrero, P.; Aracil, M.; Gayoso, M.J.; Garrosa, M.; Cordoba-Diaz, M.; Girbés, T. Plasma accumulation of vitamin B6 from an oral dose in a new gut injury-regeneration reversible mouse model. Food Nutr. Sci. 2013, 4, 908–917. [Google Scholar]
- Girbes, T.; Citores, L.; Iglesias, R.; Ferreras, J.M.; Muñoz, R.; Rojo, M.A.; Arias, F.J.; Garcia, J.R.; Mendez, E.; Calonge, M. Ebulin 1, a nontoxic novel type 2 ribosome-inactivating protein from Sambucus ebulus L. leaves. J. Biol. Chem. 1993, 268, 18195–18199. [Google Scholar]
- Pascal, J.M.; Day, P.J.; Monzingo, A.F.; Ernst, S.R.; Robertus, J.D.; Iglesias, R.; Pérez, Y.; Ferreras, J.M.; Citores, L.; Girbes, T. 2.8-A crystal structure of a nontoxic type-II ribosome-inactivating protein, ebulin l. Proteins 2001, 43, 319–326. [Google Scholar] [CrossRef]
- Citores, L.; Rojo, M.A.; Jimenez, P.; Ferreras, J.M.; Iglesias, R.; Aranguez, I.; Girbes, T. Transient occurrence of an ebulin-related d-galactose-lectin in shoots of Sambucus ebulus L. Phytochemistry 2008, 69, 857–864. [Google Scholar] [CrossRef]
- Citores, L.; de Benito, F.M.; Iglesias, R.; Miguel Ferreras, J.; Argüeso, P.; Jimenez, P.; Méndez, E.; Girbes, T. Presence of polymerized and free forms of the non-toxic Type 2 ribosome-inactivating protein ebulin and a structurally related new homodimeric lectin in fruits of Sambucus ebulus L. Planta 1998, 204, 310–319. [Google Scholar] [CrossRef]
- Stirpe, F. Ribosome-inactivating proteins. Toxicon 2004, 44, 371–383. [Google Scholar] [CrossRef]
- Rojo, M.A.; Citores, L.; Arias, F.J.; Ferreras, J.M.; Jiménez, P.; Girbés, T. Molecular cloning of a cDNA coding for the d-galactose-binding dimeric lectin of dwarf elder (Sambucus ebulus L.) leaves. Int. J. Biochem. Cell Biol. 2003, 35, 1061–1065. [Google Scholar] [CrossRef]
- Jimenez, P.; Tejero, J.; Cabrero, P.; Cordoba-Diaz, D.; Girbes, T. Differential sensitivity of d-galactose-binding lectins from fruits of dwarf elder (Sambucus ebulus L.) to a simulated gastric fluid. Food Chem. 2013, 136, 794–802. [Google Scholar] [CrossRef]
- Citores, L.; de Benito, F.M.; Iglesias, R.; Ferreras, J.M.; Argüeso, P.; Jimenez, P.; Testera, A.; Camafeita, E.; Méndez, E.; Girbes, T. Characterization of a new non-Toxic two-chain ribosome-inactivating protein and a structurally-related lectin from rhizomes of dwarf elder (Sambucus ebulus L.). Cell Mol. Biol. 1997, 43, 485–499. [Google Scholar]
- Förster-Waldl, E.; Marchetti, M.; Schöll, I.; Focke, M.; Radauer, C.; Kinaciyan, T.; Nentwich, I.; Jäger, S.; Schmid, E.R.; Boltz-Nitulescu, G.; Scheiner, O.; Jensen-Jarolim, E. Type I allergy to elderberry (Sambucus nigra) is elicited by a 33.2 kDa allergen with significant homology to ribosomal inactivating proteins. Clin. Exp. Allergy 2003, 33, 1703–1710. [Google Scholar] [CrossRef]
- Griffiths, G.D. Understanding ricin from a defensive viewpoint. Toxins 2011, 3, 1373–1392. [Google Scholar] [CrossRef]
- Bradberry, S.M.; Dickers, K.J.; Rice, P.; Griffiths, G.D.; Vale, J.A. Ricin poisoning. Toxicol Rev. 2003, 22, 65–70. [Google Scholar] [CrossRef]
- Jiménez, P.; Gayoso, M.J.; Tejero, J.; Cabrero, P.; Cordoba-Diaz, D.; Basterrechea, J.E.; Girbés, T. Toxicity in mice of lectin ebulin f present in dwarf elderberry (Sambucus ebulus L.). Toxicon 2013, 61, 26–29. [Google Scholar] [CrossRef]
- Lord, J.M.; Spooner, R.A. Ricin trafficking in plant and mammalian cells. Toxins 2011, 3, 787–801. [Google Scholar]
- Lord, M.J.; Jolliffe, N.A.; Marsden, C.J.; Pateman, C.S.; Smith, D.C.; Spooner, R.A.; Watson, P.D.; Roberts, L.M. Ricin. Mechanisms of cytotoxicity. Toxicol. Rev. 2003, 22, 53–64. [Google Scholar] [CrossRef]
- Svinth, M.; Steinghardt, J.; Hernandez, R.; Duh, J.K.; Kelly, C.; Day, P.; Lord, M.; Girbés, T.; Robertus, J.D. Differences in cytotoxicity of native and engineered RIPs can be used to assess their ability to reach the cytoplasm. Biochem. Biophys. Res. Commun. 1998, 249, 637–642. [Google Scholar] [CrossRef]
- Battelli, M.G.; Musiani, S.; Buonamici, L.; Santi, S.; Riccio, M.; Maraldi, N.M.; Girbés, T.; Stirpe, F. Interaction of volkensin with HeLa cells: Binding, uptake, intracellular localization, degradation and exocytosis. Cell Mol. Life Sci. 2004, 61, 1975–1984. [Google Scholar]
- Horrix, C.; Raviv, Z.; Flescher, E.; Voss, C.; Berger, M.R. Plant ribosome-inactivating proteins type II induce the unfolded protein response in human cancer cells. Cell Mol. Life Sci. 2011, 68, 1269–1281. [Google Scholar] [CrossRef]
- Pita, R.; Anadón, A.; Martínez-Larrañaga, M.R. Ricina: Una fitotoxina de uso potencial como arma. Revista de Toxicología 2004, 21, 51–63. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jimenez, P.; Cabrero, P.; Basterrechea, J.E.; Tejero, J.; Cordoba-Diaz, D.; Girbes, T. Isolation and Molecular Characterization of Two Lectins from Dwarf Elder (Sambucus ebulus L.) Blossoms Related to the Sam n1 Allergen. Toxins 2013, 5, 1767-1779. https://doi.org/10.3390/toxins5101767
Jimenez P, Cabrero P, Basterrechea JE, Tejero J, Cordoba-Diaz D, Girbes T. Isolation and Molecular Characterization of Two Lectins from Dwarf Elder (Sambucus ebulus L.) Blossoms Related to the Sam n1 Allergen. Toxins. 2013; 5(10):1767-1779. https://doi.org/10.3390/toxins5101767
Chicago/Turabian StyleJimenez, Pilar, Patricia Cabrero, José E. Basterrechea, Jesús Tejero, Damian Cordoba-Diaz, and Tomas Girbes. 2013. "Isolation and Molecular Characterization of Two Lectins from Dwarf Elder (Sambucus ebulus L.) Blossoms Related to the Sam n1 Allergen" Toxins 5, no. 10: 1767-1779. https://doi.org/10.3390/toxins5101767