Faces of a Changing Climate: Semi-Quantitative Multi-Mycotoxin Analysis of Grain Grown in Exceptional Climatic Conditions in Norway

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
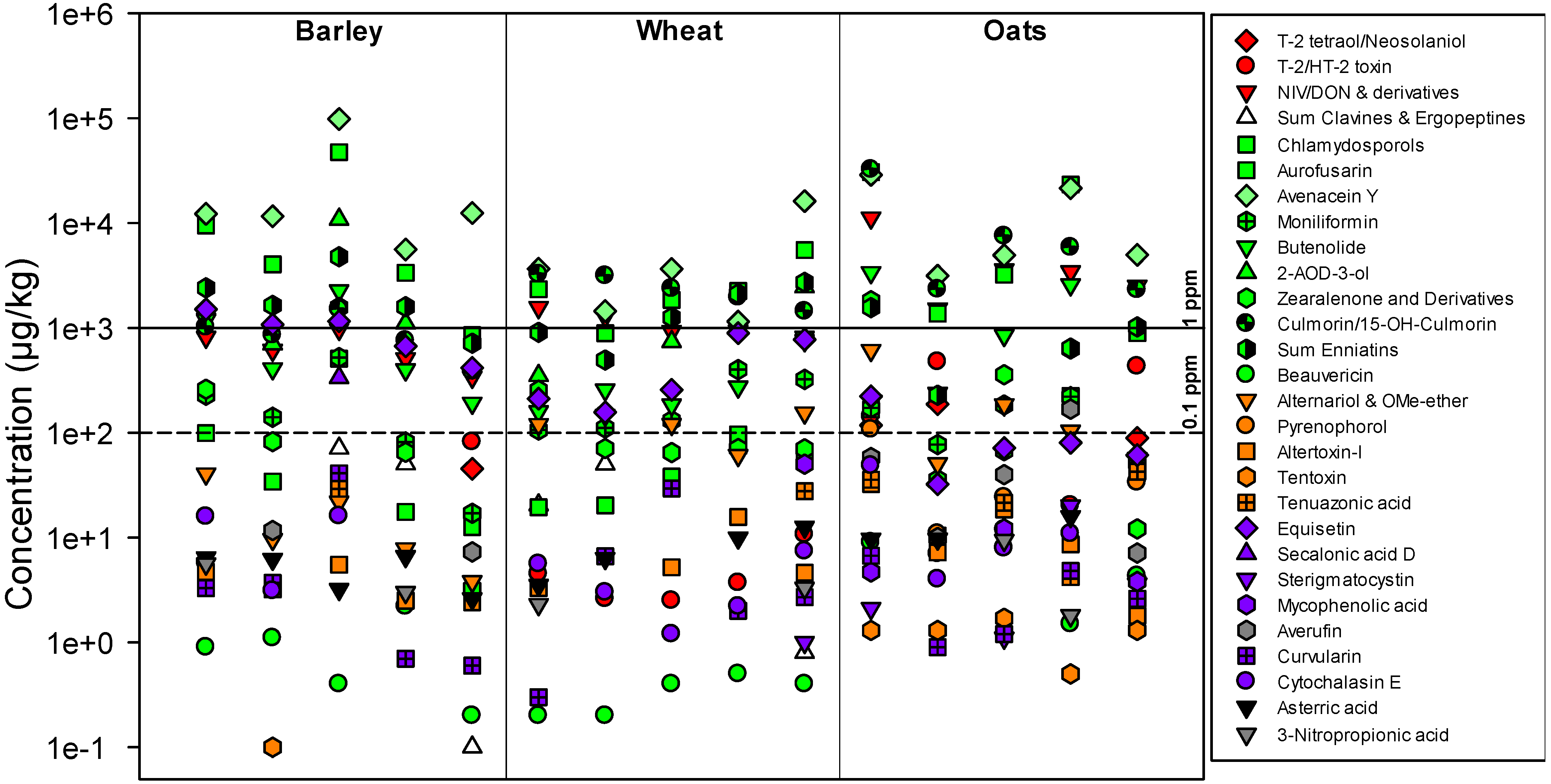
2.1. Methodological Considerations
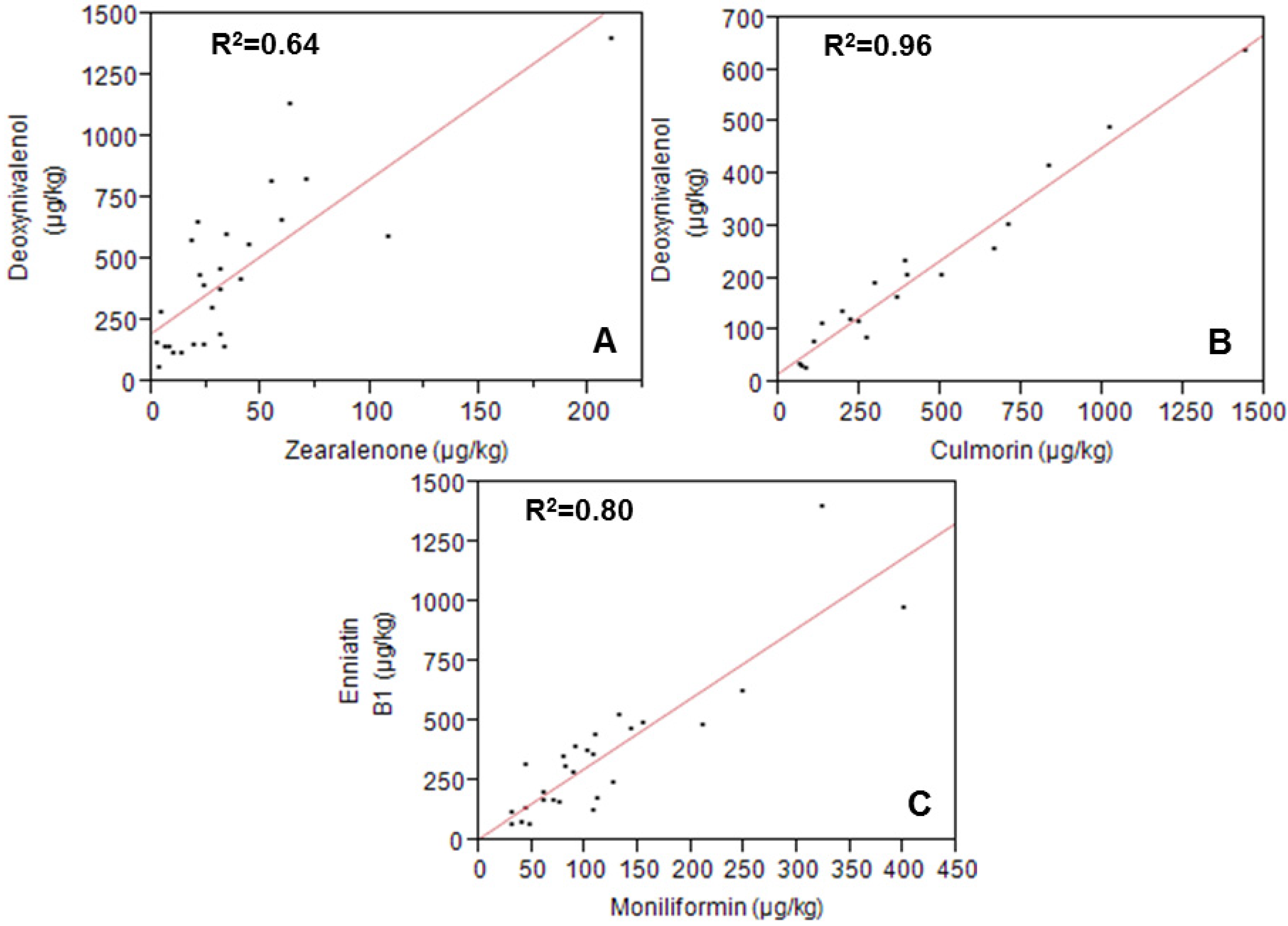
2.2. Metabolites Related to Fusarium
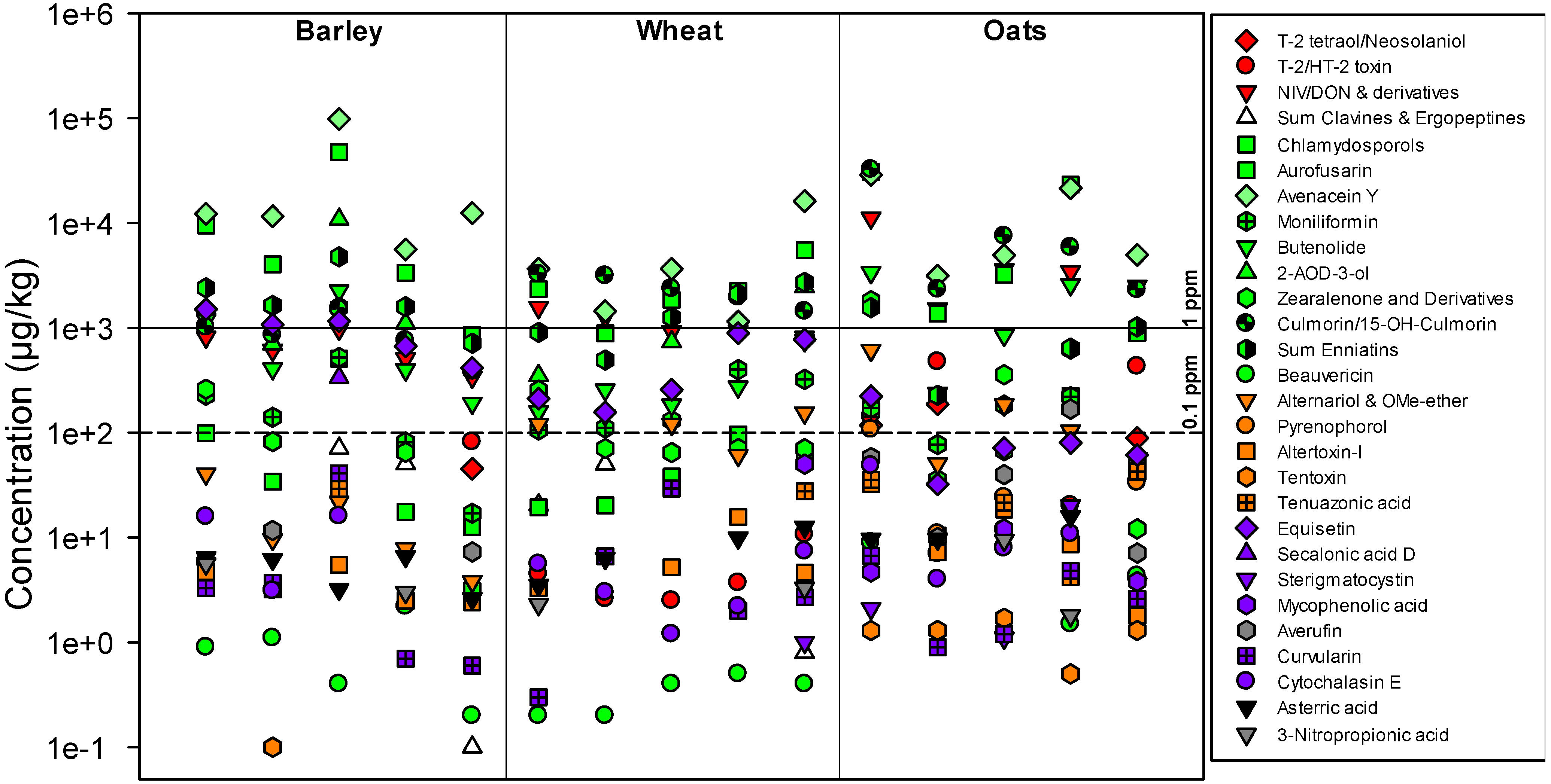

{kind=link}
{kind=link}
{kind=link}
| Barley (n = 20) | Oats (n = 28) | Wheat (n = 28) | ||||||||
| Metabolite | Positive (%) | Median (µg/kg) | Maximum (µg/kg) | Positive (%) | Median (µg/kg) | Maximum (µg/kg) | Positive (%) | Median (µg/kg) | Maximum (µg/kg) | |
| Type-A trichothecenes | T-2 tetraol | 30 | 12.9 | 43.1 | 79 | 54.7 | 174 | 0 | - | - |
| T-2 toxin | 30 | 5.1 | 14.1 | 96 | 20.1 | 143 | 79 | 3.4 | 5.3 | |
| HT-2 toxin | 25 | 14.2 | 67.6 | 82 | 92.8 | 333 | 4 | 6.1 | 6.1 | |
| Neosolaniol | 5 | 2.2 | 2.2 | 46 | 3.4 | 13.8 | 0 | - | - | |
| Type-B trichothecenes | Nivalenol | 55 | 2.9 | 13.6 | 93 | 5.2 | 45.5 | 14 | 2.2 | 2.4 |
| Deoxynivalenol | 100 | 150 | 636 | 100 | 1070 | 7230 | 100 | 383 | 1400 | |
| DON-3-glucoside | 100 | 67.8 | 270 | 100 | 252 | 2580 | 100 | 56.4 | 152 | |
| 3-Acetyl-DON | 60 | 17.8 | 141 | 100 | 128 | 1380 | 68 | 14.0 | 49.5 | |
| Depsipeptides | Enniatin A | 100 | 17.1 | 185 | 100 | 3.7 | 30.0 | 100 | 4.1 | 92.7 |
| Enniatin A1 | 100 | 145 | 1,180 | 100 | 21.4 | 263 | 100 | 48.0 | 276 | |
| Enniatin B | 100 | 440 | 807 | 100 | 69.6 | 662 | 100 | 347 | 874 | |
| Enniatin B1 | 100 | 529 | 2,820 | 100 | 65.5 | 706 | 100 | 296 | 1400 | |
| Enniatin B2 | 100 | 23.9 | 133 | 100 | 3.2 | 24.5 | 100 | 15.1 | 79.3 | |
| Enniatin B3 | 100 | 0.2 | 1.0 | 100 | 0.05 | 0.21 | 100 | 0.16 | 0.62 | |
| Beauvericin | 100 | 0.4 | 2.2 | 100 | 3.5 | 15.1 | 100 | 0.4 | 1.1 | |
| Zearalenone and related compounds | Zearalenone | 95 | 11.4 | 1340 | 93 | 89.9 | 1670 | 96 | 27.5 | 210 |
| β-Zearalenol | 35 | 6.8 | 158 | 79 | 7.2 | 97.2 | 36 | 5.4 | 18.2 | |
| ZEN-4-sulphate | 55 | 3.1 | 76.7 | 93 | 2.6 | 43.5 | 82 | 1.1 | 18.2 | |
| Other Fusarium metabolites | Chlamydosporols | 75 | 26.1 | 509 | 61 | 33.7 | 225 | 86 | 16.7 | 96.2 |
| Aurofusarin | 100 | 1090 | 47,300 | 100 | 1,350 | 30,500 | 100 | 934 | 5510 | |
| Avenacein Y | 90 | 6030 | 98,100 | 100 | 4630 | 28,800 | 100 | 1520 | 16,200 | |
| Moniliformin | 100 | 86.0 | 522 | 100 | 57.2 | 220 | 100 | 88.4 | 400 | |
| Butenolide | 100 | 231 | 2260 | 97 | 154 | 3370 | 86 | 198 | 818 | |
| Culmorin | 95 | 292 | 1440 | 100 | 2000 | 31,500 | 100 | 986 | 3160 | |
| 15-Hydroxy-culmorin | 40 | 49.5 | 70.9 | 71 | 117 | 924 | 36 | 70.6 | 105 | |
| 2-AOD-3-ol | 35 | 1100 | 10,800 | 0 | - | - | 36 | 532 | 2460 | |
| Equisetin | 100 | 433 | 2,470 | 100 | 56.3 | 311 | 100 | 204 | 890 | |
| Barley | Oats | Wheat | ||||||||
| Metabolite | Positive (%) | Median (µg/kg) | Maximum (µg/kg) | Positive (%) | Median (µg/kg) | Maximum (µg/kg) | Positive (%) | Median (µg/kg) | Maximum (µg/kg) | |
| Alternaria metabolites | Alternariol | 80 | 10.4 | 37.7 | 93 | 53.6 | 449 | 100 | 116 | 305 |
| Alternariol-methylether | 95 | 0.5 | 5.2 | 100 | 21.6 | 177 | 100 | 0.8 | 2.5 | |
| Tenuazonic acid | 15 | 59 | 247 | 89 | 21 | 82 | 21 | 18 | 35 | |
| Altertoxin-I | 95 | 2.7 | 5.6 | 96 | 7.3 | 36.1 | 96 | 4.5 | 15.7 | |
| Tentoxin | 25 | 0.2 | 0.4 | 93 | 1.3 | 3.6 | 0 | - | - | |
| Pyrenophorol | 0 | - | - | 93 | 29.4 | 108 | 0 | - | - | |
| Ergot alkaloids | Chanoclavine | 40 | 0.1 | 0.2 | 0 | - | - | 18 | 0.1 | 0.9 |
| Ergometrine/-metrinine | 10 | 0.6 | 0.6 | 0 | - | - | 11 | 0.6 | 4.9 | |
| Ergocristine/-cristinine | 25 | 20.2 | 68.3 | 0 | - | - | 4 | 44.2 | 44.2 | |
| Ergocornine/-corninine | 12 | 2.5 | 10.3 | 4 | 0.2 | 0.2 | 21 | 4.2 | 55.0 | |
| α-Ergocryptine/-cryptinine | 15 | 25.5 | 58.7 | 4 | 0.3 | 0.3 | 21 | 8.6 | 78.9 | |
| Penicillium and Aspergillus metabolites | Sterigmatocystin | 15 | 1.0 | 1.2 | 57 | 2.1 | 20.1 | 7 | 1.0 | 1.0 |
| Mycophenolic acid | 10 | 31.6 | 56.2 | 25 | 8.7 | 13.5 | 36 | 12.8 | 166.7 | |
| Averufin | 35 | 7.3 | 25.6 | 82 | 32.0 | 168 | 11 | 10.4 | 72.0 | |
| Cytochalasin E | 40 | 4.1 | 16.1 | 71 | 9.1 | 48.6 | 55 | 5.6 | 38.2 | |
| Asterric acid | 60 | 6.7 | 28.6 | 69 | 9.7 | 39.7 | 54 | 5.3 | 12.6 | |
| Other fungal metabolites | Curvularin | 60 | 1.8 | 41.1 | 79 | 2.5 | 10.7 | 71 | 2.4 | 29.4 |
| 3-Nitropropionic acid | 15 | 3.0 | 5.6 | 11 | 2.0 | 9.4 | 36 | 4.1 | 22.8 | |
| Emodin | 100 | 15.6 | 75.3 | 100 | 22.3 | 111 | 100 | 5.4 | 71.5 | |
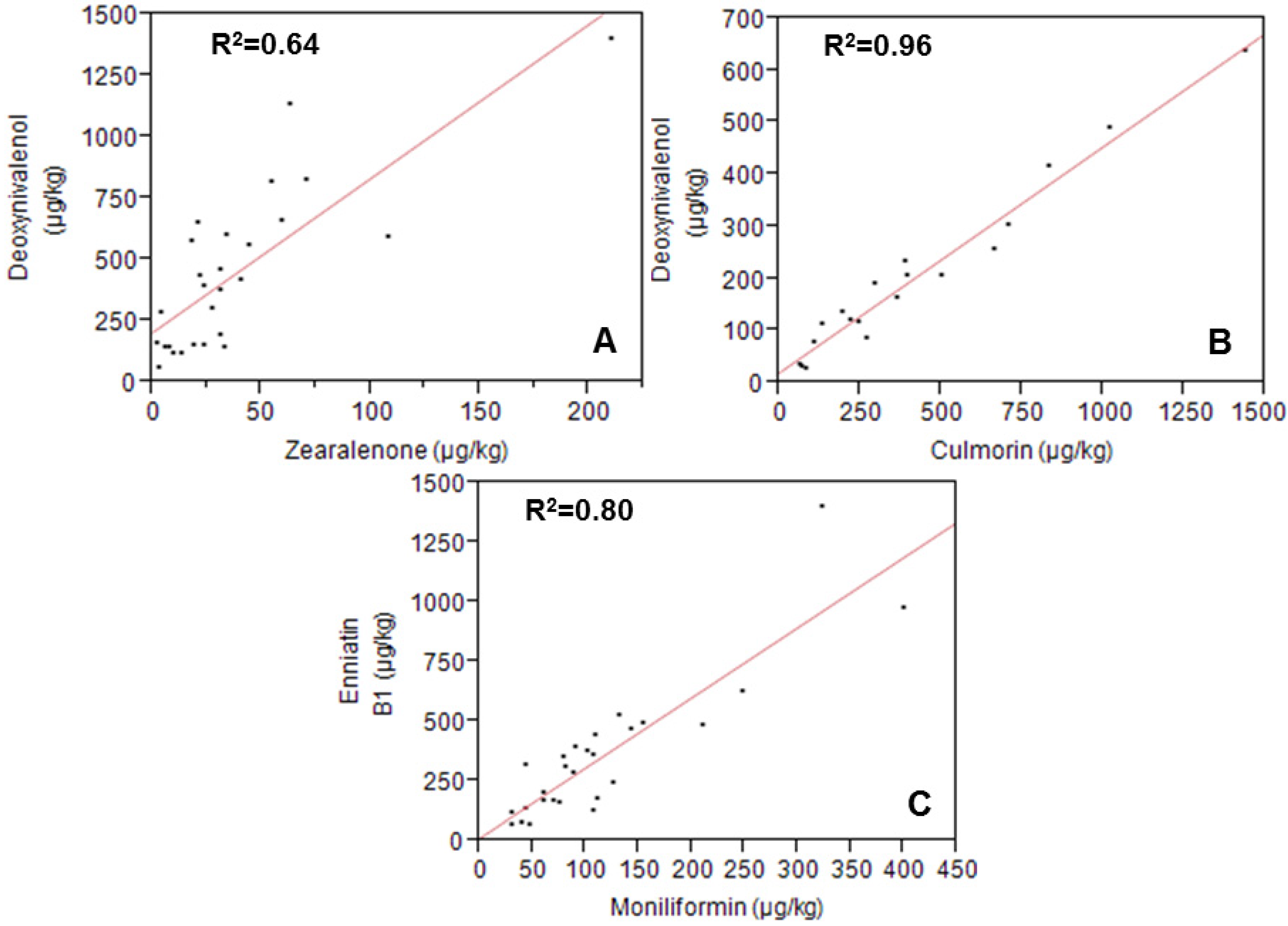
2.3. Metabolites Related to Alternaria
2.4. Ergot Alkaloids
2.5. Metabolites Related to Penicillium and Aspergillus
2.6. Toxicological Considerations
3. Experimental Section
3.1. Chemicals and Reagents
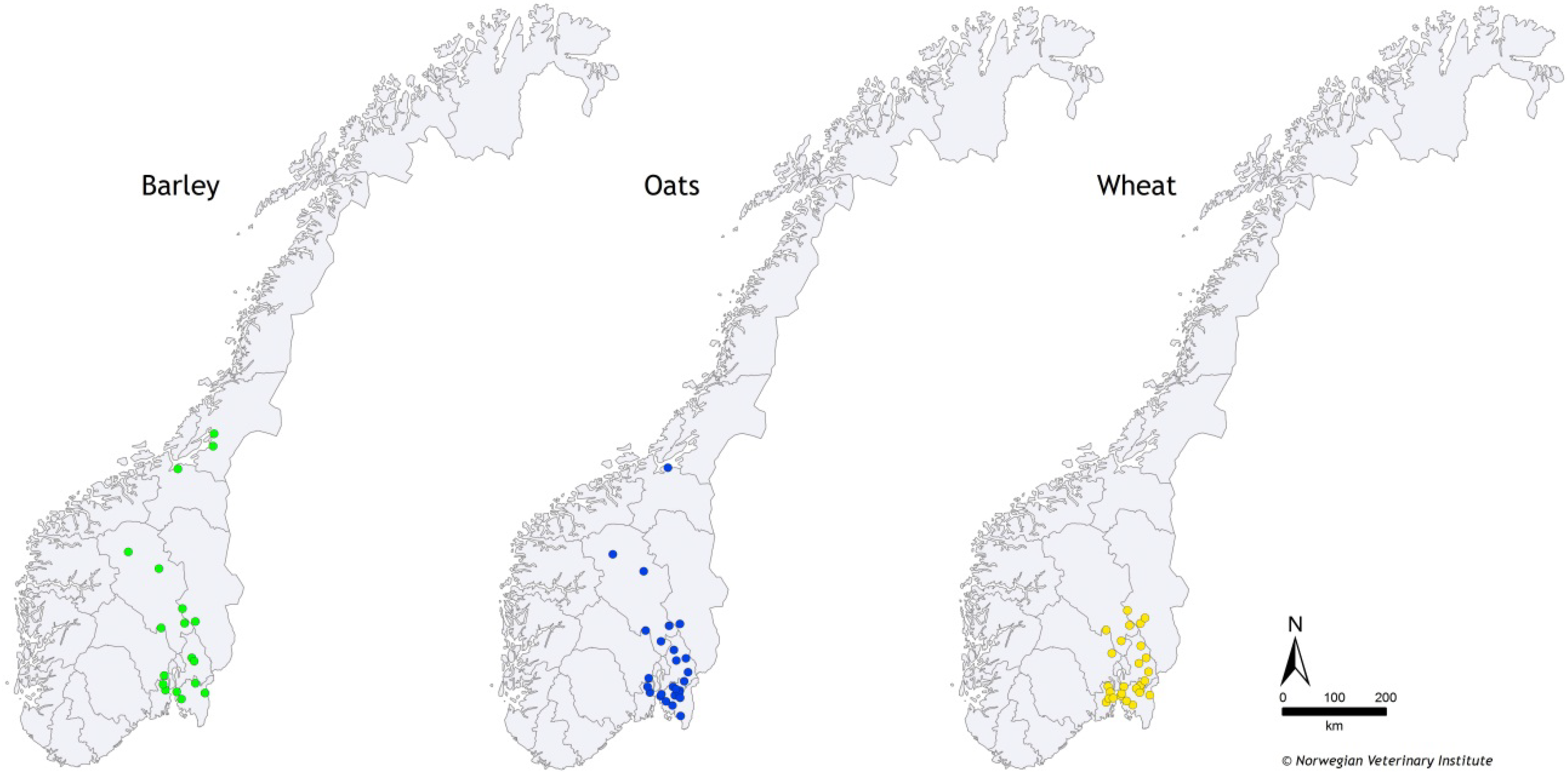
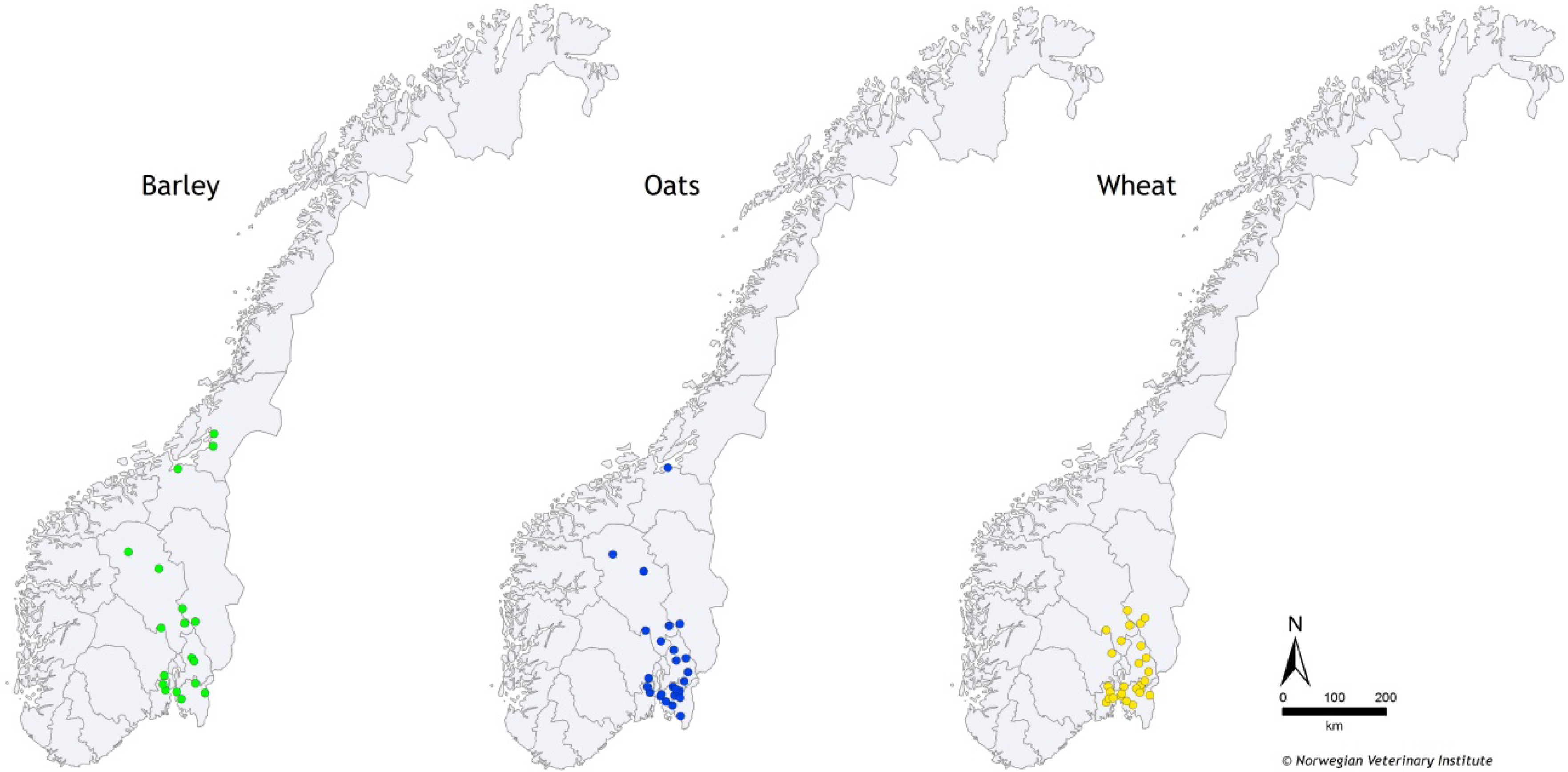
3.2. Samples
3.3. Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Været i Norge Klimatologisk oversikt Året 2011. Available online: http://met.no/Klima/Klimastatistikk/Varet_i_Norge/2011/Aret_2011/filestore/20112.pdf (accessed on 20 August 2013).
- Bernhoft, A.; Eriksen, G.S.; Sundheim, L.; Berntssen, M.; Brantsæter, A.L.; Brodal, G.; Fæste, C.K.; Hofgaard, I.S.; Rafoss, T.; Sivertsen, T.; et al. Risk Assessment of Mycotoxins in Cereal Grain in Norway. Opinion of the Scientific Steering Committee of the Norwegian Scientific Committee for Food Safety: Oslo, Norway, 2013. [Google Scholar]
- Brodal, G.; Rafoss, T.; Elen, O.; Tangerås, H. Trends and variations in the occurrence of Fusarium in cereal seeds in Norway 1970-2010. Seed Sci. Technol. 2013. submitted for publication. [Google Scholar]
- Abia, W.A.; Warth, B.; Sulyok, M.; Krska, R.; Tchana, A.N.; Njobeh, P.B.; Dutton, M.F.; Moundipa, P.F. Determination of multi-mycotoxin occurrence in cereals, nuts and their products in Cameroon by liquid chromatography tandem mass spectrometry (LC-MS/MS). Food Control 2013, 31, 438–453. [Google Scholar] [CrossRef]
- Sulyok, M.; Berthiller, F.; Krska, R.; Schumacher, R. Development and validation of a liquid chromatography/tandem mass spectrometric method for the determination of 39 mycotoxins in wheat and maize. Rapid Commun. Mass Spectrom. 2006, 20, 2649–2659. [Google Scholar] [CrossRef]
- Sulyok, M.; Krska, R.; Schuhmacher, R. Application of an LC-MS/MS based multi-mycotoxin method for the semi-quantitative determination of mycotoxins occurring in different types of food infected by moulds. Food Chem. 2010, 119, 408–416. [Google Scholar] [CrossRef]
- Vishwanath, V.; Sulyok, M.; Labuda, R.; Bicker, W.; Krska, R. Simultaneous determination of 186 fungal and bacterial metabolites in indoor matrices by liquid chromatography/tandem mass spectrometry. Anal. Bioanal. Chem. 2009, 395, 1355–1372. [Google Scholar] [CrossRef]
- Bernhoft, A.; Clasen, P.E.; Kristoffersen, A.B.; Torp, M. Less Fusarium infestation and mycotoxin contamination in organic than in conventional cereals. Food Addit. Contam. Part A 2010, 27, 842–852. [Google Scholar] [CrossRef]
- Uhlig, S.; Jestoi, M.; Parikka, P. Fusarium avenaceum—The North European situation. Int. J. Food Microbiol. 2007, 119, 17–24. [Google Scholar] [CrossRef]
- Chandelier, A.; Nimal, C.; Andre, F.; Planchon, V.; Oger, R. Fusarium species and DON contamination associated with head blight in winter wheat over a 7-year period (2003–2009) in Belgium. Eur. J. Plant Pathol. 2011, 130, 403–414. [Google Scholar] [CrossRef]
- Nielsen, L.K.; Jensen, J.D.; Rodriguez, A.; Jorgensen, L.N.; Justesen, A.F. TRI12 based quantitative real-time PCR assays reveal the distribution of trichothecene genotypes of F. graminearum and F. culmorum isolates in Danish small grain cereals. Int. J. Food Microbiol. 2012, 157, 384–392. [Google Scholar] [CrossRef]
- Waalwijk, C.; Kastelein, P.; de Vries, I.; Kerenyi, Z.; van der Lee, T.; Hesselink, T.; Kohl, J.; Kema, G. Major changes in Fusarium spp. in wheat in the Netherlands. Eur. J. Plant Pathol. 2003, 109, 743–754. [Google Scholar] [CrossRef]
- Streit, E.; Naehrer, K.; Rodrigues, I.; Schatzmayr, G. Mycotoxin occurrence in feed and feed raw materials worldwide: Long-term analysis with special focus on Europe and Asia. J. Sci. Food Agric. 2013, 93, 2892–2899. [Google Scholar] [CrossRef]
- Van der Fels-Klerx, H.J.; Klemsdal, S.; Hietaniemi, V.; Lindblad, M.; Ioannou-Kakouri, E.; van Asselt, E.D. Mycotoxin contamination of cereal grain commodities in relation to climate in North West Europe. Food Addit. Contam. Part A 2012, 29, 1581–1592. [Google Scholar] [CrossRef]
- Brandfass, C. Establishment and Application of Real-Time PCR-Based Methods to Study the Epidemiology of Fusarium Head Blight. Ph.D. thesis, Georg-August-University Göttingen, Göttingen, Germany, 2006. [Google Scholar]
- Rodrigues, I.; Naehrer, K. A three-year survey on the worldwide occurrence of mycotoxins in feedstuffs and feed. Toxins 2012, 4, 663–675. [Google Scholar] [CrossRef]
- Fredlund, E.; Gidlund, A.; Sulyok, M.; Börjesson, T.; Krska, R.; Olsen, M.; Lindblad, M. Deoxynivalenol and other selected Fusarium toxins in Swedish oats—Occurrence and correlation to specific Fusarium species. Int. J. Food Microbiol. 2013. [Google Scholar] [CrossRef]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; de Saeger, S.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef] [Green Version]
- Edwards, S.G. Fusarium mycotoxin content of UK organic and conventional barley. Food Addit. Contam. Part A 2009, 26, 1185–1190. [Google Scholar] [CrossRef]
- Ghebremeskel, M.; Langseth, W. The occurrence of culmorin and hydroxy-culmorins in cereals. Mycopathologia 2001, 152, 103–108. [Google Scholar] [CrossRef]
- Santini, A.; Meca, G.; Uhlig, S.; Ritieni, A. Fusaproliferin, beauvericin and enniatins: Occurrence in food—A review. World Mycotoxin J. 2012, 5, 71–81. [Google Scholar] [CrossRef]
- Jestoi, M.; Rokka, M.; Yli-Mattila, T.; Parikka, P.; Rizzo, A.; Peltonen, K. Presence and concentrations of the Fusarium-related mycotoxins beauvericin, enniatins and moniliformin in Finnish grain samples. Food Addit. Contam. 2004, 21, 794–802. [Google Scholar] [CrossRef]
- Uhlig, S.; Torp, M.; Heier, B.T. Beauvericin and enniatins A, A1, B and B1 in Norwegian grain: A survey. Food Chem. 2006, 94, 193–201. [Google Scholar] [CrossRef]
- Uhlig, S.; Petersen, D.; Flåøyen, A.; Wilkins, A. 2-Amino-14,16-dimethyloctadecan-3-ol, a new sphingosine analogue toxin in the fungal genus Fusarium. Toxicon 2005, 46, 513–522. [Google Scholar] [CrossRef]
- EFSA panel on contaminants in the food chain. Scientific opinion on the risks for animal and public health related to the presence of Alternaria toxins in feed and food. EFSA J. 2011, 9, 2407–2505. [Google Scholar]
- Häggblom, P.; Stepinska, A.; Solyakov, A. Alternaria Mycotoxins in Swedish Feed Grain. In Proceedings of the 29th Mycotoxin-Workshop of the Society of Mycotoxin Research, Fellbach, Germany, May 2007; p. 35.
- Müller, M.E.H.; Steier, I.; Koppen, R.; Siegel, D.; Proske, M.; Korn, U.; Koch, M. Cocultivation of phytopathogenic Fusarium and Alternaria strains affects fungal growth and mycotoxin production. J. Appl. Microbiol. 2012, 113, 874–887. [Google Scholar] [CrossRef]
- Flieger, M.; Wurst, M.; Shelby, R. Ergot alkaloids—Sources, structures and analytical methods. Folia Microbiol. 1997, 42, 3–30. [Google Scholar] [CrossRef]
- Kawai, K.; Nozawa, Y.; Maebayashi, Y.; Yamazaki, M.; Hamasaki, T. Averufin, an anthraquinone mycotoxin possessing a potent uncoupling effect on mitochondrial respiration. Appl. Environ. Microbiol. 1984, 47, 481–483. [Google Scholar]
- Udagawa, T.; Yuan, J.; Panigrahy, D.; Chang, Y.H.; Shah, J.; D’Amato, R.J. Cytochalasin E, an epoxide containing Aspergillus-derived fungal metabolite, inhibits angiogenesis and tumor growth. J. Pharmacol. Exp. Ther. 2000, 294, 421–427. [Google Scholar]
- Regueira, T.B.; Kildegaard, K.R.; Hansen, B.G.; Mortensen, U.H.; Hertweck, C.; Nielsen, J. Molecular basis for mycophenolic acid biosynthesis in Penicillium brevicompactum. Appl. Environ. Microbiol. 2011, 77, 3035–3043. [Google Scholar] [CrossRef]
- EFSA panel on contaminants in the food chain. Opinion of the scientific panel on contaminants in the food chain related to deoxynivalenol (DON) as undesirable substance in animal feed. EFSA J. 2004, 73, 1–43. [Google Scholar]
- Berthiller, F.; Schuhmacher, R.; Adam, G.; Krska, R. Formation, determination and significance of masked and other conjugated mycotoxins. Anal. Bioanal. Chem. 2009, 395, 1243–1252. [Google Scholar] [CrossRef]
- World Health Organization; Food and Agriculture Organization. Deoxynivalenol. WHO Tech. Rep. Ser. 2010, 959, 37–48. [Google Scholar]
- Nagl, V.; Schwartz, H.; Krska, R.; Moll, W.D.; Knasmüller, S.; Ritzmann, M.; Adam, G.; Berthiller, F. Metabolism of the masked mycotoxin deoxynivalenol-3-glucoside in rats. Toxicol. Lett. 2012, 213, 367–373. [Google Scholar] [CrossRef]
- Berthiller, F.; Krska, R.; Domig, K.J.; Kneifel, W.; Juge, N.; Schuhmacher, R.; Adam, G. Hydrolytic fate of deoxynivalenol-3-glucoside during digestion. Toxicol. Lett. 2011, 206, 264–267. [Google Scholar] [CrossRef]
- Tran, S.T.; Smith, T.K. Determination of optimal conditions for hydrolysis of conjugated deoxynivalenol in corn and wheat with trifluoromethanesulfonic acid. Anim. Feed Sci. Technol. 2011, 163, 84–92. [Google Scholar] [CrossRef]
- Ivanova, L.; Egge-Jacobsen, W.M.; Solhaug, A.; Thoen, E.; Fæste, C.K. Lysosomes as a possible target of enniatin B-induced toxicity in Caco-2 cells. Chem. Res. Toxicol. 2012, 25, 1662–1674. [Google Scholar] [CrossRef]
- Gammelsrud, A.; Solhaug, A.; Dendele, B.; Sandberg, W.J.; Ivanova, L.; Kocbach Bolling, A.; Lagadic-Gossman, D.; Refsnes, M.; Becher, R.; Eriksen, G.S.; et al. Enniatin B-induced cell death and inflammatory responses in RAW 267.4 murine macrophages. Toxicol. Appl. Pharmacol. 2012, 261, 74–87. [Google Scholar] [CrossRef]
- Müller, M.E.H.; Korn, U. Alternaria mycotoxins in wheat—A 10 years survey in the Northeast of Germany. Food Control 2013, 34, 197–198. [Google Scholar]
- Jaksic, D.; Puel, O.; Canlet, C.; Kopjar, N.; Kosalec, I.; Klaric, M.S. Cytotoxicity and genotoxicity of versicolorins and 5-methoxysterigmatocystin in A549 cells. Arch. Toxicol. 2012, 86, 1583–1591. [Google Scholar] [CrossRef]
- Fehr, M.; Pahlke, G.; Fritz, J.; Christensen, M.O.; Boege, F.; Altemoller, M.; Podlech, J.; Marko, D. Alternariol acts as a topoisomerase poison, preferentially affecting the IIalpha isoform. Mol. Nutr. Food Res. 2009, 53, 441–451. [Google Scholar] [CrossRef]
- Solhaug, A.; Holme, J.A.; Haglund, K.; Dendele, B.; Sergent, O.; Pestka, J.; Lagadic-Gossmann, D.; Eriksen, G.S. Alternariol induces abnormal nuclear morphology and cell cycle arrest in murine RAW 264.7 macrophages. Toxicol. Lett. 2013, 219, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Solhaug, A.; Vines, L.L.; Ivanova, L.; Spilsberg, B.; Holme, J.A.; Pestka, J.; Collins, A.; Eriksen, G.S. Mechanisms involved in alternariol-induced cell cycle arrest. Mutat. Res. 2012, 738/739, 1–11. [Google Scholar] [CrossRef]
- Instructions for Sampling of Feedstuffs; Norwegian Food Safety Authority: Oslo, Norway, 2005.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Uhlig, S.; Eriksen, G.S.; Hofgaard, I.S.; Krska, R.; Beltrán, E.; Sulyok, M. Faces of a Changing Climate: Semi-Quantitative Multi-Mycotoxin Analysis of Grain Grown in Exceptional Climatic Conditions in Norway. Toxins 2013, 5, 1682-1697. https://doi.org/10.3390/toxins5101682
Uhlig S, Eriksen GS, Hofgaard IS, Krska R, Beltrán E, Sulyok M. Faces of a Changing Climate: Semi-Quantitative Multi-Mycotoxin Analysis of Grain Grown in Exceptional Climatic Conditions in Norway. Toxins. 2013; 5(10):1682-1697. https://doi.org/10.3390/toxins5101682
Chicago/Turabian StyleUhlig, Silvio, Gunnar Sundstøl Eriksen, Ingerd Skow Hofgaard, Rudolf Krska, Eduardo Beltrán, and Michael Sulyok. 2013. "Faces of a Changing Climate: Semi-Quantitative Multi-Mycotoxin Analysis of Grain Grown in Exceptional Climatic Conditions in Norway" Toxins 5, no. 10: 1682-1697. https://doi.org/10.3390/toxins5101682