Multimodal Protein Constructs for Herbivore Insect Control

Abstract
:1. Introduction
2. Transgene Stacking and Pesticidal Protein Pyramiding in Plants
2.1. Multimodal Fusion Proteins
2.1.1. Protein Fusions for Insect Control

{kind=link}
{kind=link}
| Fusion partners | Intended uses / Improved effects | Refs. |
|---|---|---|
| Hybrid proteins | ||
| Cry1Ab toxin modified with domain III of Cry1C | Improved efficacy against Spodoptera exigua | [63] |
| Cry1Ab toxin modified with C-terminal region of Cry1Ac | Improved efficacy and range against Lepidoptera | [29] |
| Cry1Ab toxin modified with part of Cry3A variable region | Resistance to insects of different orders | [64] |
| Cry1Ac or Cry1E modified with domain III of Cry1C | Improved efficacy against Spodoptera exigua | [65] |
| Cry1Ba toxin modified with domain II of Cry1Ia | Resistance to insects of different orders | [31,66] |
| Cry1Ca, Cry1Fb and Cry1Ba modified with Cry1Ac domain III | Improved efficacy against Heliothis virescens | [67] |
| Cry1Ea toxin modified with part of Cry1Ca toxin domain III | Improved efficacy against Spodoptera litura | [68] |
| Cry hybrid SN19 modified with domain II of Cry1Ba | Resistance to insects of different orders | [69] |
| Sunflower multicystatin integrating gourd trypsin inhibitor | Broader inhibition of Lepidoptera midgut proteases | [70] |
| Bi- or multimodal translational fusions | ||
| Cry1B and Cry1Ab toxins | Durability and broader range against Lepidoptera | [30] |
| Cry1Ac toxin and galactose-binding domain of ricin B chain | Improved efficacy and broader insecticidal range | [71] |
| Cry1Ac toxin and cowpea trypsin inhibitor CpTI | Dual effect against cabbage worms and durability | [37] |
| Cry1Ac toxin and spider venom neurotoxin HWTX-I | Dual effect against Plutella xylostella | [72] |
| Cry1Ac toxin and Beauveria bassiana subtilisin CDEP2 | Dual effect against Helicoverpa armigera | [73] |
| Cry1Ac toxin and baculoviral polyhedrin | Increased stability and expression in E. coli | [74] |
| Cry1Ab toxin C-ter peptide and spider toxin ACTX-Ar1 | Improved efficacy and range against Lepidoptera | [75,76,77,78] |
| Snowdrop lectin and arthropod peptide toxins | Delivery of toxic peptides to the haemolymph | [79,80,81,82,83] |
| Soybean cystatin N2 and GSII lectin | Dual effect against Callosobruchus maculatus | [84] |
| Various plant and animal cysteine protease inhibitors | Broader inhibition of Thysanoptera midgut proteases | [85] |
| Tomato cathepsin D inhibitor and corn cystatin II | Broader inhibition of Coleoptera midgut proteases | [86] |
| Oryzacystatin I and potato carboxypeptidase inhibitor domains | Broader inhibition of Coleoptera midgut proteases | [87] |
2.1.2. Non-Cleavable Linker Peptides
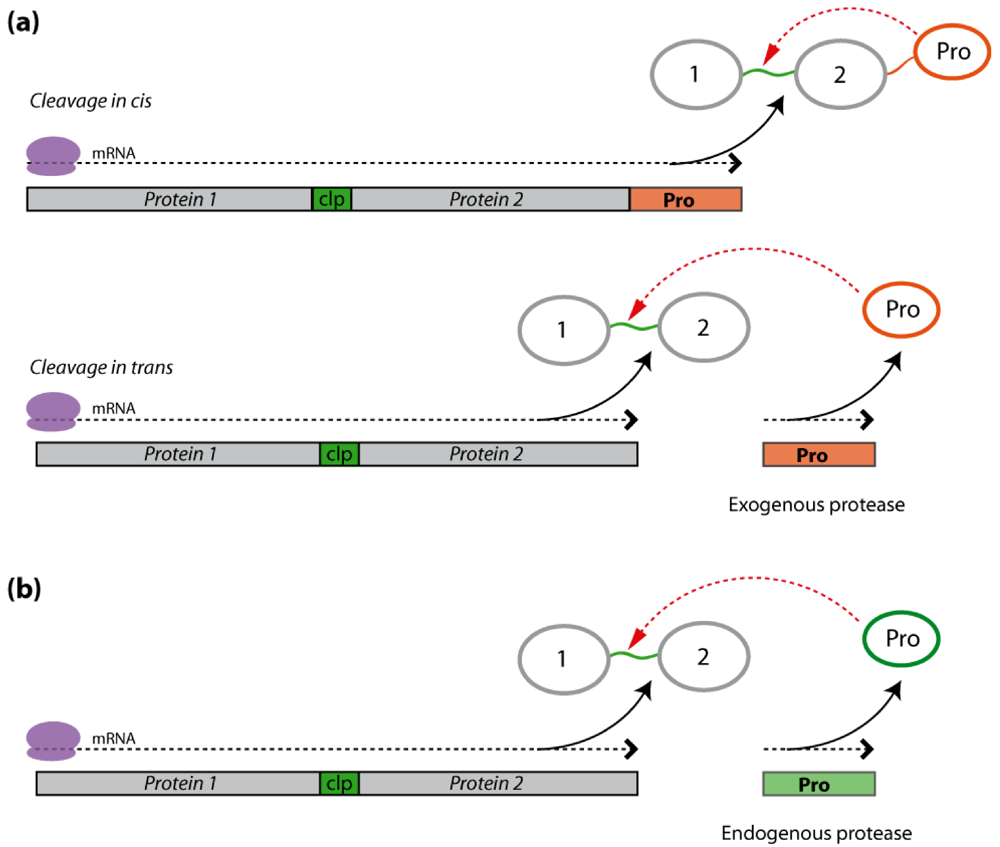
2.2. Polyprotein Precursors
2.2.1. Exogenous Protease Cleavage
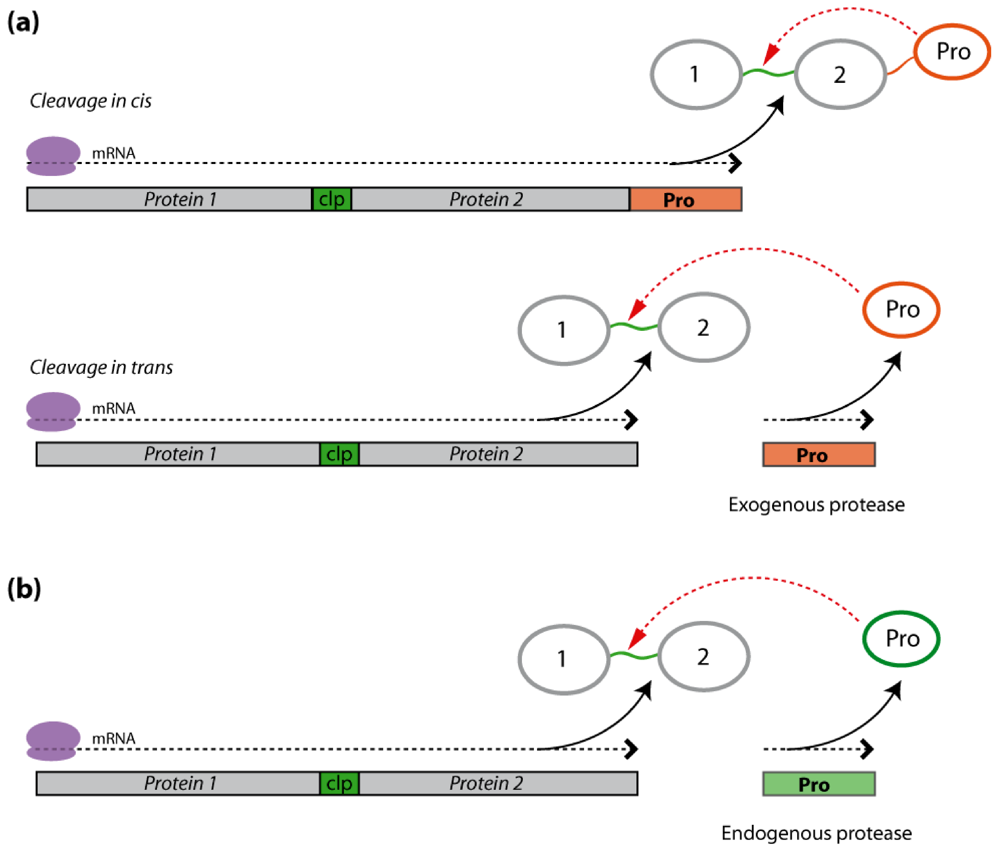
2.2.2. Endogenous Protease Cleavage
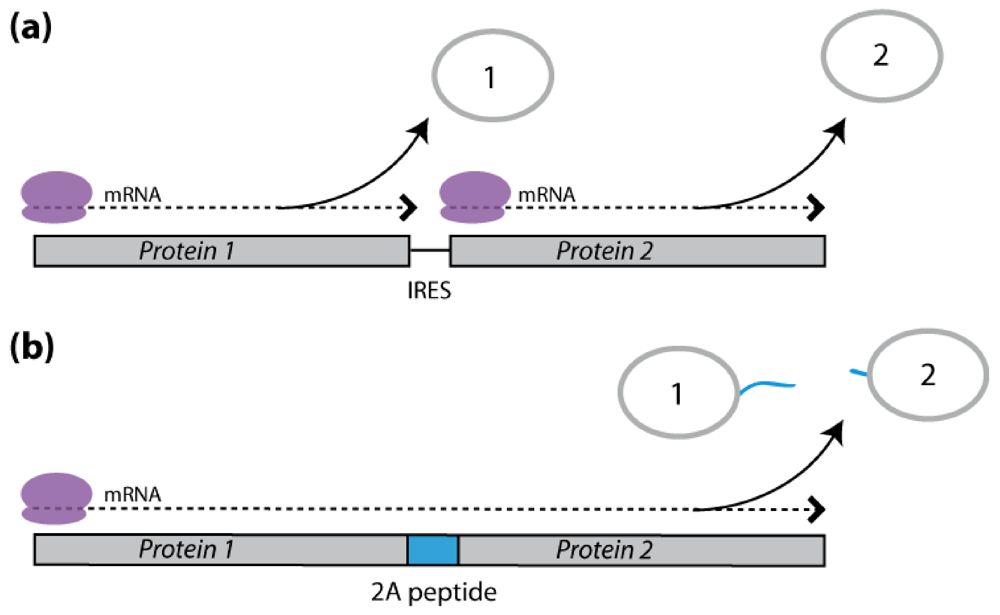
2.3. Polycistronic Constructs
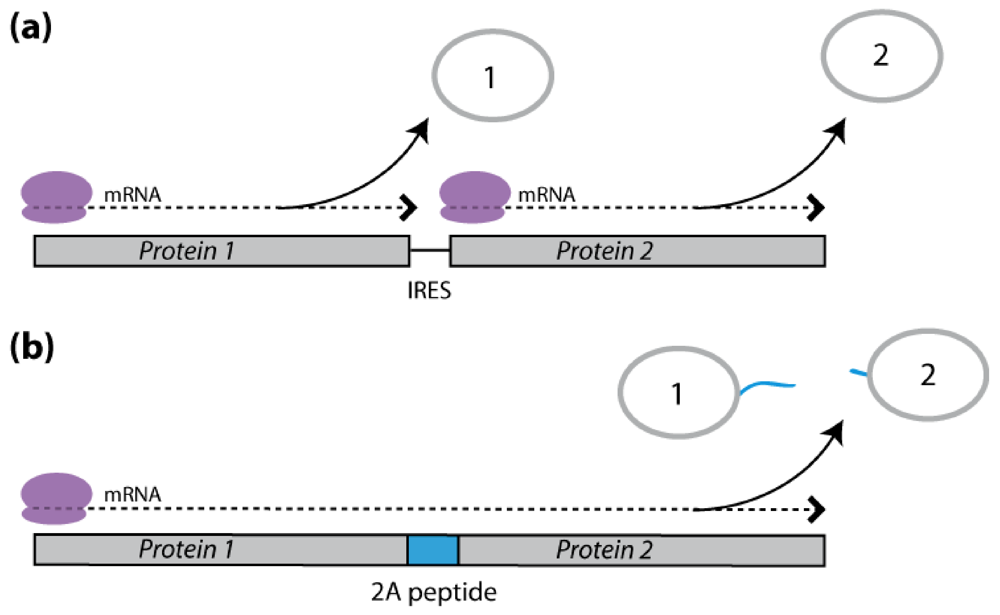
2.3.1. IRES-Mediated mRNA Translation
2.3.2. 2A Peptide-Mediated mRNA Translation
3. Concluding Remarks
Acknowledgments
Conflict of Interest
References
- James, C. Brief 42: Global Status of Commercialized Biotech/GM Crops: 2010; ISAAA: Ithaca, New York, NY, USA, 2010. [Google Scholar]
- Sanahuja, G.; Banakar, R.; Twyman, R.M.; Capell, T.; Christou, P. Bacillus tuhringiensis: A century of research, development and commercial applications. Plant Biotechnol. J. 2011, 9, 283–300. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Gassmann, A.J.; Crowder, D.W.; Carrière, Y. Insect resistance to Bt crops: Evidence versus theory. Nat. Biotechnol. 2008, 26, 199–202. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; van Rensburg, J.B.J.; Carrière, Y. Field-evolved insect resistance to Bt crops: Definition, theory, and data. J. Econ. Entomol. 2009, 102, 2011–2025. [Google Scholar] [CrossRef]
- Huang, F.; Parker, R.; Leonard, B.R.; Yang, Y.; Liu, J. Frequency of resistance alleles to Bacillus thuringiensis-corn in Texas populations of the sugarcane borer, Diatraea saccharalis (F.) (Lepidoptera: Crambidae). Crop Protect. 2009, 28, 174–180. [Google Scholar] [CrossRef]
- Downes, S.; Parker, T.L.; Mahon, R.J. Frequence of alleles conferring resistance to the Bacillus thuringiensis toxins Cry1Ac and Cry2Ab in Australian populations of Helicoverpa punctigera (Lepidoptera: Noctuidae) from 2002 to 2006. J. Econ. Entomol. 2009, 102, 733–742. [Google Scholar] [CrossRef]
- Griffitts, J.S.; Aroian, R.V. Many roads to resistance: How invertebrates adapt to Bt toxins. Bioessays 2005, 27, 614–624. [Google Scholar] [CrossRef]
- Meihls, L.N.; Higdon, M.L.; Ellersieck, M.; Hibbard, B.E. Selection for resistance to mCry3A-expressing transgenic corn in western corn rootworm. J. Econ. Entomol. 2011, 104, 1045–1054. [Google Scholar] [CrossRef]
- Oswald, K.J.; French, B.W.; Nielson, C.; Bagley, M. Selection for Cry3Bb1 resistance in a genetically diverse population of nondiapausing western corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2011, 104, 1038–1044. [Google Scholar] [CrossRef]
- Kruger, M.; van Rensburg, J.B.J.; van den Berg, J. Perspective on the development of stem borer resistance to Bt maize and refuge compliance at the Vaalharts irrigation scheme in South Africa. Crop Protect. 2009, 28, 684–689. [Google Scholar] [CrossRef]
- Bagla, P. Hardy cotton-munching pests are latest blow to GM crops. Science 2010, 327. [Google Scholar] [CrossRef]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef]
- Dhura, S.; Gujan, G.T. Field-evolved resistance to Bt toxin Cry1Ac in the pink bollworm, Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechiidae), from India. Pest Manag. Sci. 2011, 67, 898–903. [Google Scholar] [CrossRef]
- Gassmann, A.J.; Petzold-Maxwell, J.L.; Keweshan, R.S.; Dunbar, M.W. Field-evolved resistance to Bt maize by western corn rootworm. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Zhang, H.; Yin, W.; Zhao, J.; Jin, L.; Yang, Y.; Wu, S.; Tabashnik, B.E.; Wu, Y. Early warning of cotton bollworm resistance associated with intensive planting of Bt cotton in China. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Bates, S.L.; Zhao, J.-Z.; Roush, R.T.; Shelton, A.M. Insect resistance management in GM crops: Past, present and future. Nat. Biotechnol. 2005, 23, 57–62. [Google Scholar] [CrossRef]
- Naranjo, S.E. Impacts of Bt transgenic cotton on integrated pest management. J. Agric. Food Chem. 2011, 59, 5842–5851. [Google Scholar] [CrossRef]
- Gray, M.E. Relevance of traditional integrated pest management (IPM) strategies for commercial corn producers in a transgenic agroecosystem: A bygone era? J. Agric. Food Chem. 2011, 59, 5852–5858. [Google Scholar] [CrossRef]
- Roush, R.T. Two-toxin strategies for management of insecticidal transgenic crops: Can pyramiding succeed where pesticide mixtures have not? Phil. Trans. R. Soc. Lond. B 1998, 353, 1777–1786. [Google Scholar] [CrossRef]
- Moar, W.J.; Anilkumar, K.J. The power of the pyramid. Science 2007, 318, 1561–1562. [Google Scholar] [CrossRef]
- Gould, F. Bt resistance management—theory meets data. Nat. Biotechnol. 2003, 21, 1450–1451. [Google Scholar] [CrossRef]
- Zhao, J.-Z.; Cao, J.; Li, Y.; Collins, H.L.; Roush, R.T.; Earle, E.D.; Shelton, A.M. Transgenic plants expressing two Bacillus thuringiensis toxins delay insect resistance evolution. Nat. Biotechnol. 2003, 21, 1493–1497. [Google Scholar] [CrossRef]
- Zhao, J.-Z.; Cao, J.; Collins, H.L.; Bates, S.L.; Roush, R.T.; Earle, E.D.; Shelton, A.M. Concurrent use of transgenic plants expressing a single and two Bacillus thuringiensis genes speeds insect adaptation to pyramided plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8426–8430. [Google Scholar]
- Alyokhin, A. Scant evidence supports EPA’s pyramided Bt corn refuge size of 5%. Nat. Biotechnol. 2011, 29, 577–578. [Google Scholar] [CrossRef]
- Ives, A.R.; Glaum, P.R.; Ziebarth, N.L.; Andow, D.A. The evolution of resistance to two-toxin pyramid transgenic crops. Ecol. Appl. 2011, 21, 503–515. [Google Scholar] [CrossRef]
- Huang, F.; Ghimire, M.N.; Leonard, B.R.; Wang, J.; Daves, C.; Levy, R.; Cook, D.; Head, G.P.; Yang, Y.; Temple, J.; et al. F2 screening for resistance to pyramided Bacillus thuringiensis maize in Louisiana and Mississippi populations of Diatraea saccharalis (Lepidoptera: Crambidae). Pest Manag. Sci. 2011, 67, 1269–1276. [Google Scholar] [CrossRef]
- Chitkowski, R.L.; Turnispeed, S.G.; Sullivan, M.J.; Bridges, W.C., Jr. Field and laboratory evaluations of transgenic cottons expressing one or two Bacillus thuringiensis var. kurstaki Berliner proteins for management of noctuid (Lepidopteran) pests. J. Econ. Entomol. 2003, 96, 755–762. [Google Scholar] [CrossRef]
- Jackson, R.E.; Bradley, J.R.; van Duyn, J.W. Performance of feral and Cry1Ac-selected Helicoverpa zea (Lepidoptera: Noctuidae) strains on transgenic cottons expressing one or two Bacillus thuringiensis ssp. kurstaki proteins under greenhouse conditions. J. Entomol. Sci. 2004, 39, 46–55. [Google Scholar]
- Tu, J.; Zhang, G.; Datta, K.; Xu, C.; He, Y.; Zhang, Q.; Khush, G.S.; Datta, S.K. Field performance of transgenic elite commercial hybrid rice expressing Bacillus thuringiensis ∂-endotoxin. Nat. Biotechnol. 2000, 18, 1101–1104. [Google Scholar] [CrossRef]
- Bohorova, N.; Frutos, R.; Royer, M.; Estanol, P.; Pacheco, M.; Rascon, Q.; McLean, S.; Hoisington, D. Novel synthetic Bacillus thuringiensis cry1B gene and the cry1B-cry1Ab translational fusion confer resistance to southwestern corn borer, sugarcane borer and fall armyworm in transgenic tropical maize. Theor. Appl. Genet. 2001, 103, 817–826. [Google Scholar] [CrossRef]
- Naimov, S.; Dukiandjiev, S.; de Maagd, R.A. A hybrid Bacillus thuringiensis delta-endotoxin gives resistance against a coleopteran and a lepidopteran pest in transgenic potato. Plant Biotechnol. J. 2003, 1, 51–57. [Google Scholar]
- Lee, M.K.; Walters, F.S.; Hart, H.; Palekar, N.; Chen, J.-S. The mode of action of the Bacillus thuringiensis vegetative insecticidal protein Vip3A differs from that of Cry1Ab ∂-endotoxin. Appl. Environ. Microbiol. 2003, 69, 4648–4657. [Google Scholar] [CrossRef]
- Jackson, R.E.; Marcus, M.A.; Gould, F.; Bradley, J.R.; van Duyn, J.W. Cross-resistance responses of CryIAc-selected Heliothis virescens (Lepidoptera: Noctuidae) to the Bacillus thuringiensis protein vip3A. J. Econ. Entomol. 2007, 100, 180–186. [Google Scholar] [CrossRef]
- Kurtz, R.W.; McCaffery, A.; O’Reilly, D. Insect resistance management for Syngenta’s VipCot™ transgenic cotton. J. Inverteb. Pathol. 2007, 95, 227–230. [Google Scholar] [CrossRef]
- Burkness, E.C.; Dively, G.; Patton, T.; Morey, A.C.; Hutchison, W.D. Novel Vip3A Bacillus thuringiensis (Bt) maize approaches high-dose efficacy against Helicoverpa zea (Lepidoptera: Noctuidae) under field conditions: Implications for resistance management. GM Crops 2010, 1, 337–343. [Google Scholar] [CrossRef]
- Maqbool, S.B.; Riazzudin, S.; Loc, N.T.; Gatehouse, A.M.R.; Gatehouse, J.A.; Christou, P. Expression of multiple insecticidal genes confers broad resistance against a range of different rice pests. Mol. Breed. 2001, 7, 85–93. [Google Scholar] [CrossRef]
- Yang, G.D.; Zhu, Z.; Li, Y.O.; Zhu, Z.J. Transformation of Bt-CpTi fusion protein gene to cabbage (Brassica oleracea var. capitata) mediated by Agrobacterium tumefaciens and particle bombardment. Shi Yan Sheng Wu Xue Bao 2002, 35, 117–122. [Google Scholar]
- Datta, K.; Baisakh, N.; Thet, K.M.; Tu, J.; Datta, S.K. Pyramiding transgenes for multiple resistance in rice against bacterial blight, yellow stem borer and sheath blight. Theor. Appl. Genet. 2002, 106, 1–8. [Google Scholar]
- Su, X.; Chu, Y.; Li, H.; Hou, Y.; Zhang, B.; Huang, Q.; Hu, Z.; Huang, R.; Tian, Y. Expression of multiple resistance genes enhances tolerance to environmental stressors in transgenic poplar (Populus x euramericana ‘Guariento’). PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, M.; Zhang, X.; Luan, H.; Diao, S.; Tian, Y.; Su, X. Laboratory and field evaluation of the transgenic Populus alba ×Populus glandulosa expressing double coleopteran-resistance genes. Tree Physiol. 2011, 31, 567–573. [Google Scholar] [CrossRef]
- Moellenbeck, D.J.; Peters, M.L.; Bing, J.W.; Rouse, J.R.; Higgins, L.S.; Sims, L.; Nevshemal, T.; Marshall, L.; Ellis, R.T.; Bystrak, P.G.; et al. Insecticidal proteins from Bacillus thuringiensis protect corn from corn rootworms. Nat. Biotechnol. 2001, 19, 668–672. [Google Scholar] [CrossRef]
- Boulter, D.; Edwards, G.A.; Gatehouse, A.M.R.; Gatehouse, J.A.; Hilder, V.A. Additive protective effects of different plant-derived insect resistance genes in transgenic tobacco plants. Crop Protect. 1990, 9, 351–354. [Google Scholar] [CrossRef]
- Tang, K.; Tinjuangjun, P.; Xu, Y.; Sun, X.; Gatehouse, J.A.; Ronald, P.C.; Qi, H.; Lu, X.; Christou, P.; Kohli, A. Particle-bombardment-mediated co-transformation of elite Chinese rice cultivars with genes conferring resistance to bacterial blight and sap-sucking insect pests. Planta 1999, 208, 552–563. [Google Scholar] [CrossRef]
- Campbell, M.A.; Fitzgerald, H.A.; Ronald, P.C. Engineering pathogen resistance in crop plants. Transg. Res. 2002, 11, 599–613. [Google Scholar] [CrossRef]
- Abdeen, A.; Virgos, A.; Olivella, E.; Villanueva, J.; Aviles, X.; Gabarra, R.; Prat, S. Multiple insect resistance in transgenic tomato plants over-expressing two families of plant proteinase inhibitors. Plant Mol. Biol. 2005, 57, 189–202. [Google Scholar]
- Senthilkumar, R.; Cheng, C.-P.; Yeh, K.-W. Genetically pyramiding protease-inhibitor genes for dual braod-spectrum resistance against insect and phytopathogens in transgenic tobacco. Plant Biotechnol. J. 2010, 8, 65–75. [Google Scholar] [CrossRef]
- Cao, J.; Zhao, J.-Z.; Tang, J.D.; Shelton, A.M.; Earle, E.D. Broccoli plants with pyramided cry1Ac and cry1C Bt genes control diamondback moths resistant to Cry1A and Cry1C proteins. Theor. Appl. Genet. 2002, 105, 258–264. [Google Scholar] [CrossRef]
- Yang, Z.; Chen, H.; Tang, W.; Hua, H.; Lin, Y. Development and characterisation of transgenic rice expressing two Bacillus thuringiensis genes. Pest Manag. Sci. 2011, 67, 414–422. [Google Scholar] [CrossRef]
- Ramesh, S.; Nagadhara, D.; Pasalu, I.C.; Padma Kumari, A.; Sarma, N.P.; Reddy, V.D.; Rao, K.V. Development of stem borer resistant transgenic parental lines involved in the production of hybrid rice. J. Biotechnol. 2004, 111, 131–141. [Google Scholar]
- Vemanna, R.S.; Chandrashekar, B.K.; Hanumantha Rao, H.M.; Sathyanarayanagupta, S.K.; Sarangi, K.S.; Nataraja, K.N.; Udayakumar, M. A modified multisite Gateway cloning strategy for consolidation of genes in plants. Mol. Biotechnol. 2012. [Google Scholar] [CrossRef]
- Halpin, C. Gene stacking in transgenic plants–the challenge for 21st century plant biotechnology. Plant Biotechnol. J. 2005, 3, 141–155. [Google Scholar] [CrossRef]
- Dafny-Yelin, M.; Tzfira, T. Delivery of multiple transgenes to plant cells. Plant Physiol. 2007, 145, 1118–1128. [Google Scholar] [CrossRef]
- Naqvi, S.; Farre, G.; Sanahuja, G.; Capell, T.; Zhu, C.; Christou, P. When more is better: Multigene engineering in plants. Trends Plant Sci. 2009, 15, 48–56. [Google Scholar]
- Saxena, P.; Hsieh, Y.C.; Alvarado, V.Y.; Sainsbury, F.; Saunders, K.; Lomonossoff, G.P.; Scholthof, H.B. Improved foreign gene expression in plants using a virus-encoded suppressor of RNA silencing modified to be developmentally harmless. Plant Biotechnol. J. 2011, 9, 703–712. [Google Scholar] [CrossRef]
- Miki, B.; Abdeen, A.; Manaba, Y.; MacDonald, P. Selectable marker genes and unintended changes to the plant transcriptome. Plant Biotechnol. J. 2009, 7, 211–218. [Google Scholar] [CrossRef]
- Uhlen, M.; Forsberg, G.; Moks, T.; Hartmanis, M.; Nilsson, B. Fusion proteins in biotechnology. Curr. Opin. Biotechnol. 1992, 3, 363–369. [Google Scholar]
- Bouchard, É.; Cloutier, C.; Michaud, D. Oryzacystatin I expressed in transgenic potato induces digestive compensation in an insect natural predator via its herbivorous prey feeding on the plant. Mol. Ecol. 2003, 12, 2439–2446. [Google Scholar] [CrossRef]
- Waldron, C.; Wegrich, L.M.; Owens Merlo, P.A.; Walsh, T.A. Characterization of a genomic sequence coding for potato multicystatin, an eight-domain cysteine proteinase inhibitor. Plant Mol. Biol. 1993, 23, 801–812. [Google Scholar] [CrossRef]
- Benchabane, M.; Schlüter, U.; Vorster, J.; Goulet, M.-C.; Michaud, D. Plant cystatins. Biochimie 2010, 92, 1657–1666. [Google Scholar] [CrossRef]
- Kiggundu, A.; Goulet, M.-C.; Goulet, C.; Dubuc, J.-F.; Rivard, D.; Benchabane, M.; Pépin, G.; van der Vyver, C.; Kunert, K.; Michaud, D. Modulating the proteinase inhibitory profile of a plant cystatin by single mutations at positively selected amino acid sites. Plant J. 2006, 48, 403–413. [Google Scholar] [CrossRef]
- Goulet, M.-C.; Dallaire, C.; Vaillancourt, L.-P.; Khalf, M.; Badri, A.M.; Preradov, A.; Duceppe, M.-O.; Goulet, C.; Cloutier, C.; Michaud, D. Tailoring the specificity of a plant cystatin toward herbivorous insect digestive cysteine proteases by single mutations at positively selected amino acid sites. Plant Physiol. 2008, 146, 1010–1019. [Google Scholar] [CrossRef]
- Walsh, T.A.; Strickland, J.A. Proteolysis of the 85-kilodalton crystalline cysteine proteinase inhibitor from potato releases functional cystatin domains. Plant Physiol. 1993, 103, 1227–1234. [Google Scholar]
- de Maagd, R.A.; Kwa, M.S.G.; van der Klei, H.; Yamamoto, T.; Schipper, B.; Vlak, J.M.; Stiekema, W.J.; Bosch, D. Domain III substitution in Bacillus thuringiensis delta-endotoxin Cry1A(b) results in superior toxicity for Spodoptera exigua and altered membrane protein recognition. Appl. Environ. Microbiol. 1996, 62, 1537–1543. [Google Scholar]
- Walters, F.S.; deFontes, C.M.; Hart, H.; Warren, G.W.; Chen, J.S. Lepidopteran-active variable-region sequence imparts Coleopteran activity in eCry3.1Ab, an engineered Bacillus thuringiensis hybrid insecticidal protein. Appl. Environ. Microbiol. 2010, 76, 3082–3088. [Google Scholar]
- de Maagd, R.A.; Weemen-Hendriks, M.; Stiekema, W.J.; Bosch, D. Bacillus thuringiensis delta-endotoxin Cry1C domain III can function as a specificity determinant for Spodoptera exigua in different, but not all, Cry1–Cry1C hybrids. Appl. Environ. Microbiol. 2000, 66, 1559–1563. [Google Scholar] [CrossRef]
- Naimov, S.; Weemen-Hendriks, M.; Dukiandjiev, S.; de Maagd, R.A. Bacillus thuringiensis delta-endotoxin Cry1 hybrid proteins with increased activity against the Colorado potato beetle. Appl. Environ. Microbiol. 2001, 67, 5328–5330. [Google Scholar] [CrossRef]
- Karlova, R.; Weemen-Hendriks, M.; Naimov, S.; Ceron, J.; Dukiandjiev, S.; de Maagd, R.A. Bacillus thuringiensis ∂-endotoxin Cry1Ac domain III enhances activity against Heliothis virescens in some, but not all Cry1–Cry1Ac hybrids. J. Invertebr. Pathol. 2005, 88, 169–172. [Google Scholar] [CrossRef]
- Singh, P.K.; Kumar, M.; Chaturvedi, C.P.; Yadav, D.; Tuli, R. Development of a hybrid ∂-endotoxin and its expression in tobacco and cotton for control of a polyphagous pest Spodoptera litura. Transg. Res. 2004, 13, 397–410. [Google Scholar] [CrossRef]
- Lopez-Pazos, S.A.; Rojas Arias, A.C.; Ospina, S.A.; Ceron, J. Activity of Bacillus thuringiensis hybrid protein against a lepidopteran and a coleopteran pest. FEMS Microbiol. Lett. 2010, 302, 93–98. [Google Scholar] [CrossRef]
- Inanaga, H.; Kobayashi, D.; Kouzuma, Y.; Aoki-Yasunaga, C.; Kimura, M. Protein engineering of novel proteinase inhibitors and their effects on the growth of Spodoptera exigua larvae. Biosci. Biotechnol. Biochem. 2001, 65, 2259–2264. [Google Scholar] [CrossRef]
- Mehlo, L.; Gahakwa, D.; Nghia, P.T.; Loc, N.T.; Capell, T.; Gatehouse, J.A.; Gatehouse, A.M.R.; Christou, P. An alternative strategy for sustainable pest resistance in genetically enhanced crops. Proc. Natl. Acad. Sci. USA 2005, 102, 7812–7816. [Google Scholar]
- Xia, L.; Long, X.; Ding, X.; Zhang, Y. Increase in insecticidal toxicity by fusion of the cry1Ac gene from Bacillus thuringiensis with the neurotoxin gene hwtx-I. Curr. Microbiol. 2009, 58, 52–57. [Google Scholar]
- Xia, L.; Zheng, Z.; Ding, X.; Huang, F. The expression of a recombinant cry1Ac gene with subtilisin-like protease CDEP2 gene in acrystalliferous Bacillus thuringiensis by Red/ET homologous recombination. Curr. Microbiol. 2009, 59, 386–392. [Google Scholar] [CrossRef]
- Seo, J.H.; Yeo, J.S.; Cha, H.J. Baculoviral polyhedrin—Bacillus thuringiensis toxin fusion protein: A protein-based bio-insecticide expressed in Escherichia coli. Biotechnol. Bioeng. 2005, 92, 166–172. [Google Scholar] [CrossRef]
- Cao, C.-W.; Liu, G.-F.; Wang, Z.-Y.; Yan, S.-C.; Ma, L.; Yang, C.-P. Response of the gypsy moth, Lymantria dispar to transgenic poplar, Populus simonii ×P. nigra, expressing fusion protein gene of the spider insecticidal peptide and Bt-toxin C-peptide. J. Insect Sci. 2010, 10. [Google Scholar] [CrossRef]
- Zheng, S.S.; An, C.C.; Li, Q.R.; Chen, Z.L. Study of transferring fusion protein gene of the spider insecticidal peptide and Bt-toxin C-peptide into cotton. Cotton Sci. 2002, 14, 348–351. [Google Scholar]
- Fan, H.J.; Hu, C.X.; Wang, Z.Y.; Liu, G.F. Resistance of transgenic Xiaohei poplars with fusion protein gene of the spider insecticidal peptide and Bt-toxin C-peptide to Clostera anachoreta (Fabricius) (Lepidoptera; Notodontidae). Acta Entomol. Sin. 2006, 49, 780–785. [Google Scholar]
- Wang, Z.Y.; Xue, Z.; Fang, H.J.; Zhan, Y.G. Resistance of transgenic Betula platyphylla to the defoliator Lymantria dispar. Sci. Silvae Sci. 2007, 43, 116–120. [Google Scholar]
- Fitches, E.; Audsley, N.; Gatehouse, J.A.; Edwards, J.P. Fusion proteins containing neuropeptides as novel insect control agents: Snowdrop lectin delivers fused allatostatin to insect haemolymph following oral ingestion. Insect Biochem. Mol. Biol. 2002, 32, 1653–1661. [Google Scholar] [CrossRef]
- Fitches, E.; Edwards, M.G.; Mee, C.; Grishin, E.; Gatehouse, A.M.R.; Edwards, J.P.; Gatehouse, J.A. Fusion proteins containing insect-specific toxins as pest control agents: Snowdrop lectin delivers fused insecticidal spider venom toxin to insect haemolymph following oral ingestion. J. Insect Physiol. 2004, 50, 61–71. [Google Scholar] [CrossRef]
- Fitches, E.C.; Bell, H.A.; Powell, M.E.; Back, E.; Sargiotti, C.; Weaver, R.J.; Gatehouse, J.A. Insecticidal activity of scorpion toxin (ButalT) and snowdrop lectin (GNA) containing fusion proteins towards pest species of different orders. Pest Manag. Sci. 2010, 66, 74–83. [Google Scholar] [CrossRef]
- Trung, N.P.; Fitches, E.; Gatehouse, J.A. A fusion protein containing a lepidopteran-specific toxin from the South Indian red scorpion (Mesobuthus tamulus) and snowdrop lectin shows oral toxicity to target insects. BMC Biotechnol. 2006, 6. [Google Scholar] [CrossRef] [Green Version]
- Down, R.E.; Fitches, E.C.; Wiles, D.P.; Corti, P.; Bell, H.A.; Gatehouse, J.A.; Edwards, J.P. Insecticidal spider venom toxin fused to snowdrop lectin is toxic to the peach-potato aphid, Myzus persicae (Hemiptera: Aphididae) and the rice brown planthopper, Nilaparvata lugens (Hemiptera: Delphacidae). Pest Manag. Sci. 2006, 62, 77–85. [Google Scholar]
- Zhu-Salzman, K.; Ahn, J.-E.; Salzman, R.A.; Koiwa, H.; Shade, H.; Balfe, S. Fusion of a soybean cysteine protease inhibitor and a legume lectin enhances anti-insect activity synergistically. Agric. For. Entomol. 2003, 5, 317–323. [Google Scholar] [CrossRef]
- Outchkourov, N.S.; de Kogel, W.J.; Wiegers, G.L.; Abrahamson, M.; Jongsma, M.A. Engineered multidomain cysteine protease inhibitors yield resistance against western flower thrips (Frankliniella occidentalis) in greenhouse trials. Plant Biotechnol. J. 2004, 2, 449–458. [Google Scholar] [CrossRef]
- Brunelle, F.; Girard, C.; Cloutier, C.; Michaud, D. A hybrid, broad-spectrum inhibitor of Colorado potato beetle aspartate and cysteine digestive proteinases. Arch. Insect Biochem. Physiol. 2005, 60, 20–31. [Google Scholar] [CrossRef]
- Benchabane, M.; Goulet, M.-C.; Dallaire, C.; Côté, P.-L.; Michaud, D. Hybrid protease inhibitors for pest and pathogen control–a functional cost for the fusion partners? Plant Physiol. Biochem. 2008, 46, 701–708. [Google Scholar] [CrossRef]
- Michaud, D. Avoiding protease-mediated resistance in herbivorous pests. Trends Biotechnol. 1997, 15, 4–6. [Google Scholar] [CrossRef]
- Tajne, S.; Sanam, R.; Gundla, R.; Gandhi, N.S.; Mancera, R.L.; Boddupally, D.; Vudem, D.R.; Khareedu, V.R. Molecular modeling of Bt Cry1Ac (DI-DII)-ASAL (Allium sativum lectin)-fusion protein and its interaction with aminopeptidase N (APN) receptor of Manduca sexta. J. Mol. Graph. Model. 2012, 33, 61–76. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Saurabh, S.; Rai, P.; Singh, R.; Chandrashekar, K.; Verma, P.C.; Singh, P.K.; Tuli, R. SUMO fusion facilitates expression and purification of garlic leaf lectin but modifies some of its properties. J. Biotechnol. 2010, 146, 1–8. [Google Scholar] [CrossRef]
- Arai, R.; Ueda, H.; Kitayama, A.; Kamiya, N.; Nagamune, T. Design of the linkers which effectively separate domains of a bifunctional fusion protein. Prot. Eng. 2001, 14, 529–532. [Google Scholar] [CrossRef]
- Wriggers, W.; Chakravarty, S.; Jennings, P.A. Control of protein functional dynamics by peptide linkers. Biopolymers 2005, 80, 736–746. [Google Scholar] [CrossRef]
- Crasto, C.J.; Feng, J. LINKER: A program to generate linker sequences for fusion proteins. Prot. Eng. 2000, 13, 309–312. [Google Scholar] [CrossRef]
- Xue, F.; Gu, Z.; Feng, J. LINKER: A web server to generate peptide sequences with extended conformation. Nucl. Acids Res. 2004, 32, W562–W565. [Google Scholar] [CrossRef]
- Kavoosi, M.; Creagh, A.L.; Kilburn, D.G.; Haynes, C.A. Strategy for selecting and characterizing linker peptides for CBM9-tagged fusion proteins expressed in Escherichia coli. Biotechnol. Bioeng. 2007, 98, 599–610. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Morton, F.R.; Kok, C.Y.; Kong, J.; Barrett, A.J. MEROPS: The peptidase database. Nucleic.. Acids Res. 2006, 36, D320–D325. [Google Scholar]
- George, R.A.; Heringa, J. An analysis of protein domain linkers: Their classification and role in protein folding. Prot. Eng. 2003, 15, 871–879. [Google Scholar]
- Arai, R.; Wriggers, W.; Nishikawa, Y.; Nagamune, T.; Fujisawa, T. Conformations of variably linked chimeric proteins evaluated by synchrotron X-ray small-angle scattering. Proteins 2004, 57, 829–838. [Google Scholar] [CrossRef]
- Peschen, D.; Li, H.-P.; Fischer, R.; Kreuzaler, F.; Liao, Y.-C. Fusion proteins comprising a Fusarium-specific antibody linked to antifungal peptides protect plants against a fungal pathogen. Nat. Biotechnol. 2004, 22, 732–738. [Google Scholar] [CrossRef]
- Dandekar, A.M.; Gouran, H.; Ibanez, A.M.; Uratsu, S.L.; Agüero, C.B.; McFarland, S.; Borhani, Y.; Feldstein, P.A.; Bruening, G.; Nascimento, R.; et al. An engineered innate immune defense protects grapevines from Pierce disease. Proc. Natl. Acad. Sci. USA 2012, 109, 3721–3725. [Google Scholar]
- Urwin, P.E.; McPherson, M.J.; Atkinson, H.J. Enhanced transgenic plant resistance to nematodes by dual proteinase inhibitor constructs. Planta 1998, 204, 472–479. [Google Scholar] [CrossRef]
- Lu, P.; Feng, M.G. Bifunctional enhancement of a beta-glucanase-xylanase fusion enzyme by optimization of peptide linkers. Appl. Microbiol. Biotechnol. 2008, 79, 579–587. [Google Scholar] [CrossRef]
- Zhao, H.L.; Yao, X.Q.; Xue, C.; Wang, Y.; Xiong, X.H.; Liu, Z.M. Increasing the homogeneity, stability and activity of human serum albumin and interferon-alpha 2b fusion protein by linker engineering. Protein Expr. Purif. 2008, 61, 73–77. [Google Scholar] [CrossRef]
- Sauer, J.; Christensen, T.; Frandsen, T.P.; Mirgorodskaya, E.; McGuire, K.A.; Driguez, H.; Roepstorff, P.; Sigurskjold, B.W.; Svensson, B. Stability and function of interdomain linker variants of glucoamylase 1 from Aspergillus niger. Biochemistry 2001, 40, 9336–9346. [Google Scholar]
- Wu, Y.J.; Fan, C.Y.; Li, Y.K. Protein purification involving a unique auto-cleavage feature of a repeated EAAAK peptide. J. Biosci. Bioeng. 2009, 108, S63–S63. [Google Scholar]
- Maia, I.G.; Seron, K.; Haenni, A.L.; Bernardi, F. Gene expression from viral RNA genomes. Plant Mol. Biol. 1996, 32, 367–391. [Google Scholar] [CrossRef]
- Dougherty, W.G.; Semler, B.L. Expression of virus-encoded proteinases: Functional and structural similarities with cellular enzymes. Microbiol. Rev. 1993, 57, 781–822. [Google Scholar]
- Adams, M.J.; Antoniw, J.F.; Beaudoin, F. Overview and analysis of the polyprotein cleavage sites in the family Potyviridae. Mol. Plant Pathol. 2005, 6, 471–487. [Google Scholar] [CrossRef]
- Marcos, J.F.; Beachy, R.N. In vitro characterization of a cassette to accumulate multiple proteins through synthesis of a self-processing polypeptide. Plant Mol. Biol. 1994, 24, 495–503. [Google Scholar] [CrossRef]
- Saunders, K.; Sainsbury, F.; Lomonossoff, G.P. Efficient generation of cowpea mosaic virus empty virus-like particles by the proteolytic processing of precursors in insect cells and plants. Virology 2009, 393, 329–337. [Google Scholar] [CrossRef]
- Carrington, J.; Dougherty, W. Processing of the tobacco etch virus 49K protease requires autoproteolysis. Virology 1987, 160, 355–362. [Google Scholar] [CrossRef]
- Goldbach, R.W.; Wellink, J. Comovirus: Molecular Biology and Replication. In The Plant Viruses; Harrison, B.D., Murrant, A.F., Eds.; Plenum Press: New York, NY, USA, 1996; Volume 5, pp. 35–76. [Google Scholar]
- Carrington, J.; Dougherty, W. A viral cleavage site cassette: Identification of amino acid sequences required for tobacco etch virus polyprotein processing. Proc. Natl. Acad. Sci. USA 1988, 85, 3391–3395. [Google Scholar] [CrossRef]
- Phan, J.; Zdanov, A.; Evdokimov, A.G.; Tropea, J.E.; Peters, H.K., III; Kapust, R.B.; Li, M.; Wlodawer, A.; Waugh, D.S. Structural basis for the substrate specificity of tobacco etch virus protease. J. Biol. Chem. 2002, 277, 50564–50572. [Google Scholar]
- von Bodman, S.B.; Domier, L.L.; Farrand, S.K. Expression of multiple eukaryotic genes from a single promoter in Nicotiana. Bio/Technology 1995, 13, 587–591. [Google Scholar]
- Bedoya, L.; Martinez, F.; Rubio, L.; Daros, J.-A. Simultaneous equimolar expression of multiple proteins in plants from a disarmed potyvirus vector. J. Biotechnol. 2010, 150, 268–275. [Google Scholar]
- Dasgupta, S.; Collins, G.B.; Hunt, A.G. Co-ordinated expression of multiple enzymes in different subcellular compartments in plants. Plant J. 1998, 16, 107–116. [Google Scholar] [CrossRef]
- Marcos, J.F.; Beachy, R.N. Transgenic accumulation of two plant virus coat proteins on a single self-processing polypeptide. J. Gen. Virol. 1997, 78, 1771–1778. [Google Scholar]
- Ceriani, M.F.; Marcos, J.F.; Hopp, H.E.; Beachy, R.N. Simultaneous accumulation of multiple viral coat proteins from a TEV-NIa based expression vector. Plant Mol. Biol. 1998, 36, 239–248. [Google Scholar] [CrossRef]
- Powell, W.A.; Catranis, C.M.; Maynard, C.A. Design of self-processing antimicrobial peptides for plant protection. Lett. Appl. Microbiol. 2000, 31, 163–168. [Google Scholar]
- Liang, H.; Gao, H.; Maynard, C.A.; Powell, W.A. Expression of a self-processing, pathogen resistance-enhancing gene construct in Arabidopsis. Biotechnol. Lett. 2005, 27, 435–442. [Google Scholar] [CrossRef]
- Zhang, B.; Rapolu, M.; Huang, L.; Su, W.W. Coordinate expression of multiple proteins in plant cells by exploiting endogenous kex2p-like protease activity. Plant Biotechnol. J. 2011, 9, 970–981. [Google Scholar] [CrossRef]
- François, I.E.J.A.; Dwyer, G.I.; De Bolle, M.F.C.; Goderis, I.J.W.M.; van Hemelrijck, W.; Proost, P.; Wouters, P.; Broekaert, W.F.; Cammue, B.P.A. Processing in transgenic Arabidopsis thaliana plants of polyproteins with linker peptide variants derived from the Impatiens balsamina antimicrobial polyprotein precursor. Plant Physiol. Biochem. 2002, 40, 871–879. [Google Scholar] [CrossRef]
- François, I.E.J.A.; de Bolle, M.F.C.; Dwyer, G.; Goderis, I.J.W.M.; Woutors, P.F.J.; Verhaert, P.D.; Proost, P.; Schaaper, W.M.M.; Cammue, B.P.A.; Broekaert, W.F. Transgenic expression in Arabidopsis of a polyprotein construct leading to production of two different antimicrobial proteins. Plant Physiol. 2002, 128, 1346–1358. [Google Scholar] [CrossRef]
- François, I.E.J.A.; van Hemelrijck, W.; Aerts, A.M.; Wouters, P.F.J.; Proost, P.; Broekaert, W.F.; Cammue, B.P.A. Processing in Arabidopsis thaliana of a heterologous polyprotein resulting in differential targeting of the individual plant defensins. Plant Sci. 2004, 166, 113–121. [Google Scholar] [CrossRef]
- Walker, J.M.; Vierstra, R.D. A ubiquitin-based vector for the co-ordinate synthesis of multiple proteins in plants. Plant Biotechnol. J. 2007, 5, 413–421. [Google Scholar] [CrossRef]
- Hunt, A.G.; Maiti, I.B. Strategies for expressing multiple foreign genes in plants as polycistronic constructs. In Vitro Cell. Dev. Biol. Plant 2001, 37, 313–320. [Google Scholar] [CrossRef]
- Luke, G.A. Translating 2A Research into Practice. In Innovations in Biotechnology; Agbo, E.C., Ed.; InTech Open: Rijeka, Croatia, 2012; pp. 165–186. [Google Scholar]
- Martinez-Salas, E. Internal ribosome entry site biology and its use in expression vectors. Curr. Opin. Biotechnol. 1999, 10, 458–464. [Google Scholar] [CrossRef]
- Toth, R.L.; Chapman, S.; Carr, F.; Santa Cruz, S. A novel strategy for the expression of foreign genes from plant virus vectors. FEBS Lett. 2001, 489, 215–219. [Google Scholar] [CrossRef]
- Dorokhov, Y.L.; Skulachev, M.V.; Ivanov, P.A.; Zvereva, S.D.; Tjulkina, L.G.; Merits, A.; Gleba, Y.Y.; Hohn, T.; Atabekov, J.G. Polypurine (A)-rich sequences promote cross-kingdom conservation of internal ribosome entry. Proc. Natl. Acad. Sci. USA 2002, 99, 5301–5306. [Google Scholar]
- Groppelli, E.; Belsham, G.J.; Roberts, L.O. Identification of minimal sequences of the Rhopalosiphum padi virus 5' untranslated region required for internal initiation of protein synthesis in mammalian, plant and insect translation systems. J. Gen. Virol. 2007, 88, 1583–1588. [Google Scholar] [CrossRef] [Green Version]
- Urwin, P.E.; Zubko, E.I.; Atkinson, H.J. The biotechnological application and limitation of IRES to deliver multiple defence genes to plant pathogens. Physiol. Mol. Plant Pathol. 2002, 61, 103–108. [Google Scholar]
- Ali, Z.; Schumacher, H.M.; Heine-Dobbernack, E.; El-Banna, A.; Hafeez, F.Y.; Jacobsen, H.J.; Kiesecker, H. Dicistronic binary vector system—A versatile tool for gene expression studies in cell cultures and plants. J. Biotechnol. 2010, 145, 9–16. [Google Scholar]
- El-Banna, A.; Hajirezaei, M.R.; Wissing, J.; Ali, Z.; Vaas, L.; Heine-Dobbernack, E.; Jacobsen, H.J.; Schumacher, H.M.; Kiesecker, H. Over-expression of PR-10a leads to increased salt and osmotic tolerance in potato cell cultures. J. Biotechnol. 2010, 150, 277–287. [Google Scholar]
- Urwin, P.E.; Yi, L.; Martin, H.; Atkinson, H.J.; Gilmartin, P.M. Functional characterization of the EMCV IRES in plants. Plant J. 2000, 24, 583–589. [Google Scholar] [CrossRef]
- Ha, S.-H.; Liang, Y.S.; Jung, H.; Ahn, M.-J.; Suh, S.-C.; Kweon, S.-J.; Kim, D.-H.; Kim, Y.-M.; Kim, J.-K. Application of two bicistronic systems involving 2A and IRES sequences to the biosynthesis of carotenoids in rice endosperm. Plant Biotechnol. J. 2010, 8, 928–938. [Google Scholar] [CrossRef]
- de Felipe, P.; Luke, G.A.; Hughes, L.E.; Gani, D.; Halpin, C.; Ryan, M.D. E unum pluribus: Multiple proteins from a self-processing polyprotein. Trends Biotechnol. 2006, 24, 68–75. [Google Scholar] [CrossRef]
- Donnelly, M.L.L.; Luke, G.; Mehrotra, A.; Li, X.; Hughes, L.E.; Gani, D.; Ryan, M.D. Analysis of the aphthovirus 2A/2B polyprotein “cleavage” mechanism indicates not a proteolytic reaction, but a novel translational effect: A putative ribosomal “skip”. J. Gen. Virol. 2001, 82, 1013–1025. [Google Scholar]
- Sharma, P.; Yan, F.; Doronina, V.A.; Escuin-Ordinas, H.; Ryan, M.D.; Brown, J.D. 2A peptides provide distinct solutions to driving stop-carry on translational recoding. Nucl. Acids Res. 2012, 40, 3143–3151. [Google Scholar] [CrossRef]
- Halpin, C.; Cooke, S.E.; Barakate, A.; El Amrani, A.; Ryan, M.D. Self-processing 2A-polyproteins—A system for co-ordinate expression of multiple proteins in transgenic plants. Plant J. 1999, 17, 453–459. [Google Scholar] [CrossRef]
- Ma, C.; Mitra, A. Expressing multiple genes in a single open reading frame with the 2A region of foot-and-mouth disease virus as a linker. Mol. Breed. 2002, 9, 191–199. [Google Scholar] [CrossRef]
- Smolenska, L.; Roberts, I.M.; Learmonth, D.; Porter, A.J.; Harris, W.J.; Michael, T.; Wilson, A.; Santa Cruz, S. Production of a functional single chain antibody attached to the surface of a plant virus. FEBS Lett. 1998, 441, 379–382. [Google Scholar] [CrossRef]
- Sainsbury, F.; Lavoie, P.-O.; D’Aoust, M.-A.; Vézina, L.-P.; Lomonossoff, G.P. Expression of multiple proteins using full-length and deleted versions of cowpea mosaic virus RNA-2. Plant Biotechnol. J. 2008, 6, 82–92. [Google Scholar]
- Marconi, G.; Albertini, E.; Barone, P.; DeMarchis, F.; Lico, C.; Marusic, C.; Rutili, D.; Veronesi, F.; Porceddu, A. In planta production of two peptides of the Classical Swine Fever Virus (CSFV) E2 glycoprotein fused to the coat protein of potato virus X. BMC Biotechnol. 2006, 6. [Google Scholar] [CrossRef]
- Ralley, L.; Enfissi, E.M.A.; Misawa, N.; Schuch, W.; Bramley, P.M.; Fraser, P.D. Metabolic engineering of ketocarotenoid formation in higher plants. Plant J. 2004, 39, 477–486. [Google Scholar] [CrossRef]
- Randall, J.; Sutton, D.; Ghoshroy, S.; Bagga, S.; Kemp, J.D. Co-ordinate expression of β- and ∂-zeins in transgenic tobacco. Plant Sci. 2004, 167, 367–372. [Google Scholar] [CrossRef]
- Kwon, S.-J.; Hwang, E.-W.; Kwon, H.-B. Genetic engineering of drought resistant potato plants by co-introduction of genes encoding trehalose-6-phosphate synthase and trehalose-6-phosphate phosphatase of Zygosaccharomyces rouxii. Kor. J. Genet. 2004, 26, 199–206. [Google Scholar]
- Geu-Flores, F.; Olsen, C.E.; Halkier, B.A. Towards engineering glucosinolates into non-cruciferous plants. Planta 2009, 229, 261–270. [Google Scholar] [CrossRef]
- Fang, J.; Qian, J.J.; Yi, S.; Harding, T.C.; Tu, G.H.; VanRoey, M.; Jooss, K. Stable antibody expression at therapeutic levels using the 2A peptide. Nat. Biotechnol. 2005, 23, 584–590. [Google Scholar] [CrossRef]
- Fang, J.; Yi, S.; Simmons, A.; Tu, G.; Nguyen, M.; Harding, T.C.; VanRoey, M.; Jooss, K. An antibody delivery system for regulated expression of therapeutic levels of monoclonal antibodies in vivo. Mol. Ther. 2007, 15, 1153–1159. [Google Scholar]
- Luke, G.; Escuin, H.; de Filipe, P.; Ryan, M. 2A to the fore-research, technology and applications. Biotechnol. Genet. Eng. Rev. 2010, 26, 223–260. [Google Scholar]
- El Amrani, A.; Barakate, A.; Askari, B.M.; Li, X.; Roberts, A.G.; Ryan, M.D.; Halpin, C. Coordinate expression and independent subcellular targeting of multiple proteins from a single transgene. Plant Physiol. 2004, 135, 16–24. [Google Scholar] [CrossRef]
- Szymczak, A.L.; Workman, C.J.; Wang, Y.; Vignali, K.M.; Dilioglou, S.; Vanin, E.F.; Vignali, D.A.A. Correction of multi-gene deficiency in vivo using a single “self-cleaving” 2A peptide-based retroviral vector. Nat. Biotechnol. 2004, 22, 589–594. [Google Scholar] [CrossRef]
- de Felipe, P.; Luke, G.A.; Brown, J.D.; Ryan, M.D. Inhibition of 2A-mediated “cleavage” of certain artificial polyproteins bearing N-terminal signal sequences. Biotechnol. J. 2010, 5, 213–223. [Google Scholar]
- Schlüter, U.; Benchabane, M.; Munger, A.; Kiggundu, A.; Vorster, J.; Goulet, M.-C.; Cloutier, C.; Michaud, D. Recombinant protease inhibitors for herbivore pest control: A multitrophic perspective. J. Exp. Bot. 2010, 61, 4169–4183. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sainsbury, F.; Benchabane, M.; Goulet, M.-C.; Michaud, D. Multimodal Protein Constructs for Herbivore Insect Control. Toxins 2012, 4, 455-475. https://doi.org/10.3390/toxins4060455
Sainsbury F, Benchabane M, Goulet M-C, Michaud D. Multimodal Protein Constructs for Herbivore Insect Control. Toxins. 2012; 4(6):455-475. https://doi.org/10.3390/toxins4060455
Chicago/Turabian StyleSainsbury, Frank, Meriem Benchabane, Marie-Claire Goulet, and Dominique Michaud. 2012. "Multimodal Protein Constructs for Herbivore Insect Control" Toxins 4, no. 6: 455-475. https://doi.org/10.3390/toxins4060455