Effects of Cyanobacterial Lipopolysaccharides from Microcystis on Glutathione-Based Detoxification Pathways in the Zebrafish (Danio rerio) Embryo



Abstract
:1. Introduction
2. Materials and Methods
2.1. Microcystis Culture and LPS Extraction
2.2. Endotoxin Activity and Microcystin Content of LPS Extractions
2.3. Breeding and Maintenance of Zebrafish
2.4. Exposure Studies and Enzyme Assays
2.5. Statistics
3. Results and Discussion
3.1. LPS from Microcystis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cyanobacterial Strain | Source | % Yield of LPS | Endotoxin Activity (×103 EU/mg) |
|---|---|---|---|
| M. aeruginosa 299 | UTCC a | 0.1 | 107.5 |
| MC 36-1 | Doctors Lake, FL | 0.4 | 0.1619 |
| MC 81-11 | Lake Ontario | 4.1 | 36.65 |
| MC 95-11 | Lake Erie | 2.4 | 6.327 |
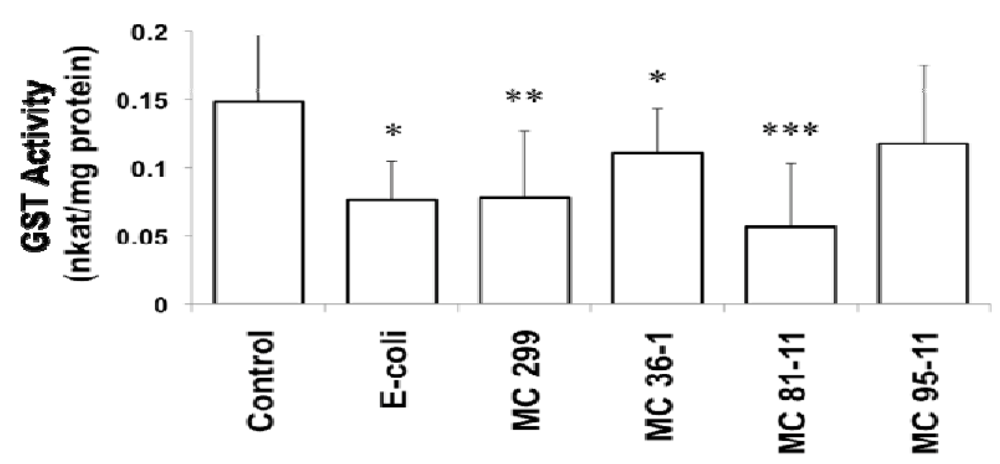
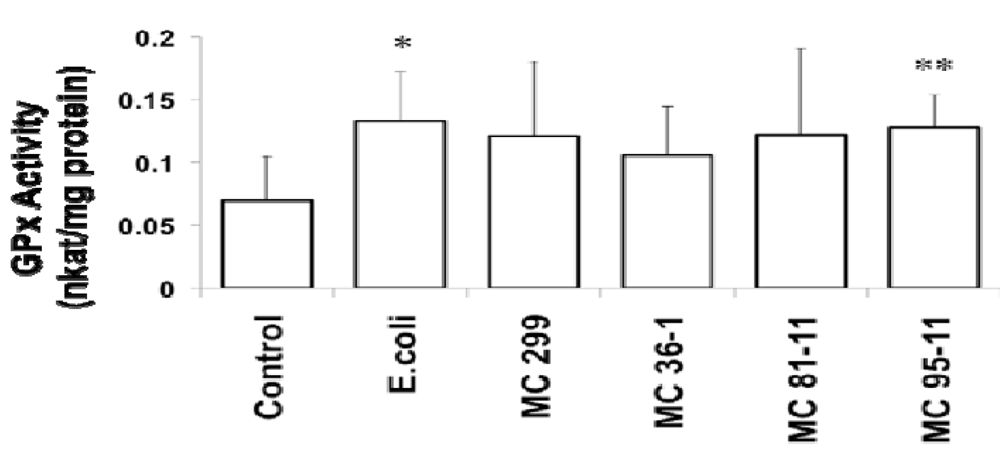
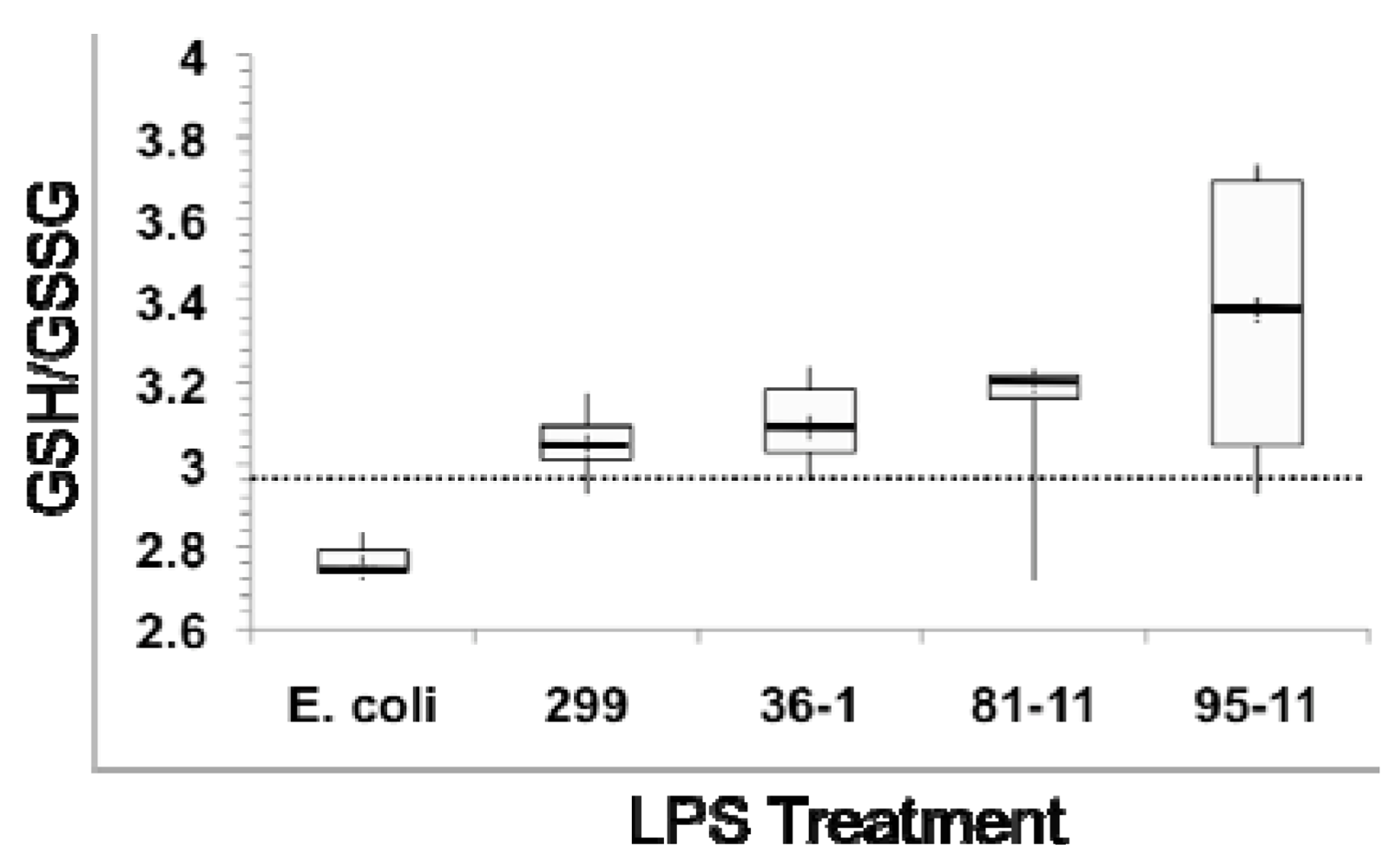
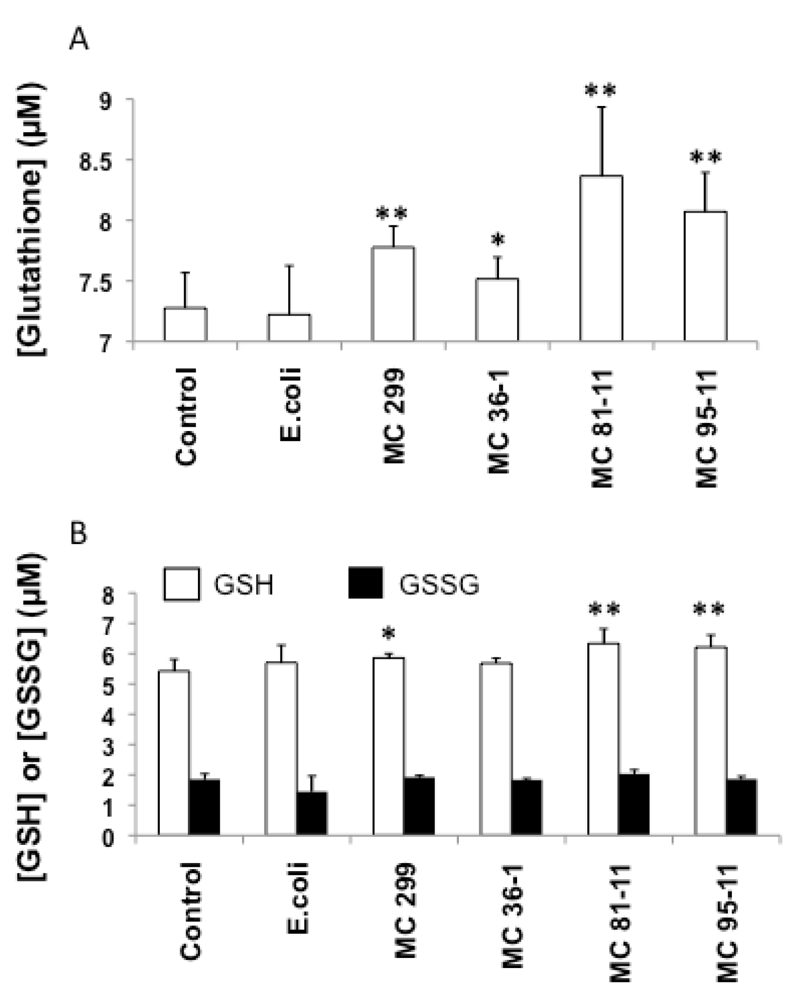
3.2. Effects of Microcystis LPS on Detoxification Pathways in the Zebrafish Embryo
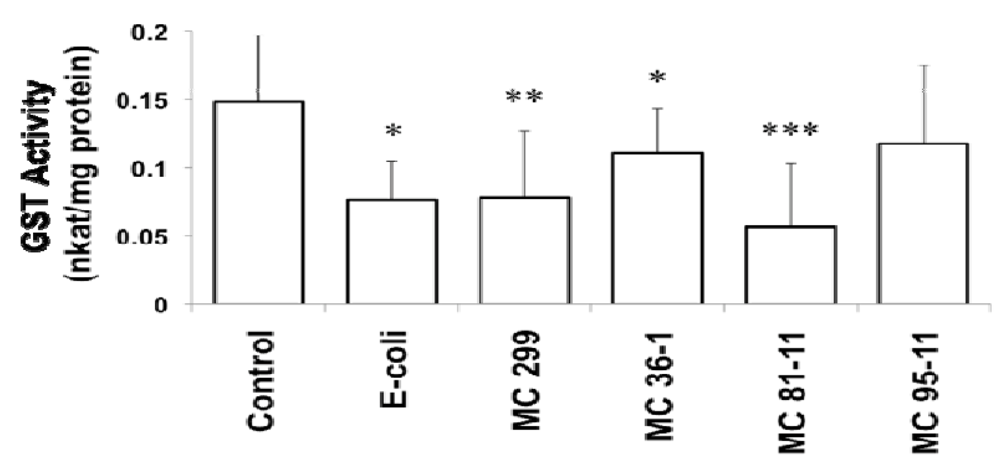
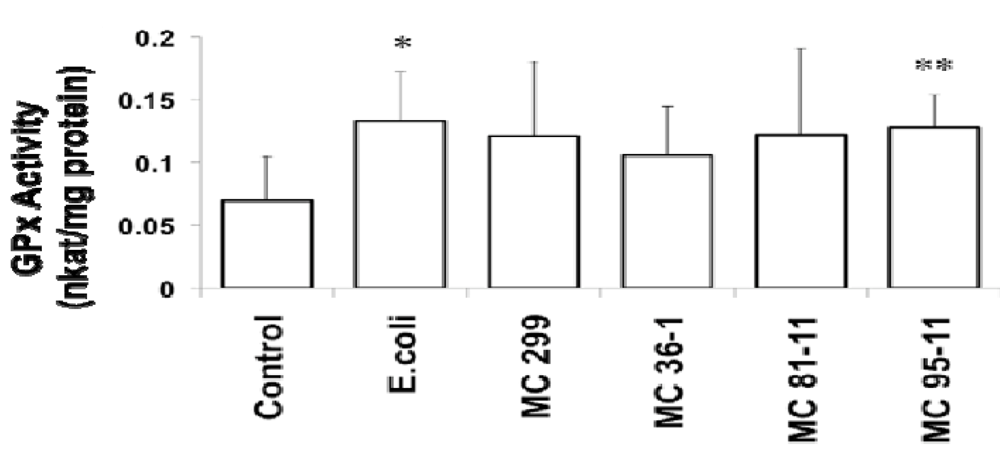


4. Conclusions
Conflict of Interest
Acknowledgements
References
- Codd, G.; Morrison, L.; Metcalf, J. Cyanobacterial toxins: Risk management for health protection. Toxicol. Appl. Pharmacol. 2005, 203, 264–272. [Google Scholar] [CrossRef]
- Brooks, G.F.; Carroll, K.C.; Butel, J.S.; Morse, S.A.; Mietzner, T.A. Jawetz, Melnick & Adelberg’s Medical Microbiology, 24th ed; McGraw-Hill Medical: New York, NY, USA, 2007. [Google Scholar]
- Saluk-Jusczak, J.; Wachowicz, B. The proinflammatory activity of lipopolysaccharide. Postepy. Biochem. 2005, 51, 280–287. [Google Scholar]
- Rada, B.; Leto, T.L. Oxidative innate immune defenses by Nox/Duox family NADPH oxidases. Contrib. Microbiol. 2008, 15, 164–187. [Google Scholar] [CrossRef]
- Poltorak, A.; He, X.; Smirnova, I.; Liu, M.Y.; Van Huffel, C.; Du, X.; Birdwell, D.; Alejos, E.; Silva, M.; Galanos, C. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: Mutations in Tlr4 gene. Science 1998, 282, 2085–2088. [Google Scholar]
- Yamamoto, M.; Akira, S. Lipid A Receptor TLR4-Mediated Signaling Pathways. In Lipid A in Cancer Therapy, 1st; Jeannin, J.F., Ed.; Springer Verlag: Berlin, Germany, 2009; pp. 59–68. [Google Scholar]
- Stewart, I.; Schluter, P.; Shaw, G. Cyanobacterial lipopolysaccharides and human health—A review. Environ. Health 2006, 5, 7. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Clifford, J.A.; Aldulescu, M.; Frenkel, J.A.; Holland, M.A.; Hall, M.L.; Glaser, K.B.; Berry, J. Cyanobacterial Microcystis aeruginosa lipopolysaccharide elicits release of superoxide anion, thromboxane B2, cytokines, chemokines and matrix metalloproteinase-9 by rat microglia. Toxicol. Sci. 2011, 121, 63–72. [Google Scholar] [CrossRef]
- Linney, E.; Upchurch, L.; Donerly, S. Zebrafish as a Neurotoxicological Model. Neurotoxicol. Teratol. 2004, 26, 709–718. [Google Scholar] [CrossRef]
- Berry, J.; Gantar, M.; Gibbs, P.; Schmale, M. The zebrafish (Danio rerio) embryo as a model system for identification and characterization of developmental toxins from marine and freshwater microalgae. Comp. Biochem. Physiol. 2007, 145, 61–72. [Google Scholar]
- Berry, J.; Gibbs, P.; Schmale, M.; Saker, M. Toxicity of cylindrospermopsin, and other apparent metabolites from Cylindrospermopsis raciborskii and Aphanizomenon ovalisporum, to the zebrafish (Danio rerio) embryo. Toxicon 2009, 53, 289–299. [Google Scholar] [CrossRef]
- Dodd, A.; Curtis, P.; Williams, L.; Love, D. Zebrafish: Bridging the gap between development and disease. Hum. Molec. Genet. 2000, 9, 2443–2449. [Google Scholar] [CrossRef]
- Zhu, H.; Zon, L.I. Use of zebrafish models for the analysis of human disease. Curr. Protoc. Hum. Genet. 2002, 15, 1–9. [Google Scholar]
- Rocke, J.; Lees, J.; Packham, I.; Chico, T. The zebrafish as a novel tool for cardiovascular drug discovery. Recent Pat. Cardiovasc. Drug Discov. 2009, 4, 1–5. [Google Scholar]
- Best, J.H.; Pflugmacher, S.; Weigand, C.; Eddy, F.B.; Metcalf, J.S.; Codd, G.A. Effects of enteric bacterial and cyanobacterial lipopolysaccharides, and of microcystin-LR, on glutathione S-transferase activities in zebrafish (Danio rerio). Aquat. Toxicol. 2002, 60, 223–231. [Google Scholar] [CrossRef]
- Notch, E.G.; Miniutti, D.M.; Patenaude, C.A.; Berry, J.P.; Mayer, G.D. Cyanobacterial LPS potentiates cadmium toxicity in zebrafish (Danio rerio) embryos. Environ. Toxicol. 2011, 26, 498–505. [Google Scholar] [CrossRef]
- Berczi, I.; Bertok, L.; Berznai, T. Comparative studies on the toxicity of Escherichia coli lipopolysaccharide endotoxin in various animal species. Can. J. Microbiol. 1966, 12, 1070–1071. [Google Scholar] [CrossRef]
- Novoa, B.; Bowman, T.V.; Zon, L.; Figueras, A. LPS response and tolerance in the zebrafish. Fish Shellfish Immunol. 2009, 26, 326–331. [Google Scholar] [CrossRef] [Green Version]
- Sepulcre, M.P.; Alcaraz-Perez, F.; Lopez-Munoz, A.; Roca, F.J.; Meseguer, J.; Cayuela, M.L.; Mulero, V. Evolution of lipopolysaccharide (LPS) recognition and signaling: Fish TLR4 does not recognize LPS and negatively regulates NF-kappaB activation. J. Immunol. 2009, 182, 1836–1845. [Google Scholar]
- Komarek, J.; Anagnostidis, K. Modern approach to the classification system of cyanophytes 2: Chroococcales. Arch. Hydrobiol. Algol. Stud. 1986, 43, 157–226. [Google Scholar]
- Komarek, J.; Anagnostidis, K. Modern approach to the classification system of cyanophytes 2: Nostocales. Arch. Hydrobiol. Algol. Stud. 1989, 56, 247–345. [Google Scholar]
- Gantar, M.; Berry, J.; Thomas, S.; Wang, M.; Perez, R.; Rein, K. Cyanobacterial toxins as allelochemicals with potential applications as algaecides, herbicides and insecticides. FEMS Microbiol. Ecol. 2008, 64, 55–64. [Google Scholar] [CrossRef]
- Bernadova, K.; Babica, P.; Marsalek, B.; Blaha, L. Isolation and endotoxin activities of lipopolysaccharides from cyanobacterial cultures and complex water blooms and comparison with the effects of heterotrophic bacteria and green alga. J. App. Toxicol. 2008, 28, 72–77. [Google Scholar] [CrossRef]
- Brand, M.; Granato, M.; Nusslein-Volhard, C. Zebrafish; Nusslein-Volhard, C., Dahm, R., Eds.; Oxford University Press: Tubingen, Germany, 2002; pp. 7–37. [Google Scholar]
- Cazenave, J.; Bistoni, M.; Zwirnmann, E.; Wunderlin, D.; Wiegand, C. Attenuating effects on natural organic matter on microcystin toxicity in zebrafish (Danio rerio) embryos. Envir. Toxicol. 2006, 21, 22–32. [Google Scholar] [CrossRef]
- Wiegand, C.; Pflugmacher, S.; Oberemm, A.; Meems, N.; Beattie, K.; Steinberg, C.; Codd, G. Uptake and effects of microcystin-LR on detoxication enzymes of early life stages of the zebrafish (Danio rerio). Environ. Toxicol. 1999, 14, 89–95. [Google Scholar] [CrossRef]
- Bradford, M.M. Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Moran, A.P.; Helander, I.M.; Kosunen, T.U. Compositional analysis of Helicobacter pylori rough-form lipopolysaccharides. J. Bacteriol. 1992, 174, 1370–1377. [Google Scholar]
- Lindsay, J.; Metcalf, J.S.; Codd, G.A. Comparison of four methods for the extraction of lipopolysaccharide from cyanobacteria. Toxicol. Environ. Chem. 2009, 91, 1253–1262. [Google Scholar] [CrossRef]
- Keleti, G.; Sykora, J. Production and properties of cyanobacterial endotoxins. Appl. Environ. Microbiol. 1982, 43, 104–109. [Google Scholar]
- Papageorgiou, J.; Linke, T.A.; Kapralos, C.; Nicholson, B.C.; Steffensen, D.A. Extraction of cyanobacterial endotoxin. Environ. Toxicol. 2004, 19, 82–87. [Google Scholar] [CrossRef]
- Raziuddin, S.; Siegelman, H.; Tornabene, T. Lipopolysaccharides of the cyanobacterium Microcystis aeruginosa. Eur. J. Biochem. 1983, 137, 333–336. [Google Scholar] [CrossRef]
- Snyder, D.S.; Brahamsha, B.; Azadi, P.; Palenik, B. Structure of compositionally simple lipopolysaccharide from marine Synechococcus. J. Bacteriol. 2009, 191, 5499–5509. [Google Scholar]
- Chen, J.; Song, L.; Dai, J.; Gan, N.; Liu, Z. Effects of microcystins on the growth and activity of superoxide dismutase and peroxidase of rape (Brassica napus) and rice (Oryza sativa). Toxicon 2004, 43, 393–400. [Google Scholar] [CrossRef]
- Gehringer, M.M.; Shephard, E.G.; Downing, T.G.; Wiegand, C.; Neilan, B.A. An investigation into the detoxification of microcystin-LR by the glutathione pathway in Balb/c mice. Int. J. Biochem. Cell Biol. 2004, 36, 931–941. [Google Scholar]
- Cazenave, J.; Bisotoni, M.; Pesce, S.F.; Wunderlin, A. Differential detoxification and antioxidant response in diverse organs of Corydoras paleatus experimentally exposed to microcystin-RR. Aquat. Toxicol. 2006, 76, 1–12. [Google Scholar] [CrossRef]
- Wang, L.; Liang, X.-F.; Liao, W.-Q.; Lei, L.-M.; Han, B.-P. Structural and functional characterization of microcystins detoxification-related liver genes in a phytoplanktivorous fish, nile tilapia (Oreochromis niloticus). Comp. Biochem. Physicol. Part C Toxicol. Pharmacol. 2006, 144, 216–227. [Google Scholar]
- Mayer, G.; Berry, J.; Patenaude, C.; Walsh, P. Effect of lipopolysaccharides from Microcystis and Lyngbya on metal toxicity in Fundulus heteroclitus. Bull. MDIBL 2004, 43, 143–144. [Google Scholar]
- Aderem, A.; Ulevitch, R.J. Toll-Like receptors in the induction of the innate immune response. Nature 2000, 406, 782–787. [Google Scholar]
- Müller, J.M.; Ziegler-Heitbrock, H.W.; Baeuerle, P.A. Nuclear factor kappa B, a mediator of lipopolysaccharide effects. Immunobiology 1993, 187, 233–256. [Google Scholar] [CrossRef]
- Owen, J.B.; Butterfield, D.A. Measurement of oxidized/reduced glutathione ratio. Meth. Mol. Biol. 2010, 648, 269–277. [Google Scholar] [CrossRef]
- Carvan, M.J.; Sonntag, D.M.; Cmar, C.B.; Cook, R.S.; Curran, M.A.; Miller, G.L. Oxidative stress in zebrafish cells: potential utility of transgenic zebrafish as deployable sentinel for site hazard ranking. Sci. Total Environ. 2001, 274, 183–196. [Google Scholar] [CrossRef]
- Kondo, F.; Matsumoto, H.; Yamada, S.; Ishikawa, N.; Ito, E.; Nagata, S.; Ueno, Y.; Suzuki, M.; Harada, K. Detection and identification of metabolites of microcystins formed in vivo in mouse and rat livers. Chem. Res. Toxicol. 1996, 9, 1355–1359. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Wiegand, C.; Oberemm, A.; Keattie, K.A.; Krause, E.; Codd, G.A.; Steinberg, C.E.W. Identification of an enzymatically formed glutathione conjugate of the cyanobacterial hepatotoxin microcystin LR; the first step in detoxification. Biochim. Biophys. Acta 1998, 1425, 527–533. [Google Scholar] [CrossRef]
- Takenaka, S. Covalent glutathione conjugation to cyanobacterial hepatotoxin microcystin LR by F344 rat cytosolic and microsomal glutathione-S-transferases. Environ. Toxicol. Pharmacol. 2001, 9, 135–139. [Google Scholar] [CrossRef]
- Best, J.H.; Eddy, F.B.; Codd, G.A. Effects of microcystis cells, cell extracts and lipopolysaccharide on drinking and liver function in rainbow trout oncorhynchus mykiss walbaum. Aquat. Toxicol. 2003, 64, 419–426. [Google Scholar] [CrossRef]
- Lindsay, J.; Metcalf, J.S.; Codd, G.A. Protection against the toxicity of microcystin-LR and cylindrospermopsin in Artemia salina and Daphnia spp. by pre-treatment with cyanobacterial lipopolysaccharide (LPS). Toxicon 2006, 48, 995–1001. [Google Scholar]
- Dionisio-Pires, L.M.; Sarpe, D.; Brehm, M.; Ibelings, B.W. Potential synergistic effects of microcystins and bacterial lipopolysaccharides on life history traits of Daphnia galeata raised on low and high food levels. Aquat. Toxicol. 2011, 104, 230–242. [Google Scholar] [CrossRef]
- Runnegar, M.T.; Kong, S.M.; Zhong, Y.Z.; Lu, S.C. Inhibition of reduced glutathione synthesis by cyanobacterial alkaloid cylindrospermopsin in cultured rat hepatocytes. Biochem. Pharmacol. 1995, 49, 219–225. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jaja-Chimedza, A.; Gantar, M.; Mayer, G.D.; Gibbs, P.D.L.; Berry, J.P. Effects of Cyanobacterial Lipopolysaccharides from Microcystis on Glutathione-Based Detoxification Pathways in the Zebrafish (Danio rerio) Embryo. Toxins 2012, 4, 390-404. https://doi.org/10.3390/toxins4060390
Jaja-Chimedza A, Gantar M, Mayer GD, Gibbs PDL, Berry JP. Effects of Cyanobacterial Lipopolysaccharides from Microcystis on Glutathione-Based Detoxification Pathways in the Zebrafish (Danio rerio) Embryo. Toxins. 2012; 4(6):390-404. https://doi.org/10.3390/toxins4060390
Chicago/Turabian StyleJaja-Chimedza, Asha, Miroslav Gantar, Gregory D. Mayer, Patrick D. L. Gibbs, and John P. Berry. 2012. "Effects of Cyanobacterial Lipopolysaccharides from Microcystis on Glutathione-Based Detoxification Pathways in the Zebrafish (Danio rerio) Embryo" Toxins 4, no. 6: 390-404. https://doi.org/10.3390/toxins4060390