Spider-Venom Peptides as Bioinsecticides


 ,
,  and
and
Abstract
:1. The Global Insect Pest Problem
1.1. Agricultural Pests
1.2. Vectors of Disease
2. Agrochemical Insecticides: Current Challenges to Insect Pest Control
2.1. Health Consequences and Environmental Impacts
2.2. Insecticide Resistance
3. Bioinsecticides as Natural Insect Pest Control Agents
4. Spider Venoms: Sources of Novel Bioinsecticides
5. Spider-Venom Peptide Nomenclature
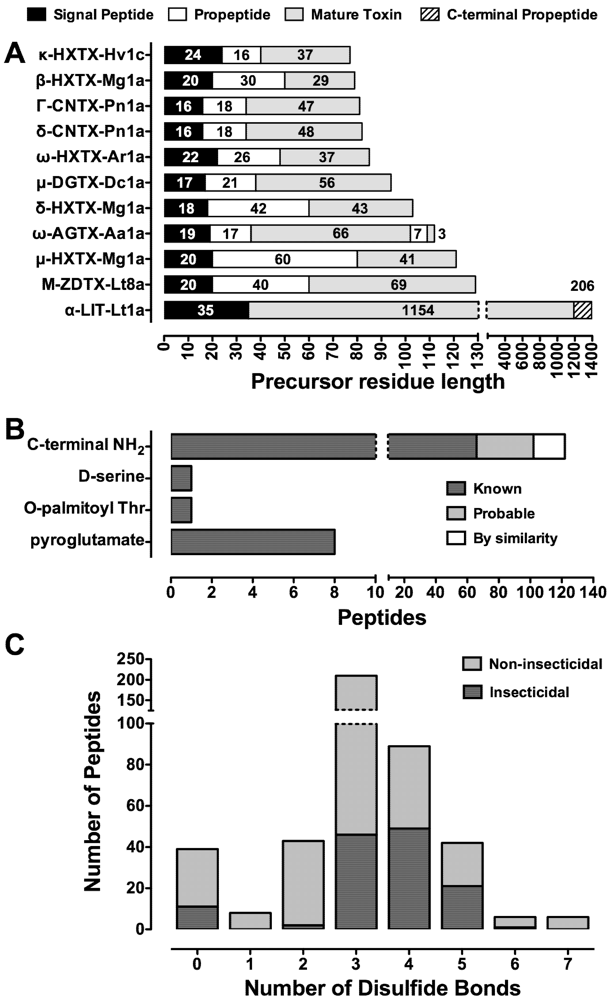
6. Structure of the Precursor Spider-Venom Peptide and Post-Translational Processing
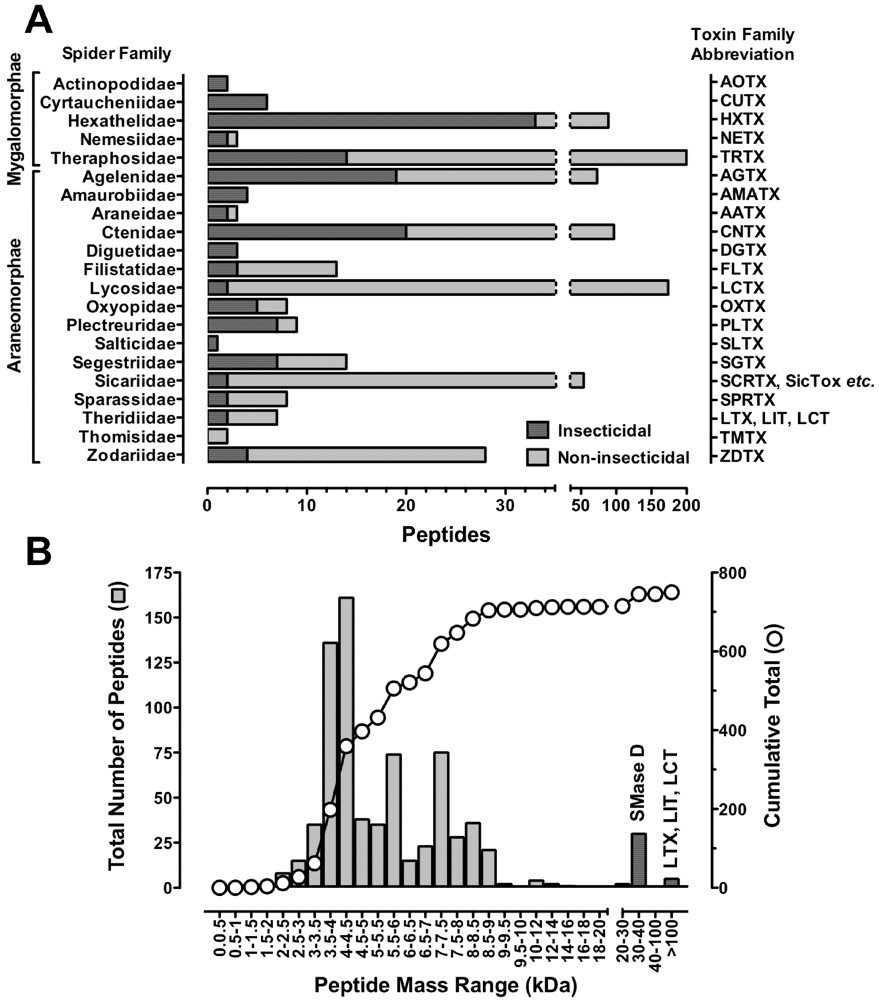
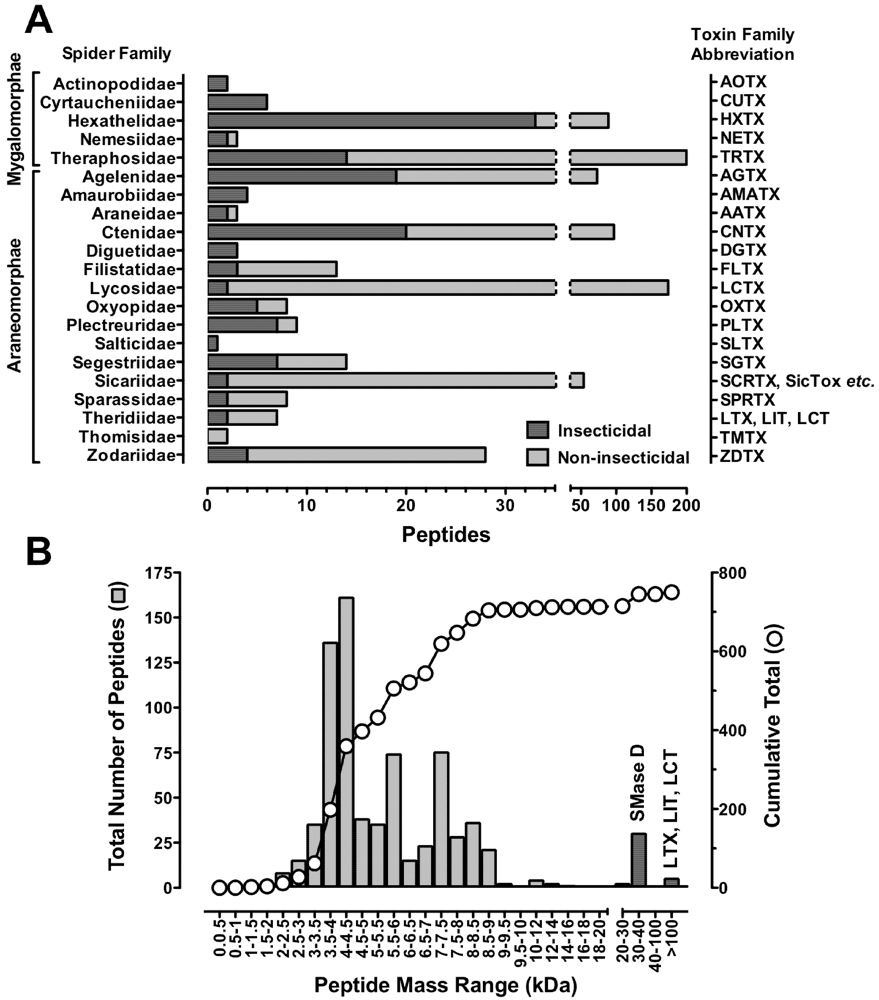
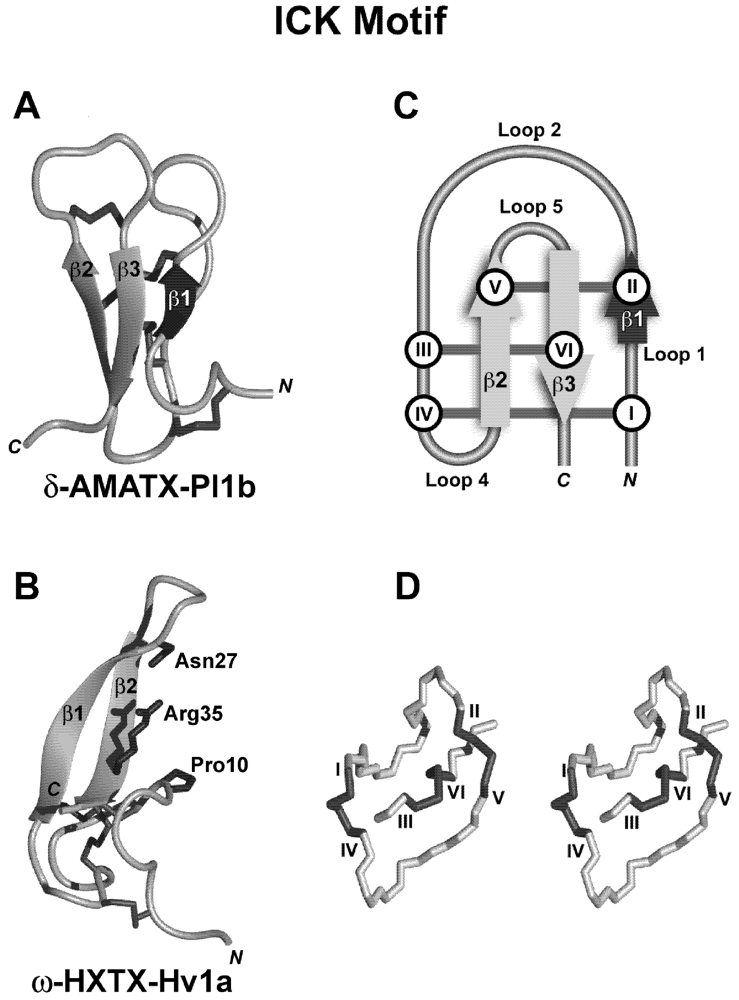
7. Structural Motifs of Spider-Venom Peptides: Variations on an Ancestral Fold
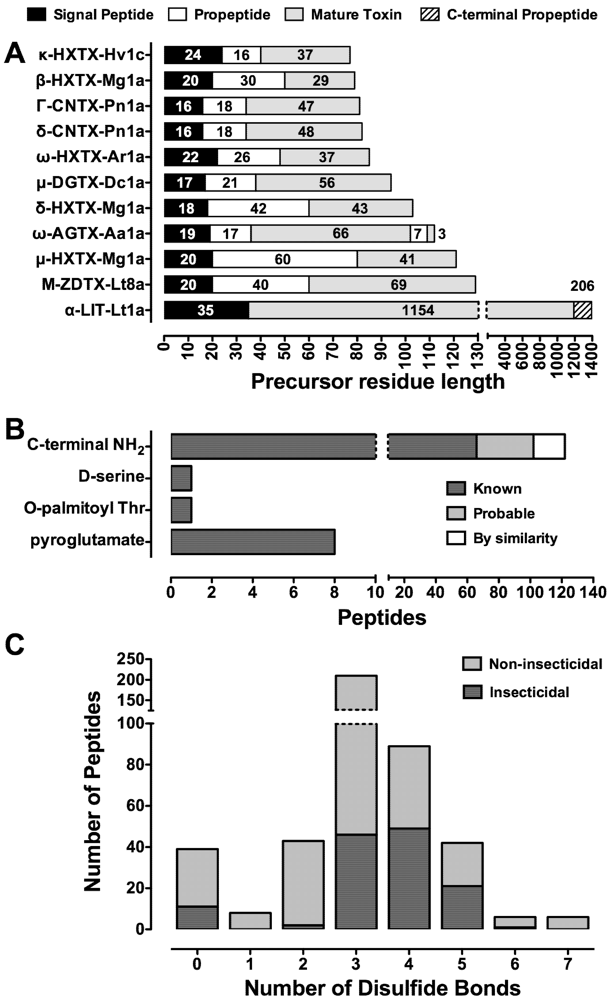
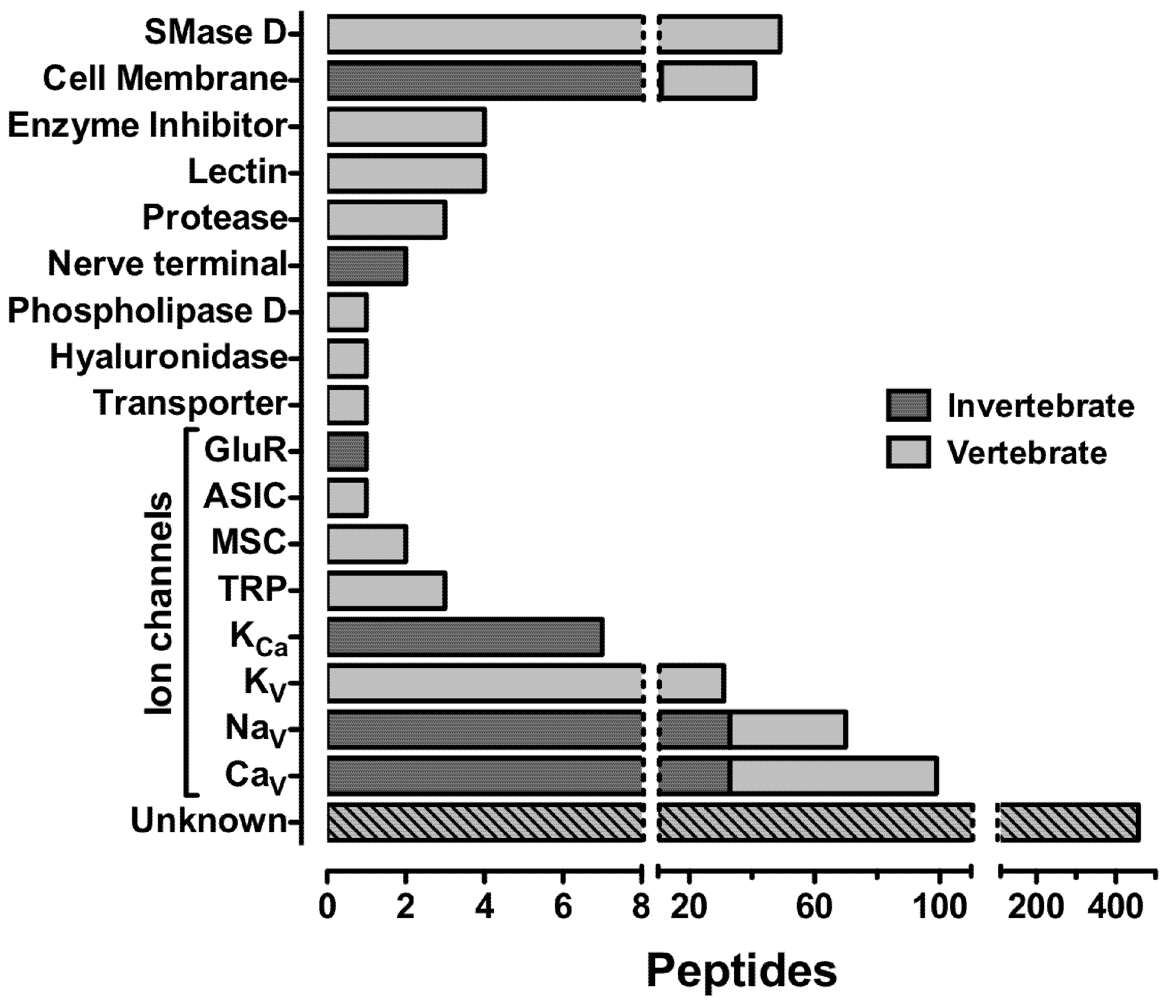
8. Insecticidal Targets of Spider Neurotoxins
8.1. Spider-Venom Peptides Targeting Insect NaV Channels
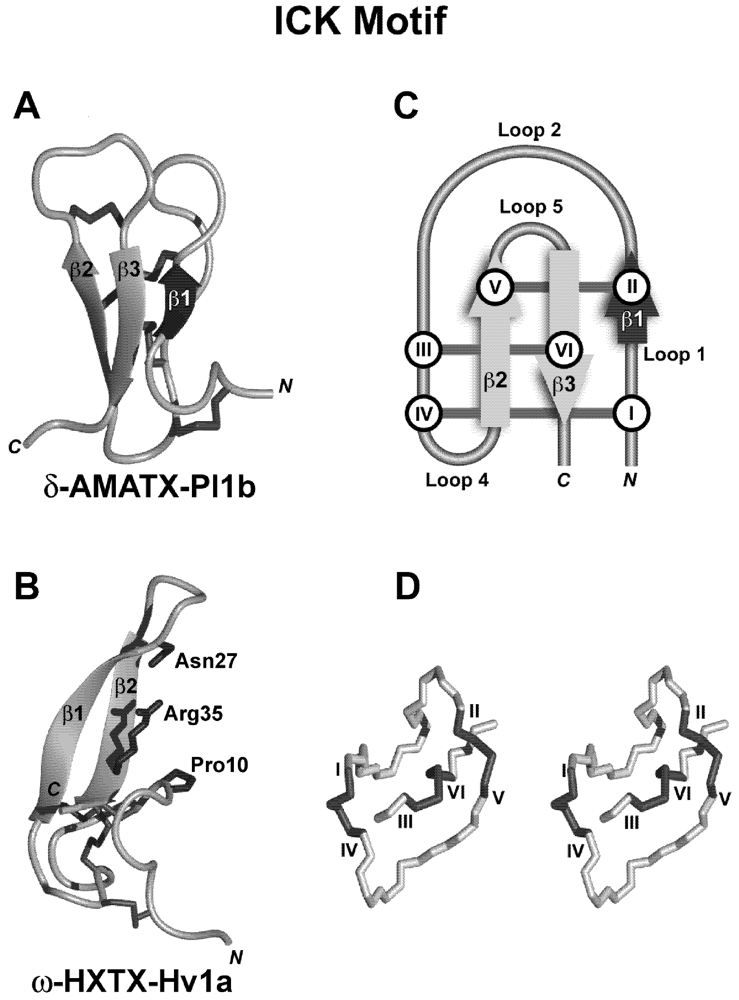
8.1.1. Spider-Venom Peptides Targeting Insect NaV Channel Site-1: Pore Blockers

8.1.2. Spider-Venom Peptides Targeting Insect NaV Channel Site-3: Gating Modifiers of Inactivation
8.1.3. Spider-Venom Peptides Targeting Insect NaV Channel Site-4: Gating Modifiers of Activation
8.1.4. Spider-Venom Toxins with an Unknown Site of Action on Insect NaV Channels
8.2. Spider-Venom Peptides Targeting Insect CaV Channels
8.2.1. Spider-Venom Peptides that Block Insect CaV1 Channels
8.2.2. Spider-Venom Peptides that Block Insect CaV2 Channels
8.2.3. Spider-Venom Peptides that Block Insect CaV3 Channels
8.3. Spider-Venom Peptides Targeting Insect KV Channels
8.4. Membrane-Acting Linear Peptides
8.5. Spider-Venom Toxins Targeting Presynaptic Nerve Terminals
8.6. Spider-Venom Toxins Targeting Glutamate Receptors
9. Bioinsecticide Lead Selection
10. Commercialisation of Spider-Venom Toxins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin Name | Source | Insect Target | Acute toxicity test species (Order †: Genus species) | ED50 or PD50 (pmol/g) | LD50 (pmol/g) | Paralogs/orthologs | |
|---|---|---|---|---|---|---|---|
| δ-CNTX-Pn1a | Phoneutria nigriventer | NaV channel | B: Periplaneta americana | 95 ‡ | 2 | ||
| D: Musca domestica | 36 | ||||||
| Γ-CNTX-Pn1a | Phoneutria nigriventer | GluR | B: Periplaneta americana | 48 ‡ | 0 | ||
| D: Musca domestica | 10 ‡ | ||||||
| O: Acheta domesticus | 29 ‡ | ||||||
| κ-HXTX-Hv1c | Hadronyche versuta | BKCa channel | D: Musca domestica | 91 | 6 | ||
| D: Musca domestica | 319 # | ||||||
| D: Lucilia cuprina | 117 # | ||||||
| L: Heliothis virescens | 3195 # | ||||||
| L: Spodoptera frugiperda | 3070 # | ||||||
| O: Acheta domesticus | 167 | ||||||
| O: Acheta domesticus | 1022 # | ||||||
| μ-AGTX-Aa1d | Agelenopsis aperta | NaV channel | D: Musca domestica | 30 | 11 | ||
| L: Manduca sexta | 9524 | ||||||
| μ-DGTX-Dc1a | Diguetia canities | NaV channel | L: Heliothis virescens | 380 | 3 | ||
| ω-HXTX-Hv1a | Hadronyche versuta | CaV channel | D: Musca domestica | 250 # | 77 | 27 | |
| L: Heliothis virescens | |||||||
| 89 | |||||||
| O: Acheta domesticus |
11. Concluding Remarks
| Goals | OPs † | Carbamates | Pyrethroids | Insect-selective spider toxins |
|---|---|---|---|---|
| Broad pest-species specificity | +++ | +++ | +++ | +++ |
| Low toxicity in non-target organisms | + | + | ++ | +++ |
| Remain in the environment long enough to be effective | +++ | +++ | ++ | ++ |
| Do not persist in environment to induce resistance development | + | + | ++ | +++ |
| Cheap to produce | +++ | +++ | ++ | ++ |
| Easy to formulate and deliver | +++ | +++ | ++ | + |
| Publicly perceived as innocuous | + | + | +++ | + |
| Accessible to small farmers and agribusinesses | + | ++ | +++ | + |
Acknowledgments
References
- Ødegaard, F. How many species of arthropods? Erwin’s estimate revised. Biol. J. Linn. Soc. 2000, 71, 583–597. [Google Scholar] [CrossRef]
- Pimentel, D. Pesticides and pest control. In Integrated Pest Management: Innovation-Development Process; Peshin, R., Dhawan, A.K., Eds.; Springer Verlag: Dordrecht, The Netherland, 2009; Volume 1, pp. 83–87. [Google Scholar]
- Oerke, E.; Dehne, H. Safeguarding production-losses in major crops and the role of crop protection. Crop Prot. 2004, 23, 275–285. [Google Scholar]
- Carlini, C.R.; Grossi-de-Sá, M.F. Plant toxic proteins with insecticidal properties. A review on their potentialities as bioinsecticides. Toxicon 2002, 40, 1515–1539. [Google Scholar] [CrossRef]
- Novotny, V.; Basset, Y.; Miller, S.E.; Weiblen, G.D.; Bremer, B.; Cizek, L.; Drozd, P. Low host specificity of herbivorous insects in a tropical forest. Nature 2002, 416, 841–844. [Google Scholar]
- McCaffery, A.R. Resistance to insecticides in heliothine Lepidoptera: a global view. Philos. Trans. R. Soc. Lond. Ser. B 1998, 353, 1735–1750. [Google Scholar]
- Brooks, E.; Hines, E. Viral biopesticides for heliothine control-fact or fiction. Todays Life Sci. 1999, Jan/Feb, 38–44. [Google Scholar]
- Nicholson, G.M. Fighting the global pest problem: Preface to the special Toxicon issue on insecticidal toxins and their potential for insect pest control. Toxicon 2007, 49, 413–422. [Google Scholar]
- Nauen, R. Insecticide resistance in disease vectors of public health importance. Pest. Manag. Sci. 2007, 63, 628–633. [Google Scholar]
- Gratz, N.G. Emerging and resurging vector-borne diseases. Annu. Rev. Entomol. 1999, 44, 51–75. [Google Scholar]
- Gubler, D.J. The global emergence/resurgence of arboviral diseases as public health problems. Arch. Med. Res. 2002, 33, 330–342. [Google Scholar]
- Hall, R.D.; Gerhardt, R.R. Flies (Diptera). In Medical and Veterinary Entomology, 2nd; Mullen, G.R., Durden, L.A., Eds.; Elsevier: Burlington, NJ, USA, 2009; pp. 127–161. [Google Scholar]
- Lounibos, L.P. Invasions by insect vectors of human disease. Annu. Rev. Entomol. 2002, 47, 233–266. [Google Scholar]
- Gayle, A.; Ringdahl, E. Tick-borne diseases. Am. Fam. Physician 2001, 64, 461–466. [Google Scholar]
- Schofield, C.; Kabayo, J.P. Trypanosomiasis vector control in Africa and Latin America. Parasites Vectors 2008, 1, 1–7. [Google Scholar]
- Billeter, S.A.; Levy, M.G.; Chomel, B.B.; Breitschwerdt, E.B. Vector transmission of Bartonella species with emphasis on the potential for tick transmission. Med. Vet. Entomol. 2008, 22, 1–15. [Google Scholar]
- Brogdon, W.G.; McAllister, J.C. Insecticide resistance and vector control. Emerg. Infect. Dis. 1998, 4, 605–613. [Google Scholar]
- Centers for Disease Control and Prevention Malaria Facts: Malaria Worldwide. Available online: http://www.cdc.gov/malaria/about/facts.html (accessed on 20 January 2012).
- World Health Organization. World Malaria Report 2010. WHO Global Malaria Programme 2010, 1–62.
- World Health Organization Media centre: Malaria. Available online: http://www.who.int/mediacentre/factsheets/fs094/en/ (accessed on 20 January 2012).
- Committee on Foreign Animal Diseases of the United States Animal Health Association Foreign animal diseases, The Gray Book; Pat Campbell & Associates and Carter Printing Company: Richmond, VA, USA, 1998.
- Attaran, A.; Roberts, D.R.; Curtis, C.F.; Kilama, W.L. Balancing risks on the backs of the poor. Nat. Med. 2000, 6, 729–731. [Google Scholar]
- Casida, J.E.; Quistad, G.B. Golden age of insecticide research: past, present, or future? Annu. Rev. Entomol. 1998, 43, 1–16. [Google Scholar] [CrossRef]
- Gunnell, D.; Eddleston, M.; Phillips, M.R.; Konradsen, F. The global distribution of fatal pesticide self-poisoning: systematic review. BMC Public Health 2007, 7, 357. [Google Scholar]
- Gunnell, D.; Eddleston, M. Suicide by intentional ingestion of pesticides: a continuing tragedy in developing countries. Int. J. Epidemiol. 2003, 32, 902–909. [Google Scholar]
- Bertolote, J.M.; Fleischmann, A.; Eddleston, M.; Gunnell, D. Deaths from pesticide poisoning: a global response. Brit. J. Psychiatry 2006, 189, 201–203. [Google Scholar]
- Pimentel, D. Environmental and economic costs of the application of pesticides primarily in the United States. Environ. Dev. Sustain. 2005, 7, 229–252. [Google Scholar]
- Eriksson, P. Developmental neurotoxicity of environmental agents in the neonate. Neurotox. 1997, 18, 719–726. [Google Scholar]
- Alavanja, M.C.; Hoppin, J.A.; Kamel, F. Health effects of chronic pesticide exposure: cancer and neurotoxicity. Annu. Rev. Public Health 2004, 25, 155–197. [Google Scholar]
- Barone, S., Jr.; Das, K.P.; Lassiter, T.L.; White, L.D. Vulnerable processes of nervous system development: a review of markers and methods. Neurotoxicology 2000, 21, 15–36. [Google Scholar]
- Longnecker, M.P.; Klebanoff, M.A.; Zhou, H.; Brock, J.W. Association between maternal serum concentration of the DDT metabolite DDE and preterm and small-for-gestational-age babies at birth. TheLancet 2001, 358, 110–114. [Google Scholar]
- Priyadarshi, A.; Khuder, S.A.; Schaub, E.A.; Shrivastava, S. A meta-analysis of Parkinson’s disease and exposure to pesticides. Neurotoxicology 2000, 21, 435–440. [Google Scholar]
- Gorell, J.M.; Johnson, C.C.; Rybicki, B.A.; Peterson, E.L.; Richardson, R.J. The risk of Parkinson’s disease with exposure to pesticides, farming, well water, and rural living. Neurology 1998, 50, 1346–1350. [Google Scholar]
- Semchuk, K.M.; Love, E.J.; Lee, R.G. Parkinson’s disease and exposure to agricultural work and pesticide chemicals. Neurology 1992, 42, 1328–1335. [Google Scholar]
- Betarbet, R.; Sherer, T.B.; MacKenzie, G.; Garcia-Osuna, M.; Panov, A.V.; Greenamyre, J.T. Chronic systemic pesticide exposure reproduces features of Parkinson’s disease. Nat. Neurosci. 2000, 3, 1301–1306. [Google Scholar]
- Sherer, T.B.; Kim, J.H.; Betarbet, R.; Greenamyre, J.T. Subcutaneous rotenone exposure causes highly selective dopaminergic degeneration and α-synuclein aggregation. Exp. Neurol. 2003, 179, 9–16. [Google Scholar]
- Kamel, F.; Hoppin, J.A. Association of pesticide exposure with neurologic dysfunction and disease. Environ. Health Perspect. 2004, 112, 950–958. [Google Scholar]
- Pain, D.J.; Gargi, R.; Cunningham, A.A.; Jones, A.; Prakash, V. Mortality of globally threatened Sarus cranes Grus antigon from monocrotophos poisoning in India. Sci. Total Environ. 2004, 326, 55–61. [Google Scholar]
- Schuler, T.H.; Denholm, I.; Jouanin, L.; Clark, S.J.; Clark, A.J.; Poppy, G.M. Population-scale laboratory studies of the effect of transgenic plants on nontarget insects. Mol. Ecol. 2001, 10, 1845–1853. [Google Scholar]
- Van Wijngaarden, R.P.; Brock, T.C.; Van den Brink, P.J. Threshold levels for effects of insecticides in freshwater ecosystems: a review. Ecotoxicology 2005, 14, 355–380. [Google Scholar]
- Dale, K. Personal communication, U.S. Environmental Protection Agency: Washington, DC, USA, 2012.
- Casida, J.E. Pest toxicology: the primary mechanisms of pesticide action. Chem. Res. Toxicol. 2009, 22, 609–619. [Google Scholar]
- Sattelle, D.B.; Cordova, D.; Cheek, T.R. Insect ryanodine receptors: molecular targets for novel pest control chemicals. Invert. Neurosci. 2008, 8, 107–119. [Google Scholar]
- Feyereisen, R. Molecular biology of insecticide resistance. Toxicol. Lett. 1995, 82–90. [Google Scholar]
- Hemingway, J.; Hawkes, N.J.; McCarroll, L.; Ranson, H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem. Mol. Biol. 2004, 34, 653–665. [Google Scholar]
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human disease. Annu. Rev. Entomol. 2000, 45, 371–391. [Google Scholar]
- World Health Organisation. Vector resistance to insecticides. 15th Report of the WHO Expert Committee on Vector Biology and Control. World Health Org. Tech. Rep. Ser. 1992, 818, 1–62.
- Georghiou, G.P. Overview of insecticide resistance. ACS Symp. Ser. 1990, 421, 18–41. [Google Scholar]
- Krogstad, D.J. Malaria as a reemerging disease. Epidemiol. Rev. 1996, 18, 77–89. [Google Scholar]
- Nauen, R.; Bretschneider, T. New modes of action of insecticides. Pestic. Outlook 2002, 13, 241–245. [Google Scholar]
- Wratten, S.D. Chapter 17: Conservation biological control and biopesticides in agriculture. In Applications in Ecological Engineering; Jorgensen, S.E., Ed.; Elsevier Academic Press: Maryland Heights, MO, USA, 2009; pp. 130–134. [Google Scholar]
- Lehr, P. Biopesticides: The Global Market; BCC Research: Wellesley, MA, USA, 2010; p. 160. [Google Scholar]
- Thakore, Y. The New Biopesticide Market; BCC Research: Wellesley, MA, USA, 2006; p. 168. [Google Scholar]
- Copping, L.; Menn, J. Biopesticides: a review of their action, applications and efficacy. Pest Manag. Sci. 2000, 56, 651–676. [Google Scholar]
- Froy, O.; Zilberberg, N.; Chejanovsky, N.; Anglister, J.; Loret, E.; Shaanan, B.; Gordon, D.; Gurevitz, M. Scorpion neurotoxins: structure/function relationships and application in agriculture. Pest Manag. Sci. 2000, 56, 472–474. [Google Scholar]
- Quistad, G.B.; Skinner, W.S. Isolation and sequencing of insecticidal peptides from the primitive hunting spider, Plectreurys tristis (Simon). J. Biol. Chem. 1994, 269, 11098–11101. [Google Scholar]
- Tomalski, M.D.; Bruce, W.A.; Travis, J.; Blum, M.S. Preliminary characterization of toxins from the straw itch mite, Pyemotes tritici, which induce paralysis in the larvae of a moth. Toxicon 1988, 26, 127–132. [Google Scholar]
- Nicholson, G.M. Fighting the global pest problem: preface to the special Toxicon issue on insecticidal toxins and their potential for insect pest control. Toxicon 2007, 49, 413–422. [Google Scholar]
- King, G.F. Modulation of insect CaV channels by peptidic spider toxins. Toxicon 2007, 49, 513–530. [Google Scholar]
- Platnick, N.I. The world spider catalog, version 12.0. American Museum of Natural History, 2012. Available online: http://research.amnh.org/iz/spiders/catalog (accessed on 20 March 2012).
- Coddington, J.A.; Levi, H.W. Systematics and evolution of spiders (Araneae). Annu. Rev. Ecol. System. 1991, 22, 565–592. [Google Scholar]
- Escoubas, P.; Quinton, L.; Nicholson, G.M. Venomics: unravelling the complexity of animal venoms with mass spectrometry. J. Mass Spectrom. 2008, 43, 279–295. [Google Scholar]
- Escoubas, P.; Rash, L.D. Tarantulas: Eight-legged pharmacists and combinatorial chemists. Toxicon 2004, 43, 555–574. [Google Scholar]
- Rash, L.D.; Hodgson, W.C. Pharmacology and biochemistry of spider venoms. Toxicon 2002, 40, 225–254. [Google Scholar]
- Tedford, H.W.; Sollod, B.L.; Maggio, F.; King, G.F. Australian funnel-web spiders: master insecticide chemists. Toxicon 2004, 43, 601–618. [Google Scholar]
- Estrada, G.; Garcia, B.I.; Schiavon, E.; Ortiz, E.; Cestèle, S.; Wanke, E.; Possani, L.D.; Corzo, G. Four disulfide-bridged scorpion beta neurotoxin CssII: heterologous expression and proper folding in vitro. Biochim. Biophys. Acta, Gen. Subj. 2007, 1770, 1161–1168. [Google Scholar] [CrossRef]
- Vassilevski, A.A.; Kozlov, S.A.; Grishin, E.V. Molecular diversity of spider venom. Biochem. Mosc. 2009, 74, 1505–1534. [Google Scholar]
- Sollod, B.L.; Wilson, D.T.; Zhaxybayeva, O.; Gogarten, J.P.; Drinkwater, R.; King, G.F. Were arachnids the first to use combinatorial peptide libraries? Peptides 2005, 26, 131–139. [Google Scholar] [CrossRef]
- Kuhn-Nentwig, L.; Stöcklin, R.; Nentwig, W. Venom composition and strategies in spiders: is everything possible? Adv. Insect Physiol. 2011, 40, 1–86. [Google Scholar] [CrossRef]
- Escoubas, P.; Sollod, B.; King, G.F. Venom landscapes: mining the complexity of spider venoms via a combined cDNA and mass spectrometric approach. Toxicon 2006, 47, 650–663. [Google Scholar]
- Escoubas, P.; King, G.F. Venomics as a drug discovery platform. Expert Rev. Proteomics 2009, 6, 221–224. [Google Scholar]
- King, G.F.; Sollod McFarland, B.; Nicholson, G.M.; Gunning, S. Insecticidal polypeptides and methods of use thereof. U.S. Patent 2011/0237502 A1, 29 September 2011. [Google Scholar]
- King, G.F.; Gentz, M.C.; Escoubas, P.; Nicholson, G.M. A rational nomenclature for naming peptide toxins from spiders and other venomous animals. Toxicon 2008, 52, 264–276. [Google Scholar]
- Herzig, V.; Wood, D.L.A.; Newell, F.; Chaumeil, P.A.; Kaas, Q.; Binford, G.J.; Nicholson, G.M.; Gorse, D.; King, G.F. ArachnoServer 2.0, an updated online resource for spider toxin sequences and structures. Nucleic Acids Res. 2011, 39, D653–D657. [Google Scholar]
- Wood, D.L.; Miljenovic, T.; Cai, S.; Raven, R.J.; Kaas, Q.; Escoubas, P.; Herzig, V.; Wilson, D.; King, G.F. ArachnoServer: a database of protein toxins from spiders. BMC Genomics 2009, 10, 375. [Google Scholar]
- Norton, R.S.; Pallaghy, P.K. The cystine knot structure of ion channel toxins and related polypeptides. Toxicon 1998, 36, 1573–1583. [Google Scholar]
- Pallaghy, P.K.; Norton, R.S.; Nielsen, K.J.; Craik, D.J. A common structural motif incorporating a cystine knot and a triple-stranded β-sheet in toxic and inhibitory polypeptides. Protein Sci. 1994, 3, 1833–1839. [Google Scholar]
- King, G.F.; Tedford, H.W.; Maggio, F. Structure and function of insecticidal neurotoxins from Australian funnel-web spiders. J. Toxicol. Toxin Rev. 2002, 21, 359–389. [Google Scholar]
- Wang, X.H.; Connor, M.; Smith, R.; Maciejewski, M.W.; Howden, M.E.H.; Nicholson, G.M.; Christie, M.J.; King, G.F. Discovery and characterization of a family of insecticidal neurotoxins with a rare vicinal disulfide bridge. Nat. Struct. Biol. 2000, 7, 505–513. [Google Scholar]
- Smith, J.J.; Hill, J.M.; Little, M.J.; Nicholson, G.M.; King, G.F.; Alewood, P.F. Unique scorpion toxin with a putative ancestral fold provides insight into evolution of the inhibitor cystine knot motif. Proc. Natl. Acad. Sci. USA 2011, 108, 10478–10483. [Google Scholar]
- Wen, S.; Wilson, D.C.; Kuruppu, S.; Korsinczky, M.L.; Hedrick, J.; Pang, L.; Szeto, T.H.; Hodgson, W.C.; Alewood, P.; Nicholson, G.M. Discovery of an MIT-like atracotoxin family: spider venom peptides that share sequence homology but not pharmacological properties with AVIT family proteins. Peptides 2005, 26, 2412–2426. [Google Scholar]
- Colgrave, M.L.; Craik, D.J. Thermal, chemical, and enzymatic stability of the cyclotide kalata B1: the importance of the cyclic cystine knot. Biochemistry 2004, 43, 5965–5975. [Google Scholar]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; Renjifo, C.; de la Vega, R.C. The toxicogenomic multiverse: convergent recruitment of proteins into animal venoms. Annu. Rev. Genomics Hum. Genet. 2009, 10, 483–511. [Google Scholar]
- Vetter, I.; Davis, J.L.; Rash, L.D.; Anangi, R.; Mobli, M.; Alewood, P.F.; Lewis, R.J.; King, G.F. Venomics: a new paradigm for natural products-based drug discovery. Amino Acids 2011, 40, 15–28. [Google Scholar]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. (Lond.) 1952, 117, 500–544. [Google Scholar]
- Hille, B. Ion Channels of Excitable Membranes, 3rd ed; Sinauer Associates: Sunderland, MA, USA, 2001; pp. 75–81. [Google Scholar]
- Payandeh, J.; Scheuer, T.; Zheng, N.; Catterall, W.A. The crystal structure of a voltage-gated sodium channel. Nature 2011, 475, 353–358. [Google Scholar]
- Catterall, W.A. From ionic currents to molecular mechanisms: the structure and function of voltage-gated sodium channels. Neuron 2000, 26, 13–25. [Google Scholar]
- Catterall, W.A. Ion channel voltage sensors: structure, function, and pathophysiology. Neuron 2010, 67, 915–928. [Google Scholar]
- West, J.W.; Patton, D.E.; Scheuer, T.; Wang, Y.; Goldin, A.L.; Catterall, W.A. A cluster of hydrophobic amino acid residues required for fast Na+-channel inactivation. Proc. Natl. Acad. Sci. USA 1992, 89, 10910–10914. [Google Scholar]
- Davies, T.G.; Field, L.M.; Usherwood, P.N.R.; Williamson, M.S. A comparative study of voltage-gated sodium channels in the Insecta: implications for pyrethroid resistance in Anopheline and other Neopteran species. Insect Mol. Biol. 2007, 16, 361–375. [Google Scholar]
- Goldin, A.L.; Barchi, R.L.; Caldwell, J.H.; Hofmann, F.; Howe, J.R.; Hunter, J.C.; Kallen, R.G.; Mandel, G.; Meisler, M.H.; Netter, Y.B.; Noda, M.; Tamkun, M.M.; Waxman, S.G.; Wood, J.N.; Catterall, W.A. Nomenclature of voltage-gated sodium channels. Neuron 2000, 28, 365–368. [Google Scholar]
- Song, W.; Liu, Z.; Tan, J.; Nomura, Y.; Dong, K. RNA editing generates tissue-specific sodium channels with distinct gating properties. J. Biol. Chem. 2004, 279, 32554–32561. [Google Scholar]
- Tan, J.; Liu, Z.; Nomura, Y.; Goldin, A.L.; Dong, K. Alternative splicing of an insect sodium channel gene generates pharmacologically distinct sodium channels. J. Neurosci. 2002, 22, 5300–5309. [Google Scholar]
- King, G.F.; Escoubas, P.; Nicholson, G.M. Peptide toxins that selectively target insect NaV and CaV channels. Channels (Austin, Tex) 2008, 2, 100–116. [Google Scholar]
- Bloomquist, J.R. Ion channels as targets for insecticides. Annu. Rev. Entomol. 1996, 41, 163–190. [Google Scholar]
- Raymond-Delpech, V.; Matsuda, K.; Sattelle, B.M.; Rauh, J.J.; Sattelle, D.B. Ion channels: Molecular targets of neuroactive insecticides. Invert. Neurosci. 2005, 5, 119–133. [Google Scholar]
- Cestèle, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar]
- Catterall, W.A.; Cestèle, S.; Yarov-Yarovoy, V.; Yu, F.H.; Konoki, K.; Scheuer, T. Voltage-gated ion channels and gating modifier toxins. Toxicon 2007, 49, 124–141. [Google Scholar] [Green Version]
- Nicholson, G.M. Insect-selective spider toxins targeting voltage-gated sodium channels. Toxicon 2007, 49, 490–512. [Google Scholar]
- Liang, S.P.; Peng, X.J.; Huang, R.H.; Chen, P. Biochemical identification of Selenocosmia hainana sp. nov. from south China (Araneae Theraphosidae). Life Sci. Res. 1999, 3, 299–303. [Google Scholar]
- Li, D.; Xiao, Y.; Hu, W.; Xie, J.; Bosmans, F.; Tytgat, J.; Liang, S.P. Function and solution structure of hainantoxin-I, a novel insect sodium channel inhibitor from the Chinese bird spider Selenocosmia hainana. FEBS Lett. 2003, 555, 616–622. [Google Scholar]
- Li, D.; Xiao, Y.; Xu, X.; Xiong, X.; Lu, S.; Liu, Z.; Zhu, Q.; Wang, M.; Gu, X.; Liang, S.P. Structure-activity relationships of hainantoxin-IV and structure determination of active and inactive sodium channel blockers. J. Biol. Chem. 2004, 279, 37734–37740. [Google Scholar]
- Figueiredo, S.G.; Garcia, M.E.; Valentim, A.C.; Cordeiro, M.N.; Diniz, C.R.; Richardson, M. Purification and amino acid sequence of the insecticidal neurotoxin Tx4(6–1) from the venom of the “armed” spider Phoneutria nigriventer (Keys). Toxicon 1995, 33, 83–93. [Google Scholar]
- de Lima, M.E.; Stankiewicz, M.; Hamon, A.; de Figueiredo, S.G.; Cordeiro, M.N.; Diniz, C.R.; Martin-Eauclaire, M.F.; Pelhate, M. The toxin Tx4(6–1) from the spider Phoneutria nigriventer slows down Na+ current inactivation in insect CNS via binding to receptor site 3. J. Insect Physiol. 2002, 48, 53–61. [Google Scholar]
- Corzo, G.; Gilles, N.; Satake, H.; Villegasa, E.; Dai, L.; Nakajimaa, T.; Haupt, J. Distinct primary structures of the major peptide toxins from the venom of the spider Macrothele gigas that bind to sites 3 and 4 in the sodium channel. FEBS Lett. 2003, 547, 43–50. [Google Scholar]
- Corzo, G.; Escoubas, P.; Stankiewicz, M.; Pelhate, M.; Kristensen, C.P.; Nakajima, T. Isolation, synthesis and pharmacological characterization of δ-palutoxins IT, novel insecticidal toxins from the spider Paracoelotes luctuosus (Amaurobiidae). Eur. J. Biochem. 2000, 267, 5783–5795. [Google Scholar]
- Ferrat, G.; Bosmans, F.; Tytgat, J.; Pimentel, C.; Chagot, B.; Gilles, N.; Nakajima, T.; Darbon, H.; Corzo, G. Solution structure of two insect-specific spider toxins and their pharmacological interaction with the insect voltage-gated Na+ channel. Proteins 2005, 59, 368. [Google Scholar]
- Corzo, G.; Escoubas, P.; Villegas, E.; Karbat, I.; Gordon, D.; Gurevitz, M.; Nakajima, T.; Gilles, N. A spider toxin that induces a typical effect of scorpion α-toxins but competes with β-toxins on binding to insect sodium channels. Biochimie 2005, 44, 1542–1549. [Google Scholar]
- Cohen, L.; Karbat, I.; Gilles, N.; Froy, O.; Corzo, G.; Angelovici, R.; Gordon, D.; Gurevitz, M. Dissection of the functional surface of an anti-insect excitatory toxin illuminates a putative “hot spot” common to all scorpion β-toxins affecting Na+ channels. J. Biol. Chem. 2004, 279, 8206–8211. [Google Scholar]
- Adams, M.E.; Herold, E.E.; Venema, V.J. Two classes of channel-specific toxins from funnel web spider venom. J. Comp. Physiol. (A) 1989, 164, 333–342. [Google Scholar] [CrossRef]
- Skinner, W.S.; Adams, M.E.; Quistad, G.B.; Kataoka, H.; Cesarin, B.J.; Enderlin, F.E.; Schooley, D.A. Purification and characterization of two classes of neurotoxins from the funnel web spider, Agelenopsis aperta. J. Biol. Chem. 1989, 264, 2150–2155. [Google Scholar]
- Kozlov, S.; Malyavka, A.; McCutchen, B.; Lu, A.; Schepers, E.; Herrmann, R.; Grishin, E. A novel strategy for the identification of toxinlike structures in spider venom. Proteins 2005, 59, 131–140. [Google Scholar]
- Quistad, G.B.; Reuter, C.C.; Skinner, W.S.; Dennis, P.A.; Suwanrumpha, S.; Fu, E.W. Paralytic and insecticidal toxins from the funnel web spider, Hololena curta. Toxicon 1991, 29, 329–336. [Google Scholar]
- Stapleton, A.; Blankenship, D.; Ackermann, B.; Chen, T.; Gorder, G.; Manley, G.; Palfreyman, M.; Coutant, J.; Cardin, A.C. Neurotoxic insecticidal polypeptides isolated from the funnel-web spider Hololena curta. J. Biol. Chem. 1990, 265, 2054–2059. [Google Scholar]
- Cohen, C.; Bale, T.; Ertel, E.; Warren, V.; Smith, M. μ-Aga-IV: a spider toxin specific for insect Na channels. Biophys. J. 1993, 64, A4. [Google Scholar]
- Norris, T.; Lee, A.; Adams, M.E. Modulation of sodium channels by insect-selective scorpion and spider toxins. Soc. Neurosci. Abstr. 1995, 21, 1820. [Google Scholar]
- Wang, G.; Strichartz, G. Purification and physiological characterization of neurotoxins from venoms of the scorpions Centruroides sculpturatus and Leiurus quinquestriatus. Mol. Pharmacol. 1983, 23, 519–533. [Google Scholar]
- Krapcho, K.J.; Kral, R.M.; Vanwagenen, B.C.; Eppler, K.G.; Morgan, T.K. Characterization and cloning of insecticidal peptides from the primitive weaving spider Diguetia canities. Insect Biochem. Mol. Biol. 1995, 25, 991–1000. [Google Scholar]
- Bloomquist, J.R.; Kinne, L.P.; Deutsch, V.; Simpson, S.F. Mode of action of an insecticidal peptide toxin from the venom of a weaving spider (Diguetia canities). Toxicon 1996, 34, 1072–1075. [Google Scholar]
- Catterall, W.A.; Perez-Reyes, E.; Snutch, T.P.; Striessnig, J. International Union of Pharmacology. XLVIII. Nomenclature and structure-function relationships of voltage-gated calcium channels. Pharmacol. Rev. 2005, 57, 411–425. [Google Scholar] [CrossRef]
- Bourinet, E.; Zamponi, G.W. Voltage gated calcium channels as targets for analgesics. Curr. Top. Med. Chem. 2005, 5, 539–546. [Google Scholar]
- Tanabe, T.; Takeshima, H.; Mikami, A.; Flockerzi, V.; Takahashi, H.; Kangawa, K.; Kojima, M.; Matsuo, H.; Hirose, T.; Numa, S. Primary structure of the receptor for calcium channel blockers from skeletal muscle. Nature 1987, 328, 313–318. [Google Scholar]
- Littleton, J.T.; Ganetzky, B. Ion channels and synaptic organization: analysis of the Drosophila genome. Neuron 2000, 26, 35–43. [Google Scholar]
- Eberl, D.F.; Ren, D.; Feng, G.; Lorenz, L.J.; Van Vactor, D.; Hall, L.M. Genetic and developmental characterization of Dmca1D, a calcium channel α1 subunit gene in Drosophila melanogaster. Genetics 1998, 148, 1159–1169. [Google Scholar]
- Kawasaki, F.; Collins, S.C.; Ordway, R.W. Synaptic calcium-channel function in Drosophila: analysis and transformation rescue of temperature-sensitive paralytic and lethal mutations of cacophony. J. Neurosci. 2002, 22, 5856–5864. [Google Scholar]
- Wicher, D.; Walther, C.; Wicher, C. Non-synaptic ion channels in insects-basic properties of currents and their modulation in neurons and skeletal muscles. Prog. Neurobiol. 2001, 64, 431–525. [Google Scholar]
- Fletcher, J.I.; Smith, R.; O’Donoghue, S.I.; Nilges, M.; Connor, M.; Howden, M.E.; Christie, M.J.; King, G.F. The structure of a novel insecticidal neurotoxin, ω-atracotoxin-HV1, from the venom of an Australian funnel web spider. Nat. Struct. Biol. 1997, 4, 559–566. [Google Scholar]
- Atkinson, R.; Vonarx, E.; Howden, M. Effects of whole venom and venom fractions from several Australian spiders, including Atrax (Hadronyche) species, when injected into insects. Comp. Biochem. Physiol. 1996, 114, 113–117. [Google Scholar]
- Wang, X.H.; Smith, R.; Fletcher, J.I.; Wilson, H.; Wood, C.J.; Howden, M.E.; King, G.F. Structure-function studies of ω-atracotoxin, a potent antagonist of insect voltage-gated calcium channels. Eur. J. Biochem. 1999, 264, 488–494. [Google Scholar]
- Chong, Y.; Hayes, J.L.; Sollod, B.; Wen, S.; Wilson, D.T.; Hains, P.G.; Hodgson, W.C.; Broady, K.W.; King, G.F.; Nicholson, G.M. The ω-atracotoxins: selective blockers of insect M-LVA and HVA calcium channels. Biochem. Pharmacol. 2007, 74, 623–638. [Google Scholar]
- Bloomquist, J.R. Mode of action of atracotoxin at central and peripheral synapses of insects. Invert. Neurosci. 2003, 5, 45–50. [Google Scholar]
- Khan, S.A.; Zafar, Y.; Briddon, R.W.; Malik, K.A.; Mukhtar, Z. Spider venom toxin protects plants from insect attack. Transgenic Res. 2006, 15, 349–357. [Google Scholar]
- Tedford, H.W.; Maggio, F.; Reenan, R.A.; King, G.F. A model genetic system for testing the in vivo function of peptide toxins. Peptides 2007, 28, 51–56. [Google Scholar]
- Tedford, H.W.; Gilles, N.; Ménez, A.; Doering, C.J.; Zamponi, G.W.; King, G.F. Scanning mutagenesis of ω-atracotoxin-Hv1a reveals a spatially restricted epitope that confers selective activity against insect calcium channels. J. Biol. Chem. 2004, 279, 44133–44140. [Google Scholar]
- Mukherjee, A.K.; Sollod, B.L.; Wikel, S.K.; King, G.F. Orally active acaricidal peptide toxins from spider venom. Toxicon 2006, 47, 182–187. [Google Scholar]
- Branton, W.D.; Rudnick, M.S.; Zhou, Y.; Eccleston, E.D.; Fields, G.B.; Bowers, L.D. Fatty acylated toxin structure. Nature 1993, 365, 496–497. [Google Scholar]
- Branton, W.D.; Kolton, L.; Jan, Y.; Jan, L. Neurotoxins from Plectreurys spider venom are potent presynaptic blockers in Drosophila. J. Neurosci. 1987, 7, 4195–4200. [Google Scholar]
- Bodi, J.; Nishio, H.; Zhou, Y.; Branton, W.D.; Kimura, T.; Sakakibara, S. Synthesis of an O-palmitoylated 44-residue peptide amide (PLTX II) blocking presynaptic calcium channels in Drosophila. Peptide Res. 1995, 8, 228–235. [Google Scholar]
- Leung, H.T.; Branton, W.D.; Phillips, H.S.; Jan, L.; Byerly, L. Spider toxins selectively block calcium currents in Drosophila. Neuron 1989, 3, 767–772. [Google Scholar]
- Kuromi, H.; Honda, A.; Kidokoro, Y. Ca2+ influx through distinct routes controls exocytosis and endocytosis at Drosophila presynaptic terminals. Neuron 2004, 41, 101–111. [Google Scholar]
- Wang, X.H.; Connor, M.; Wilson, D.C.; Wilson, H.I.; Nicholson, G.M.; Smith, R.; Shaw, D.; Mackay, J.P.; Alewood, P.F.; Christie, M.J.; King, G.F. Discovery and structure of a potent and highly specific blocker of insect calcium channels. J. Biol. Chem. 2001, 276, 40306–40312. [Google Scholar]
- Zhang, P.F.; Chen, P.; Hu, W.J.; Liang, S.P. Huwentoxin-V, a novel insecticidal peptide toxin from the spider Selenocosmia huwena, and a natural mutant of the toxin: indicates the key amino acid residues related to the biological activity. Toxicon 2003, 42, 15–20. [Google Scholar]
- Deng, M.; Luo, X.; Meng, E.; Xiao, Y.; Liang, S. Inhibition of insect calcium channels by huwentoxin-V, a neurotoxin from Chinese tarantula Ornithoctonus huwena venom. Eur. J. Pharmacol. 2008, 582, 12–16. [Google Scholar]
- Wei, A.D.; Gutman, G.A.; Aldrich, R.; Chandy, K.G.; Grissmer, S.; Wulff, H. International Union of Pharmacology. LII. Nomenclature and molecular relationships of calcium-activated potassium channels. Pharmacol. Rev. 2005, 57, 463–472. [Google Scholar] [CrossRef]
- Coetzee, W.A.; Amarillo, Y.; Chiu, J.; Chow, A.; Lau, D.; McCormack, T.; Moreno, H.; Nadal, M.S.; Ozaita, A.; Pountney, D.; Saganich, M.; Vega-Saenz de Miera, E.; Rudy, B. Molecular diversity of K+ channels. Ann. N. Y. Acad. Sci. 1999, 868, 233–285. [Google Scholar]
- Chandy, K.G.; Gutman, G.A. Nomenclature for mammalian potassium channel genes. Trends Pharmacol. Sci. 1993, 14, 434. [Google Scholar]
- Adelman, J.P.; Shen, K.Z.; Kavanaugh, M.P.; Warren, R.A.; Wu, Y.N.; Lagrutta, A.; Bond, C.T.; North, R.A. Calcium-activated potassium channels expressed from cloned complementary DNAs. Neuron 1992, 9, 209–216. [Google Scholar]
- Meera, P.; Wallner, M.; Song, M.; Toro, L. Large conductance voltage- and calcium-dependent K+ channel, a distinct member of voltage-dependent ion channels with seven N-terminal transmembrane segments (S0–S6), an extracellular N terminus, and an intracellular (S9–S10) C terminus. Proc. Natl. Acad. Sci. USA 1997, 94, 14066–14071. [Google Scholar]
- Pongs, O.; Leicher, T.; Berger, M.; Roeper, J.; Bähring, R.; Wray, D.; Giese, K.P.; Silva, A.J.; Storm, J.F. Functional and molecular aspects of voltage-gated K+ channel β subunits. Ann. N. Y. Acad. Sci. 1999, 868, 344–355. [Google Scholar]
- Gunning, S.J.; Maggio, F.; Windley, M.J.; Valenzuela, S.M.; King, G.F.; Nicholson, G.M. The Janus-faced atracotoxins are specific blockers of invertebrate KCa channels. FEBS J. 2008, 275, 4045–4059. [Google Scholar]
- Maggio, F.; King, G.F. Scanning mutagenesis of a Janus-faced atracotoxin reveals a bipartite surface patch that is essential for neurotoxic function. J. Biol. Chem. 2002, 277, 22806–22813. [Google Scholar]
- Maggio, F.; King, G.F. Role of the structurally disordered N- and C-terminal residues in the Janus-faced atracotoxins. Toxicon 2002, 40, 1355–1361. [Google Scholar]
- Diochot, S. Precious natural peptides from spider venoms: new tools for studying potassium channels. Toxin Rev. 2005, 24, 289–312. [Google Scholar]
- Corzo, G.; Diego-García, E.; Clement, H.; Peigneur, S.; Odell, G.; Tytgat, J.; Possani, L.D.; Alagón, A. An insecticidal peptide from the theraposid Brachypelma smithi spider venom reveals common molecular features among spider species from different genera. Peptides 2008, 29, 1901–1908. [Google Scholar]
- Windley, M.J.; Escoubas, P.; Valenzuela, S.M.; Nicholson, G.M. A novel family of insect-selective peptide neurotoxins targeting insect large-conductance calcium-activated K+ channels isolated from the venom of the theraphosid spider Eucratoscelus constrictus. Mol. Pharmacol. 2011, 80, 1–13. [Google Scholar]
- Vassilevski, A.A.; Kozlov, S.A.; Samsonova, O.V.; Egorova, N.S.; Karpunin, D.V.; Pluzhnikov, K.A.; Feofanov, A.V.; Grishin, E.V. Cyto-insectotoxins, a novel class of cytolytic and insecticidal peptides from spider venom. Biochem. J. 2008, 411, 687–696. [Google Scholar]
- Grishin, E.V. Black widow spider toxins: the present and the future. Toxicon 1998, 36, 1693–1701. [Google Scholar]
- Graudins, A.; Little, M.J.; Pineda, S.S.; Hains, P.G.; King, G.F.; Broady, K.W.; Nicholson, G.M. Cloning and activity of a novel α-latrotoxin from red-back spider venom. Biochem. Pharmacol. 2011, 83, 170–183. [Google Scholar]
- Ushkaryov, Y.A.; Rohou, A.; Sugita, S. α-Latrotoxin and its receptors. Pharmacol. Neurotransmitter Release 2008, 184, 171–206. [Google Scholar] [CrossRef]
- Krasnoperov, V.G.; Shamotienko, O.G.; Grishin, E.V. A crustacean-specific neurotoxin from the venom of the black widow spider Latrodectus mactans tredecimguttatus. Biorgan. Khim. 1990, 16, 1567–1569. [Google Scholar]
- Kiyatkin, N.I.; Dulubova, I.E.; Grishin, E.V. Cloning and structural analysis of α-latroinsectotoxin cDNA. Abundance of ankyrin-like repeats. Eur. J. Biochem. 1993, 213, 121–127. [Google Scholar] [CrossRef]
- Dulubova, I.E.; Krasnoperov, V.G.; Khvotchev, M.V.; Pluzhnikov, K.A.; Volkova, T.M.; Grishin, E.V.; Vais, H.; Bell, D.R.; Usherwood, P.N. Cloning and structure of δ-latroinsectotoxin, a novel insect-specific member of the latrotoxin family: functional expression requires C-terminal truncation. J. Biol. Chem. 1996, 271, 7535–7543. [Google Scholar]
- Rohou, A.; Nield, J.; Ushkaryov, Y.A. Insecticidal toxins from black widow spider venom. Toxicon 2007, 49, 531–549. [Google Scholar]
- Krasnoperov, V.G.; Shamotienko, O.G.; Grishin, E.V. Isolation and properties of insect-specific neurotoxins from venoms of the spider Lactodectus mactans tredecimguttatus. Biorgan. Khim. 1990, 16, 1138–1140. [Google Scholar]
- Stawski, P.; Janovjak, H.; Trauner, D. Pharmacology of ionotropic glutamate receptors: A structural perspective. Bioorg. Med. Chem. 2010, 18, 7759–7772. [Google Scholar]
- Nicoletti, F.; Bockaert, J.; Collingridge, G.L.; Conn, P.J.; Ferraguti, F.; Schoepp, D.D.; Wroblewski, J.T.; Pin, J.P. Metabotropic glutamate receptors: from the workbench to the bedside. Neuropharmacology 2011, 60, 1017–1041. [Google Scholar]
- Usherwood, P.N.R. Insect glutamate receptors. Adv. Insect Physiol. 1994, 24, 309–339. [Google Scholar]
- DiAntonio, A. Glutamate receptors at the Drosophila neuromuscular junction. Int. Rev. Neurobiol. 2006, 75, 165–179. [Google Scholar]
- Raymond, V.; Sattelle, D.B.; Lapied, B. Co-existence in DUM neurones of two GluCl channels that differ in their picrotoxin sensitivity. Neuroreport 2000, 11, 2695–2701. [Google Scholar]
- Kane, N.S.; Hirschberg, B.; Qian, S.; Hunt, D.; Thomas, B.; Brochu, R.; Ludmerer, S.W.; Zheng, Y.; Smith, M.; Arena, J.P.; Cohen, C.J.; Schmatz, D.; Warmke, J.; Cully, D.F. Drug-resistant Drosophila indicate glutamate-gated chloride channels are targets for the antiparasitics nodulisporic acid and ivermectin. Proc. Natl. Acad. Sci. USA 2000, 97, 13949–13954. [Google Scholar]
- Adams, M.E. Agatoxins: ion channel specific toxins from the American funnel web spider, Agelenopsis aperta. Toxicon 2004, 43, 509–525. [Google Scholar]
- de Figueiredo, S.G.; de Lima, M.E.; Nascimento Cordeiro, M.; Diniz, C.R.; Patten, D.; Halliwell, R.F.; Gilroy, J.; Richardson, M. Purification and amino acid sequence of a highly insecticidal toxin from the venom of the brazilian spider Phoneutria nigriventer which inhibits NMDA-evoked currents in rat hippocampal neurones. Toxicon 2001, 39, 309–317. [Google Scholar]
- Nicholson, G.M.; Graudins, A.; Wilson, H.I.; Little, M.; Broady, K.W. Arachnid toxinology in Australia: from clinical toxicology to potential applications. Toxicon 2006, 48, 872–898. [Google Scholar]
- Hughes, P.R.; Wood, H.A.; Breen, J.P.; Simpson, S.F.; Duggan, A.J.; Dybas, J.A. Enhanced bioactivity of recombinant baculoviruses expressing insect-specific spider toxins in lepidopteran crop pests. J. Invert. Pathol. 1997, 69, 112–118. [Google Scholar]
- Thiem, S.M. Prospects for altering host range for baculovirus bioinsecticides. Curr. Opin. Biotechnol. 1997, 8, 317–322. [Google Scholar]
- Elazar, M.; Levi, R.; Zlotkin, E. Targeting of an expressed neurotoxin by its recombinant baculovirus. J. Exp. Biol. 2001, 204, 2637–2645. [Google Scholar]
- Kamita, S.G.; Kang, K.D.; Hammock, B.D.; Inceoglu, A.B. Genetically modified baculoviruses for pest insect control. In Comprehensive Molecular Insect Science; Gilbert, L.I., Iatrou, K., Gill, S.S., Eds.; Elsevier: Amsterdam, The Netherland, 2005; pp. 271–322. [Google Scholar]
- Prikhod’ko, G.; Popham, H.; Felcetto, T.; Ostlind, D.; Warren, V.; M.M., S.; Garsky, V.M.; Warmke, J.W.; Cohen, C.; Miller, L. Effects of simultaneous expression of two sodium channel toxin genes on the properties of baculoviruses as biopesticides. Biol. Control 1998, 12, 66–78. [Google Scholar]
- Prikhod’ko, G.; Robson, M.; Warmke, J.; Cohen, C.; Smith, M.; Wang, P.; Warren, V.; Kaczorowski, G.; Van der Ploeg, L.; Miller, L. Properties of three baculovirus-expressing genes that encode insect-selective toxins: μ-Aga-IV, As II, and Sh I. Biol. Control 1996, 7, 236–244. [Google Scholar]
- Tomalski, M.D.; Kutney, R.; Bruce, W.A.; Brown, M.R.; Blum, M.S.; Travis, J. Purification and characterization of insect toxins derived from the mite, Pyemotes tritici. Toxicon 1989, 27, 1151–1167. [Google Scholar]
- Wang, C.; St Leger, R.J. A scorpion neurotoxin increases the potency of a fungal insecticide. Nat. Biotechnol. 2007, 25, 1455–1456. [Google Scholar]
- Bravo, A.; Gill, S.S.; Soberón, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar]
- Qaim, M.; Zilberman, D. Yield effects of genetically modified crops in developing countries. Science 2003, 299, 900–902. [Google Scholar]
- Huang, J.; Rozelle, S.; Pray, C.; Wang, Q. Plant biotechnology in China. Science 2002, 295, 674–676. [Google Scholar]
- Fitches, E.; Philip, J.; Hinchliffe, G.; Vercruysse, L.; Chougule, N.; Gatehouse, J.A. An evaluation of garlic lectin as an alternative carrier domain for insecticidal fusion proteins. Insect Sci. 2008, 15, 483–495. [Google Scholar]
- Fitches, E.; Wiles, D.; Douglas, A.E.; Hinchliffe, G.; Audsley, N.; Gatehouse, J.A. The insecticidal activity of recombinant garlic lectins towards aphids. Insect Biochem. Mol. Biol. 2008, 38, 905–915. [Google Scholar]
- Fitches, E.; Edwards, M.G.; Mee, C.; Grishin, E.; Gatehouse, A.M.; Edwards, J.; Gatehouse, J.A. Fusion proteins containing insect-specific toxins as pest control agents: snowdrop lectin delivers fused insecticidal spider venom toxin to insect haemolymph following oral ingestion. J. Insect Physiol. 2004, 50, 61–71. [Google Scholar]
- Down, R.E.; Fitches, E.C.; Wiles, D.P.; Corti, P.; Bell, H.A.; Gatehouse, J.A.; Edwards, J. Insecticidal spider venom toxin fused to snowdrop lectin is toxic to the peach-potato aphid, Myzus persicae (Hemiptera: Aphididae) and the rice brown planthopper, Nilaparvata lugens (Hemiptera: Delphacidae). Pest Manag. Sci. 2006, 62, 77–85. [Google Scholar]
- Pence, R.J. The antimetabolite imidazole as a pesticide. California Agric. 1965, 13–15. [Google Scholar]
- Baell, J.B.; Duggan, P.; Forsyth, S.; Lewis, R.; Lok, Y.; Schroeder, C.; Shepherd, N. Synthesis and biological evaluation of anthranilamide-based non-peptide mimics of ω-conotoxin GVIA. Tetrahedron 2006, 62, 7284–7292. [Google Scholar]
- Menzler, S.; Bikker, J.A.; Suman-Chauhan, N.; Horwell, D.C. Design and biological evaluation of non-peptide analogues of ω-conotoxin MVIIA. Bioorg. Med. Chem. Lett. 2000, 10, 345–347. [Google Scholar]
- McCutchen, B.F.; Hoover, K.; Preisler, H.K.; Betana, M.D.; Herrmann, R.; Robertson, J.L.; Hammock, B.D. Interactions of recombinant and wild-type baculoviruses with classical insecticides and pyrethroid-resistant tobacco budworm (Lepidoptera: Noctuidae). J. Econ. Entomol. 1997, 90, 1170–1180. [Google Scholar]
- Elzen, G.W.; Hardee, D.D. United States Department of Agriculture–Agricultural Research Service research on managing insect resistance to insecticides. Pest Manag. Sci. 2003, 59, 770–776. [Google Scholar]
- Whetstone, P.A.; Hammock, B.D. Delivery methods for peptide and protein toxins in insect control. Toxicon 2007, 49, 576–596. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Windley, M.J.; Herzig, V.; Dziemborowicz, S.A.; Hardy, M.C.; King, G.F.; Nicholson, G.M. Spider-Venom Peptides as Bioinsecticides. Toxins 2012, 4, 191-227. https://doi.org/10.3390/toxins4030191
Windley MJ, Herzig V, Dziemborowicz SA, Hardy MC, King GF, Nicholson GM. Spider-Venom Peptides as Bioinsecticides. Toxins. 2012; 4(3):191-227. https://doi.org/10.3390/toxins4030191
Chicago/Turabian StyleWindley, Monique J., Volker Herzig, Sławomir A. Dziemborowicz, Margaret C. Hardy, Glenn F. King, and Graham M. Nicholson. 2012. "Spider-Venom Peptides as Bioinsecticides" Toxins 4, no. 3: 191-227. https://doi.org/10.3390/toxins4030191