Natural Toxins for Use in Pest Management

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
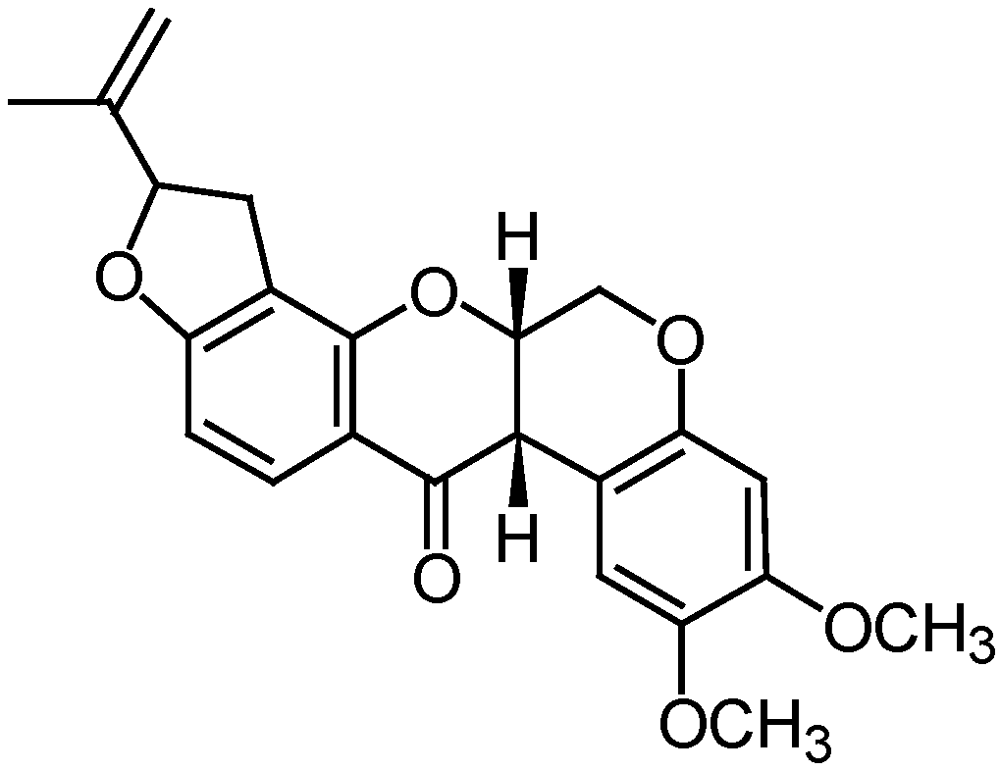{kind=link}
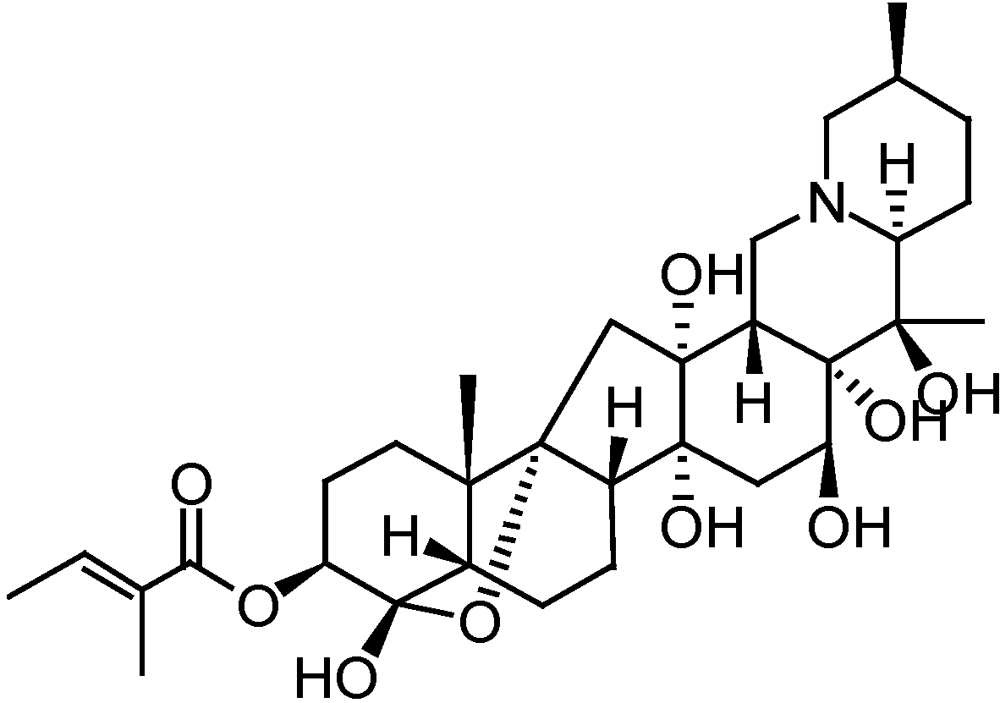{kind=link}
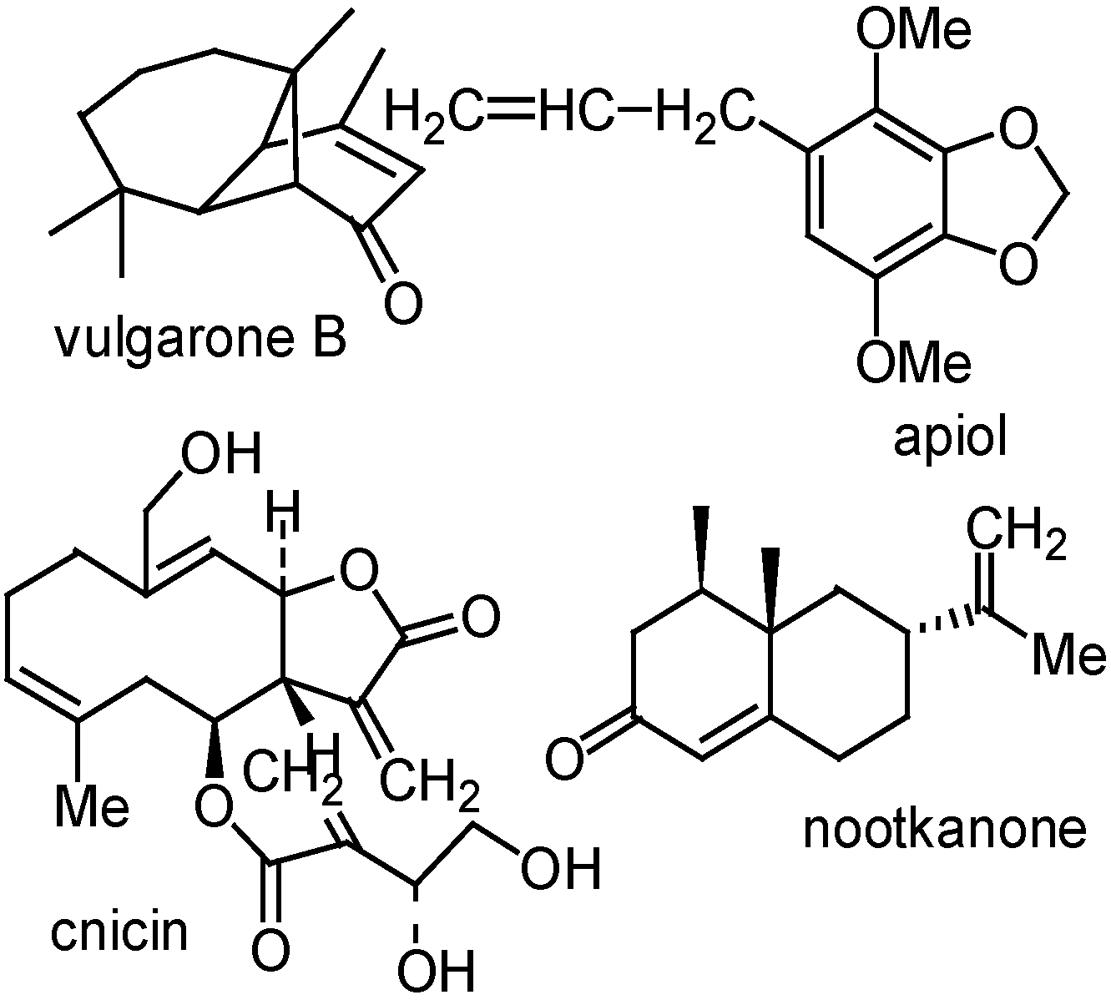{kind=link}
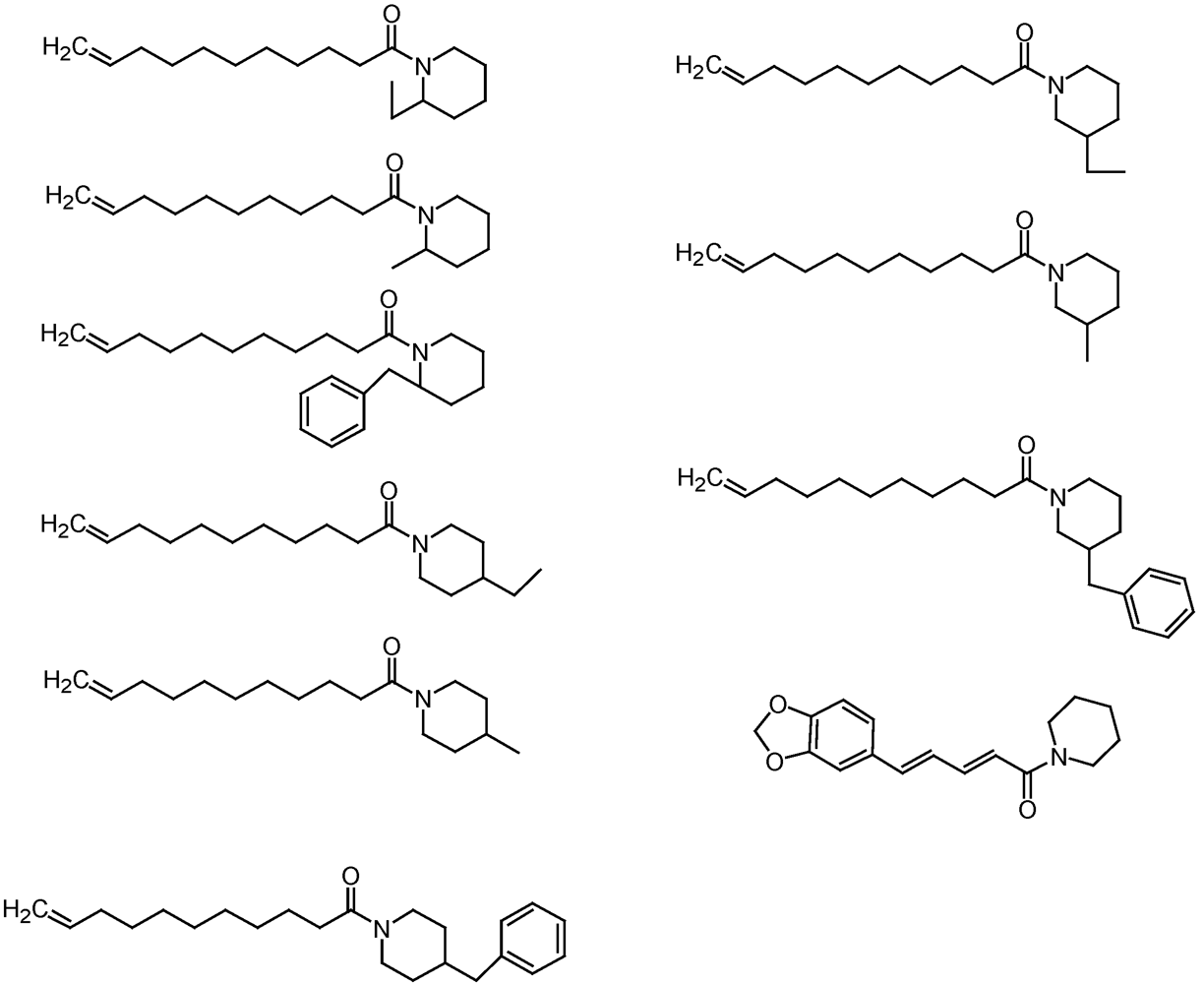{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Weed Management
2.1. Overview
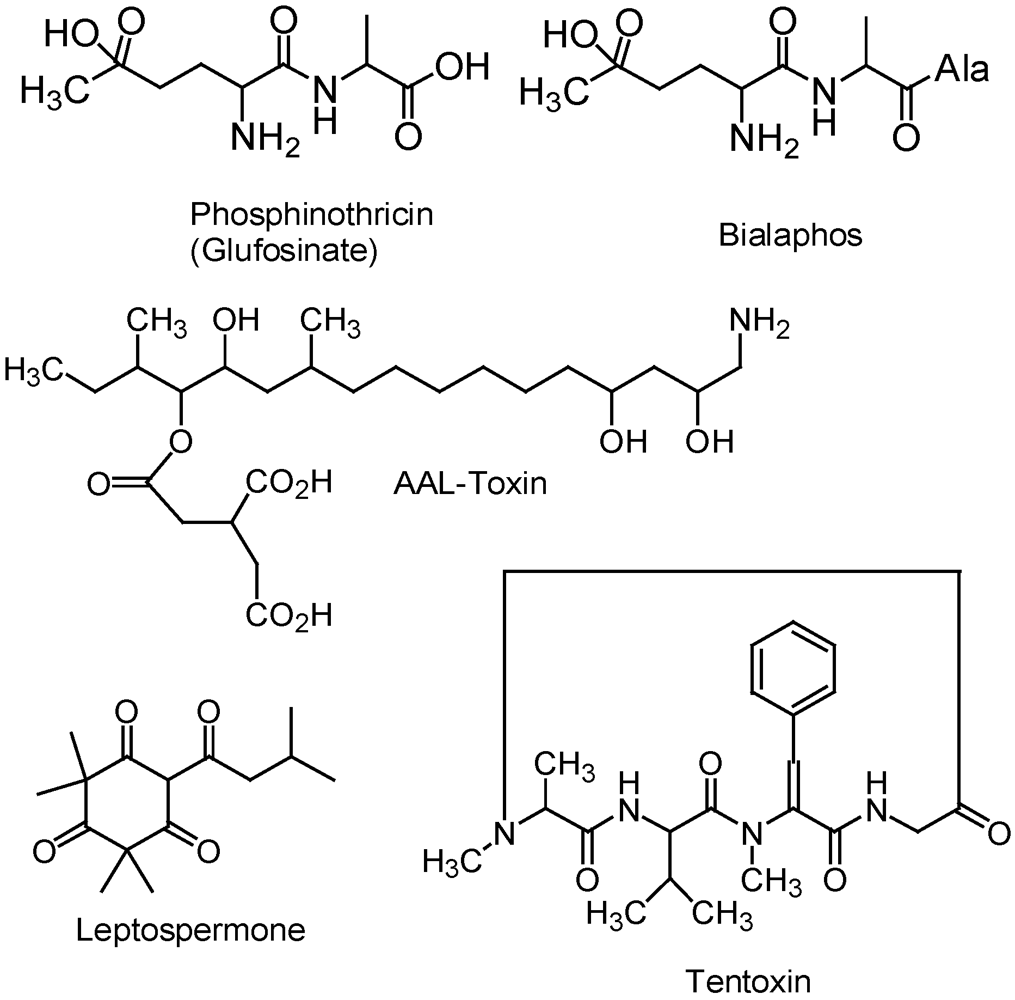
2.2. Phosphinothricin and Glufosinate
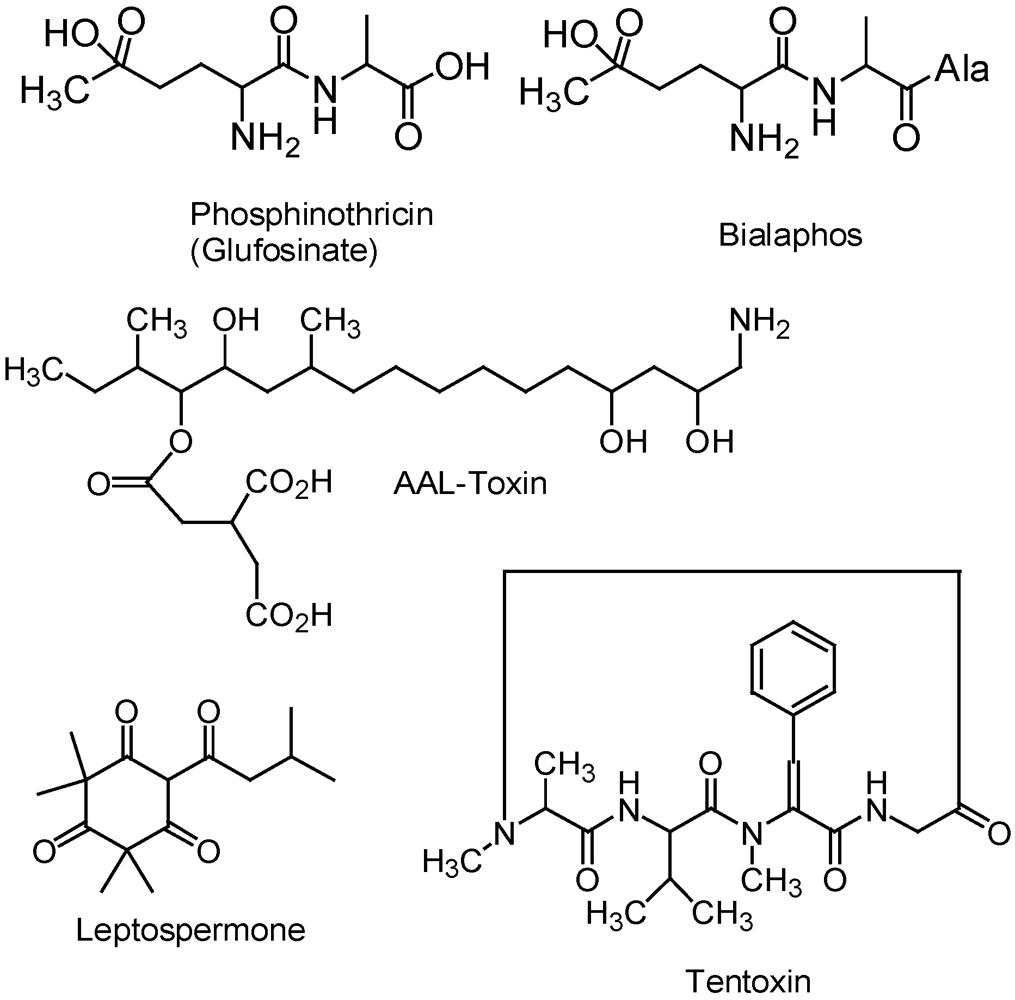
2.3. Phytotoxins from Microbes
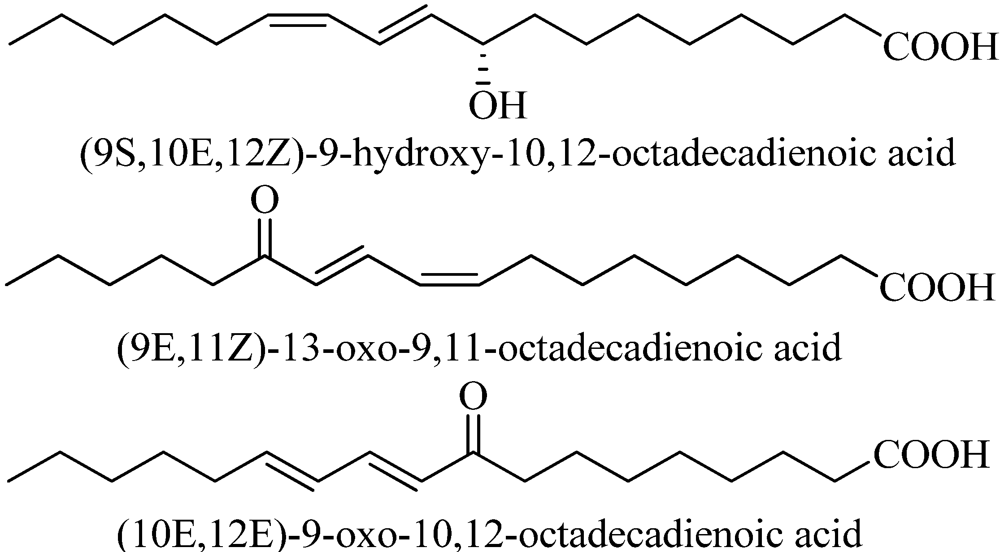
2.4. Allelochemicals
3. Insect Management
3.1. Overview
3.2. Insecticides for Agricultural Use
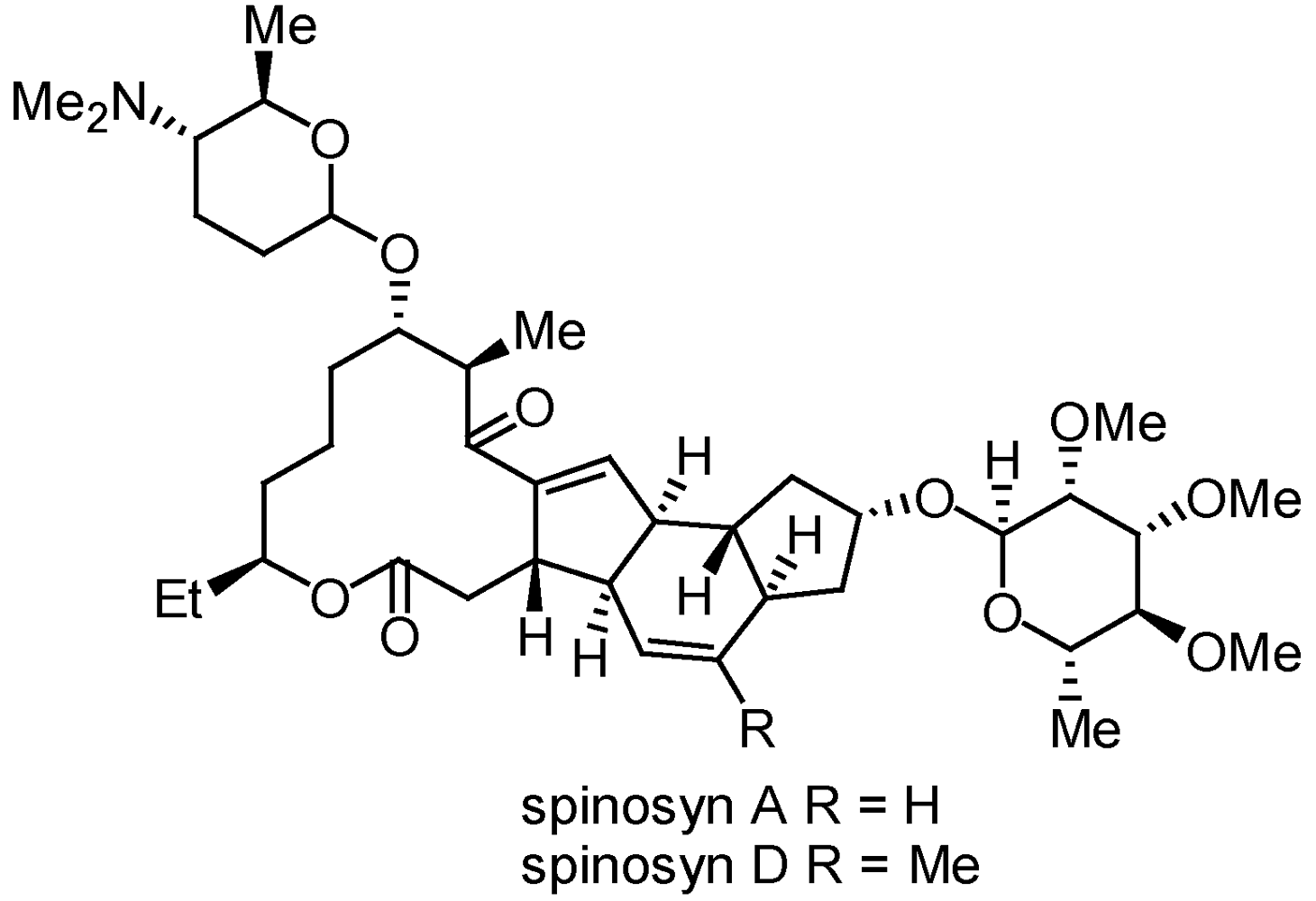
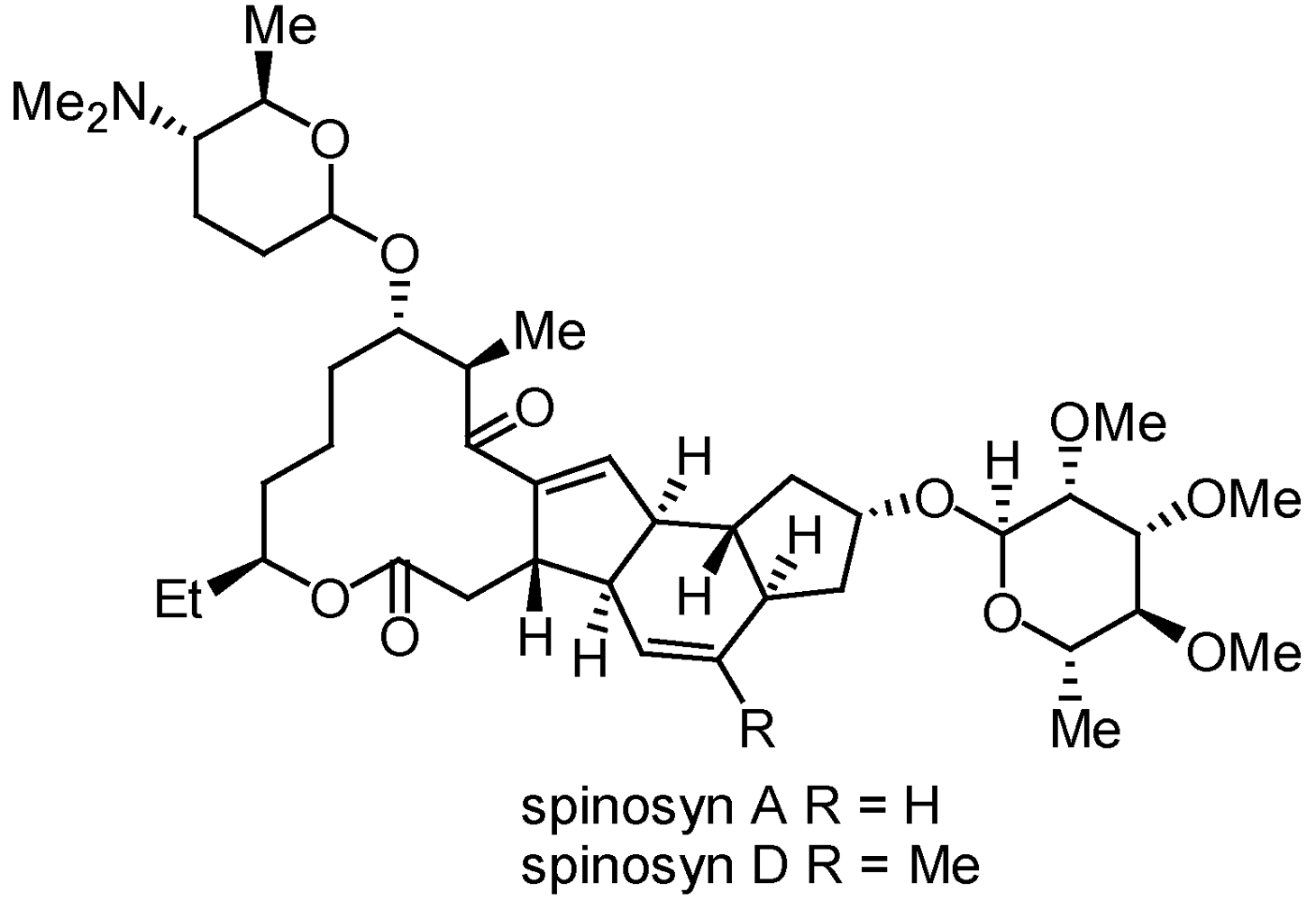
3.2.1. Spinosads
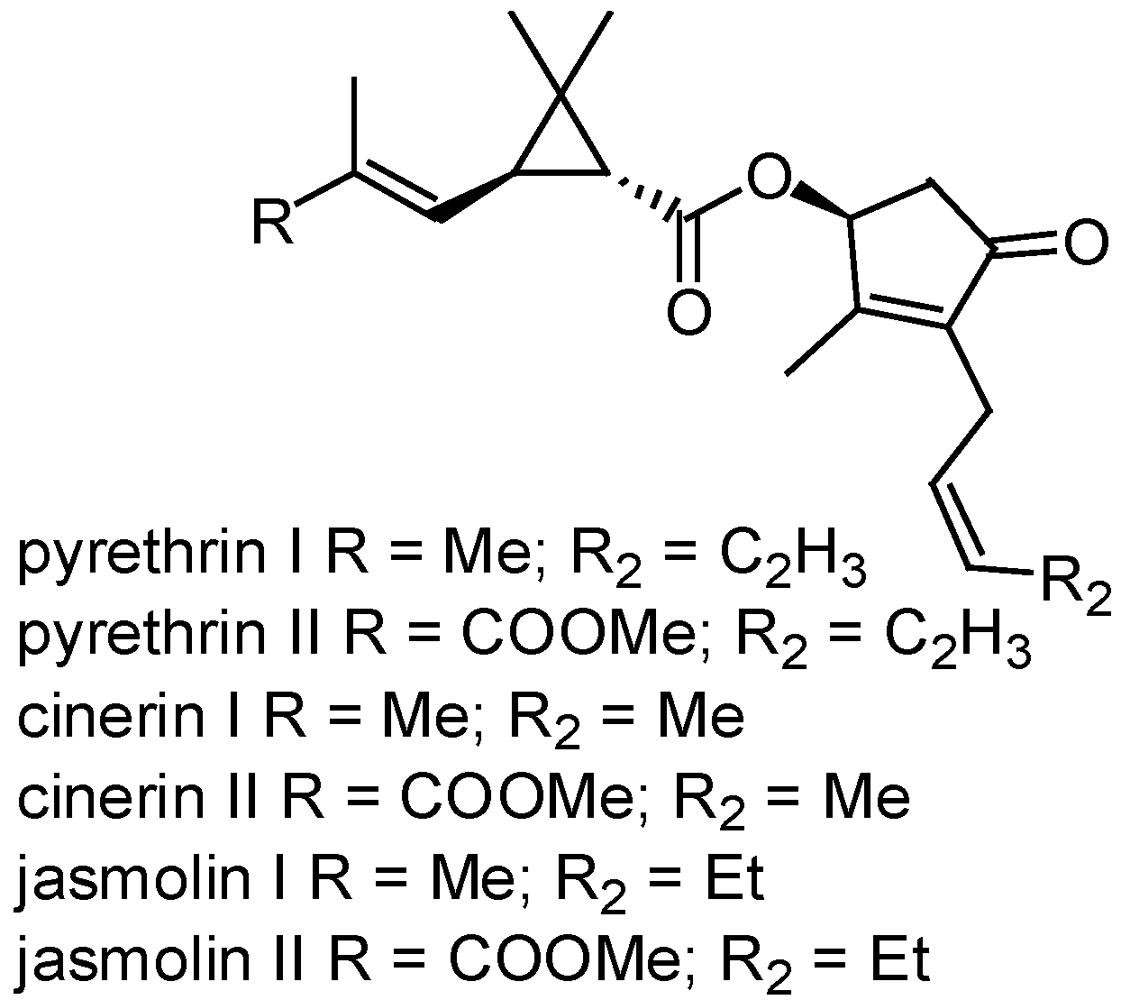
3.2.2. Pyrethrum
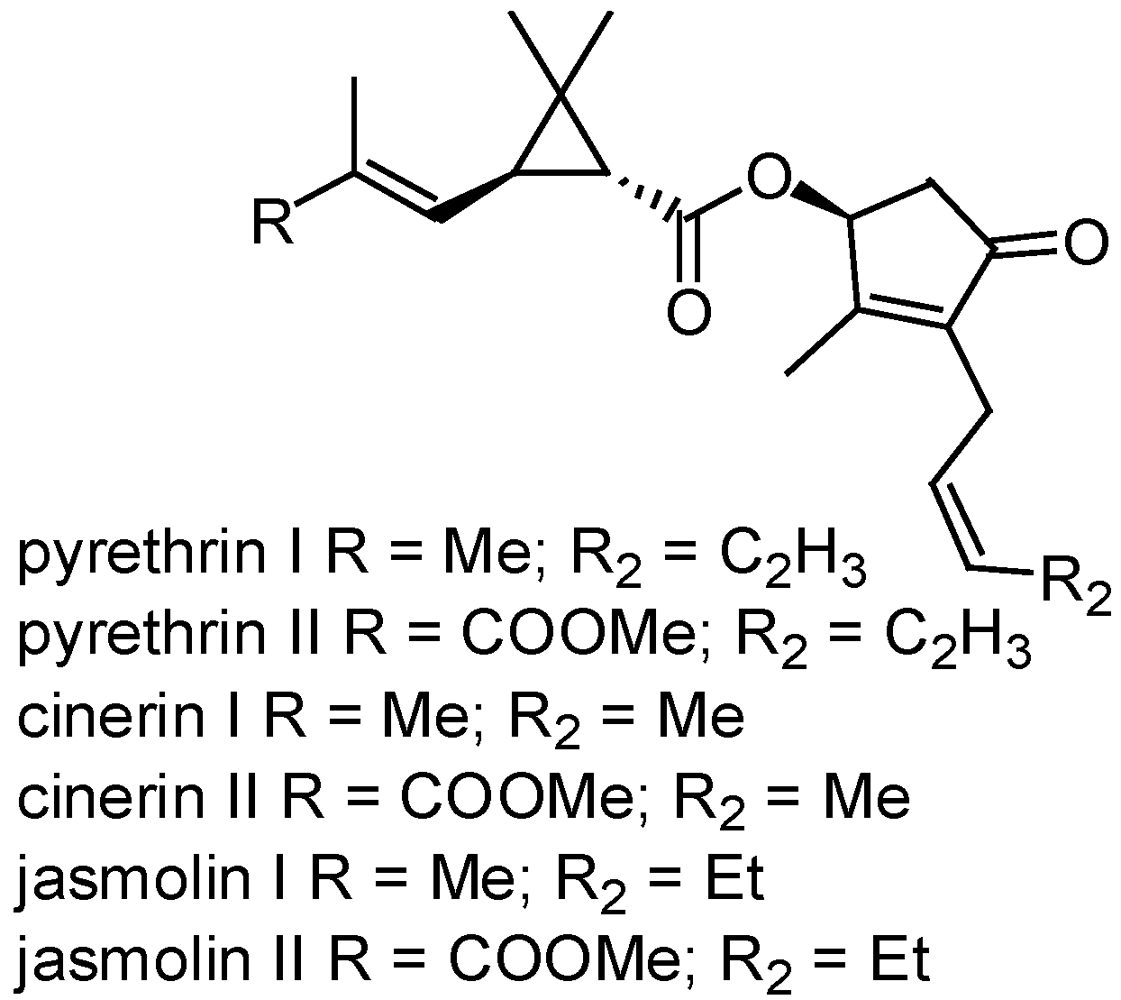
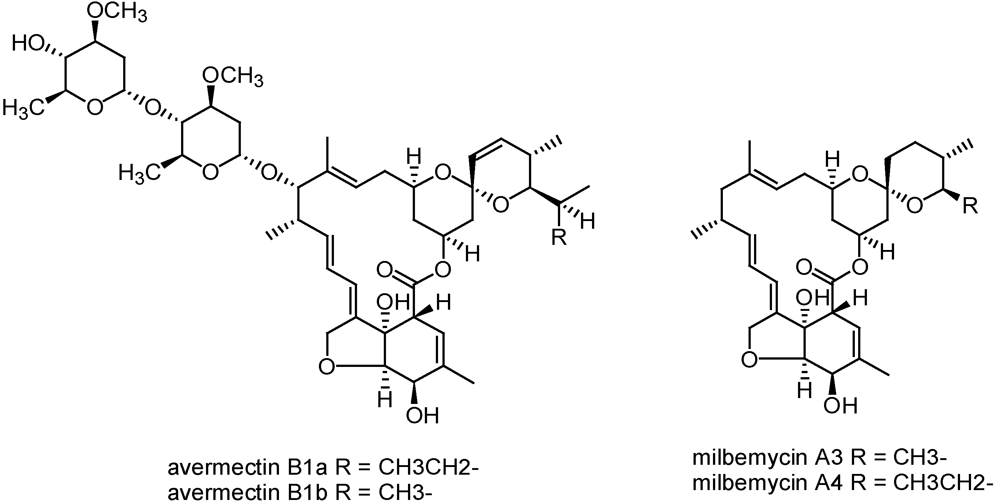
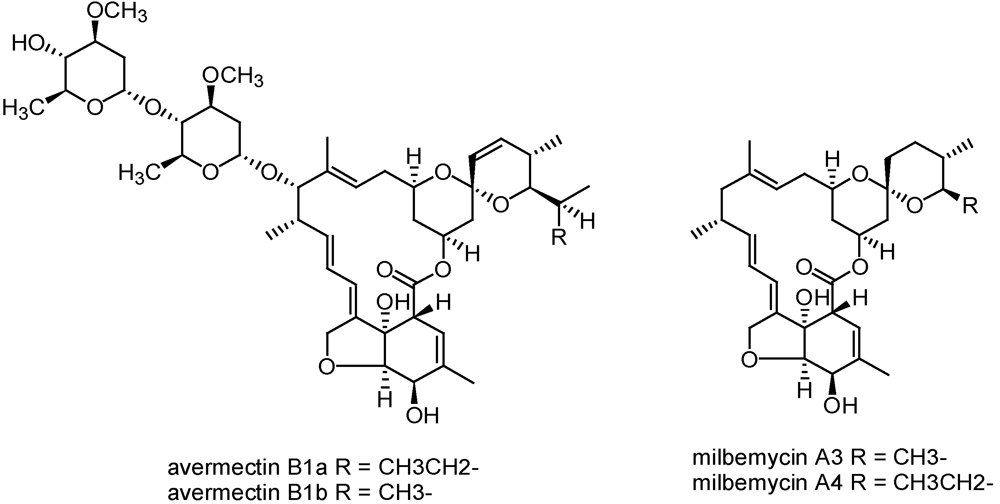
3.2.3. Avermectins and milbemycins
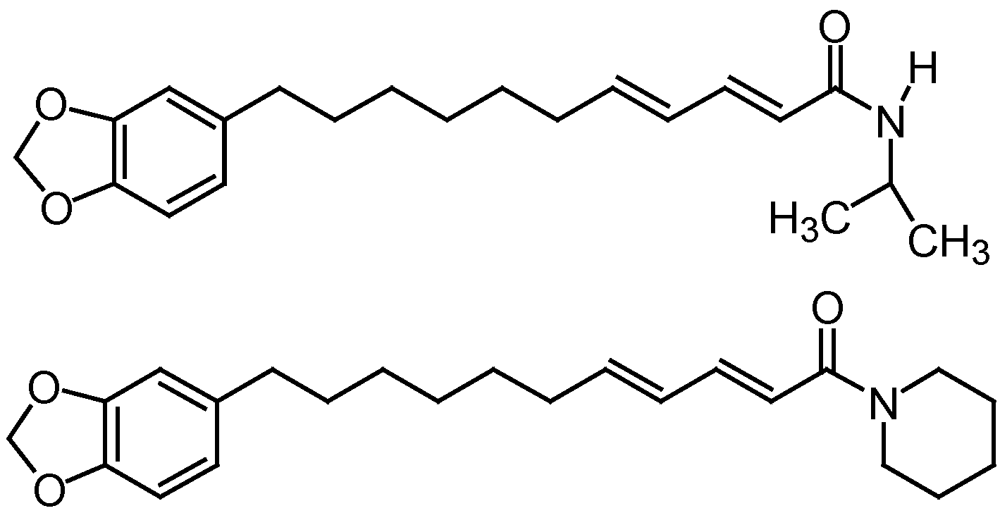
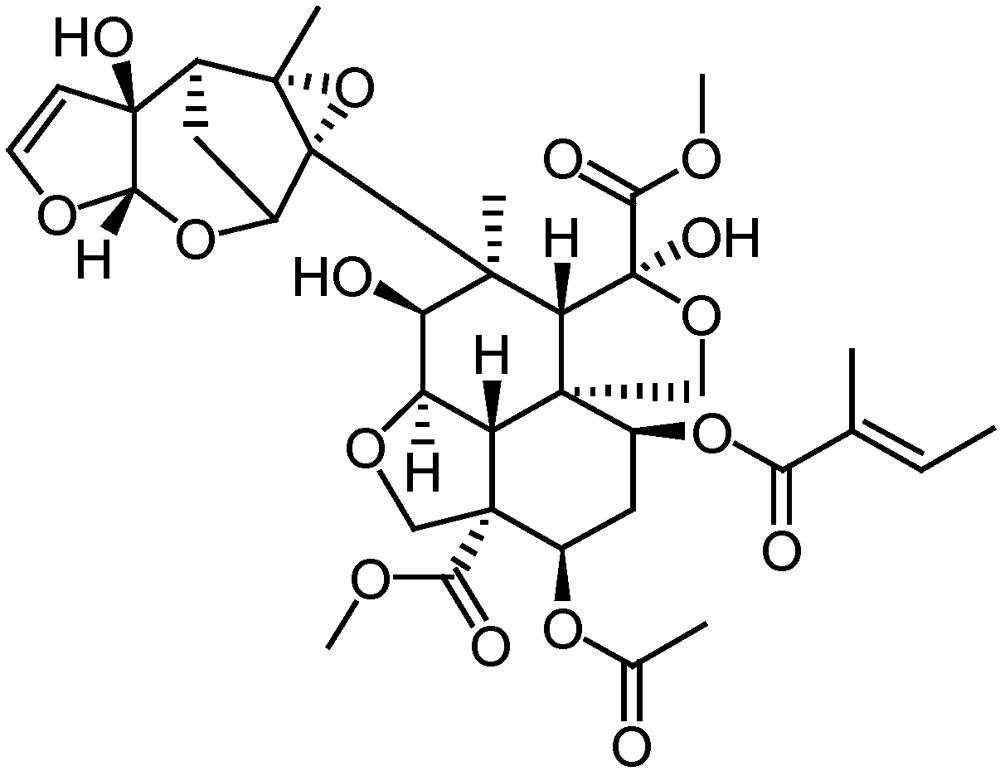
3.2.4. Neem (Azadiracta indica) based products
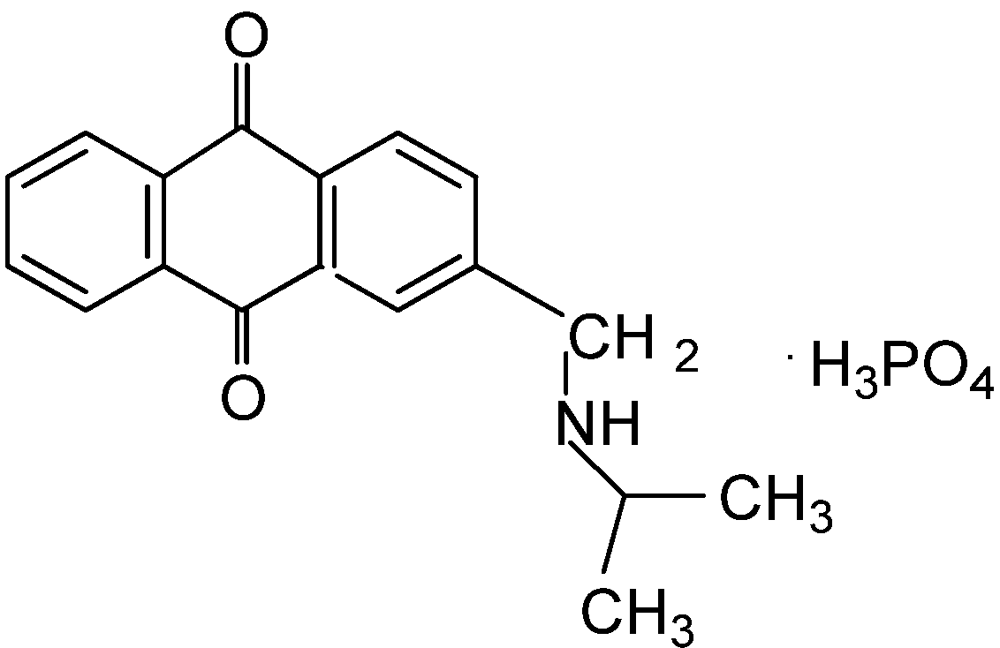
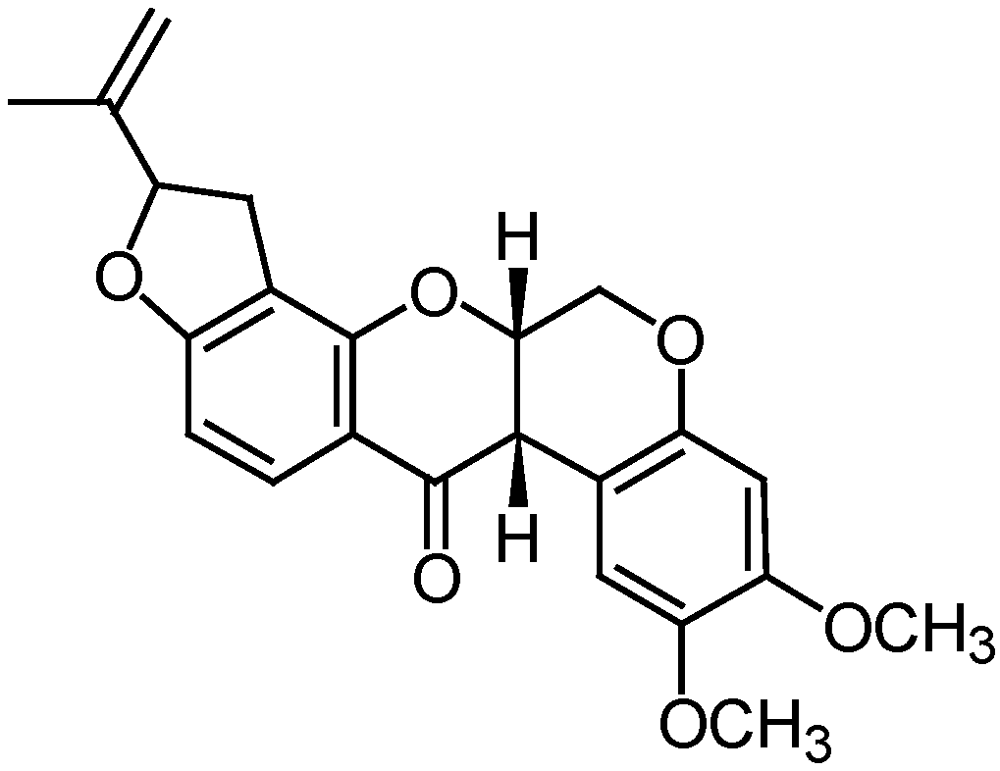
3.2.5. Rotenone
3.2.6. Sabadilla
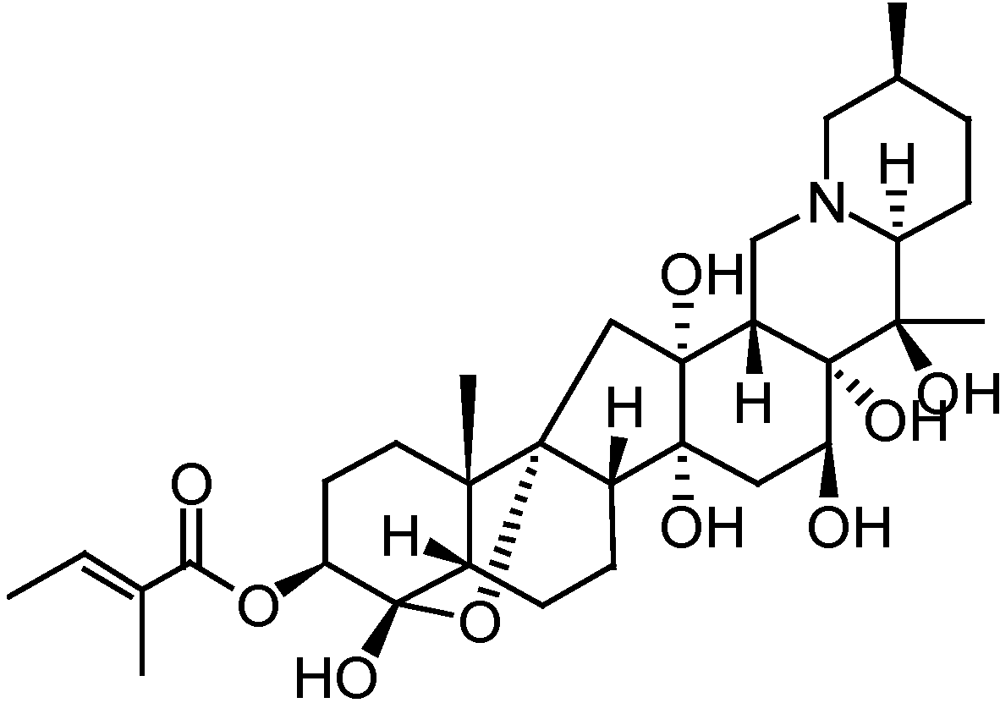
3.2.7. Additional products and preparations
3.3. Insecticides for Urban Use
3.3.1. Termites

3.3.2. Mosquitoes
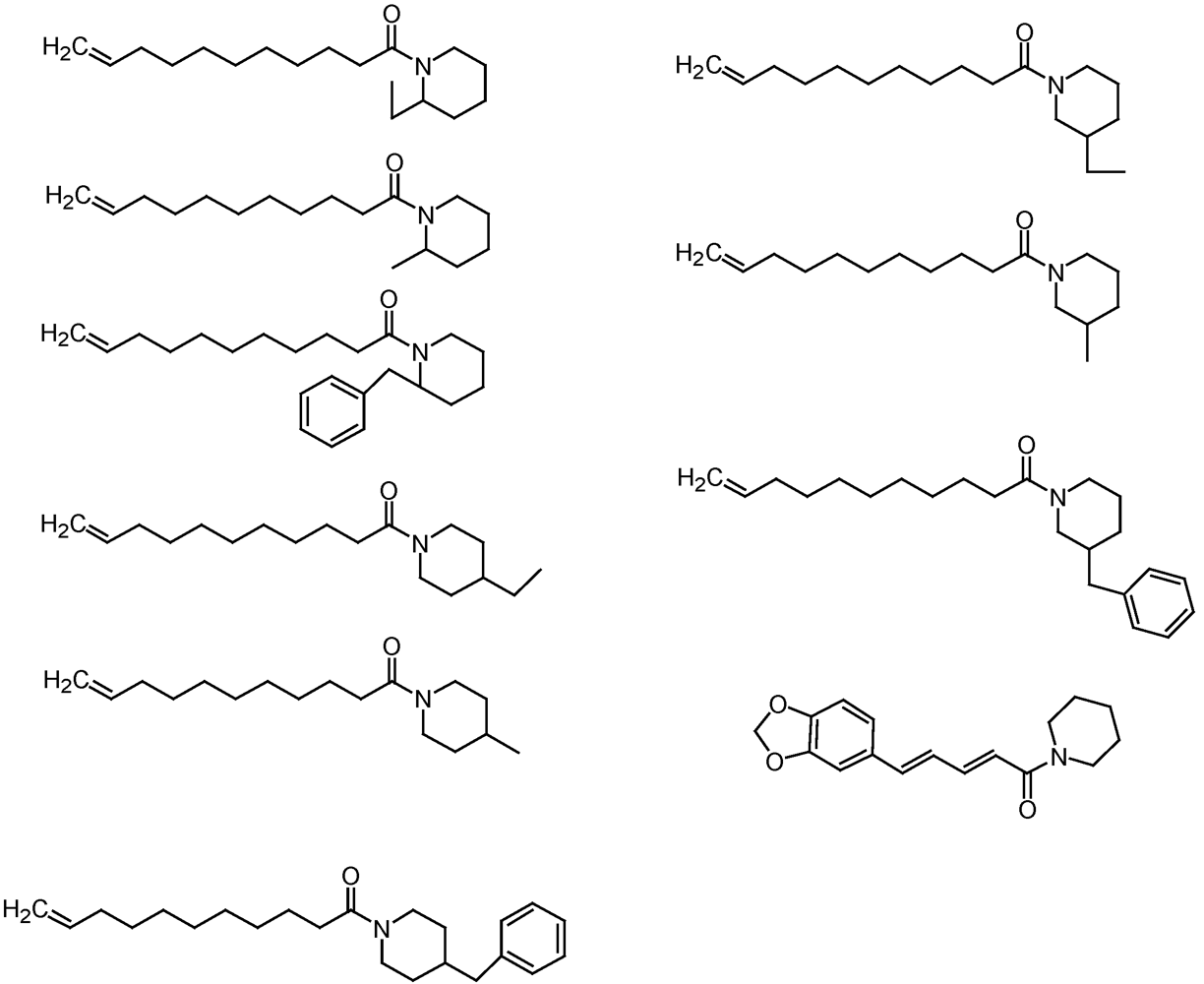
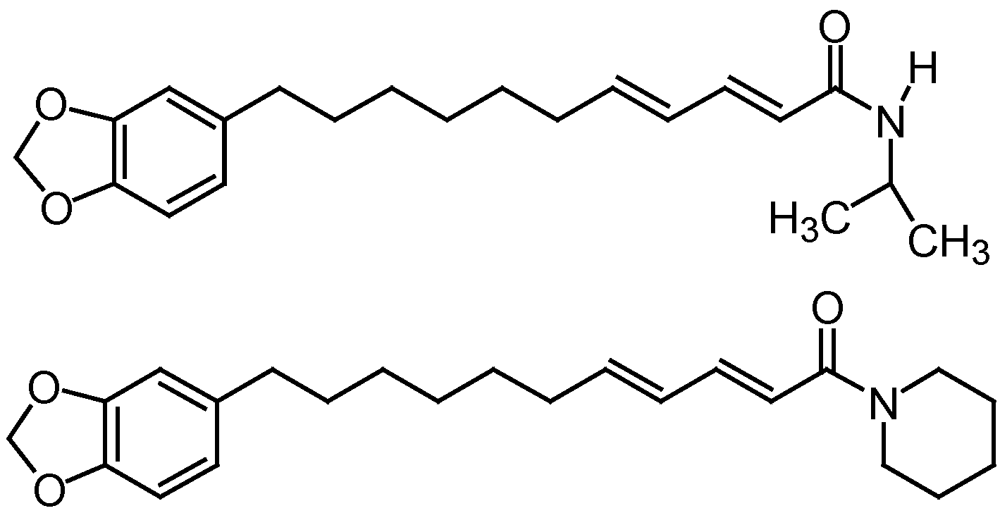
4. Mollusc Management
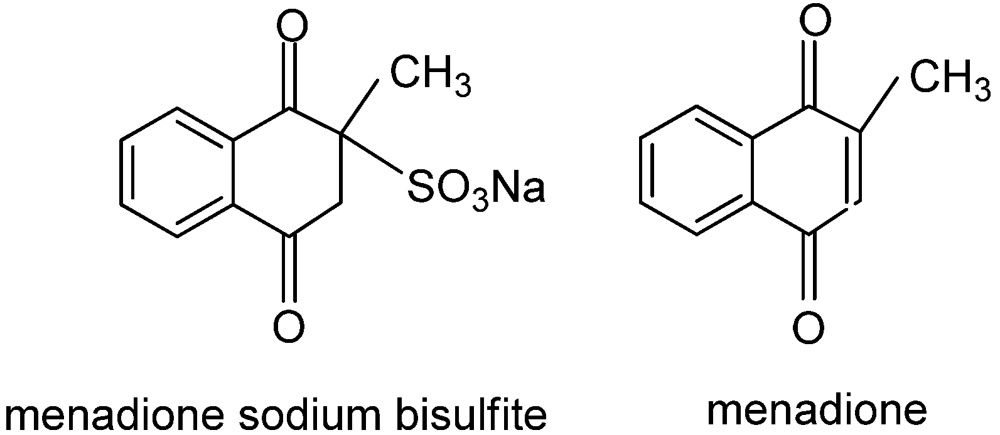
5. Algae Management
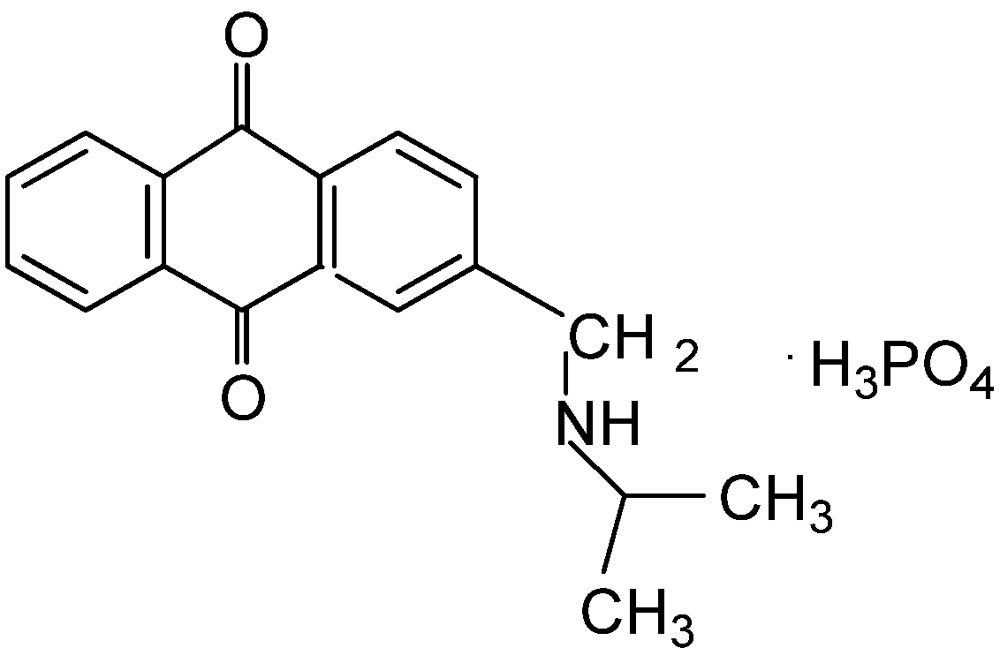
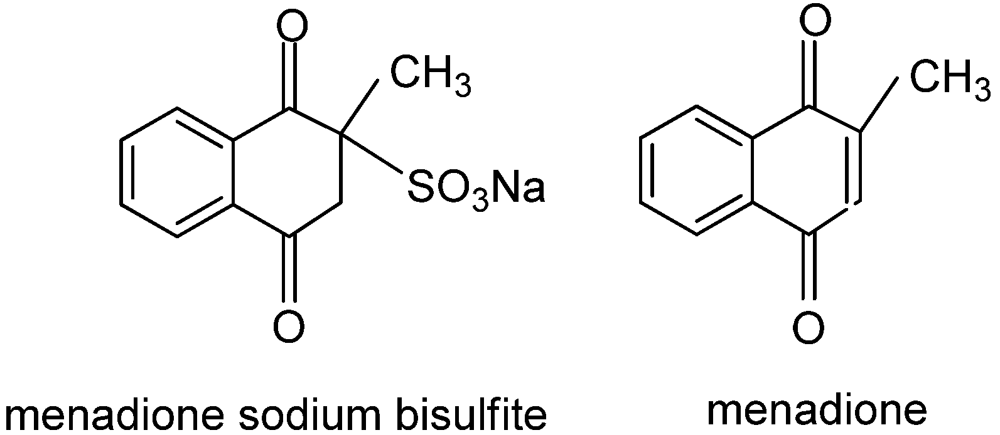
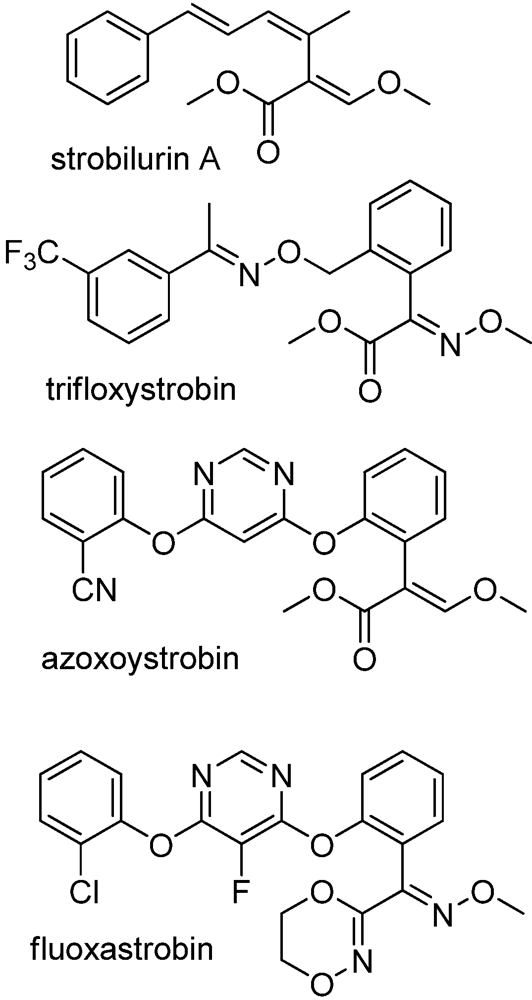
6. Plant Pathogen Management
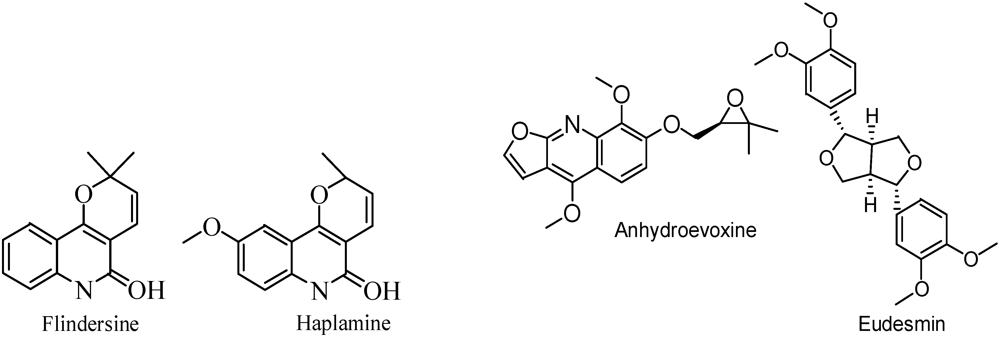
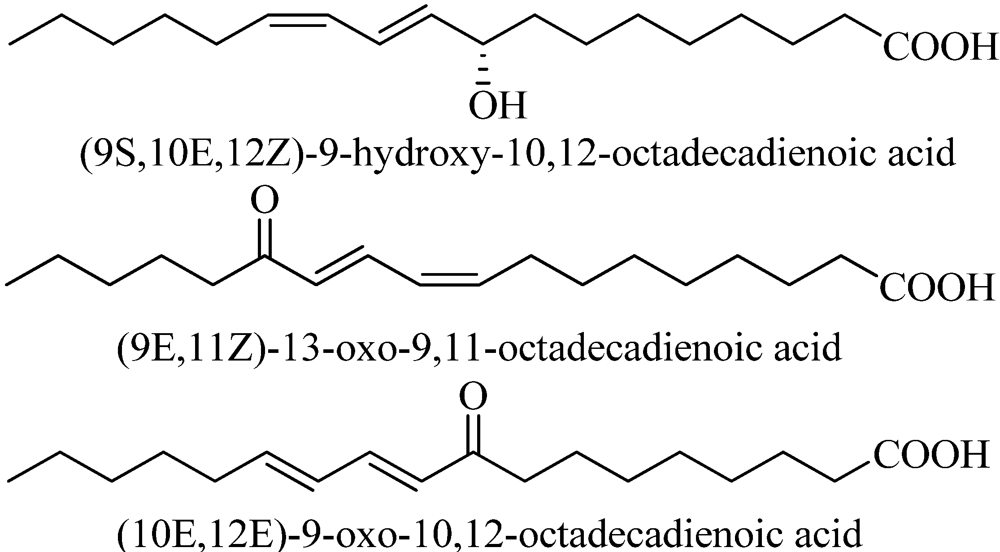
7. Conclusions
References
- Duke, S.O.; Abbas, H.K.; Amagasa, T.; Tanaka, T. Phytotoxins of microbial origin with potential for use as herbicides. In Crop Protection Agents from Nature: Natural Products and Analogues, Critical Reviews on Applied Chemistry; Copping, L.G., Ed.; Society for Chemical Industries: Cambridge, UK, 1996; Volume 35, pp. 82–113. [Google Scholar]
- Copping, L.G.; Duke, S.O. Natural products that have been used commercially as crop protection agents—a review. Pest Manag. Sci. 2007, 63, 524–554. [Google Scholar]
- Dayan, F.E.; Cantrell, C.L.; Duke, S.O. Natural products in crop protection. Bioorg. Med. Chem. 2009, 17, 4022–4034. [Google Scholar]
- Duke, S.O.; Rimando, A.M.; Schrader, K.K.; Cantrell, C.L.; Meepagala, K.M.; Wedge, D.E.; Tabanca, N.; Dayan, F.E. Natural products for pest management. In Selected Topics in the Chemistry of Natural Products; Ikan, R., Ed.; World Scientific: Singapore, Singapore, 2008; pp. 209–251. [Google Scholar]
- Duke, S.O.; Dayan, F.E.; Rimando, A.M.; Schrader, K.K.; Aliotta, G.; Oliva, A.; Romagni, J.G. Chemicals from nature for weed management. Weed Sci. 2002, 50, 138–151. [Google Scholar]
- Lydon, J.; Duke, S.O. Inhibitors of glutamine biosynthesis. In Plant Amino Acids: Biochemistry and Biotechnology; Singh, B.K., Ed.; Marcel Dekker: New York, NY, USA, 1999; pp. 445–464. [Google Scholar]
- Duke, S.O.; Cerdeira, A.L. Transgenic crops for herbicide resistance. In Transgenic Crop Plants: Utilization and Biosafety; Kole, C., Michler, C.H., Abbott, A.G., Hall, T.C., Eds.; Springer-Verlag: Berlin, Germany, 2010; Volume 2, pp. 133–166. [Google Scholar]
- Duke, S.O.; Powles, S.B. Glyphosate: A once in a century herbicide. Pest Manag. Sci. 2008, 64, 319–325. [Google Scholar]
- Duke, S.O. Microbial phytotoxins as herbicides - a perspective. In The Science of Allelopathy; Putnam, A.R., Tang, C.S., Eds.; John Wiley: New York, NY, USA, 1986; pp. 287–304. [Google Scholar]
- Abbas, H.K.; Tanaka, T.; Duke, S.O.; Boyette, C.D. Susceptibility of various crop and weed species to AAL-toxin, a natural herbicide. Weed Technol. 1995, 9, 125–130. [Google Scholar]
- Abbas, H.K.; Tanaka, Y.; Duke, S.O.; Porter, J.K.; Wray, E.M.; Hodges, L.; Sessions, A.E.; Wang, E.; Merrill, A.H.; Riley, R.T. Fumonisin and AAL-toxin-induced disruption of sphingolipid metabolism with accumulation of free sphingoid bases: Involvement in plant disease. Plant Physiol. 1994, 106, 1085–1093. [Google Scholar]
- Abbas, H.K.; Duke, S.O.; Shier, W.T.; Riley, R.T.; Kraus, G.A. The chemistry and biological activities of the natural products AAL-toxin and the fumonisins. In Natural Toxins 2: Structure, Mechanism of Action, and Detection (Advances in Experimental Medicine and Biology); Singh, B.R., Tu, A.T., Eds.; Plenum: New York, NY, USA, 1996; Volume 391, pp. 293–308. [Google Scholar]
- Lee, D.L.; Prisbylla, M.P.; Cromatie, T.H.; Dagarin, D.P.; Howard, S.W.; Provan, W.M.; Ellis, M.K.; Fraser, T.; Mutter, L.C. The discovery and structural requirements of inhibitors of p-hydroxyphenylpyruvate dioxygenase. Weed Sci. 1997, 45, 601–609. [Google Scholar]
- Dayan, F.E.; Duke, S.O.; Sauldubois, A.; Singh, N.; McCurdy, C.; Cantrell, C.L. p-Hydroxyphenylpyruvate dioxygenase is a target site for β-triketones from Leptospermum scoparium. Phytochemistry 2007, 68, 2004–2014. [Google Scholar]
- Nauen, R. Insecticide mode of action: return of the ryanodine receptor. Pest Manag. Sci. 2006, 62, 690–692. [Google Scholar]
- Orr, N.; Shaffner, A.J.; Richey, K.; Crouse, G.D. Novel mode of action of spinosad: Receptor binding studies demonstrating lack of interaction with known insecticidal target sites. Pestic. Biochem. Physiol. 2009, 95, 1–5. [Google Scholar]
- Sparks, T.C.; Crouse, G.D.; Durst, G. Natural products as insecticides: the biology, biochemistry and quantitative structure–activity relationships of spinosyns and spinosoids. Pest Manag. Sci. 2001, 57, 896–905. [Google Scholar]
- Snyder, D.E.; Meyer, J.; Zimmermann, A.G.; Qiao, M.; Gissendanner, S.J.; Cruthers, L.R.; Slone, R.L; Young, D.R. Preliminary studies on the effectiveness of the novel pulicide, spinosad, for the treatment and control of fleas on dogs. Vet. Parasitol. 2007, 150, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. The role of botanical insecticides, deterrents and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar]
- Casida, J.E.; Quistad, G.B. Pyrethrums, a benefit to human welfare. In Pyrethrum Flowers: Production, Chemistry, Toxicology and Uses; Casida, J.E., Quistad, G.B., Eds.; Oxford University: New York, NY, USA, 1995; pp. 345–350. [Google Scholar]
- Chang, C.P.; Plapp, F.W., Jr. DDT and Synthetic pyrethroids: Mode of action, selectivity, and mechanism of synergism in the tobacco budworm (Lepidoptera: Noctuidae) and a predator, Chrysopa carnea Stephens (Neuroptera: Chrysopidae). J. Econ. Entomol. 1983, 76, 1206–1210. [Google Scholar]
- Clark, J.M. Insecticides as Tools in probing vital receptors and enzymes in excitable membranes. Pestic. Biochem. Physiol. 1997, 57, 235–254. [Google Scholar]
- Campbell, W.C. Ivermectin and Abamectin; Springer-Verlag: New York, NY, USA, 1989; p. 361. [Google Scholar]
- Kornis, G.I. Avermectins and milbemycins. In Agrochemicals from Natural Products; Godfrey, C.R.A., Ed.; Marcel Dekker: New York, NY, USA, 1995; pp. 215–255. [Google Scholar]
- Meister, R.T. Farm Chemicals Handbook '92; Meister: Willoughby, OH, USA, 1992. [Google Scholar]
- Hayes, W.J.; Laws, E.R. Handbook of Pesticide Toxicology, Classes of Pesticides; Academic: New York, NY, USA, 1990; Volume 3. [Google Scholar]
- Arena, J.P.; Liu, K.K.; Paress, P.S.; Frazier, E.G.; Cully, D.F.; Mrozik, H.; Schaeffer, J. The mechanism of action of avermectins in Caenorhabditis elegans: Correlation between activation of glutamate-sensitive chloride current, membrane binding, and biological Activity. J. Parasitol. 1995, 81, 286–294. [Google Scholar]
- Mordue, A.J.; Blackwell, A. Azadirachtin: an update. J. Insect Phys. 1993, 39, 903–924. [Google Scholar]
- Fang, N.; Casida, J.E. Cube resin insecticide: Identification and biological activity of 29 rotenoid constituents. J. Agric. Food Chem. 1999, 47, 2130–2136. [Google Scholar]
- Barton, D.H.R.; Jeger, O.; Prelog, V.; Woodward, R.B. The constitutions of cevine and some related alkaloids. Experientia 1954, 10, 81–90. [Google Scholar]
- Schmelz, I. Nicotine and other tobacco alkaloids. In Naturally Occurring Insecticides; Jacobson, M., Crosby, D.G., Eds.; Marcel Dekker: New York, NY, USA, 1971; pp. 99–136. [Google Scholar]
- Liu, B.; Chen, C.; Wu, D.; Su, Q. Enantiomeric analysis of anatabine, nornicotine and anabasine in commercial tobacco by multi-dimensional gas chromatography and mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2008, 865, 13–17. [Google Scholar]
- Lax, A.R.; Osbrink, W.L.A. Research on targeted management of the Formosan subterranean termite Coptotermes formosanus Shiraki Isoptera: Rhinotermitidae. Pest Manag. Sci. 2003, 59, 788–800. [Google Scholar]
- Chiasson, H.; Belanger, A.; Bostanian, N.; Vincent, C.; Poliquin, A. Acaricidal properties of Artemisia absinthium and Tanacetum vulgare (Asteraceae) essential oils obtained by three methods of extraction. J. Econ. Entomol. 2001, 94, 167–171. [Google Scholar]
- Gonzalez-Coloma, A.; Guadano, A.; Tonn, C.E.; Sosa, M.E. Antifeedant/insecticidal terpenes from Asteraceae and Labiatae species native to Argentinean semi-arid lands. Z. Naturforsch., C, J. Biosci. 2005, 60, 855–861. [Google Scholar] [PubMed]
- Gupta, S.C.; Misra, A.K. Management of okra shoot of fruit borer, Earias vittella fabr. through bio-rational insecticides. Pestic. Res. J. 2006, 18, 33–34. [Google Scholar]
- Prasad, S.M.; Prasad, N.; Barnwal, M.K. Evaluation of insecticides against aphid vectors of virus diseases of French Bean. J. Plant Protect. Environ. 2006, 3, 87–90. [Google Scholar]
- Sharma, D.C.; Badiyala, A.; Choudhary, A. Bioefficacy and persistent toxicity of biopesticides and insecticides against potato tuber moth, Phthorimaea opercullela Zell. on spring potato. Pestic. Res. J. 2006, 18, 43–46. [Google Scholar]
- Hadacek, F.; Mueller, C.; Werner, A.; Greger, H.; Proksch, P. Analysis, isolation and insecticidal activity of linear furanocoumarins and other coumarin derivatives from Peucedanum (Apiaceae: Apioideae). J. Chem. Ecol. 1994, 20, 2035–2054. [Google Scholar]
- Kang, S.; Kim, H.; Lee, W.; Ahn, Y. Toxicity of bisabolangelone from Ostericum koreanum roots to Dermatophagoides farinae and Dermatophagoides pteronyssinus (Acari: Pyroglyphidae). J. Agric. Food Chem. 2006, 54, 3547–3550. [Google Scholar]
- Meepagala, K.; Osbrink, W.; Sturtz, G.; Lax, A. Plant-derived natural products exhibiting activity against Formosan subterranean termites (Coptotermes formosanus). Pest Manag. Sci. 2006, 62, 565–570. [Google Scholar]
- Meepagala, K.M.; Kuhajek, J.M.; Sturtz, G.; Wedge, D.E. Vulgarone B, the antifungal constituent in the steam-distilled fraction of Artemisia douglasiana. J. Chem. Ecol. 2003, 29, 1771–1780. [Google Scholar]
- Zhu, B.C.R.; Henderson, G.; Chen, F.; Maistrello, L.; Laine, R.A. Nootkatone is a repellent for Formosan subterranean termite (Coptotermes formosanus). J. Chem. Ecol. 2001, 27, 523–531. [Google Scholar]
- Pridgeon, J.W.; Becnel, J.J.; Wang, Z.; Katritzky, A.R.; Meepagala, K.M.; Clark, G.G. Structure-activity relationships of 33 piperidines as toxicants against female adults of Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2007, 44, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Meepagala, K.M. 2008; USDA, ARS NPURU, Oxford, MS, USA. Unpublished work.
- Goll, P.H.; Lemma, A.; Duncan, J.; Mazengia, B. Control of schistosomiasis in Adwa, Ethiopia, using the plant molluscicide endod (Phytolacca dodecandra). Tropenmedizin und Parasitologie 1983, 34, 177–183. [Google Scholar]
- Marston, A.; Hostetmann, K. Plant molluscicides. Phytochemistry 1985, 24, 639–652. [Google Scholar]
- Mitchell, A.J. Update and impact of a trematode that infects cultured channel catfish. Catfish J. 2001, 16, 17–27. [Google Scholar]
- Meepagala, K.M.; Sturtz, G.; Mischke, C.C.; Wise, D.; Duke, S.O. Molluscicidal activity of vulgarone B against ram's horn snail (Planorbella trivolvis). Pest Manag. Sci. 2004, 60, 479–482. [Google Scholar]
- Meepagala, K.M.; Sturtz, G.; Wise, D.; Wedge, D.E. Molluscicidal and antifungal activity of Erigeron speciosus steam distillate. Pest Manag. Sci. 2002, 58, 1043–1047. [Google Scholar]
- Meepagala, K.M. 2005; USDA, ARS NPURU, Oxford, MS, USA. Unpublished work.
- Joshi, R.C.; Meepagala, K.M.; Sturtz, G.; Cagauan, A.G.; Mendoza, C.O.; Dayan, F.E.; Duke, S.O. Molluscicidal activity of vulgarone B from Artemisia douglasiana (Besser) against the invasive, alien, mollusc pest, Pomacea canaliculata (Lamarck). Int. J. Pest Manag. 2005, 51, 175–180. [Google Scholar]
- Meepagala, K.M. 2004; USDA, ARS NPURU, Oxford, MS, USA. Unpublished work.
- Tucker, C.S. 2005; Mississippi State University, Stoneville, MS, USA. Unpublished work.
- Schrader, K.K.; de Regt, M.Q.; Tucker, C.S.; Duke, S.O. A rapid bioassay for selective algicides. Weed Technol. 1997, 11, 767–774. [Google Scholar]
- Schrader, K.K.; Duke, S.O.; Kingsbury, S.K.; Tucker, C.S.; Duke, M.V.; Dionigi, C.P.; Millie, D.F.; Zimba, P.V. Evaluation of ferulic acid for controlling the musty-odor cyanobacterium, Oscillatoria perornata, in aquaculture ponds. J. Appl. Aquacult. 2000, 10, 1–16. [Google Scholar]
- Schrader, K.K.; de Regt, M.Q.; Tidwell, P.R.; Tucker, C.S.; Duke, S.O. Selective growth inhibition of the musty-odor producing cyanobacterium Oscillatoria cf. chalybea by natural compounds. Bull. Environ. Contam. Toxicol. 1998, 60, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Schrader, K.K.; Nanayakkara, N.P.D. Selective algaecides for control of cyanochloronta. U.S. Patent 6,949,250,27, 27 September 2005. [Google Scholar]
- Schrader, K.K.; Nanayakkara, N.P.D.; Tucker, C.S.; Rimando, A.M.; Ganzera, M.; Schaneberg, B.T. Novel derivatives of 9,10-anthraquinone are selective algicides against the musty-odor cyanobacterium Oscillatoria perornata. Appl. Environ. Microbiol. 2003, 69, 5319–5327. [Google Scholar]
- Schrader, K.K.; Foran, C.M.; Holmes, B.D.; Schlenk, D.K.; Nanayakkara, N.P.D.; Schaneberg, B.T. Toxicological evaluation of two anthraquinone-based cyanobactericides to channel catfish. N. Am. J. Aquacult. 2004, 66, 119–124. [Google Scholar]
- Cutler, H.G.; Cutler, S.J.; Wright, D.; Dawson, R. Methods of controlling zoological and aquatic plant growth. U.S. Patent 6,340,468,22, 22 January 2002. [Google Scholar]
- Schrader, K.K.; Rimando, A.M.; Tucker, C.S.; Glinski, J.; Cutler, S.J.; Cutler, H.G. Evaluation of the natural product SeaKleen® for controlling the musty-odor-producing cyanobacterium Oscillatoria perornata in catfish ponds. N. Am. J. Aquacult. 2004, 66, 20–28. [Google Scholar]
- Wedge, D.E.; Duke, S.O. Finding new fungicides from natural sources. Am. Chem. Soc. Symp. Ser. 2006, 927, 152–167. [Google Scholar]
- Engelmeier, D.; Hadacek, F. Antifungal natural products: Assays and applications. In Advances in Phytomedicine, Naturally Occurring Bioactive Compounds; Ashok, K., Ed.; Elsevier: Amsterdam, The Netherlands, 2006; Volume 3, pp. 423–467. [Google Scholar]
- Kim, B.S.; Hwang, B.K. Microbial fungicides in the control of plant diseases. J. Phytopathol. 2007, 155, 641–653. [Google Scholar]
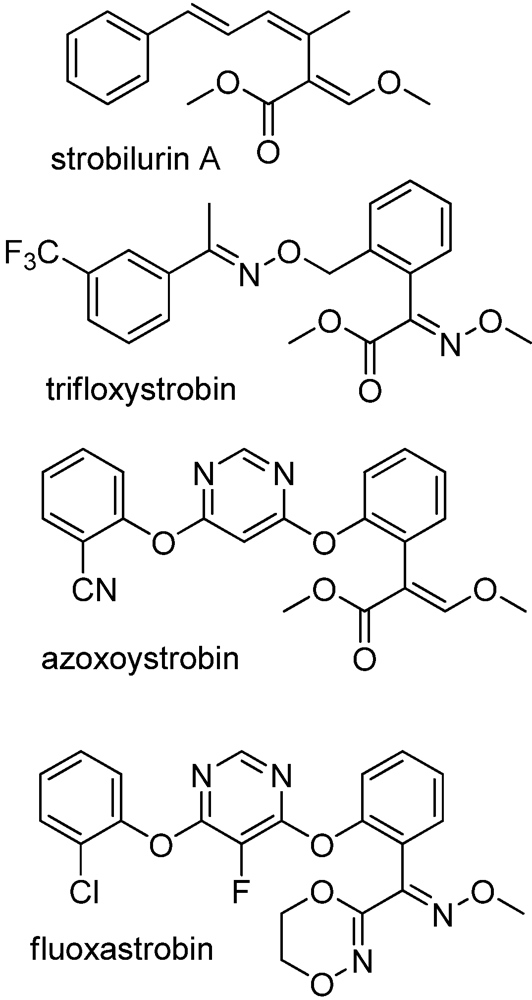
- Sauter, H.; Ammermann, E.; Roehl, F. Strobilurins- From natural products to a new class of fungicides. In Crop Protection Agents from Nature: Natural Products and Analogues; Copping, L.G., Ed.; Royal Soc. Chem.: Cambridge, UK, 1996; pp. 50–81. [Google Scholar]
- Bang, K.-H.; Lee, D.-W.; Park, H.-M.; Rhee, Y.-H. Inhibition of fungal cell wall synthesizing enzymes by trans-cinnamaldehyde. Biosci. Biotech. Biochem. 2000, 64, 1061–1063. [Google Scholar]
- Kang, T.H.; Hwang, E.I.; Yun, B.S.; Park, K.D.; Kwon, B.M.; Shin, C.S.; Kim, S.U. Inhibition of chitin synthases and antifungal activities of 2’-benzolyoxycinnamaldehyde from Pleuropterus ciliinervis and its derivatives. Biol. Pharm. Bull. 2007, 30, 598–602. [Google Scholar]
- Wedge, D.E.; Nagle, D.G. Preparation of sampangine and its analogs as fungicides. U.S. Pat. Appl. Publ. CODEN: USXXCO US 2004192721 A1 20040930 CAN 141:273006 AN 2004:803936, 2004. [Google Scholar]
- Li, X.-C.; Jacob, M.B.; Wedge, D.E. Preparation of cyclopentenedione antifungal compounds. U.S. Pat. Appl. Publ. CODEN: USXXCO US 2005215648 A1 20050929, 2005. [Google Scholar]
- Cantrell, C.L.; Schrader, K.K.; Mamonov, L.K.; Sitpaeva, G.T.; Kustova, T.S.; Dunbar, C.; Wedge, D.E. Isolation and identification of antifungal and antialgal alkaloids from Haplophyllum sieversii. J. Agric. Food Chem. 2005, 53, 7741–7748. [Google Scholar]
- Cantrell, C.L.; Case, B.P.; Mena, E.E.; Kniffin, T.M.; Duke, S.O.; Wedge, D.E. Isolation and identification of antifungal fatty acids from the basidiomycete Gomphus floccosus. J. Agric. Food Chem 2008, 56, 5062–5068. [Google Scholar]
- Fokialakis, N.; Cantrell, C.L.; Duke, S.O.; Skaltsounis, A.L.; Wedge, D.E. Antifungal activity of thiophenes from Echinops ritro. J. Agric. Food Chem. 2006, 54, 1651–1655. [Google Scholar]
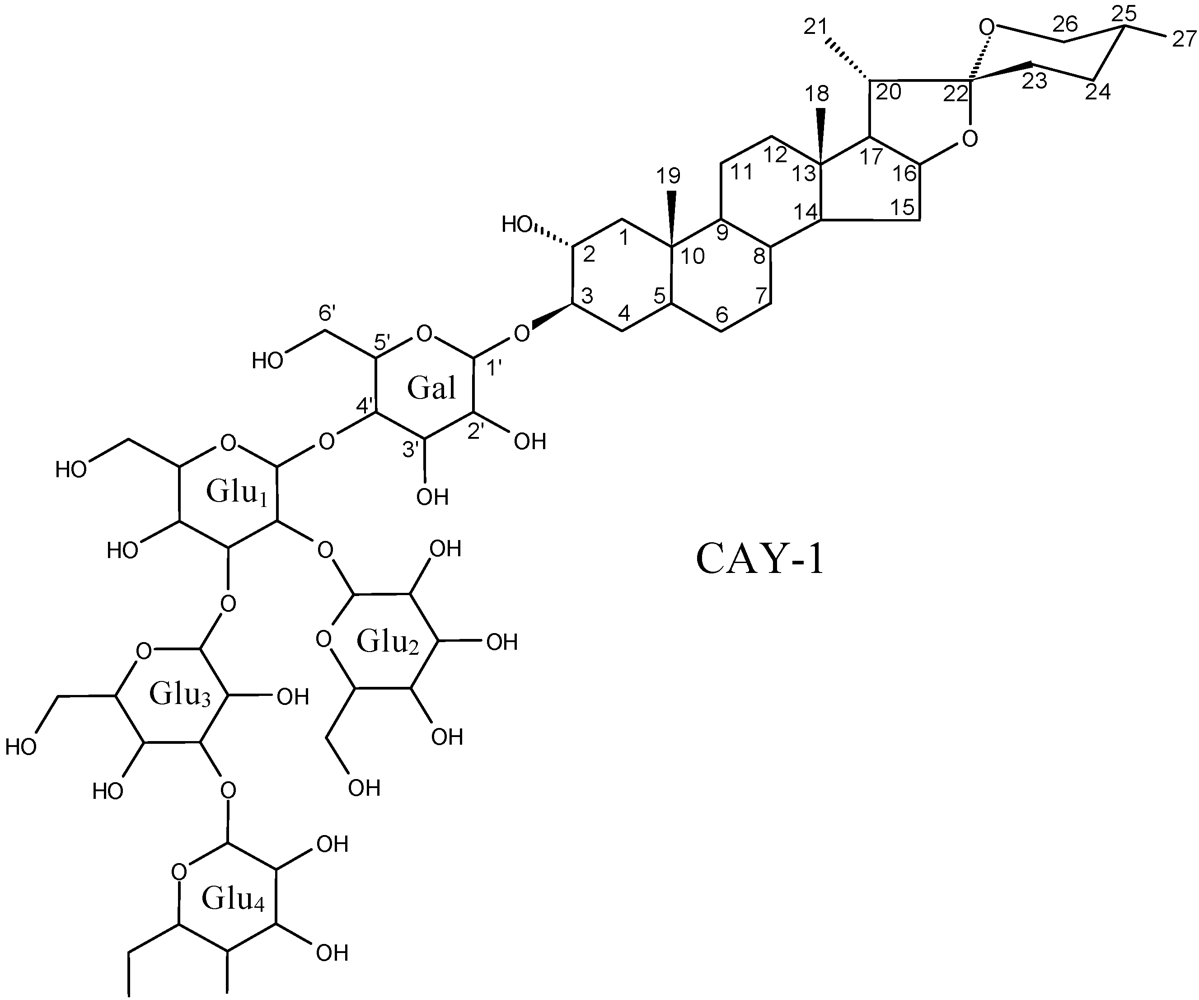
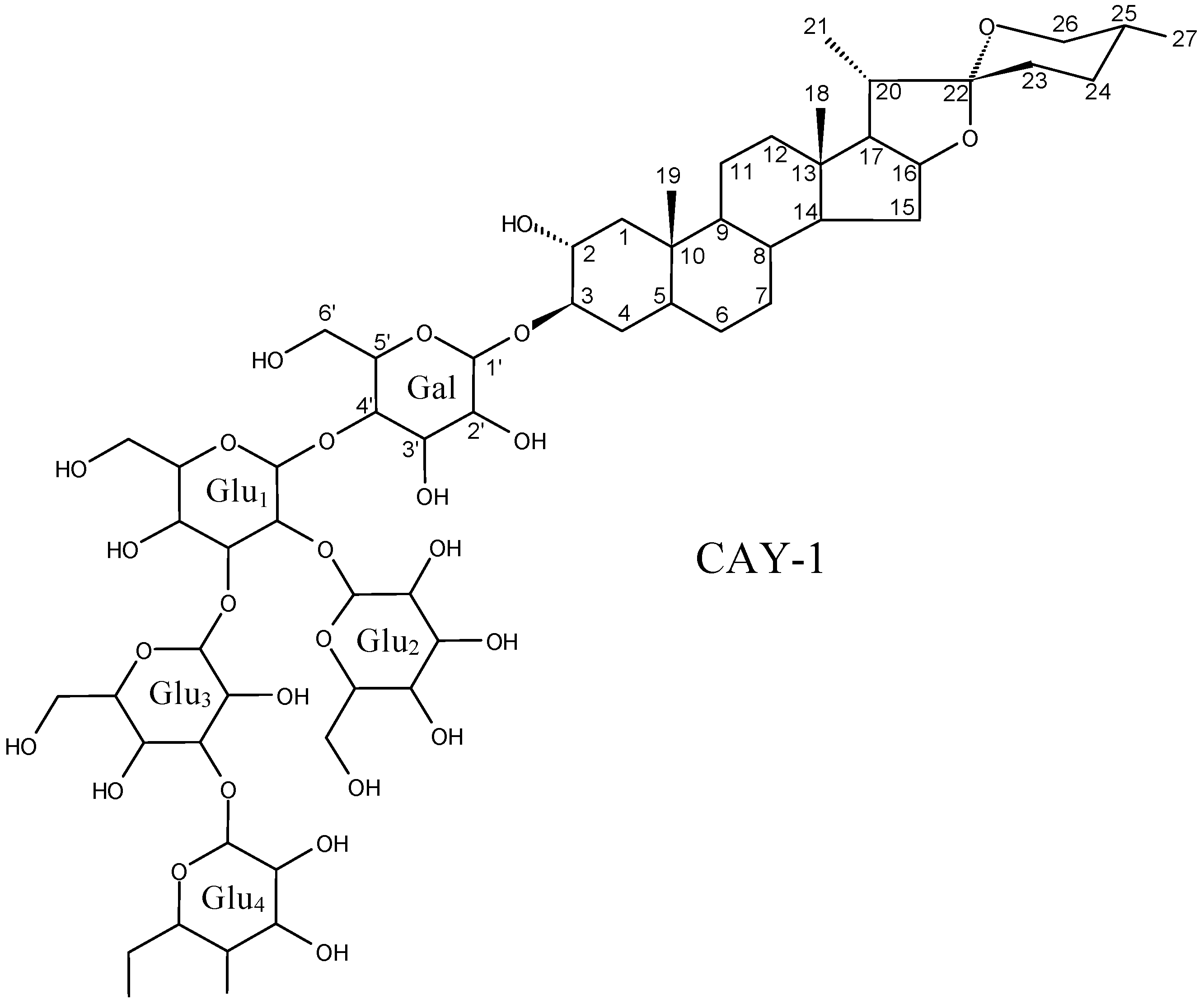
- Abril, M.; Curry, K.J.; Smith, B.J.; DeLucca, A.J.; Boue, S.; Wedge, D.E. Greenhouse and field evaluation of the natural saponin CAY-1, for control of several strawberry diseases. Int. J. Fruit Sci. 2008, 9, 211–220. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Duke, S.O.; Cantrell, C.L.; Meepagala, K.M.; Wedge, D.E.; Tabanca, N.; Schrader, K.K. Natural Toxins for Use in Pest Management. Toxins 2010, 2, 1943-1962. https://doi.org/10.3390/toxins2081943
Duke SO, Cantrell CL, Meepagala KM, Wedge DE, Tabanca N, Schrader KK. Natural Toxins for Use in Pest Management. Toxins. 2010; 2(8):1943-1962. https://doi.org/10.3390/toxins2081943
Chicago/Turabian StyleDuke, Stephen O., Charles L. Cantrell, Kumudini M. Meepagala, David E. Wedge, Nurhayat Tabanca, and Kevin K. Schrader. 2010. "Natural Toxins for Use in Pest Management" Toxins 2, no. 8: 1943-1962. https://doi.org/10.3390/toxins2081943