Role of the Osmotic Stress Regulatory Pathway in Morphogenesis and Secondary Metabolism in Filamentous Fungi

{kind=link}
Abstract
:1. Introduction
2. Osmoadaptation Mechanisms in Yeast
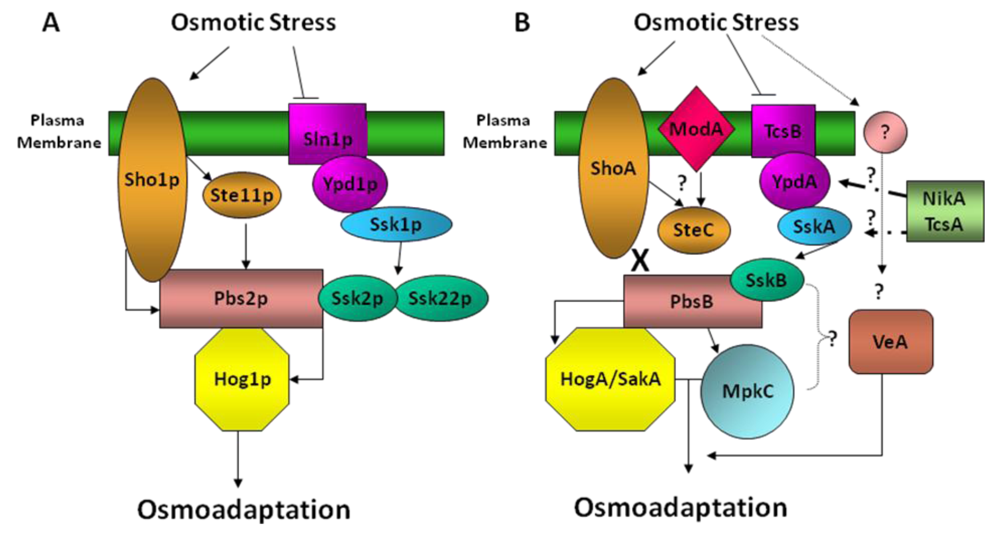
3. Hog Pathway in Aspergillus nidulans
4. Examples of the Osmoadaptation Signaling Pathway in Other Aspergilli
5. Role of the Osmotic Stress-response Pathway on Fungal Development and Secondary Metabolism
6. Conclusions
Acknowledgements
References
- Hohmann, S.; Krantz, M.; Nordlander, B. Yeast osmoregulation. Methods Enzymol. 2007, 428, 29–45. [Google Scholar] [PubMed]
- Hohmann, S. Osmotic stress signaling and osmoadaptation in yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372. [Google Scholar] [PubMed]
- Todd, R.B.; Davis, M.A.; Hynes, M.J. Genetic manipulation of Aspergillus nidulans: Meiotic progeny for genetic analysis and strain construction. Nat. Protoc. 2007, 2, 811–821. [Google Scholar] [PubMed]
- Bennett, J.W. Aspergillus: A primer for the novice. Med. Mycol. 2009, 47, S5–S12. [Google Scholar] [CrossRef] [PubMed]
- Cary, J.W.; Linz, J.E.; Bhatnagar, D. Aflatoxins: Biological significance and regulation of biosynthesis. In Microbial Foodborne Diseases: Mechanisms of Pathogenesis and Toxin Synthesis; Cary, J.W., Linz, J.E., Bhatnagar, D., Eds.; Technomic Publishing Co.: Lancaster, PA, USA, 2000; pp. 317–361. [Google Scholar]
- Roukas, T. Citric and gluconic acid production from fig by Aspergillus niger using solid-state fermentation. J. Ind. Microbiol. Biotechnol. 2000, 25, 298–304. [Google Scholar] [PubMed]
- Machida, M.; Yamada, O.; Gomi, K. Genomics of Aspergillus oryzae: Learning from the history of Koji mold and exploration of its future. DNA Res. 2008, 15, 173–183. [Google Scholar] [PubMed]
- Latge, J.P. Aspergillus fumigatus and aspergillosis. Clin. Microbiol. Rev. 1999, 12, 310–350. [Google Scholar] [PubMed]
- Lillehoj, E.B.; Wall, J.H.; Bowers, E.J. Preharvest aflatoxin contamination: Effect of moisture and substrate variation in developing cottonseed and corn kernels. Appl. Environ. Microbiol. 1987, 53, 584–586. [Google Scholar] [PubMed]
- Fiedurek, J. Effect of osmotic stress on glucose oxidase production and secretion by Aspergillus niger. J. Basic Microbiol. 1998, 38, 107–112. [Google Scholar] [PubMed]
- Kobayashi, A.; Sano, M.; Oda, K.; Hisada, H.; Hata, Y.; Ohashi, S. The glucoamylase-encoding gene (glaB) is expressed in solid-state culture with a low water content. Biosci. Biotechnol. Biochem. 2007, 71, 1797–1799. [Google Scholar] [PubMed]
- Miskei, M.; Karanyi, Z.; Pocsi, I. Annotation of stress-response proteins in the aspergilli. Fungal Genet. Biol. 2009, 46, S105–S120. [Google Scholar] [PubMed]
- Han, K.H.; Prade, R.A. Osmotic stress-coupled maintenance of polar growth in Aspergillus nidulans. Mol. Microbiol. 2002, 43, 1065–1078. [Google Scholar] [PubMed]
- Furukawa, K.; Hoshi, Y.; Maeda, T.; Nakajima, T.; Abe, K. Aspergillus nidulans HOG pathway is activated only by two-component signalling pathway in response to osmotic stress. Mol. Microbiol. 2005, 56, 1246–1261. [Google Scholar] [PubMed]
- Posas, F.; Wurgler-Murphy, S.M.; Maeda, T.; Witten, E.A.; Thai, T.C.; Saito, H. Yeast HOG1 MAP kinase cascade is regulated by a multistep phosphorelay mechanism in the SLN1-YPD1-SSK1 "two-component" osmosensor. Cell 1996, 86, 865–875. [Google Scholar] [PubMed]
- Li, S.; Ault, A.; Malone, C.L.; Raitt, D.; Dean, S.; Johnston, L.H.; Deschenes, R.J.; Fassler, J.S. The yeast histidine protein kinase, Sln1p, mediates phosphotransfer to two response regulators, Ssk1p and Skn7p. EMBO J. 1998, 17, 6952–6962. [Google Scholar] [PubMed]
- Maeda, T.; Wurgler-Murphy, S.M.; Saito, H. A two-component system that regulates an osmosensing MAP kinase cascade in yeast. Nature 1994, 369, 242–245. [Google Scholar] [PubMed]
- Posas, F.; Saito, H. Activation of the yeast SSK2 MAP kinase kinase kinase by the SSK1 two-component response regulator. EMBO J. 1998, 17, 1385–1394. [Google Scholar] [PubMed]
- Ferrigno, P.; Posas, F.; Koepp, D.; Saito, H.; Silver, P.A. Regulated nucleo/cytoplasmic exchange of HOG1 MAPK requires the importin beta homologs NMD5 and XPO1. EMBO J. 1998, 17, 5606–5614. [Google Scholar] [PubMed]
- Albertyn, J.; Hohmann, S.; Thevelein, J.M.; Prior, B.A. GPD1, which encodes glycerol-3-phosphate dehydrogenase, is essential for growth under osmotic stress in Saccharomyces cerevisiae, and its expression is regulated by the high-osmolarity glycerol response pathway. Mol. Cell Biol. 1994, 14, 4135–4144. [Google Scholar] [PubMed]
- Norbeck, J.; Pahlman, A.K.; Akhtar, N.; Blomberg, A.; Adler, L. Purification and characterization of two isoenzymes of DL-glycerol-3-phosphatase from Saccharomyces cerevisiae. Identification of the corresponding GPP1 and GPP2 genes and evidence for osmotic regulation of Gpp2p expression by the osmosensing mitogen-activated protein kinase signal transduction pathway. J. Biol. Chem. 1996, 271, 13875–13881. [Google Scholar] [PubMed]
- Mattison, C.P.; Ota, I.M. Two protein tyrosine phosphatases, Ptp2 and Ptp3, modulate the subcellular localization of the Hog1 MAP kinase in yeast. Gene. Develop. 2000, 14, 1229–1235. [Google Scholar]
- Warmka, J.; Hanneman, J.; Lee, J.; Amin, D.; Ota, I. Ptc1, a type 2C Ser/Thr phosphatase, inactivates the HOG pathway by dephosphorylating the mitogen-activated protein kinase Hog1. Mol. Cell Biol. 2001, 21, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Young, C.; Mapes, J.; Hanneman, J.; Al-Zarban, S.; Ota, I. Role of Ptc2 type 2C Ser/Thr phosphatase in yeast high-osmolarity glycerol pathway inactivation. Eukaryot Cell 2002, 1, 1032–1040. [Google Scholar] [PubMed]
- Mapes, J.; Ota, I.M. Nbp2 targets the Ptc1-type 2C Ser/Thr phosphatase to the HOG MAPK pathway. EMBO J. 2004, 23, 302–311. [Google Scholar] [PubMed]
- Saito, H.; Tatebayashi, K. Regulation of the osmoregulatory HOG MAPK cascade in yeast. J. Biochem. 2004, 136, 267–272. [Google Scholar] [PubMed]
- Tatebayashi, K.; Yamamoto, K.; Tanaka, K.; Tomida, T.; Maruoka, T.; Kasukawa, E.; Saito, H. Adaptor functions of Cdc42, Ste50, and Sho1 in the yeast osmoregulatory HOG MAPK pathway. EMBO J. 2006, 25, 3033–3044. [Google Scholar] [CrossRef] [PubMed]
- Proft, M.; Pascual-Ahuir, A.; de Nadal, E.; Arino, J.; Serrano, R.; Posas, F. Regulation of the Sko1 transcriptional repressor by the Hog1 MAP kinase in response to osmotic stress. EMBO J. 2001, 20, 1123–1133. [Google Scholar] [PubMed]
- Mas, G.; de Nadal, E.; Dechant, R.; Rodriguez de la Concepcion, M.L.; Logie, C.; Jimeno-Gonzalez, S.; Chavez, S.; Ammerer, G.; Posas, F. Recruitment of a chromatin remodelling complex by the Hog1 MAP kinase to stress genes. EMBO J. 2009, 28, 326–336. [Google Scholar] [PubMed]
- De Nadal, E.; Zapater, M.; Alepuz, P.M.; Sumoy, L.; Mas, G.; Posas, F. The MAPK Hog1 recruits Rpd3 histone deacetylase to activate osmoresponsive genes. Nature 2004, 427, 370–374. [Google Scholar] [PubMed]
- Alepuz, P.M.; de Nadal, E.; Zapater, M.; Ammerer, G.; Posas, F. Osmostress-induced transcription by Hot1 depends on a Hog1-mediated recruitment of the RNA Pol II. EMBO J. 2003, 22, 2433–2442. [Google Scholar] [PubMed]
- Moskvina, E.; Schuller, C.; Maurer, C.T.; Mager, W.H.; Ruis, H. A search in the genome of Saccharomyces cerevisiae for genes regulated via stress response elements. Yeast 1998, 14, 1041–1050. [Google Scholar] [PubMed]
- Causton, H.C.; Ren, B.; Koh, S.S.; Harbison, C.T.; Kanin, E.; Jennings, E.G.; Lee, T.I.; True, H.L.; Lander, E.S.; Young, R.A. Remodeling of yeast genome expression in response to environmental changes. Mol. Biol. Cell 2001, 12, 323–337. [Google Scholar] [PubMed]
- Gasch, A.P. Comparative genomics of the environmental stress response in ascomycete fungi. Yeast 2007, 24, 961–976. [Google Scholar] [PubMed]
- O'Rourke, S.M.; Herskowitz, I.; O'Shea, E.K. Yeast go the whole HOG for the hyperosmotic response. Trends Genet. 2002, 18, 405–412. [Google Scholar] [PubMed]
- Akhtar, N.; Pahlman, A.K.; Larsson, K.; Corbett, A.H.; Adler, L. SGD1 encodes an essential nuclear protein of Saccharomyces cerevisiae that affects expression of the GPD1 gene for glycerol 3-phosphate dehydrogenase. FEBS Lett. 2000, 483, 87–92. [Google Scholar] [PubMed]
- Lin, H.; Nguyen, P.; Vancura, A. Phospholipase C interacts with Sgd1p and is required for expression of GPD1 and osmoresistance in Saccharomyces cerevisiae. Mol. Genet. Genomics. 2002, 267, 313–320. [Google Scholar] [PubMed]
- de Nadal, E.; Casadome, L.; Posas, F. Targeting the MEF2-like transcription factor Smp1 by the stress-activated Hog1 mitogen-activated protein kinase. Mol. Cell. Biol. 2003, 23, 229–237. [Google Scholar] [PubMed]
- Bilsland-Marchesan, E.; Arino, J.; Saito, H.; Sunnerhagen, P.; Posas, F. Rck2 kinase is a substrate for the osmotic stress-activated mitogen-activated protein kinase Hog1. Mol. Cell. Biol. 2000, 20, 3887–3895. [Google Scholar] [PubMed]
- Teige, M.; Scheikl, E.; Reiser, V.; Ruis, H.; Ammerer, G. Rck2, a member of the calmodulin-protein kinase family, links protein synthesis to high osmolarity MAP kinase signaling in budding yeast. Proc. Natl. Acad. Sci. USA 2001, 98, 5625–5630. [Google Scholar] [CrossRef]
- Norbeck, J.; Blomberg, A. The level of cAMP-dependent protein kinase A activity strongly affects osmotolerance and osmo-instigated gene expression changes in Saccharomyces cerevisiae. Yeast 2000, 16, 121–137. [Google Scholar] [PubMed]
- Marchler, G.; Schuller, C.; Adam, G.; Ruis, H. A Saccharomyces cerevisiae UAS element controlled by protein kinase A activates transcription in response to a variety of stress conditions. EMBO J. 1993, 12, 1997–2003. [Google Scholar] [PubMed]
- Ruis, H.; Schuller, C. Stress signaling in yeast. Bioessays 1995, 17, 959–965. [Google Scholar] [PubMed]
- Siderius, M.M.; Mager, W.H. The general stress response in search for a common denominator. In Yeast Stress Responses, 1st; Hohmann, S.M., Mager, W.H., Eds.; R.G. Landes Company: Austin, TX, USA, 1997; pp. 213–230. [Google Scholar]
- Dove, S.K.; Cooke, F.T.; Douglas, M.R.; Sayers, L.G.; Parker, P.J.; Michell, R.H. Osmotic stress activates phosphatidylinositol-3,5-bisphosphate synthesis. Nature 1997, 390, 187–192. [Google Scholar] [PubMed]
- Kawasaki, L.; Sanchez, O.; Shiozaki, K.; Aguirre, J. SakA MAP kinase is involved in stress signal transduction, sexual development and spore viability in Aspergillus nidulans. Mol. Microbiol. 2002, 45, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, D.; Asano, Y.; Marui, J.; Furukawa, K.; Kanamaru, K.; Kato, M.; Abe, K.; Kobayashi, T.; Yamashino, T.; Mizuno, T. The SskA and SrrA response regulators are implicated in oxidative stress responses of hyphae and asexual spores in the phosphorelay signaling network of Aspergillus nidulans. Biosci. Biotechnol. Biochem. 2007, 71, 1003–1014. [Google Scholar] [PubMed]
- Vargas-Perez, I.; Sanchez, O.; Kawasaki, L.; Georgellis, D.; Aguirre, J. Response regulators SrrA and SskA are central components of a phosphorelay system involved in stress signal transduction and asexual sporulation in Aspergillus nidulans. Eukaryotic Cell 2007, 6, 1570–1583. [Google Scholar] [PubMed]
- Catlett, N.L.; Yoder, O.C.; Turgeon, B.G. Whole-genome analysis of two-component signal transduction genes in fungal pathogens. Eukaryotic Cell 2003, 2, 1151–1161. [Google Scholar] [PubMed]
- Hagiwara, D.; Matsubayashi, Y.; Marui, J.; Furukawa, K.; Yamashino, T.; Kanamaru, K.; Kato, M.; Abe, K.; Kobayashi, T.; Mizuno, T. Characterization of the NikA histidine kinase implicated in the phosphorelay signal transduction of Aspergillus nidulans, with special reference to fungicide responses. Biosci. Biotechnol. Biochem. 2007, 71, 844–847. [Google Scholar] [PubMed]
- Furukawa, K.; Katsuno, Y.; Urao, T.; Yabe, T.; Yamada-Okabe, T.; Yamada-Okabe, H.; Yamagata, Y.; Abe, K.; Nakajima, T. Isolation and functional analysis of a gene, tcsB, encoding a transmembrane hybrid-type histidine kinase from Aspergillus nidulans. Appl. Environ. Microbiol. 2002, 68, 5304–5310. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Kanamaru, K.; Azuma, N.; Kato, M.; Kobayashi, T. GFP-tagged expression analysis revealed that some histidine kinases of Aspergillus nidulans show temporally and spatially different expression during the life cycle. Biosci. Biotechnol. Biochem. 2008, 72, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Virginia, M.; Appleyard, C.L.; McPheat, W.L.; Stark, M.J. A novel 'two-component' protein containing histidine kinase and response regulator domains required for sporulation in Aspergillus nidulans. Curr. Genet. 2000, 37, 364–372. [Google Scholar] [PubMed]
- Krantz, M.; Becit, E.; Hohmann, S. Comparative genomics of the HOG-signalling system in fungi. Curr. Genet. 2006, 49, 137–151. [Google Scholar] [PubMed]
- Zarrinpar, A.; Park, S.H.; Lim, W.A. Optimization of specificity in a cellular protein interaction network by negative selection. Nature 2003, 426, 676–680. [Google Scholar] [PubMed]
- Virag, A.; Lee, M.P.; Si, H.; Harris, S.D. Regulation of hyphal morphogenesis by cdc42 and rac1 homologues in Aspergillus nidulans. Mol. Microbiol. 2007, 66, 1579–1596. [Google Scholar] [PubMed]
- Wei, H.; Requena, N.; Fischer, R. The MAPKK kinase SteC regulates conidiophore morphology and is essential for heterokaryon formation and sexual development in the homothallic fungus Aspergillus nidulans. Mol. Microbiol. 2003, 47, 1577–1588. [Google Scholar] [PubMed]
- Graessle, S.; Dangl, M.; Haas, H.; Mair, K.; Trojer, P.; Brandtner, E.M.; Walton, J.D.; Loidl, P.; Brosch, G. Characterization of two putative histone deacetylase genes from Aspergillus nidulans. Biochim. Biophys. Acta 2000, 1492, 120–126. [Google Scholar] [PubMed]
- Aguirre, J.; Rios-Momberg, M.; Hewitt, D.; Hansberg, W. Reactive oxygen species and development in microbial eukaryotes. Trends Microbiol. 2005, 13, 111–118. [Google Scholar] [PubMed]
- Furukawa, K.; Yoshimi, A.; Furukawa, T.; Hoshi, Y.; Hagiwara, D.; Sato, N.; Fujioka, T.; Mizutani, O.; Mizuno, T.; Kobayashi, T.; et al. Novel reporter gene expression systems for monitoring activation of the Aspergillus nidulans HOG pathway. Biosci. Biotechnol. Biochem. 2007, 71, 1724–1730. [Google Scholar] [PubMed]
- Borgia, P.T.; Miao, Y.; Dodge, C.L. The orlA gene from Aspergillus nidulans encodes a trehalose-6-phosphate phosphatase necessary for normal growth and chitin synthesis at elevated temperatures. Mol. Microbiol. 1996, 20, 1287–1296. [Google Scholar] [PubMed]
- d'Enfert, C.; Fontaine, T. Molecular characterization of the Aspergillus nidulans treA gene encoding an acid trehalase required for growth on trehalose. Mol. Microbiol. 1997, 24, 203–216. [Google Scholar] [PubMed]
- Fillinger, S.; Chaveroche, M.K.; van Dijck, P.; de Vries, R.; Ruijter, G.; Thevelein, J.; d'Enfert, C. Trehalose is required for the acquisition of tolerance to a variety of stresses in the filamentous fungus Aspergillus nidulans. Microbiology 2001, 147, 1851–1862. [Google Scholar] [PubMed]
- Fillinger, S.; Ruijter, G.; Tamas, M.J.; Visser, J.; Thevelein, J.M.; d'Enfert, C. Molecular and physiological characterization of the NAD-dependent glycerol 3-phosphate dehydrogenase in the filamentous fungus Aspergillus nidulans. Mol. Microbiol. 2001, 39, 145–157. [Google Scholar] [PubMed]
- de Vries, R.P.; Flitter, S.J.; van de Vondervoort, P.J.; Chaveroche, M.K.; Fontaine, T.; Fillinger, S.; Ruijter, G.J.; d'Enfert, C.; Visser, J. Glycerol dehydrogenase, encoded by gldB is essential for osmotolerance in Aspergillus nidulans. Mol. Microbiol. 2003, 49, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Sarfati, J.; Latge, J.P.; Calderone, R. The role of the sakA (Hog1) and tcsB (sln1) genes in the oxidant adaptation of Aspergillus fumigatus. Med. Mycol. 2006, 44, 211–218. [Google Scholar] [PubMed]
- Reyes, G.; Romans, A.; Nguyen, C.K.; May, G.S. Novel mitogen-activated protein kinase MpkC of Aspergillus fumigatus is required for utilization of polyalcohol sugars. Eukaryotic Cell 2006, 5, 1934–1940. [Google Scholar] [PubMed]
- Xue, T.; Nguyen, C.K.; Romans, A.; May, G.S. A mitogen-activated protein kinase that senses nitrogen regulates conidial germination and growth in Aspergillus fumigatus. Eukaryotic Cell 2004, 3, 557–560. [Google Scholar] [PubMed]
- Hagiwara, D.; Mizuno, T.; Abe, K. Characterization of NikA histidine kinase and two response regulators with special reference to osmotic adaptation and asexual development in Aspergillus nidulans. Biosci. Biotechnol. Biochem. 2009, 73, 1566–1571. [Google Scholar] [PubMed]
- Hagiwara, D.; Asano, Y.; Marui, J.; Yoshimi, A.; Mizuno, T.; Abe, K. Transcriptional profiling for Aspergillus nidulans HogA MAPK signaling pathway in response to fludioxonil and osmotic stress. Fungal Genet. Biol. 2009, 46, 868–878. [Google Scholar] [PubMed]
- Ma, Y.; Qiao, J.; Liu, W.; Wan, Z.; Wang, X.; Calderone, R.; Li, R. The sho1 sensor regulates growth, morphology, and oxidant adaptation in Aspergillus fumigatus but is not essential for development of invasive pulmonary aspergillosis. Infect Immun. 2008, 76, 1695–1701. [Google Scholar] [PubMed]
- Alonso-Monge, R.; Navarro-Garcia, F.; Roman, E.; Negredo, A.I.; Eisman, B.; Nombela, C.; Pla, J. The Hog1 mitogen-activated protein kinase is essential in the oxidative stress response and chlamydospore formation in Candida albicans. Eukaryotic Cell 2003, 2, 351–361. [Google Scholar] [PubMed]
- Hicks, J.; Lockington, R.A.; Strauss, J.; Dieringer, D.; Kubicek, C.P.; Kelly, J.; Keller, N. RcoA has pleiotropic effects on Aspergillus nidulans cellular development. Mol. Microbiol. 2001, 39, 1482–1493. [Google Scholar] [PubMed]
- Todd, R.B.; Hynes, M.J.; Andrianopoulos, A. The Aspergillus nidulans rcoA gene is required for veA-dependent sexual development. Genetics 2006, 174, 1685–1688. [Google Scholar] [PubMed]
- Han, K.H.; Seo, J.A.; Yu, J.H. A putative G protein-coupled receptor controls growth, germination and coordinated development in Aspergillus nidulans. Fungal Genet. Newsl. 2003, 50. [Google Scholar]
- Mert, H.H.; Ekmekci, S. The effect of salinity and osmotic pressure of the medium on the growth, sporulation and changes in the total organic acid content of Aspergillus flavus and Penicillium chrysogenum. Mycopathologia 1987, 100, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Cotty, P.J. Virulence and cultural characteristics of two Aspergillus flavus strains pathogenic on cotton. Phytopathology 1989, 79, 808–814. [Google Scholar]
- Calvo, A.M. The VeA regulatory system and its role in morphological and chemical development in fungi. Fungal Genet. Biol. 2008, 45, 1053–1061. [Google Scholar] [PubMed]
- El-Abyad, M.S.; Hindorf, H.; Rizk, M.A. Impact of salinity stress on soil-borne fungi of sugarbeet. Plant Soil 1988, 110, 27–32. [Google Scholar]
- Ramos, A.J.M.; Magan, N.; Sanchis, V. Osmotic and matric potential effects on growth, sclerotia and partitioning of polyols and sugars in colonies and spores of Aspergillus ochraceus. Mycol. Res. 1999, 103, 141–147. [Google Scholar] [CrossRef]
- Whipps, J.M.M.; Magan, N. Effects of nutrient status and water potential of media on fungal growth and antagonist-pathogen interactions. EPPO Bulletin 1986, 17, 581–591. [Google Scholar] [CrossRef]
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar] [PubMed]
- Hicks, J.; Lockington, R.A.; Strauss, J.; Dieringer, D.; Kubicek, C.P.; Kelly, J.; Keller, N. RcoA has pleiotropic effects on Aspergillus nidulans cellular development. Mol. Microbiol. 2001, 39, 1482–1493. [Google Scholar] [PubMed]
- Ochiai, N.; Tokai, T.; Nishiuchi, T.; Takahashi-Ando, N.; Fujimura, M.; Kimura, M. Involvement of the osmosensor histidine kinase and osmotic stress-activated protein kinases in the regulation of secondary metabolism in Fusarium graminearum. Biochem. Biophys. Res. Commun. 2007, 363, 639–644. [Google Scholar] [PubMed]
- Igbaria, A.; Lev, S.; Rose, M.S.; Lee, B.N.; Hadar, R.; Degani, O.; Horwitz, B.A. Distinct and combined roles of the MAP kinases of Cochliobolus heterostrophus in virulence and stress responses. Mol. Plant Microbe Interact. 2008, 21, 769–780. [Google Scholar] [PubMed]
- Park, S.M.; Choi, E.S.; Kim, M.J.; Cha, B.J.; Yang, M.S.; Kim, D.H. Characterization of HOG1 homologue, CpMK1, from Cryphonectria parasitica and evidence for hypovirus-mediated perturbation of its phosphorylation in response to hypertonic stress. Mol. Microbiol. 2004, 51, 1267–1277. [Google Scholar] [CrossRef] [PubMed]
- Dixon, K.P.; Xu, J.R.; Smirnoff, N.; Talbot, N.J. Independent signaling pathways regulate cellular turgor during hyperosmotic stress and appressorium-mediated plant infection by Magnaporthe grisea. Plant Cell 1999, 11, 2045–2058. [Google Scholar] [PubMed]
- Chapmann, R.F. The Insects: Structure and Function; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Wang, C.; Duan, Z.; St Leger, R.J. MOS1 osmosensor of Metarhizium anisopliae is required for adaptation to insect host hemolymph. Eukaryotic Cell 2008, 7, 302–309. [Google Scholar] [PubMed]
- Vylkova, S.; Jang, W.S.; Li, W.; Nayyar, N.; Edgerton, M. Histatin 5 initiates osmotic stress response in Candida albicans via activation of the Hog1 mitogen-activated protein kinase pathway. Eukaryotic Cell 2007, 6, 1876–1888. [Google Scholar] [PubMed]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Duran, R.; Cary, J.W.; Calvo, A.M. Role of the Osmotic Stress Regulatory Pathway in Morphogenesis and Secondary Metabolism in Filamentous Fungi. Toxins 2010, 2, 367-381. https://doi.org/10.3390/toxins2040367
Duran R, Cary JW, Calvo AM. Role of the Osmotic Stress Regulatory Pathway in Morphogenesis and Secondary Metabolism in Filamentous Fungi. Toxins. 2010; 2(4):367-381. https://doi.org/10.3390/toxins2040367
Chicago/Turabian StyleDuran, Rocio, Jeffrey W. Cary, and Ana M. Calvo. 2010. "Role of the Osmotic Stress Regulatory Pathway in Morphogenesis and Secondary Metabolism in Filamentous Fungi" Toxins 2, no. 4: 367-381. https://doi.org/10.3390/toxins2040367