Environmental Factors and Interactions with Mycobiota of Grain and Grapes: Effects on Growth, Deoxynivalenol and Ochratoxin Production by Fusarium culmorum and Aspergillus carbonarius

Abstract
:1. Introduction
2. Results
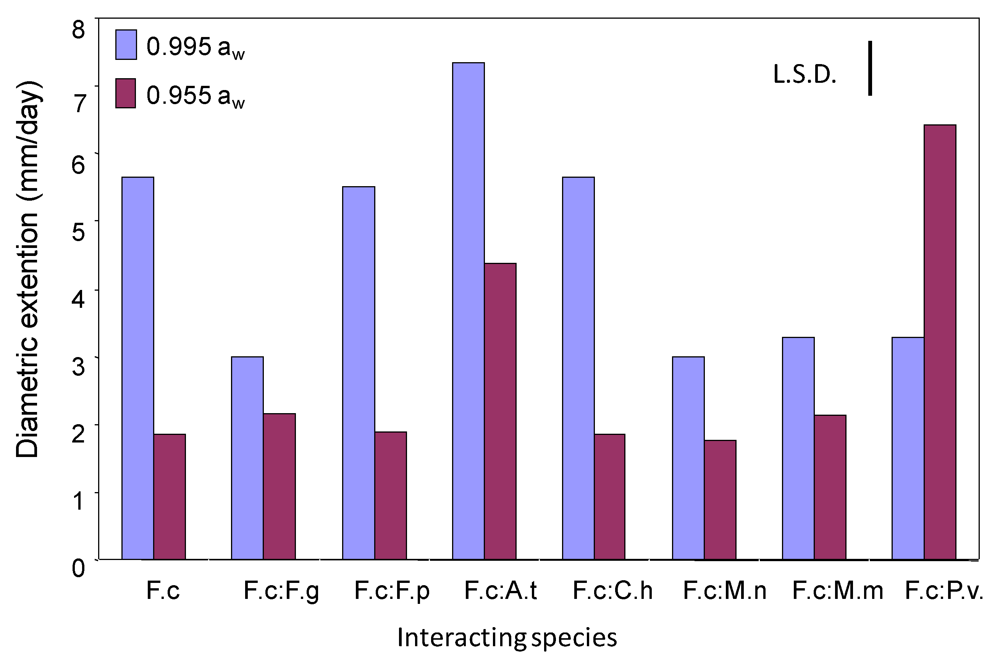
2.1. Effects of interactions between Fusarium culmorum and other mycobiota on growth and DON production on wheat-based matrices

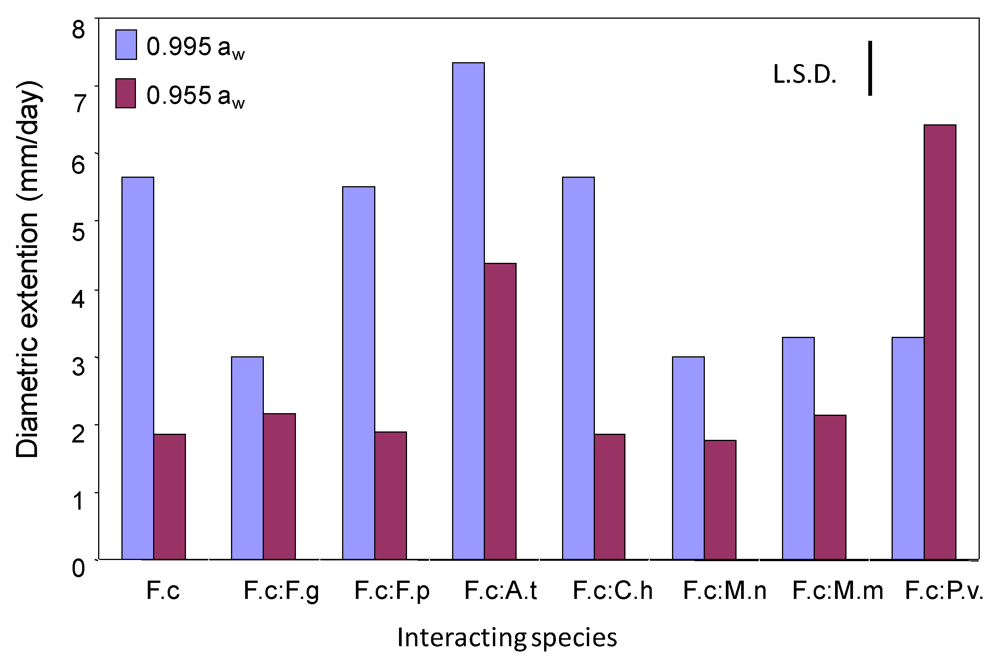{kind=link}
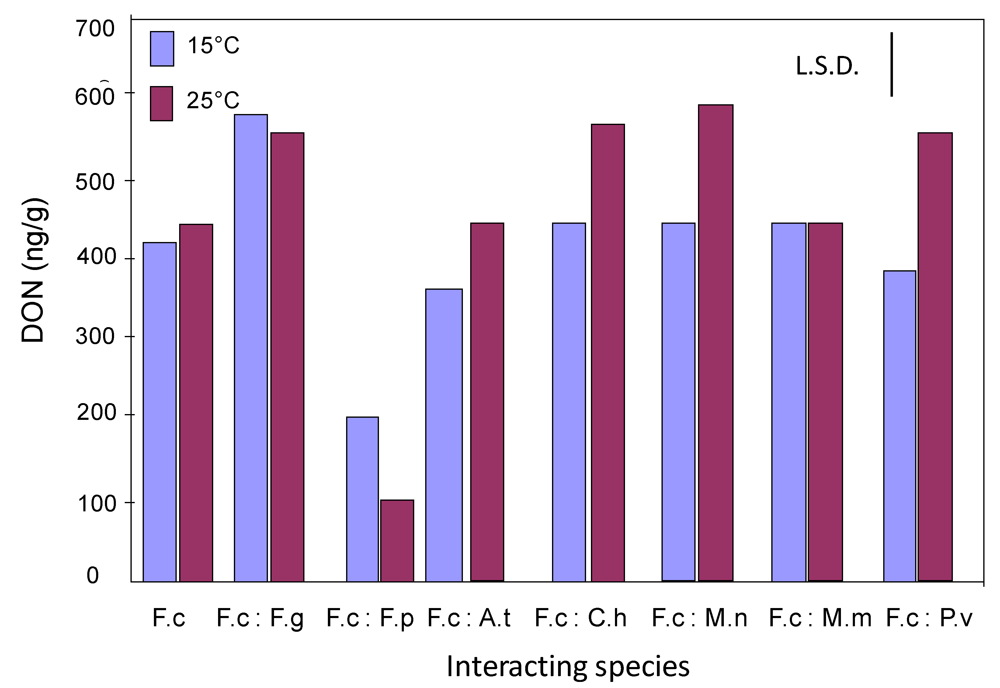{kind=link}
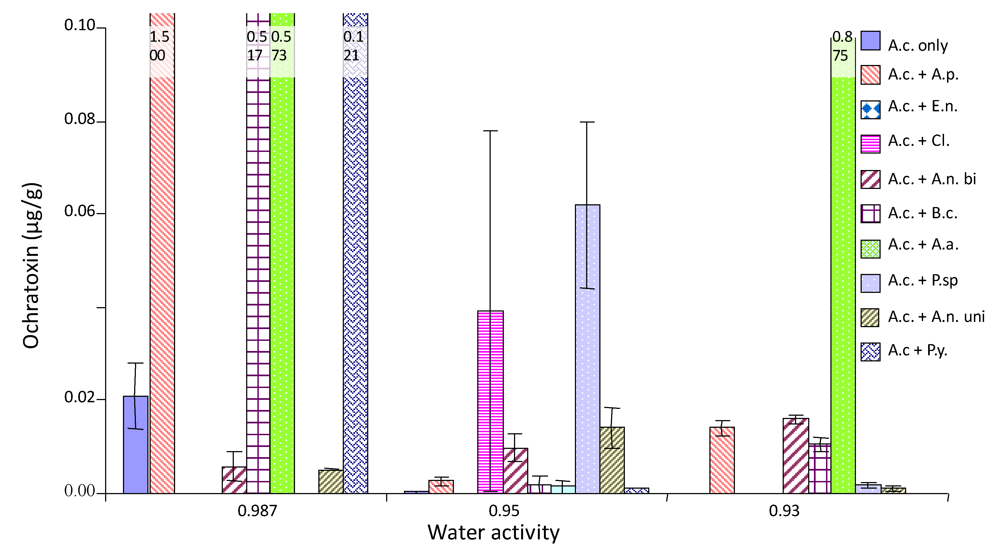{kind=link}
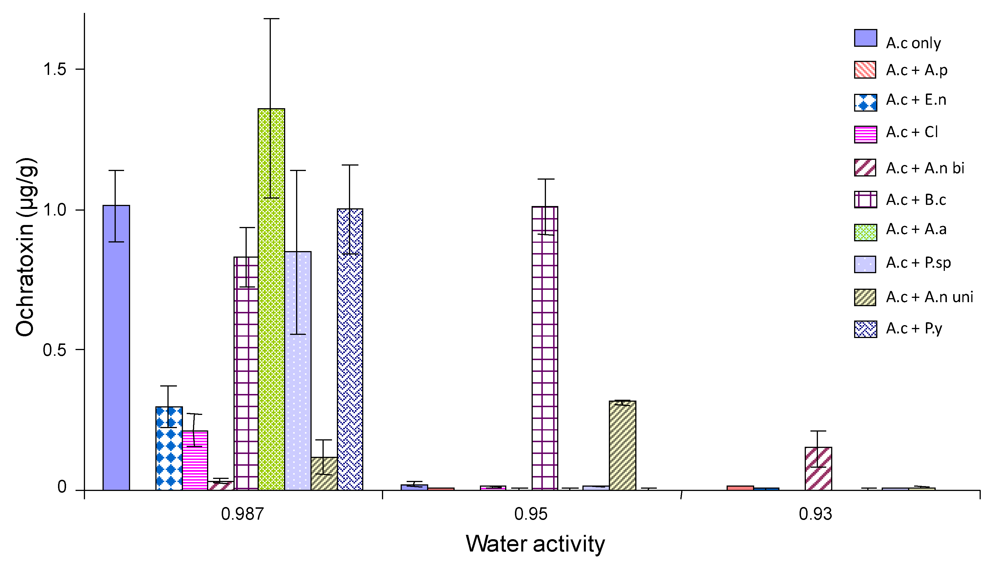{kind=link}
{kind=link}
| Species | Temperature (°C) | |||||
|---|---|---|---|---|---|---|
| 15 | 25 | |||||
| 0.995 | 0.955 | ID | 0.995 | 0.955 | ID | |
| F. graminearum | 0/4 | 0/4 | 0/8 | 0/4 | 0/4 | 0/8 |
| F. poae | 2/2 | 0/4 | 2/6 | 1/1 | 2/2 | 3/3 |
| A. tenuissima | 4/0 | 4/0 | 8/0 | 4/0 | 4/0 | 8/0 |
| C. herbarum | 4/0 | 4/0 | 8/0 | 4/0 | 4/0 | 8/0 |
| M. nivale | 2/2 | 4/0 | 6/2 | 4/0 | 4/0 | 8/0 |
| M. nivale var majus | 2/2 | 0/4 | 2/6 | 4/0 | 4/0 | 8/0 |
| P. verrucosum | 4/0 | 2/2 | 6/2 | 2/2 | 4/0 | 6/2 |
| Total ID | 18/10 | 14/14 | 32/14 | 19/7 | 22/6 | 41/13 |
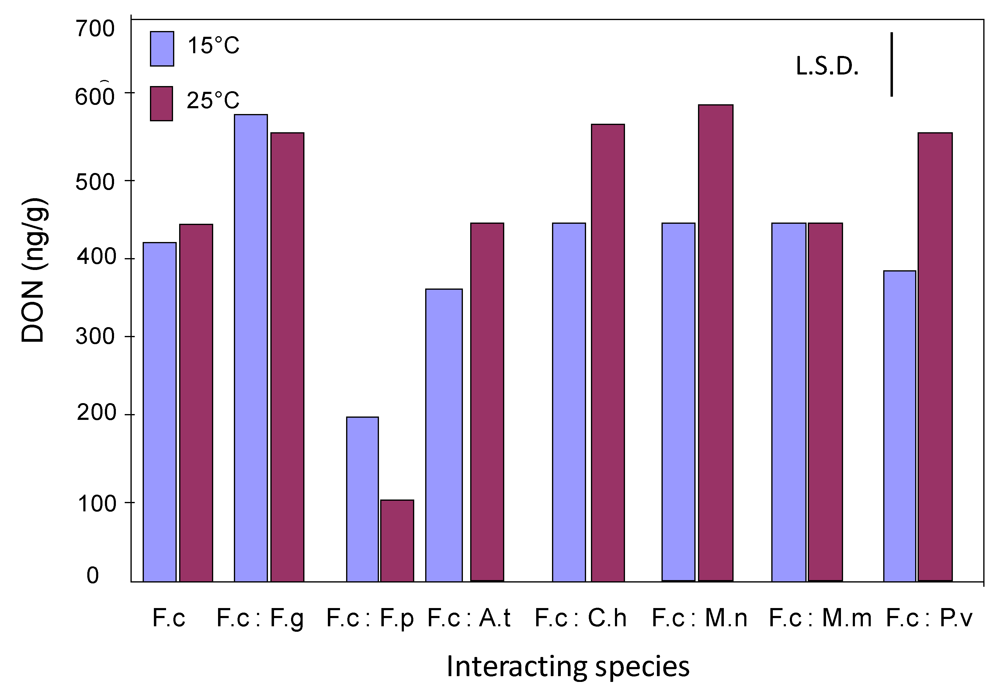
2.2. Effect of interactions between F. culmorum and other grain mycobiota on deoxynivalenol production on wheat grain
| Temperature (°C) | ||||
|---|---|---|---|---|
| 15 | 25 | |||
| Water activity | 0.995 | 0.955 | 0.995 | 0.955 |
| F.c:F.g | NS | ↑ | ↓ | ↑ |
| F.c:F.p | NS | NS | ↓ | NS |
| F.c:A.t | NS | NS | ↓ | NS |
| F.c:C.h | NS | NS | ↓ | ↑ |
| F.c:M.n | NS | NS | ↓ | ↑ |
| F.c:M.m | NS | NS | ↓ | NS |
| F.c:P.v | NS | NS | ↓ | ↑ |
2.3. Effects of interactions between A. carbonarius and other mycobiota on growth and OTA production on grape-based matrices
| 30 °C | 25 °C | 20 °C | |||||||
|---|---|---|---|---|---|---|---|---|---|
| aw/species | 0.98 | 0.95 | 0.93 | 0.98 | 0.95 | 0.93 | 0.98 | 0.95 | 0.93 |
| Cladosporium species | 4/0 | 4/0 | 4/0 | 4/0 | 4/0 | 2/2 | 4/0 | 2/2 | 2/2 |
| Epicoccum nigrum | 2/2 | 2/2 | 2/2 | 3/3 | 3/3 | 3/3 | 3/3 | 3/3 | 3/3 |
| Aureobasidium pullulans | 4/0 | 4/0 | 4/0 | 4/0 | 4/0 | 2/2 | 2/2 | 2/2 | 1/1 |
| Aspergillus section Nigri biseriate | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 | 2/2 |
| Botrytis cinerea | 4/0 | 4/0 | 4/0 | 4/0 | 4/0 | 4/0 | 2/2 | 2/2 | 2/2 |
| Alternaria alternaria | 4/0 | 4/0 | 4/0 | 4/0 | 4/0 | 2/2 | 2/2 | 2/2 | 2/2 |
| Aspergillus section Nigri uniseriate | 2/2 | 2/2 | 2/2 | 2/2 | 3/3 | 2/2 | 3/3 | 3/3 | 2/2 |
| Phoma species | 4/0 | 4/0 | 4/0 | 4/0 | 4/0 | 2/2 | 2/2 | 2/2 | 2/2 |
| Pink yeast | 4/0 | 4/0 | 4/0 | 4/0 | 4/0 | 4/0 | 4/0 | 4/0 | 4/0 |
| ID | 30/6 | 30/6 | 30/6 | 31/7 | 32/8 | 19/15 | 24/16 | 22/18 | 20/16 |
| 30 °C | 25 °C | 20 °C | |||||||
|---|---|---|---|---|---|---|---|---|---|
| aw/species | 0.987 | 0.95 | 0.93 | 0.987 | 0.95 | 0.93 | 0.987 | 0.95 | 0.93 |
| Aspergillus carbonarius | 5.1 | 4.8 | 4.0 | 5.0 | 4.5 | 3.5 | 3.9 | 2.8 | 2.2 |
| Cladosporium species | 5.4 | 5.8+ | 6.6+ | 5.2 | 5.0+ | 3.2 | 3.5 | 1.9- | 1.2- |
| Epicoccum nigrum | N/A | N/A | N/A | 5.7+ | 4.4 | 2.3- | 5.9+ | 3.4+ | 2.5 |
| Aureobasidium pullulans | 5.9+ | 6.3+ | 4.7+ | 3.8- | 5.0+ | 3.7 | 2.6- | 1.6- | 2.4 |
| Aspergillus section Nigri biseriate | 2.2- | 2.2- | 2.4- | 3.6- | 2.2- | 1.5- | 3.6 | 2.5 | 0.9- |
| Botrytis cinerea | 7.1+ | 7.6+ | 4.2 | 4.8 | 4.4 | 3.6 | 2.9- | 2.6 | 1.2- |
| Alternaria alternaria | 6.5+ | 5.8+ | 4.1 | 5.3 | 4.6 | 4.1+ | 3.6 | 3.8+ | 1.9 |
| Aspergillus section Nigri uniseriate | 4.5- | 3.4- | 4.0 | 4.9 | 3.3- | 2.7- | 3.6 | 2.2- | 0.9- |
| Phoma species | 5.2 | 4.9 | 6.4+ | 4.7 | 3.5- | 2.6- | 3.6 | 2.2- | 1.0- |
| Pink yeast | 6.7+ | 5.8+ | 4.2 | 4.9 | 4.6 | 1.8- | 3.7 | 2.6 | 1.4- |
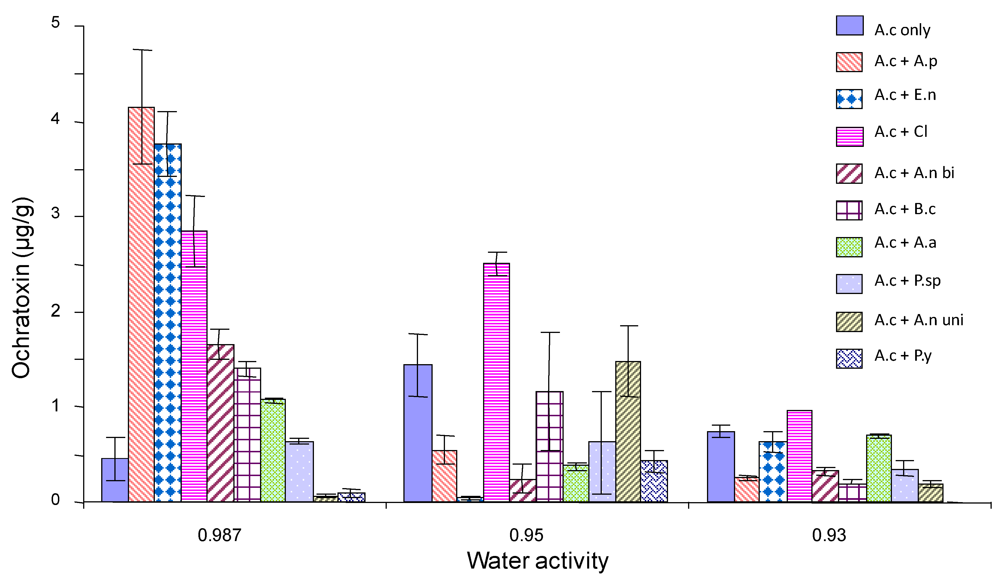
2.4. Ochratoxin A production by A. carbonarius when grown against other common grape fungi
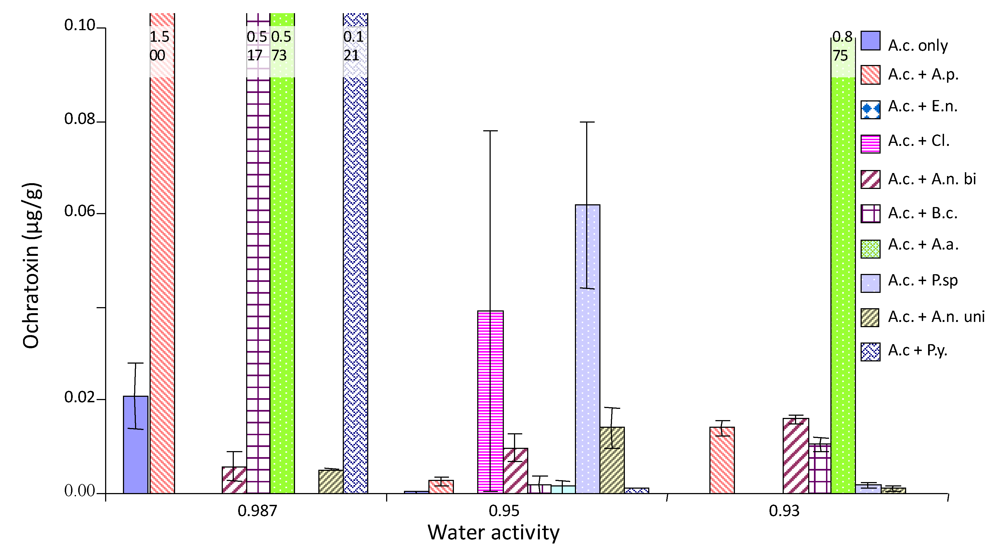

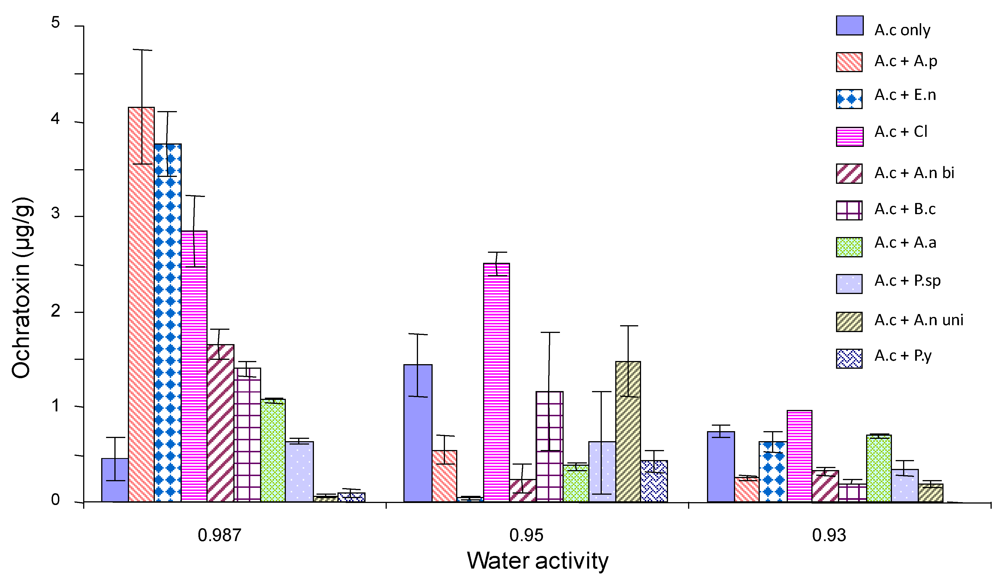
3. Discussion
3.1. F. culmorum and DON production
3.2. A. carbonarius and OTA production
4. Experimental Section
4.1. Fungal isolates used in this study
| Ecosystem type | Strain number | Cereals | Grapes |
|---|---|---|---|
| Fusarium culmorum (DON) | CC171 | + | - |
| Fusarium graminearum (DON) | CC175 | + | - |
| Fusarium poae | CC179 | + | - |
| Aspergillus carbonarius (OTA) | IMI388653 | - | + |
| Aspergillus carbonarius (uniseriate) | IMI388862 | - | + |
| Aspergillus niger (biseriate) | IMI388550 | - | + |
| Penicillium verrucosum (OTA) | IBT2266 | - | + |
| Microdochium nivale | 18/1/N | + | - |
| Microdochium nivale var majus | 1/1M | + | - |
| Epicoccum nigrum | SVF01 | + | + |
| Cladosporium herbarum | IBT7961 | + | + |
| Alternaria alternata | IBT8320 | + | + |
| Aureobasidium pullulans | SVF02 | - | + |
| Botrytis cinerea | SVF03 | - | + |
| Pink yeast (Sporobolomyces species) | SVF06 | - | + |
| Phoma species | SVF05 | - | + |
4.2. Media used
4.3. Measurements of growth rates and interaction scores
4.4. Mycotoxin analyses
5. Conclusions
Acknowledgements
References
- Magan, N.; Lacey, J. The phylloplane microbial populations of wheat and effect of late fungicide applications. Ann. Appl. Biol. 1986, 109, 109. [Google Scholar]
- Bellí, N.; Mitchell, D.; Marín, S.; Alegre, I.; Ramos, A.J.; Magan, N.; Sanchis, V. Ochratoxin A and producing fungi in Spanish wine grapes and their relationship with climatic conditions. Eur. J. Plant. Pathol. 2005, 113, 233–239. [Google Scholar] [CrossRef]
- Magan, N.; Aldred, D. Why to fungi produce mycotoxins? In Food Mycology: A Multifaceted Approach to Fungi and Food; Dijksterhuis, J., Samson, R.A., Eds.; Taylor & Francis: Boca Raton,FL,USA, 2007; pp. 121–133. [Google Scholar]
- Magan, N.; Aldred, D. Environmental fluxes and fungal interactions: maintaining a competitive edge. In Stress in Yeasts and Filamentous Fungi; van West, P., Avery, S., Stratford., M., Eds.; Elsevier Ltd.: Amsterdam,The Netherland, 2007. [Google Scholar]
- Magan, N.; Lacey, J. The effect of water activity, temperature and substrate on interactions between field and storage fungi. Trans. Br. Mycol. Soc. 2004, 82, 83–93. [Google Scholar]
- Marin, S.; Sanchis, V.; Ramos, A.G.; Magan, N. Environmental factors, interspecific interactions, and niche overlap between Fusarium moniliforme and F. proliferatum and Fusarium graminearum, Aspergillus and Penicillium spp. isolated from maize. Mycol. Res. 1998, 102, 831–837. [Google Scholar] [CrossRef]
- Lee, H.B.; Magan, N. Environmental influences on in vitro interspecific interactions between A. ochraceus and other maize spoilage fungi on growth and ochratoxin production. Mycopathologia 2000, 146, 43–47. [Google Scholar]
- Marin, S.; Magan, N.; Ramos, A.J.; Sanchis, V. Fumonisin-producing strains of Fusarium: A review of their ecophysiology. J. Food Prot. 2004, 67, 1792–1805. [Google Scholar]
- Giorni, P.; Magan, N.; Battilani, P. Environmental factors modify carbon nutritional patterns and niche overlap between Aspergillus flavus and Fusarium verticillioides strains from maize. J. Food Microbiol. 2009, 130, 213–218. [Google Scholar] [CrossRef] [Green Version]
- Jennings, P.; Turner, J.A.; Nicholson, P. Overview of Fusarium ear blight in the UK-effect of fungicide treatment on disease control and mycotoxin production. The BCPC Conference-Pests and Diseases, Brighton, UK, 13-16 November 2000; Volume 2, pp. 707–712.
- Simpson, D.R.; Weston, G.E.; Turner, J.A.; Jennings, P.; Nicholson, P. Differential control of head blight pathogens of wheat by fungicides and consequences for mycotoxin contamination of grain. Eur. J. Plant. Pathol. 2001, 107, 421–431. [Google Scholar] [CrossRef]
- Ramakrishna, N.; Lacey, J.; Smith, J.E. The effects of fungal competition on colonization of barley grain by Fusarium sporotrichioides on T-2 toxin formation. Food Addit. Contam. 1996, 13, 939–948. [Google Scholar] [PubMed]
- Edwards, S.C.; Pirgozliev, S.R.; Hare, M.C.; Jenkinson, P. Quantification of trichothecene-producing Fusarium species in harvested grain by competitive PCR to determine efficacies of fungicides against Fusarium head blight of winter wheat. Appl. Environ. Microbiol. 2001, 67, 1575–1580. [Google Scholar] [CrossRef]
- Bateman, G.L.; Murray, G. Seasonal variations in populations of Fusarium species in wheat-field soil. Appl. Soil Ecol. 2001, 18, 117–128. [Google Scholar] [CrossRef]
- Mitchell, D.; Parra, R.; Aldred, D.; Magan, N. Water and temperature relations of growth and ochratoxin A production by Aspergillus carbonarius strainsfrom grapes in Europe and Israel. J. Appl. Microbiol. 2004, 97, 439–445. [Google Scholar]
- De Cal, A.; Pascual, S.; Melgarejo, P. Nutritional requirements of antagonists to peach twig blight, Monilinia laxa, in relation to biocontrol. Mycopathologia 1993, 121, 21–26. [Google Scholar] [CrossRef]
- Cooney, J.M.; Lauren, D.R.; di Menna, M.E. Impact of competitive fungi on trichothecene production by Fusarium graminearum. J. Agric. Food Chem. 2001, 49, 522–526. [Google Scholar] [CrossRef]
- Bragulat, M.R.; Abarca, M.L.; Cabanas, F.J. An easy screening method for fungi producing ochratoxin A in pure culture. Int. J. Food Microbiol. 2001, 71, 139–144. [Google Scholar] [CrossRef]
- Schmidt-Heydt, M.; Geisen, R. A microarray for monitoring the production of mycotoxins in food. Int. J. Food Microbiol. 2007, 117, 131–140. [Google Scholar] [CrossRef]
- Jurado, M.; Marin, P.; Magan, N.; González-Jaén, M.T. Relationship between solute and matric potential stress, temperature, and growth and FUM1 gene expression in two Fusarium verticillioides strains from Spai. Appl. Environ. Microbiol. 2008, 74, 2032–2036. [Google Scholar]
- Doohan, F.M.; Weston, G.; Rezanoor, H.N.; Parry, D.W.; Nicholson, P. Development and use of a reverse transcription-PCR assay to study the expression of tri5 by Fusarium species in vitro and in planta. Appl. Environ. Microbiol. 1999, 65, 3850–3854. [Google Scholar]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Magan, N.; Aldred, D.; Hope, R.; Mitchell, D. Environmental Factors and Interactions with Mycobiota of Grain and Grapes: Effects on Growth, Deoxynivalenol and Ochratoxin Production by Fusarium culmorum and Aspergillus carbonarius. Toxins 2010, 2, 353-366. https://doi.org/10.3390/toxins2030353
Magan N, Aldred D, Hope R, Mitchell D. Environmental Factors and Interactions with Mycobiota of Grain and Grapes: Effects on Growth, Deoxynivalenol and Ochratoxin Production by Fusarium culmorum and Aspergillus carbonarius. Toxins. 2010; 2(3):353-366. https://doi.org/10.3390/toxins2030353
Chicago/Turabian StyleMagan, Naresh, David Aldred, Russell Hope, and David Mitchell. 2010. "Environmental Factors and Interactions with Mycobiota of Grain and Grapes: Effects on Growth, Deoxynivalenol and Ochratoxin Production by Fusarium culmorum and Aspergillus carbonarius" Toxins 2, no. 3: 353-366. https://doi.org/10.3390/toxins2030353