UDP-Glucosyltransferases from Rice, Brachypodium, and Barley: Substrate Specificities and Synthesis of Type A and B Trichothecene-3-O-β-d-glucosides

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Substrate Specificities of Trichothecene-Conjugating UGTs
2.2. Preparative Synthesis of Glucosides with OsUGT79 and HvUGT13248, and Purification
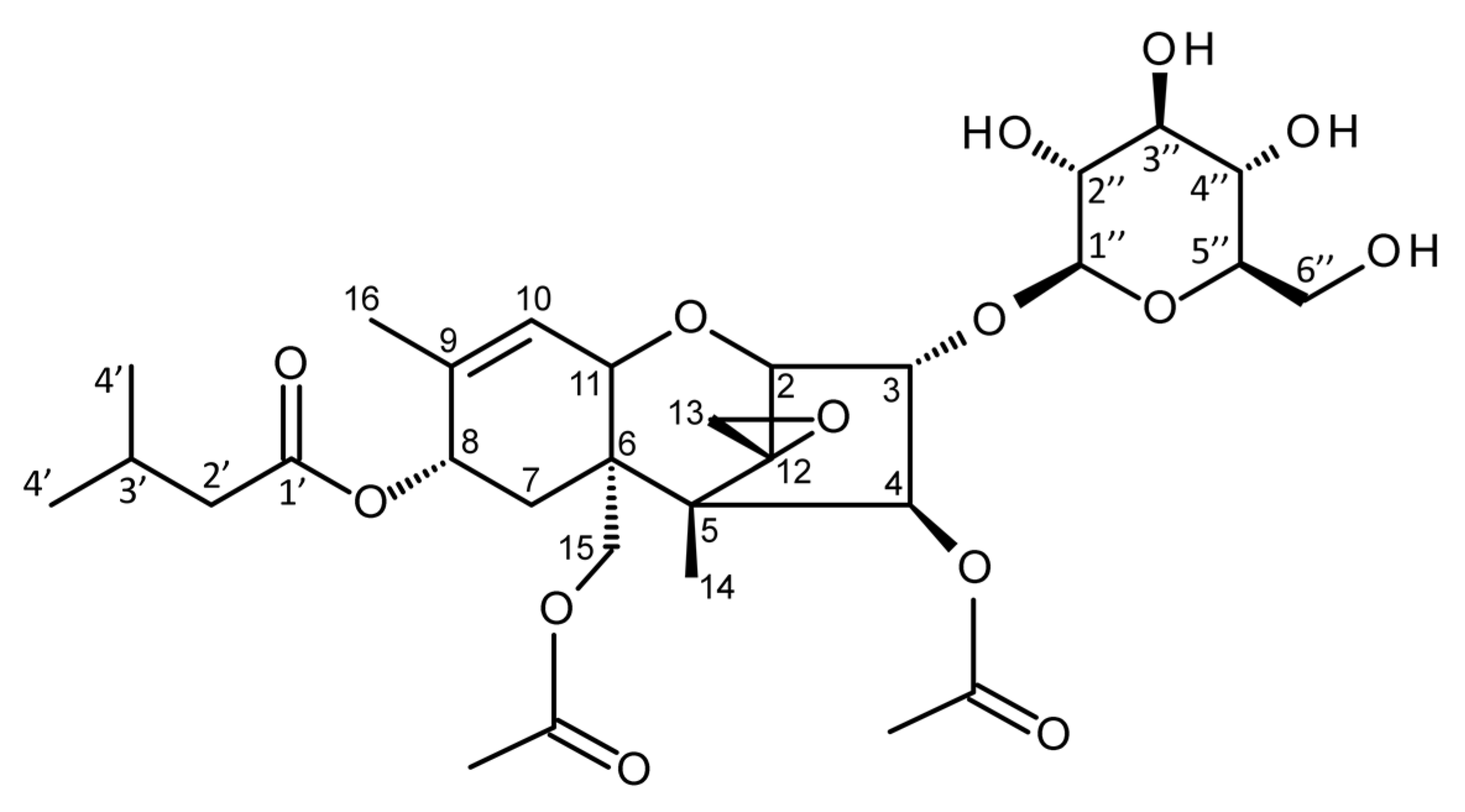
2.3. Characterization by HRMS, MS/HRMS, and NMR
3. Conclusions
4. Materials and Methods
4.1. Materials
4.2. Expression and Purification of OsUGT79, HvUGT13248, and Bradi5g03300
4.3. Glycosyltransferase Assays
4.4. LC-MS/MS Measurements
4.5. Large Scale Glucosylation of Trichothecenes and Purification Thereof
4.6. Characterization of the Produced Glucosides
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| -Glc | -glucoside |
| -3-Glc | -3-O-β-d-glucoside |
| 3-Ac-DON | 3-acetyl-deoxynivalenol |
| 15-Ac-DON | 15-acetyl-deoxynivalenol |
| DAS | 4,15-diacetoxyscirpenol |
| DON | deoxynivalenol |
| FHB | Fusarium head blight |
| FUSX | fusarenon X (4-acetyl-nivalenol) |
| HT2 | HT-2 toxin |
| IMAC | immobilized metal affinity chromatography |
| IPTG | isopropyl-β-d-1-thiogalactopyranoside |
| iso-NEO | iso-neosolaniol (15-deacetyl-8-acetyl-neosolaniol) |
| MalE | maltose binding protein |
| NEO | neosolaniol |
| NIV | nivalenol |
| LC-MS/MS | liquid chromatography coupled to tandem mass spectrometry |
| LC-(MS/)HRMS | liquid chromatography coupled to (tandem) high resolution mass spectrometry |
| T2 | T-2 toxin |
| T2 triol | T-2 triol |
| T2 tetraol | T-2 tetraol |
| QTL | quantitative trait locus |
| UGT | UDP-glucosyltransferase |
| UHPLC | ultra high performance liquid chromatography |
References
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Bai, G.-H.; Desjardins, A.; Plattner, R. Deoxynivalenol-nonproducing Fusarium graminearum causes initial infection, but does not cause Disease Spread in wheat spikes. Mycopathologia 2002, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Van der Lee, T.; Zhang, H.; van Diepeningen, A.; Waalwijk, C. Biogeography of Fusarium graminearum species complex and chemotypes: A review. Food Addit. Contam. Part A 2015, 32, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.; Barrier-Guillot, B.; Clasen, P.; Hietaniemi, V.; Pettersson, H. Emerging issues of HT-2 and T-2 toxins in European cereal production. World Mycotoxin J. 2009, 2, 173–179. [Google Scholar] [CrossRef]
- Krska, R.; Malachova, A.; Berthiller, F.; Van Egmond, H. Determination of T-2 and HT-2 toxins in food and feed: An update. World Mycotoxin J. 2014, 7, 131–142. [Google Scholar] [CrossRef]
- Thrane, U.; Adler, A.; Clasen, P.-E.; Galvano, F.; Langseth, W.; Lew, H.; Logrieco, A.; Nielsen, K.F.; Ritieni, A. Diversity in metabolite production by Fusarium langsethiae, Fusarium poae, and Fusarium sporotrichioides. Int. J. Food Microbiol. 2004, 95, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Buerstmayr, H.; Ban, T.; Anderson, J.A. QTL mapping and marker-assisted selection for Fusarium head blight resistance in wheat: A review. Plant Breed. 2009, 128, 1–26. [Google Scholar] [CrossRef]
- Schweiger, W.; Steiner, B.; Ametz, C.; Siegwart, G.; Wiesenberger, G.; Berthiller, F.; Lemmens, M.; Jia, H.; Adam, G.; Muehlbauer, G.J. Transcriptomic characterization of two major Fusarium resistance quantitative trait loci (QTLs), Fhb1 and Qfhs. ifa-5A, identifies novel candidate genes. Mol. Plant Pathol. 2013, 14, 772–785. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.; Blake-Kalff, M.; Davies, E. Detoxification of xenobiotics by plants: Chemical modification and vacuolar compartmentation. Trends Plant Sci. 1997, 2, 144–151. [Google Scholar] [CrossRef]
- Cole, D.J. Detoxification and activation of agrochemicals in plants. Pest Manag. Sci. 1994, 42, 209–222. [Google Scholar] [CrossRef]
- Warth, B.; Fruhmann, P.; Wiesenberger, G.; Kluger, B.; Sarkanj, B.; Lemmens, M.; Hametner, C.; Fröhlich, J.; Adam, G.; Krska, R. Deoxynivalenol-sulfates: Identification and quantification of novel conjugated (masked) mycotoxins in wheat. Anal. Bioanal. Chem. 2015, 407, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Schmeitzl, C.; Warth, B.; Fruhmann, P.; Michlmayr, H.; Malachová, A.; Berthiller, F.; Schuhmacher, R.; Krska, R.; Adam, G. The metabolic fate of deoxynivalenol and its acetylated derivatives in a wheat suspension culture: Identification and detection of DON-15-O-glucoside, 15-acetyl-DON-3-O-glucoside and 15-acetyl-DON-3-sulfate. Toxins 2015, 7, 3112–3126. [Google Scholar] [CrossRef] [PubMed]
- Kluger, B.; Bueschl, C.; Lemmens, M.; Berthiller, F.; Häubl, G.; Jaunecker, G.; Adam, G.; Krska, R.; Schuhmacher, R. Stable isotopic labelling-assisted untargeted metabolic profiling reveals novel conjugates of the mycotoxin deoxynivalenol in wheat. Anal. Bioanal. Chem. 2013, 405, 5031–5036. [Google Scholar] [CrossRef] [PubMed]
- Lemmens, M.; Scholz, U.; Berthiller, F.; Dall’Asta, C.; Koutnik, A.; Schuhmacher, R.; Adam, G.; Buerstmayr, H.; Mesterházy, Á.; Krska, R. The ability to detoxify the mycotoxin deoxynivalenol colocalizes with a major quantitative trait locus for Fusarium head blight resistance in wheat. Mol. Plant Microbe Interact. 2005, 18, 1318–1324. [Google Scholar] [CrossRef] [PubMed]
- Gunnaiah, R.; Kushalappa, A.C.; Duggavathi, R.; Fox, S.; Somers, D.J. Integrated metabolo-proteomic approach to decipher the mechanisms by which wheat QTL (Fhb1) contributes to resistance against Fusarium graminearum. PLoS ONE 2012, 7, e40695. [Google Scholar] [CrossRef] [PubMed]
- Rawat, N.; Pumphrey, M.O.; Liu, S.; Zhang, X.; Tiwari, V.K.; Ando, K.; Trick, H.N.; Bockus, W.W.; Akhunov, E.; Anderson, J.A. Wheat Fhb1 encodes a chimeric lectin with agglutinin domains and a pore-forming toxin-like domain conferring resistance to Fusarium head blight. Nat. Genet. 2016, 48, 1576–1580. [Google Scholar] [CrossRef] [PubMed]
- Kluger, B.; Bueschl, C.; Lemmens, M.; Michlmayr, H.; Malachova, A.; Koutnik, A.; Maloku, I.; Berthiller, F.; Adam, G.; Krska, R. Biotransformation of the mycotoxin deoxynivalenol in Fusarium resistant and susceptible near isogenic wheat lines. PLoS ONE 2015, 10, e0119656. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, W.; Steiner, B.; Vautrin, S.; Nussbaumer, T.; Siegwart, G.; Zamini, M.; Jungreithmeier, F.; Gratl, V.; Lemmens, M.; Mayer, K. Suppressed recombination and unique candidate genes in the divergent haplotype encoding Fhb1. Theor. Appl. Genet. 2016, 129, 1607–1623. [Google Scholar] [CrossRef] [PubMed]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; Saeger, S.D.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gareis, M.; Bauer, J.; Thiem, J.; Plank, G.; Grabley, S.; Gedek, B. Cleavage of Zearalenone-Glycoside, a “Masked” Mycotoxin, during Digestion in Swine. Zoonoses Public Health 1990, 37, 236–240. [Google Scholar] [CrossRef]
- Rychlik, M.; Humpf, H.-U.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a comprehensive definition of modified and other forms of mycotoxins including “masked” mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Scientific opinion on the risks for human and animal health related to the presence of modified forms of certain mycotoxins in food and feed. EFSA J. 2014, 3916. [Google Scholar] [CrossRef] [Green Version]
- Crews, C.; MacDonald, S.J. Chapter 2 Natural Occurrence of Masked Mycotoxins. In Masked Mycotoxins in Food: Formation, Occurrence and Toxicological Relevance; Dall’Asta, C., Berthiller, F., Eds.; The Royal Society of Chemistry: Croydon, UK, 2016; pp. 14–31. [Google Scholar]
- Broekaert, N.; Devreese, M.; van Bergen, T.; Schauvliege, S.; De Boevre, M.; De Saeger, S.; Vanhaecke, L.; Berthiller, F.; Michlmayr, H.; Malachová, A. In vivo contribution of deoxynivalenol-3-β-d-glucoside to deoxynivalenol exposure in broiler chickens and pigs: Oral bioavailability, hydrolysis and toxicokinetics. Arch. Toxicol. 2016, 91, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Poppenberger, B.; Berthiller, F.; Lucyshyn, D.; Sieberer, T.; Schuhmacher, R.; Krska, R.; Kuchler, K.; Glössl, J.; Luschnig, C.; Adam, G. Detoxification of the Fusarium mycotoxin deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana. J. Biol. Chem. 2003, 278, 47905–47914. [Google Scholar] [CrossRef] [PubMed]
- Nagl, V.; Schwartz, H.; Krska, R.; Moll, W.-D.; Knasmüller, S.; Ritzmann, M.; Adam, G.; Berthiller, F. Metabolism of the masked mycotoxin deoxynivalenol-3-glucoside in rats. Toxicol. Lett. 2012, 213, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Nagl, V.; Woechtl, B.; Schwartz-Zimmermann, H.E.; Hennig-Pauka, I.; Moll, W.-D.; Adam, G.; Berthiller, F. Metabolism of the masked mycotoxin deoxynivalenol-3-glucoside in pigs. Toxicol. Lett. 2014, 229, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Ohmichi, K.; Sakamoto, S.; Sago, Y.; Kushiro, M.; Nagashima, H.; Yoshida, M.; Nakajima, T. Detection of a new Fusarium masked mycotoxin in wheat grain by high-resolution LC–Orbitrap™ MS. Food Addit. Contam. Part A 2011, 28, 1447–1456. [Google Scholar] [CrossRef] [PubMed]
- Nathanail, A.V.; Syvähuoko, J.; Malachová, A.; Jestoi, M.; Varga, E.; Michlmayr, H.; Adam, G.; Sieviläinen, E.; Berthiller, F.; Peltonen, K. Simultaneous determination of major type A and B trichothecenes, zearalenone and certain modified metabolites in Finnish cereal grains with a novel liquid chromatography-tandem mass spectrometric method. Anal. Bioanal. Chem. 2015, 407, 4745–4755. [Google Scholar] [CrossRef] [PubMed]
- Yoshinari, T.; Sakuda, S.; Furihata, K.; Furusawa, H.; Ohnishi, T.; Sugita-Konishi, Y.; Ishizaki, N.; Terajima, J. Structural determination of a nivalenol glucoside and development of an analytical method for the simultaneous determination of nivalenol and deoxynivalenol, and their glucosides, in wheat. J. Agric. Food Chem. 2014, 62, 1174–1180. [Google Scholar] [CrossRef] [PubMed]
- Veprikova, Z.; Vaclavikova, M.; Lacina, O.; Dzuman, Z.; Zachariasova, M.; Hajslova, J. Occurrence of mono-and di-glycosylated conjugates of T-2 and HT-2 toxins in naturally contaminated cereals. World Mycotoxin J. 2012, 5, 231–240. [Google Scholar] [CrossRef]
- Lattanzio, V.M.; Visconti, A.; Haidukowski, M.; Pascale, M. Identification and characterization of new Fusarium masked mycotoxins, T2 and HT2 glycosyl derivatives, in naturally contaminated wheat and oats by liquid chromatography-high-resolution mass spectrometry. J. Mass Spectrom. 2012, 47, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Boddu, J.; Cho, S.; Kruger, W.M.; Muehlbauer, G.J. Transcriptome analysis of the barley-Fusarium graminearum interaction. Mol. Plant Microbe Interact. 2006, 19, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Boddu, J.; Cho, S.; Muehlbauer, G.J. Transcriptome analysis of trichothecene-induced gene expression in barley. Mol. Plant Microbe Interact. 2007, 20, 1364–1375. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, S.A.; Boddu, J.; Berthiller, F.; Hametner, C.; Stupar, R.M.; Adam, G.; Muehlbauer, G.J. Transcriptome analysis of the barley—Deoxynivalenol interaction: Evidence for a role of glutathione in deoxynivalenol detoxification. Mol. Plant Microbe Interact. 2010, 23, 962–976. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, W.; Boddu, J.; Shin, S.; Poppenberger, B.; Berthiller, F.; Lemmens, M.; Muehlbauer, G.J.; Adam, G. Validation of a candidate deoxynivalenol-inactivating UDP-glucosyltransferase from barley by heterologous expression in yeast. Mol. Plant Microbe Interact. 2010, 23, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Michlmayr, H.; Schweiger, W.; Malachova, A.; Shin, S.; Huang, Y.; Dong, Y.; Wiesenberger, G.; McCormick, S.; Lemmens, M. A barley UDP-glucosyltransferase inactivates nivalenol and provides Fusarium Head Blight resistance in transgenic wheat. J. Exp. Bot. 2017, 68, 2187–2197. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shin, S.; Heinen, S.; Dill-Macky, R.; Berthiller, F.; Nersesian, N.; Clemente, T.; McCormick, S.; Muehlbauer, G.J. Transgenic wheat expressing a barley UDP-glucosyltransferase detoxifies deoxynivalenol and provides high levels of resistance to Fusarium graminearum. Mol. Plant Microbe Interact. 2015, 28, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Torres-Acosta, J.A.; Heinen, S.J.; McCormick, S.; Lemmens, M.; Paris, M.P.K.; Berthiller, F.; Adam, G.; Muehlbauer, G.J. Transgenic Arabidopsis thaliana expressing a barley UDP-glucosyltransferase exhibit resistance to the mycotoxin deoxynivalenol. J. Exp. Bot. 2012, 63, 4731–4740. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, W.; Pasquet, J.-C.; Nussbaumer, T.; Paris, M.P.K.; Wiesenberger, G.; Macadré, C.; Ametz, C.; Berthiller, F.; Lemmens, M.; Saindrenan, P. Functional characterization of two clusters of Brachypodium distachyon UDP-glycosyltransferases encoding putative deoxynivalenol detoxification genes. Mol. Plant Microbe Interact. 2013, 26, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.; Li, Y.; Lim, E.-K.; Bowles, D.J. Higher plant glycosyltransferases. Genome Biol. 2001, 2, REVIEWS3004. [Google Scholar] [CrossRef] [PubMed]
- Pasquet, J.-C.; Changenet, V.; Macadré, C.; Boex-Fontvieille, E.; Soulhat, C.; Bouchabké-Coussa, O.; Dalmais, M.; Atanasova-Pénichon, V.; Bendahmane, A.; Saindrenan, P. A Brachypodium UDP-glycosyltransferase confers root tolerance to deoxynivalenol and resistance to Fusarium infection. Plant Physiol. 2016, 172, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Michlmayr, H.; Malachová, A.; Varga, E.; Kleinová, J.; Lemmens, M.; Newmister, S.; Rayment, I.; Berthiller, F.; Adam, G. Biochemical Characterization of a Recombinant UDP-glucosyltransferase from Rice and Enzymatic Production of Deoxynivalenol-3-O-β-d-glucoside. Toxins 2015, 7, 2685–2700. [Google Scholar] [CrossRef] [PubMed]
- Wetterhorn, K.M.; Gabardi, K.; Michlmayr, H.; Malachová, A.; Busman, M.; McCormick, S.; Berthiller, F.; Adam, G.; Rayment, I. Determinants and Expansion of Specificity in a Trichothecene UDP-glucosyltransferase from Oryza sativa. Biochemistry 2017, 56, 6585–6596. [Google Scholar] [CrossRef] [PubMed]
- Wetterhorn, K.M.; Newmister, S.A.; Caniza, R.K.; Busman, M.; McCormick, S.P.; Berthiller, F.; Adam, G.; Rayment, I. Crystal structure of Os79 (Os04g0206600) from Oryza sativa: A UDP-glucosyltransferase involved in the detoxification of deoxynivalenol. Biochemistry 2016, 55, 6175–6186. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.K.; Bowles, D.J. A class of plant glycosyltransferases involved in cellular homeostasis. EMBO J. 2004, 23, 2915–2922. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Song, J.-L.; Zhang, S.; Wang, Y.; Cui, D.-F.; Wang, C.-C. Chaperone activity of DsbC. J. Biol. Chem. 1999, 274, 19601–19605. [Google Scholar] [CrossRef] [PubMed]
- Ilus, T.; Ward, P.J.; Nummi, M.; Adlercreutz, H.; Gripenberg, J. A new mycotoxin from Fusarium. Phytochemistry 1977, 16, 1839–1840. [Google Scholar] [CrossRef]
- McCormick, S.P.; Kato, T.; Maragos, C.M.; Busman, M.; Lattanzio, V.M.; Galaverna, G.; Dall-Asta, C.; Crich, D.; Price, N.P.; Kurtzman, C.P. Anomericity of T-2 toxin-glucoside: Masked mycotoxin in cereal crops. J. Agric. Food Chem. 2015, 63, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Meng-Reiterer, J.; Bueschl, C.; Rechthaler, J.; Berthiller, F.; Lemmens, M.; Schuhmacher, R. Metabolism of HT-2 Toxin and T-2 Toxin in Oats. Toxins 2016, 8, 364. [Google Scholar] [CrossRef] [PubMed]
- Meng-Reiterer, J.; Varga, E.; Nathanail, A.V.; Bueschl, C.; Rechthaler, J.; McCormick, S.P.; Michlmayr, H.; Malachová, A.; Fruhmann, P.; Adam, G. Tracing the metabolism of HT-2 toxin and T-2 toxin in barley by isotope-assisted untargeted screening and quantitative LC-HRMS analysis. Anal. Bioanal. Chem. 2015, 407, 8019–8033. [Google Scholar] [CrossRef] [PubMed]
- Nathanail, A.V.; Varga, E.; Meng-Reiterer, J.; Bueschl, C.; Michlmayr, H.; Malachova, A.; Fruhmann, P.; Jestoi, M.; Peltonen, K.; Adam, G. Metabolism of the Fusarium mycotoxins T-2 toxin and HT-2 toxin in wheat. J. Agric. Food Chem. 2015, 63, 7862–7872. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.W.; McCormick, S.P.; Alexander, N.J.; Proctor, R.H.; Desjardins, A.E. Inactivation of a cytochrome P-450 is a determinant of trichothecene diversity in Fusarium species. Fungal Genet. Biol. 2002, 36, 224–233. [Google Scholar] [CrossRef]
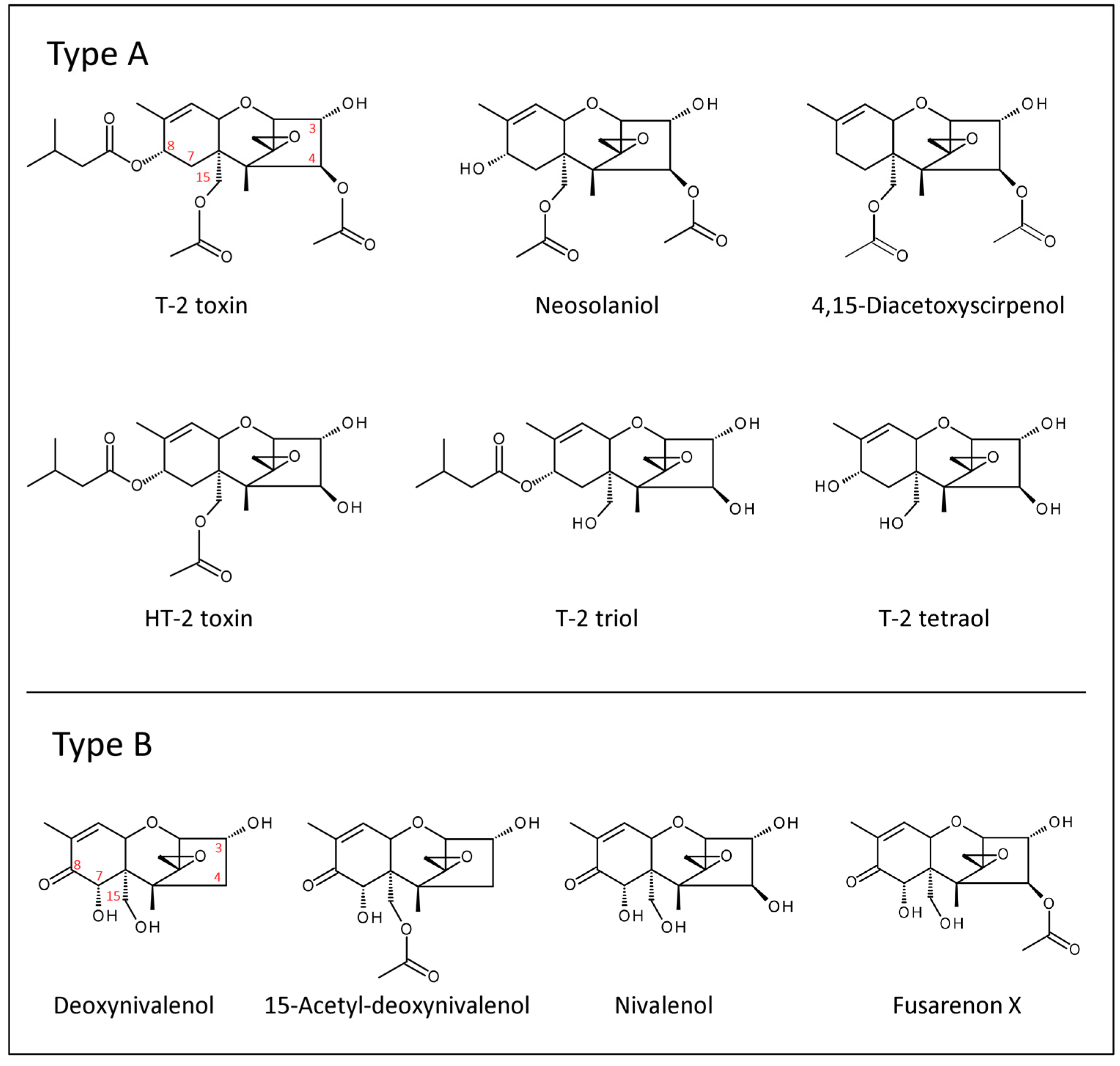

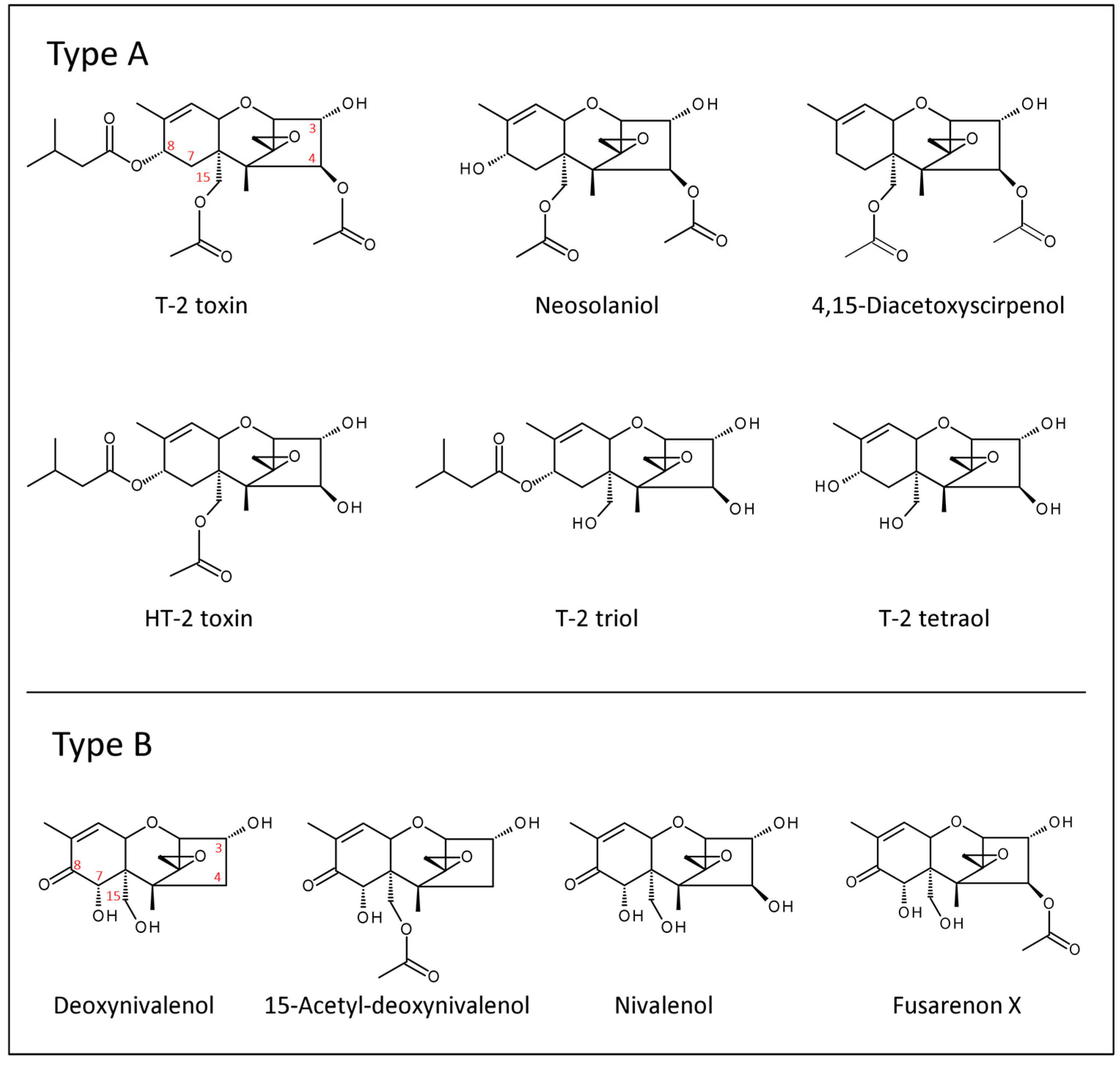{kind=link}
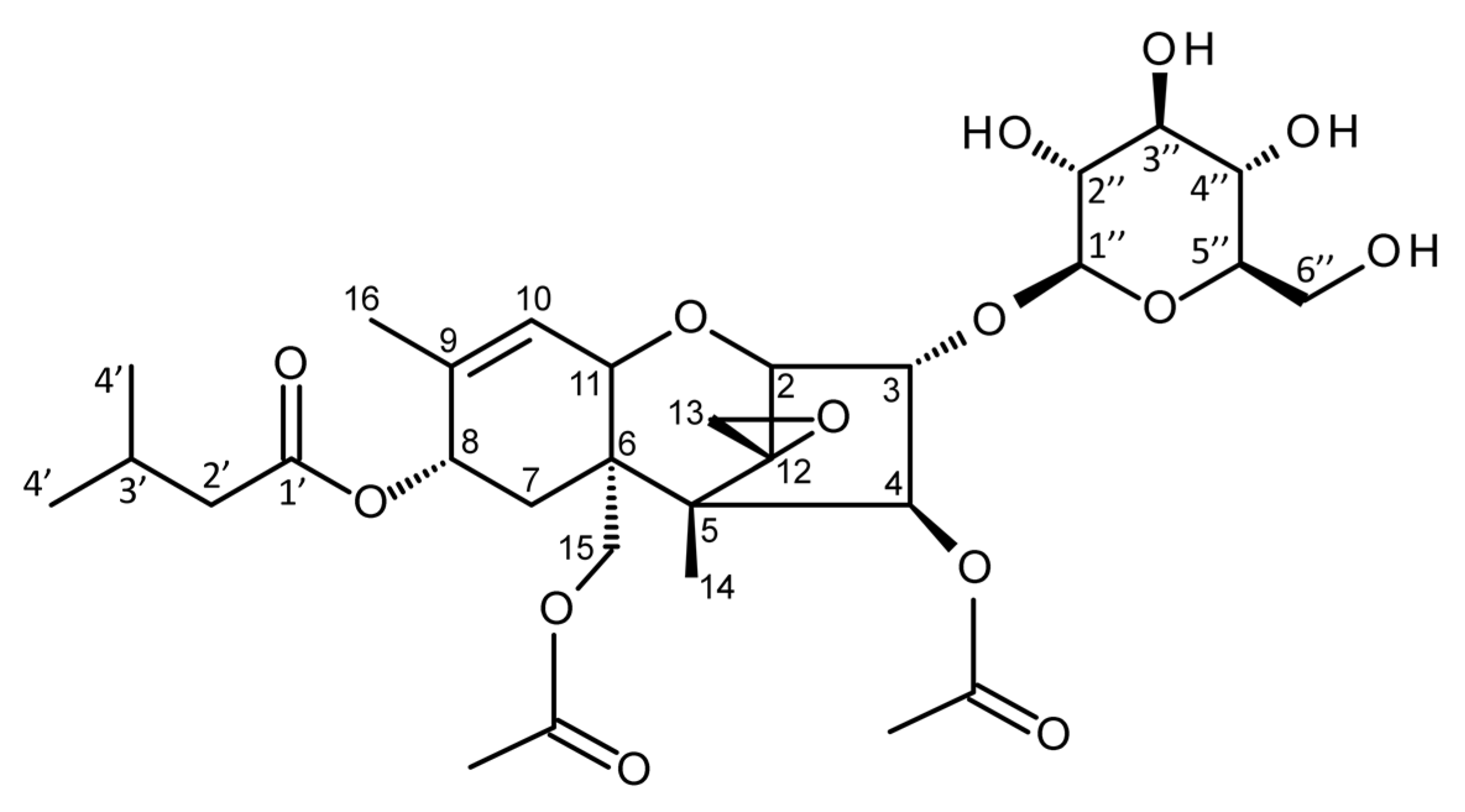
{kind=link}
| Substrate | Relative Activity (%) | ||
|---|---|---|---|
| OsUGT79 | HvUGT13248 | Bradi5g03300 | |
| Deoxynivalenol | 100 | 100 | 100 |
| 15-Acetyl-deoxynivalenol | 31 ± 3 | 424 ± 28 | 21 ± 5 |
| Nivalenol | 64 ± 3 | 360 ± 26 | 41 ± 6 |
| Fusarenon X | 4.6 ± 2.2 | 29 ± 3 | nd |
| T-2 toxin | nd | 21 ± 12 | nd |
| HT-2 toxin | 97 ± 16 | 1634 ± 177 | 1065 ± 68 |
| T-2 triol | 117 ± 21 | 1646 ± 124 | 1391 ± 69 |
| T-2 tetraol | 110 ± 16 | 382 ± 41 | 195 ± 6 |
| 4-Deoxy T-2 toxin | 63 ± 3 | 204 ± 6 | 169 ± 16 |
| Neosolaniol | nd | 19 ± 9 | nd |
| 4,15-Diacetoxyscirpenol | nd | 10 ± 3 | nd |
| Zearalenone | 89 ± 2 | 153 ± 4 | 20 ± 4 |
| Kaempferol | nd | 22 ± 3 | nd |
| Quercetin | nd | 7 ± 3 | nd |
| Enzyme | Substrate | KM (mM) | Vmax (µmol min−1 mg−1) | Vmax/KM |
|---|---|---|---|---|
| HvUGT13248 | Deoxynivalenol | 1.5 ± 0.3 | 0.22 ± 0.03 | 0.14 |
| Nivalenol | 0.64 ± 0.12 | 0.47 ± 0.05 | 0.73 | |
| HT-2 toxin | 0.053 ± 0.021 | 0.21 ± 0.02 | 4.0 | |
| Bradi5g03300 | Deoxynivalenol | 0.37 ± 0.05 | 0.019 ± 0.001 | 0.05 |
| Nivalenol | 1.3 ± 0.3 | 0.014 ± 0.001 | 0.01 | |
| HT-2 toxin | 0.12 ± 0.02 | 0.042 ± 0.004 | 0.35 |
| 15-Ac-DON | HT2 | HT2 | T2 triol | FUSX | T2 | DAS | NEO | |
|---|---|---|---|---|---|---|---|---|
| Total amount of toxin (mg) | 10 | 5 | 5 | 2.2 | 6 | 5 | 5 | 6 |
| Concentration toxin (mM) | 1.5 | 0.6 | 0.6 | 1.15 | 2.1 | 2.8 | 2.7 | 2.8 |
| UDP-glucose (mM) | 2.3 | 1 | 1.2 | 10 | 10 | 10 | 10 | 10 |
| Buffer (100 mM, pH 7) | Tris | Tris | Tris | Tris | KPP | KPP | KPP | KPP |
| Temperature (°C) | 25 | 25 | 25 | 37 | 25 | 25 | 25 | 25 |
| Reaction time (h) | 24 | 24 | 4 | 16 | 72 | 72 | 72 | 72 |
| UGT (mg/mL) | 1 (A) | 1 (A) | 1 (A) | 2 (A) | 1.7 (B) | 1.7 (B) | 3.2 (B) | 4 (B) |
| Sucrose (mM) | 50 | 50 | 50 | 50 | ||||
| Sucrose synthase (mg/mL) | 0.9 | 0.9 | 1.8 | 1.8 | ||||
| Final amount glucoside (mg) | not stable | 5.6 (C) | 7.0 | 1.9 | 2.2 | 3.4 | 5.9 | 1.5/0.4 (D) |
| Pos. | T-2 Toxin-3-O-β-d-glucoside in Methanol-d4 (A) | HT-2 Toxin-3-O-β-d-glucoside in Acetone-d6 | T-2 Triol-3-O-β-d-glucoside in Acetone-d6 | T-2 Tetraol-3-O-β-d-glucoside in Methanol-d4 | 4,15-Diacetoxyscirpenol-3-O-β-d-glucoside in Methanol-d4 | Neosolaniol-3-O-β-d-glucoside in Methanol-d4 | Iso-neosolaniol-3-O-β-d-glucoside in Methanol-d4 | Fusarenon X-3-O-β-d-glucoside in Acetonitrile-d3 |
|---|---|---|---|---|---|---|---|---|
| 2 | 3.72 (d, 5.0, 1H) | 3.54 (d, 4.7, 1H) | 3.48 (d, 4.9, 1H) | 3.57 (d, 4.8, 1H) | 3.70 (d, 4.8, 1H) | 3.69 (d, 5.0, 1H) | 3.68 (d, 5.0, 1H) | 3.81 (d, 4.8, 1H) |
| 3 | 4.48 (dd, 5.0, 3.1, 1H) | 4.31 (b, 1H) | 4.24 (dd, 4.9, 3.2, 1H) | 4.23 (dd, 4.8, 3.3, 1H) | 4.44 (dd, 4.8, 3.3 1H) | 4.46 (m, 1H) | 4.38 (dd, 5.0, 3.2, 1H) | 4.42 (dd, 4.8, 3.5, 1H) |
| 4 | 5.98 (d, 3.1, 1H) | 4.60 (b, 1H) | 4.83 (m, 1H) | 4.36 (d, 3.3, 1H) | 5.79 (d, 3.2, 1H) | 6.06 (d, 3.0, 1H) | 6.09 (d, 3.2, 1H) | 5.80 (d, 3.5, 1H) |
| 4-Ac | 2.09 (s, 3H) | - | - | - | 2.10 (s, 3H) | 2.09 (s, 3H) | 2.10 (s, 3H) (D) | 2.09 (s, 3H) |
| 7 | 2.38 (dd, 15.2, 5.8, 1H) 1.94 (bd, 15.2, 1H) | 2.35 (dd, 15.1, 5.6, 1H) 2.01 (bd, 15.0, 1H) | 2.26 (dd, 15.0, 5.8, 1H) 1.86 (d, 15.0, 1H) | 2.17 (dd, 14.3, 5.1, 1H) 1.99 (bd, 14.3, 1H) | 2.01 (m, 1H) 1.81 (m, 1H) | 2.25 (dd, 14.5, 5.6, 1H) 1.85 (bd, 14.5, 1H) | 2.24 (dd, 15.1, 5.9, 1H) 1.92 (bd, 15.1, 1H) | 4.79 (s, 1H) |
| 8 | 5.33 (d, 5.7, 1H) | 5.28 (d, 5.5, 1H) | 5.26 (d, 5.8, 1H) | 4.02 (bd, 4.9, 1H) | 2.05–1.90 (m, 2H) | 4.06 (d, 5.5, 1H) | 5.24 (d, 5.7, 1H) | - |
| 8-Ac | - | - | - | - | - | - | 2.05 (s, 3H) (D) | - |
| 10 | 5.75 (d, 5.9, 1H) | 5.69 (d, 5.5, 1H) | 5.66 (m, 1H) | 5.55 (bd, 5.6, 1H) | 5.48 (d, 5.5, H) | 5.60 (d, 5.8, 1H) | 5.77 (d, 6.0, 1H) | 6.58 (dd, 5.9, 1.5, 1H) |
| 11 | 4.38 (m, 1H) | 4.20 (d, 5.5, 1H) | 4.18 (d, 5.9, 1H) | 3.87 (bd, 5.6, 1H) | 4.15 (d, 5.5, 1H) | 4.38 (d, 5.8, 1H) | 4.31 (d, 6.0, 1H) | 4.67 (d, 5.9, 1H) |
| 13 | 3.04 (d, 3.8, 1H) 2.87 (d, 3.8, 1H) | 2.95 (d, 4.0, 1H) 2.80 (d, 4.0, 1H) | 2.89 (d, 4.2, 1H) 2.75 (d, 4.2, 1H) | 2.93 (d, 4.1, 1H) 2.80 (d, 4.1, 1H) | 3.02 (d, 3.9, 1H) 2.85 (d, 3.9, 1H) | 3.03 (d, 4.0, 1H) 2.85 (d, 4.0, 1H) | 3.02 (d, 4.0, 1H) 2.85 (d, 4.0, 1H) | 3.05 (d, 4.2, 1H) 2.98 (d, 4.2, 1H) |
| 14 | 0.74 (s, 3H) | 0.83 (s, 3H) | 0.84 (s, 3H) | 0.86 (s, 3H) | 0.75 (s, 3H) | 0.78 (s, 3H) | 0.79 (s, 3H) | 0.94 (s, 3H) |
| 15 | 4.38 (d, 12.6, 1H) 4.09 (d, 12.6, 1H) | 4.24 (d, 12.2, 1H) 3.95 (d, 12.2, 1H) | 3.84 (d, 12.1, 1H) 3.53 (d, 12.0, 1H) | 3.74 (d, 12.4, 1H) 3.41 (d, 12.4, 1H) | 4.31 (d, 12.3, 1H) 4.04 (d, 12.3, 1H) | 4.39 (d, 12.5, 1H) 4.20 (d, 12.5, 1H) | 3.97 (d, 12.2, 1H) 3.56 (d, 12.2, 1H) | 3.89 (d, 12.3, 1H) 3.66 (d, 12.3, 1H) |
| 15-Ac | 2.06 (s, 3H) | 2.03 (s, 3H) | - | - | 2.05 (s, 3H) | 2.07 (s, 3H) | - | - |
| 16 | 1.75 (s, 3H) | 1.72 (s, 3H) | 1.70 (s, 3H) | 1.83 (s, 3H) | 1.73 (s, 3H) | 1.84 (s, 3H) | 1.76 (s, 3H) | 1.82 (s, 3H) |
| 2′ | 2.15 (m, 2H) | 2.15 (d, 7.0, 2H) | 2.21 (d, 6.9, 2H) | - | - | - | - | - |
| 3′ | ca. 2.06 (m, 1H) | ca. 2.05 (1H) (B) | 2.06 (m, 1H) | - | - | - | - | - |
| 4′ | 0.97 (d, 6.6, 3H) 0.96 (d, 6.6, 3H) | 0.97 (d, 6.4, 3H) 0.96 (d, 6.4, 3H) | 0.95 (d, 6.7, 3H) 0.94 (d, 6.7, 3H) | - | - | - | - | - |
| 1″ | 4.44 (d, 7.8, 1H) | 4.72 (d, 7.9, 1H) | 4.66 (d, 7.9, 1H) | 4.57 (d, 7.9, 1H) | 4.42 (d, 7.9, 1H) | 4.45 (d, 7.8, 1H) | 4.45 (d, 7.8, 1H) | 4.39 (d, 7.8, 1H) |
| 2″ | 3.21 (m, 1H) | 3.29 (t, 8.2, 1H) | 3.26 (dd, 8.6, 7.9, 1H) | 3.25 (b, 1H) | 3.24 (dd, 8.9, 7.9, 1H) | 3.25 (dd, 9.0, 7.9, 1H) | 3.25 (m, 1H) | 3.16 (m, 1H) |
| 3″ | 3.35 (t, 9.0, 1H) | 3.43 (t, 8.8, 1H) | 3.42 (t, 8.7, 1H) | 3.37 (bt, 7.2, 2H) (C) | 3.35 (t, 8.9, 1H) | 3.35 (t, 9.0, 1H) | 3.35 (m, 1H) | 3.30 (m, 1H) |
| 4″ | ca. 3.26 (m, 1H) | 3.40 (t, 9.1, 1H) | 3.37 (dd, 9.3, 8.7, 1H) | 3.37 (bt, 7.2, 2H) (C) | 3.31 (1H) (B) | 3.30 (t, 9.0, 1H) | 3.30 (m, 1H) | 3.25 (m, 1H) |
| 5″ | 3.21 (ddd, 9.7, 5.7, 2.2, 1H) | 3.34 (b, 1H) | 3.32 (dd, 9.4, 5.4, 1H) | 3.27 (b, 1H) | 3.21 (ddd, 9.5, 5.6, 2.2, 1H) | 3.21 (ddd, 9.5, 5.7, 2.2, 1H) | 3.22 (m, 1H) | 3.25 (m, 1H) |
| 6″ | 3.83 (dd, 12.1, 2.2, 1H) 3.65 (dd, 12.1, 5.7, 1H) | 3.84 (bd, 10.6, 1H) 3.70 (m, 1H) | 3.82 (m, 1H) 3.66 (m, 1H) | 3.83 (dd, 12.1, 2.0, 1H) 3.68 (dd, 12.1, 5.0, 1H) | 3.83 (dd,12.0, 2.3, 1H) 3.65 (dd, 12.0, 5.6, 1H) | 3.83 (dd, 12.0, 2.2, 1H) 3.65 (dd, 12.0, 5.7, 1H) | 3.82 (dd, 12.2, 2.4, 1H) 3.65 (m, 1H) | 3.6 (m, 2H) |
| Pos. | T-2 Toxin-3-O-β-d-glucoside in Methanol-d4 (A) | HT-2 Toxin-3-O-β-d-glucoside in Acetone-d6 | T-2 Triol-3-O-β-d-glucoside in Acetone-d6 | T-2 Tetraol-3-O-β-d-glucoside in Methanol-d4 | 4,15-Diacetoxyscirpenol-3-O-β-d-glucoside in Methanol-d4 | Neosolaniol-3-O-β-d-glucoside in Methanol-d4 | Iso-neosolaniol-3-O-β-d-glucoside in Methanol-d4 | Fusarenon X-3-O-β-d-glucoside in Acetonitrile-d3 |
|---|---|---|---|---|---|---|---|---|
| 2 | 80.6 | 78.8 | 79.9 | 80.4 (B) | 80.8 | 80.6 | 80.7 | 80.7 |
| 3 | 84.0 | 85.3 | 86.7 | 87.7 | 84.4 | 84.1 | 84.8 | 83.3 |
| 4 | 81.3 | 78.8 | 79.7 | 80.5 (B) | 81.8 | 81.3 | 81.8 | 80.9 |
| 4-Ac | 172.3 & 20.9 | - | - | - | 172.5 & 21.0 | 172.3 & 21.0 | 172.6 & 21.0 | 172.0 & 21.2 |
| 5 | 50.2 | 48.6 | 49.4 | 50.2 | 50.4 | 50.2 | 49.9 | 50.1 |
| 6 | 44.5 | 42.6 | 45.1 | 46.2 | 45.6 | 44.9 | 45.9 | 54.4 |
| 7 | 28.9 | 27.2 | 28.3 | 30.0 | 22.3 | 31.8 | 28.4 | 74.8 |
| 8 | 69.5 | 68.0 | 69.2 | 67.0 | 29.0 | 67.2 | 70.1 | 200.4 |
| 8-Ac | - | - | - | - | - | - | 172.6 & 21.3 | - |
| 9 | 137.5 | 134.8 | 135.6 | 141.5 | 141.9 | 141.6 | 137.6 | 136.7 |
| 10 | 125.2 | 124.8 | 126.3 | 122.6 | 119.8 | 121.9 | 125.4 | 138.9 |
| 11 | 68.6 | 67.0 | 68.2 | 70.2 | 69.5 | 69.1 | 69.2 | 70.5 |
| 12 | 65.4 | 64.2 | 65.5 | 65.7 | 65.4 | 65.6 | 65.7 | 65.4 |
| 13 | 48.0 | 45.9 | 46.9 | 47.5 | 47.9 | 48.1 | 48.1 | 47.0 |
| 14 | 7.2 | 6.6 | 7.4 | 7.4 | 7.2 | 7.2 | 7.2 | 8.0 |
| 15 | 65.9 | 64.2 | 63.5 | 63.2 | 64.8 | 66.2 | 63.7 | 61.4 |
| 15-Ac | 172.4 & 21.4 | 169.6 & 20.3 | - | - | 172.6 & 21.1 | 172.8 & 21.3 | - | - |
| 16 | 20.6 | 19.5 | 20.4 | 20.9 | 23.4 | 21.0 | 20.5 | 15.4 |
| 1′ | 174.1 | 171.7 | 172.7 | - | - | - | - | - |
| 2′ | 44.7 | 43.1 | 44.1 | - | - | - | - | - |
| 3′ | 27.1 | 25.5 | 26.4 | - | - | - | - | - |
| 4′ | 22.9 | 21.8 & 21.7 | 22.7 | - | - | - | - | - |
| 1″ | 103.9 | 102.4 | 103.5 | 104.4 | 104.3 | 103.9 | 104.3 | 103.3 |
| 2″ | 74.9 | 73.6 | 74.6 | 75.0 | 74.9 | 74.9 | 75.1 | 74.4 |
| 3″ | 78.2 | 77.1 | 78.1 | 78.3 (C) | 78.2 | 78.2 | 78.1 | 77.5 |
| 4″ | 71.6 | 70.7 | 71.7 | 71.4 | 71.5 | 71.5 | 71.5 | 71.4 |
| 5″ | 78.4 | 76.8 | 77.7 | 78.3 (C) | 78.4 | 78.4 | 78.4 | 77.5 |
| 6″ | 62.8 | 61.9 | 62.9 | 62.6 | 62.7 | 62.7 | 62.7 | 62.7 |
| Compound Name | Q1 (m/z) and Ion Species | DP (V) | Q3 (m/z) | CE (eV) |
|---|---|---|---|---|
| 15-acetyl-deoxynivalenol | 339.1 [M + H]+ | 86 | 261.0/321.0 | 15/11 |
| 15-acetyl-deoxynivalenol-glucoside | 518.0 [M + NH4]+ | 56 | 339.0/321.1 | 19/27 |
| 4,15-diacetoxyscirpenol | 384.2 [M + NH4]+ | 51 | 307.2/105.1 | 17/61 |
| 4,15-diacetoxyscirpenol-3-O-β-d-glucoside | 546.0 [M + NH4]+ | 66 | 307.2/105.0 | 21/83 |
| fusarenon X | 413.3 [M + CH3COO]− | −40 | 59.1/262.9 | −44/−22 |
| fusarenon X-3-O-β-d-glucoside | 561.1 [M + CH3COO]− | −80 | 515.1/244.8 | −22/−34 |
| HT-2 toxin | 442.2 [M + NH4]+ | 70 | 215.1/197.1 | 19/25 |
| HT-2 toxin-3-O-β-d-glucoside | 604.4 [M + NH4]+ | 51 | 263.3/215.1 | 27/25 |
| neosolaniol | 400.2 [M + NH4]+ | 46 | 185.0/215.0 | 25/29 |
| neosolaniol-3-O-β-d-glucoside | 562.2 [M + NH4]+ | 71 | 305.0/185.0 | 25/39 |
| iso-neosolaniol-3-O-β-d-glucoside | 562.2 [M + NH4]+ | 66 | 215.2/202.9 | 35/27 |
| T-2 tetraol | 316.2 [M + NH4]+ | 31 | 215.3/281.4 | 13/25 |
| T-2 tetraol-3-O-β-d-glucoside | 478.3 [M+NH4]+ | 46 | 215.1/233.2 | 21/15 |
| T-2 triol | 400.2 [M + NH4]+ | 41 | 215.2/281.3 | 17/13 |
| T-2 triol-3-O-β-d-glucoside | 562.3 [M + NH4]+ | 41 | 215.5/233.5 | 20/10 |
| T-2 toxin | 484.3 [M + NH4]+ | 56 | 215.2/185.1 | 29/31 |
| T-2 toxin-3-O-β-d-glucoside | 646.3 [M + NH4]+ | 66 | 305.0/215.0 | 30/35 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michlmayr, H.; Varga, E.; Malachová, A.; Fruhmann, P.; Piątkowska, M.; Hametner, C.; Šofrová, J.; Jaunecker, G.; Häubl, G.; Lemmens, M.; et al. UDP-Glucosyltransferases from Rice, Brachypodium, and Barley: Substrate Specificities and Synthesis of Type A and B Trichothecene-3-O-β-d-glucosides. Toxins 2018, 10, 111. https://doi.org/10.3390/toxins10030111
Michlmayr H, Varga E, Malachová A, Fruhmann P, Piątkowska M, Hametner C, Šofrová J, Jaunecker G, Häubl G, Lemmens M, et al. UDP-Glucosyltransferases from Rice, Brachypodium, and Barley: Substrate Specificities and Synthesis of Type A and B Trichothecene-3-O-β-d-glucosides. Toxins. 2018; 10(3):111. https://doi.org/10.3390/toxins10030111
Chicago/Turabian StyleMichlmayr, Herbert, Elisabeth Varga, Alexandra Malachová, Philipp Fruhmann, Marta Piątkowska, Christian Hametner, Jana Šofrová, Günther Jaunecker, Georg Häubl, Marc Lemmens, and et al. 2018. "UDP-Glucosyltransferases from Rice, Brachypodium, and Barley: Substrate Specificities and Synthesis of Type A and B Trichothecene-3-O-β-d-glucosides" Toxins 10, no. 3: 111. https://doi.org/10.3390/toxins10030111
APA StyleMichlmayr, H., Varga, E., Malachová, A., Fruhmann, P., Piątkowska, M., Hametner, C., Šofrová, J., Jaunecker, G., Häubl, G., Lemmens, M., Berthiller, F., & Adam, G. (2018). UDP-Glucosyltransferases from Rice, Brachypodium, and Barley: Substrate Specificities and Synthesis of Type A and B Trichothecene-3-O-β-d-glucosides. Toxins, 10(3), 111. https://doi.org/10.3390/toxins10030111