The Impact of Diet Wheat Source on the Onset of Type 1 Diabetes Mellitus—Lessons Learned from the Non-Obese Diabetic (NOD) Mouse Model



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Wheat Varieties
2.3. Diets
2.4. Experimental Design
2.5. Statistical Analysis
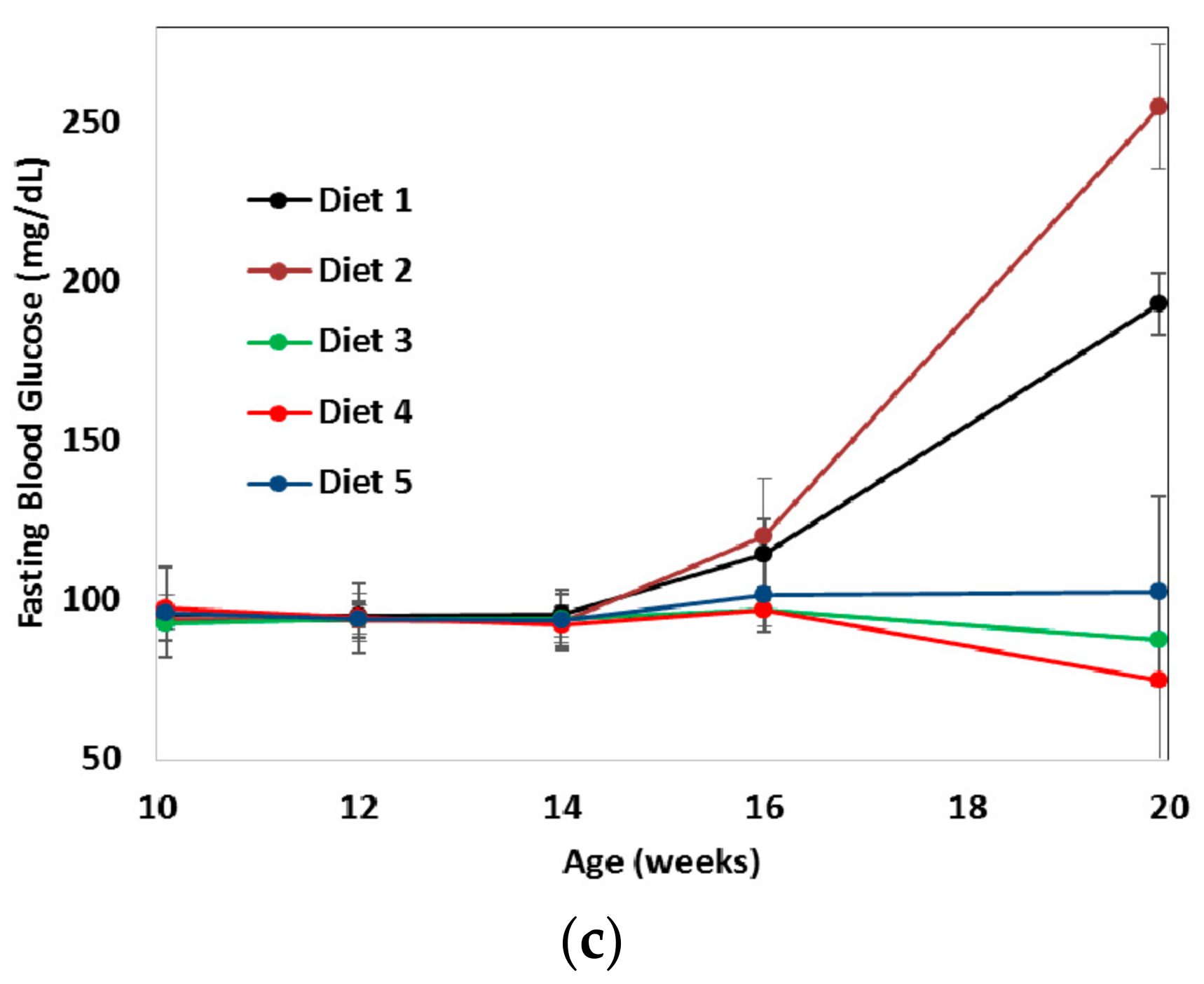
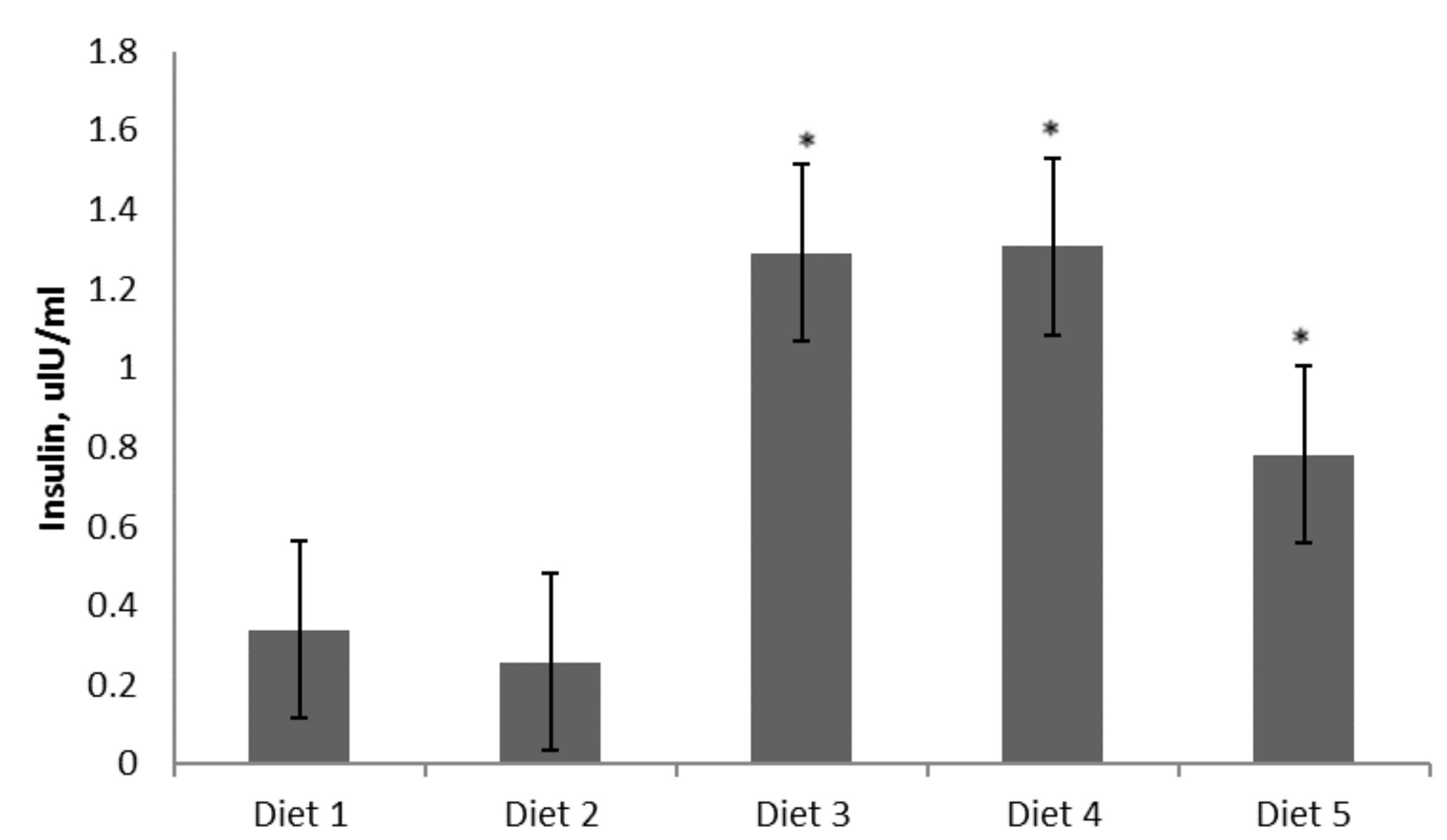
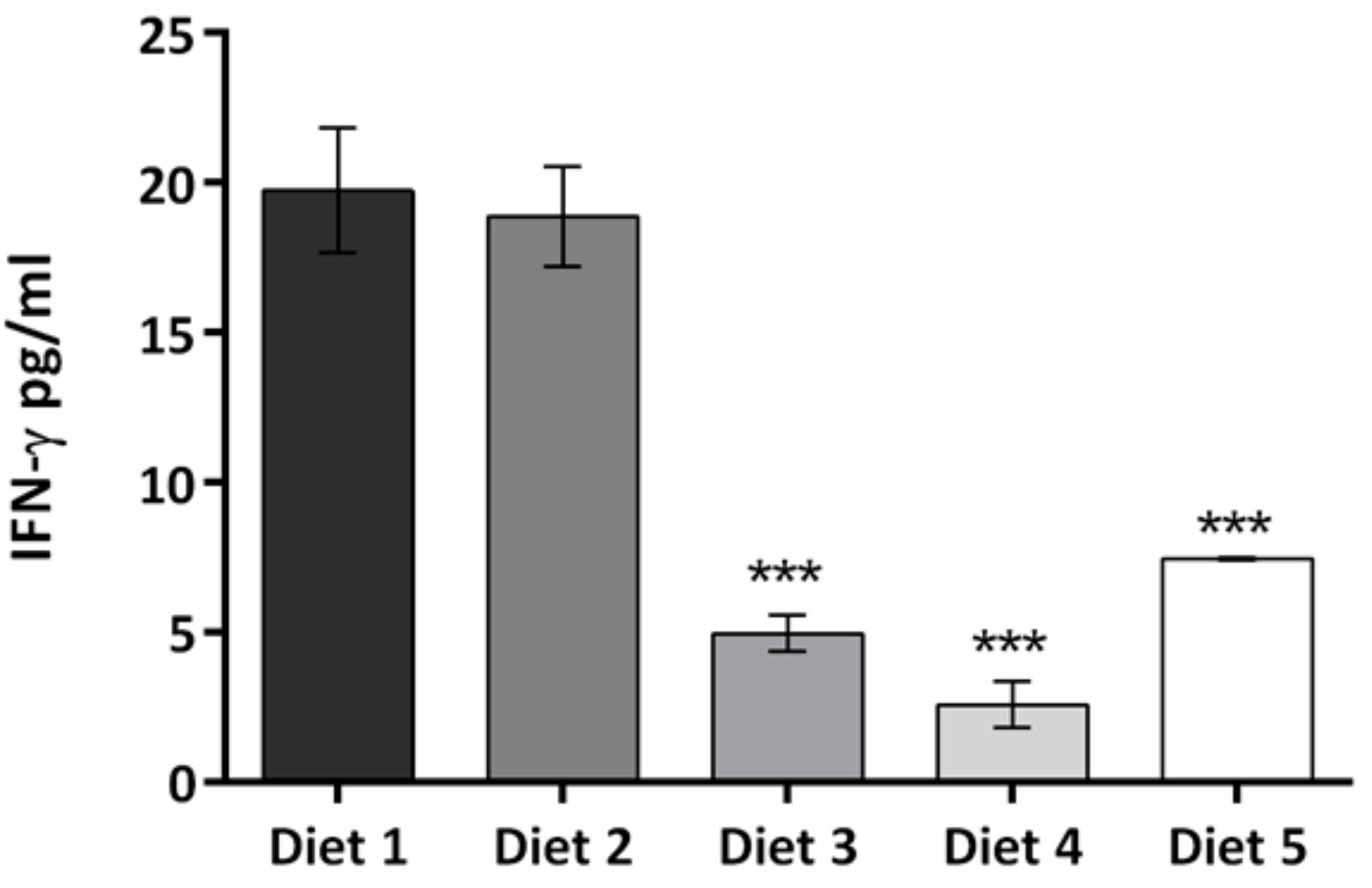
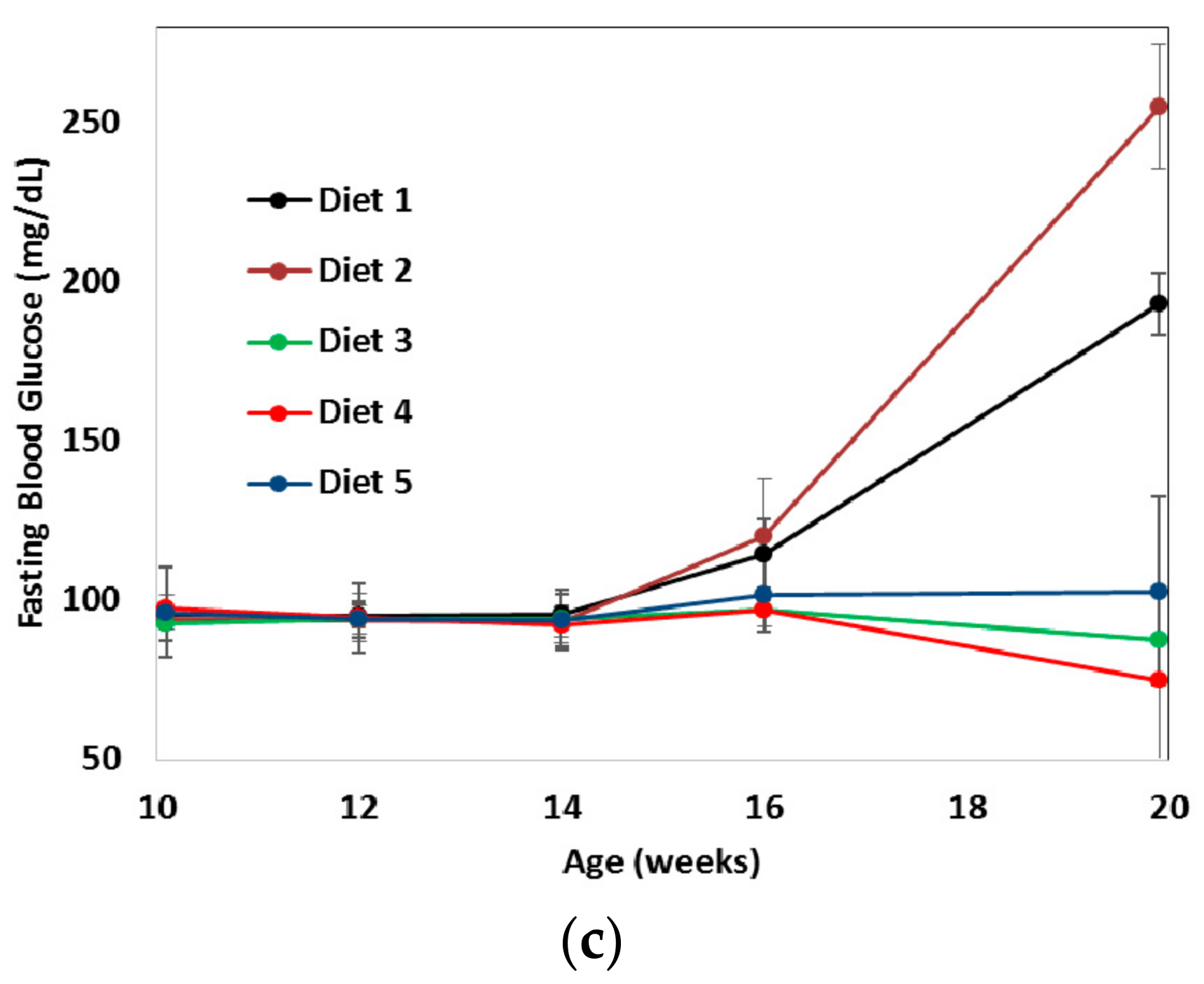
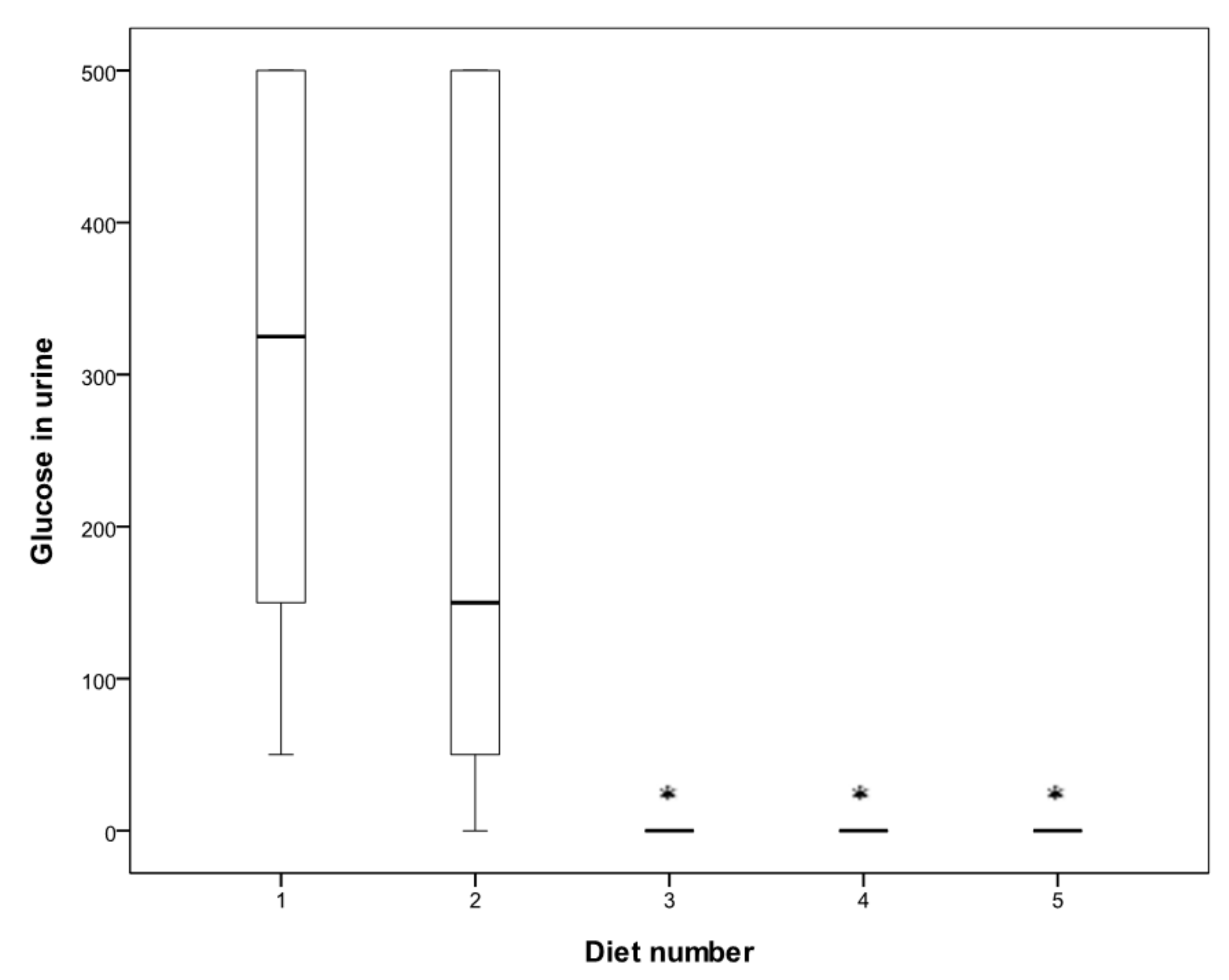
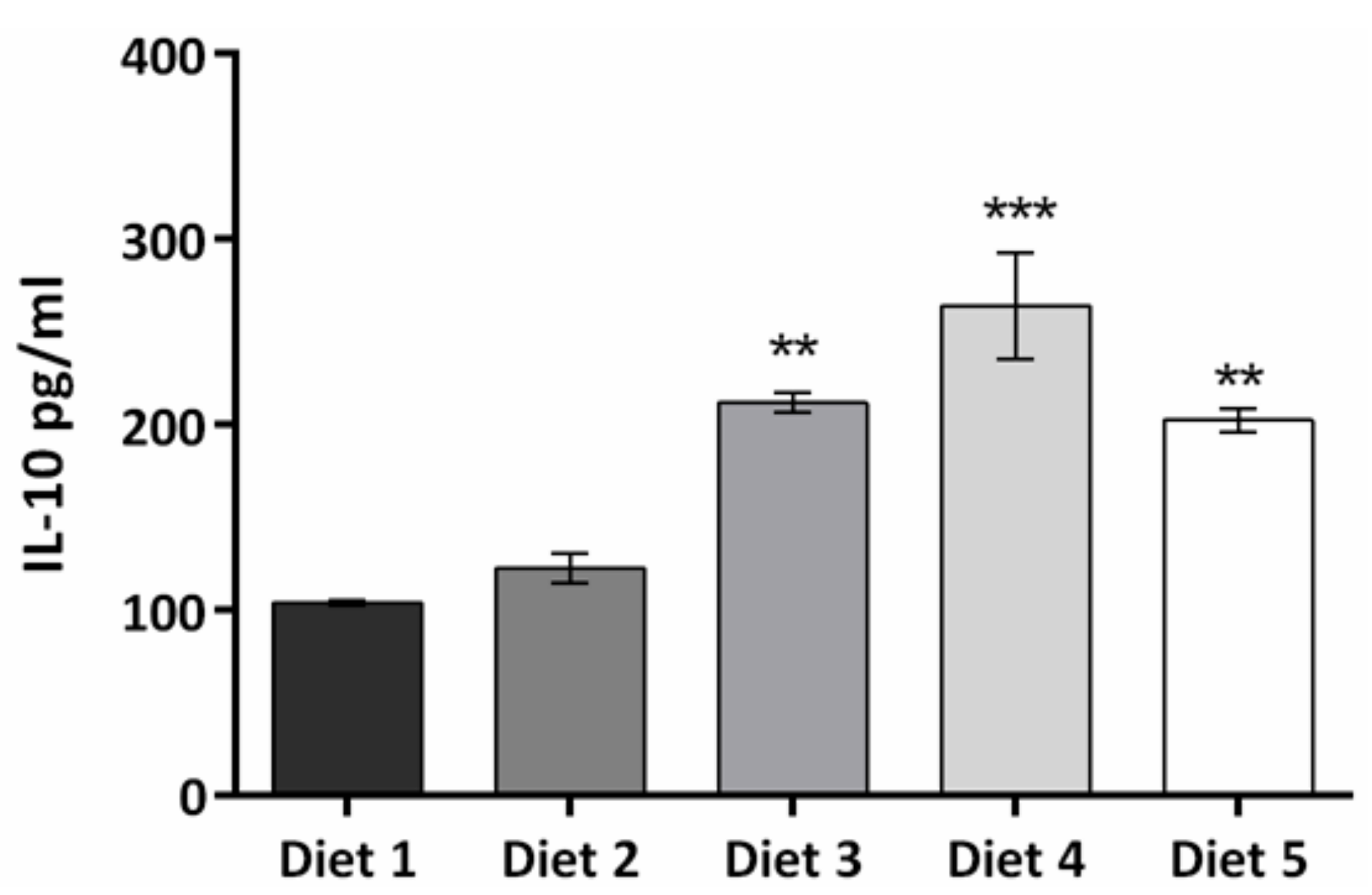
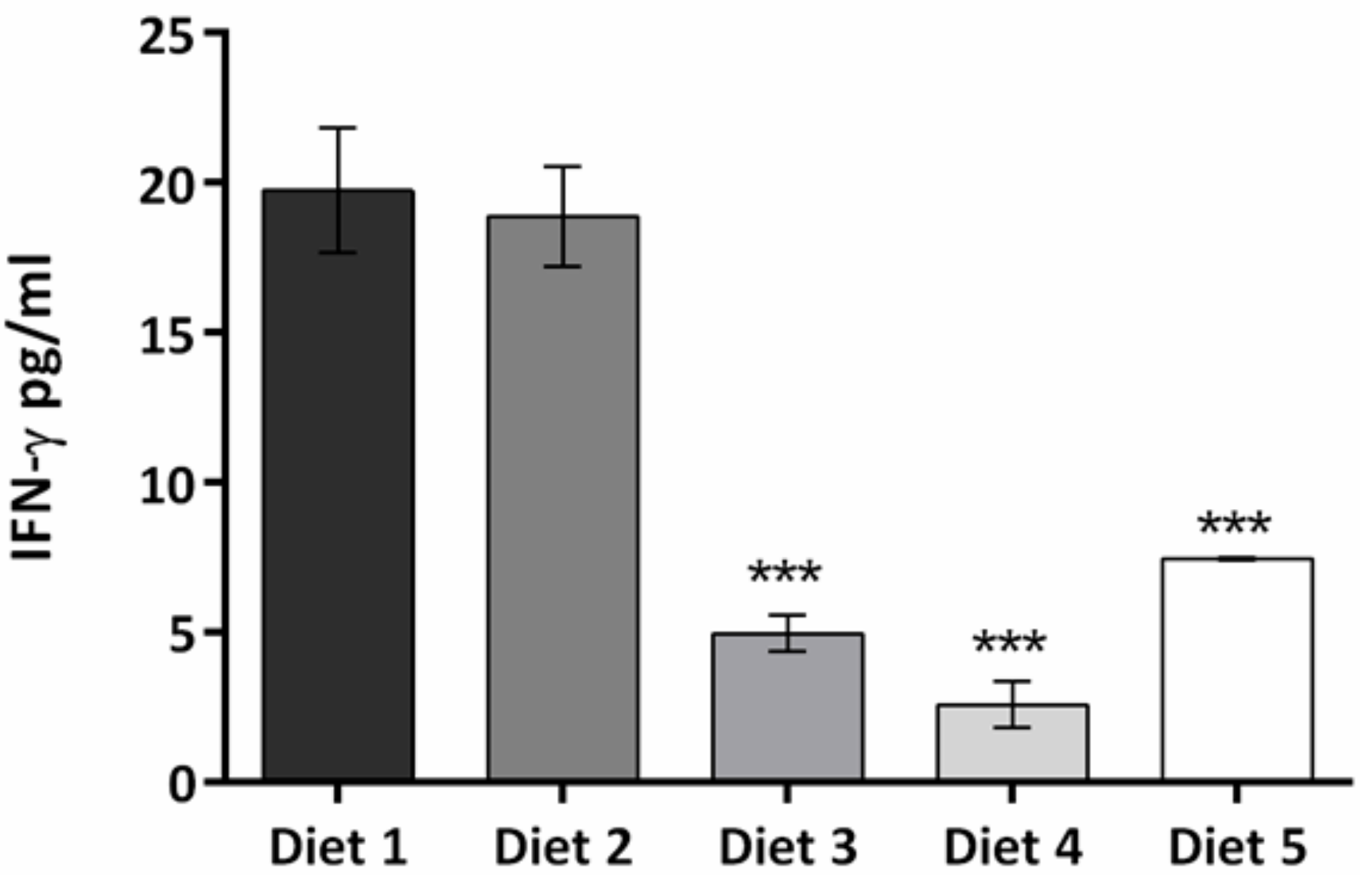
3. Results
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Shalitin, S.; Peter Chase, H. Diabetes technology and treatments in the pediatric age group. Int. J. Clin. Pract. 2011, 65 (Suppl. 170), 76–82. [Google Scholar] [CrossRef] [PubMed]
- Funda, D.; Kaas, A.; Bock, T.; Tlaskalová-Hogenová, H.; Buschard, K. Gluten-free diet prevents diabetes in NOD mice. Diabetes Metab. Res. Rev. 1999, 15, 323–327. [Google Scholar] [CrossRef]
- Landin-Olsson, M.; Hillman, M.; Erlanson-Albertsson, C. Is type 1 diabetes a food-induced disease? Med. Hypotheses 2013, 81 (Suppl. 2), 338–342. [Google Scholar] [CrossRef] [PubMed]
- Antvorskov, J.C.; Josefsen, K.; Engkilde, K.; Funda, D.P.; Buschard, K. Dietary gluten and the development of type 1 diabetes. Diabetologia 2014, 57 (Suppl. 9), 1770–1780. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, D.S.; Krych, L.; Buschaard, K.; Hansen, C.H.; Hansen, A.K. Beyond genetics. Influence of dietary factors and gut microbiota on type 1 diabetes. FEBS Lett. 2014, 588, 4234–4243. [Google Scholar] [CrossRef] [PubMed]
- Elliott, R.B.; Harris, D.P.; Hill, J.P.; Bibby, N.J.; Wasmuth, H.E. Type 1 (insulin-dependent) diabetes mellitus and cow milk: Casein variant consumption. Diabetologia 1999, 42, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Merriman, T.R. Type 1 diabetes, the A1 milk hypothesis and vitamin D deficiency. Diabetes Res. Clin. Prac. 2009, 83, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Schmid, S.; Koczwara, K.; Schwinghammer, S.; Lampasona, V.; Ziegler, A.G.; Bonifacio, E. Delayed exposure to wheat and barley proteins reduces diabetes incidence in non-obese diabetic mice. Clin. Immunol. 2004, 111, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Maurano, F.; Mazzarella, G.; Luongo, D.; Stefanile, R.; D’Arienzo, R.; Rossi, M.; Troncone, R. Small intestinal enteropathy in nonobese diabetic mice fed a diet containing wheat. Diabetologia 2005, 48, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Wolta, U.; Tovoli, F.; Caio, G. Clinical and immunological features of celiac disease in patients with type 1 diabetes mellitus. Expert Rev. Gastroenterol. Hepatol. 2011, 5 (Suppl. 4), 479–487. [Google Scholar]
- Adlercreutz, E.H.; Wingren, C.J.; Vincente, R.P.; Merlo, J.; Agardh, D. Perinatal risk factors increase the risk of being affected by both type 1 diabetes and coeliacdisease. Acta Paediatr. 2015, 104 (Suppl. 2), 178–184. [Google Scholar] [CrossRef] [PubMed]
- Mollazadegan, K.; Sanders, D.S.; Ludvigsson, J.; Ludvigsson, J.F. Long-term coeliac disease influences risk of death in patients with type 1 diabetes. J. Intern. Med. 2013, 274 (Suppl. 3), 273–280. [Google Scholar] [CrossRef] [PubMed]
- Scaramuzza, A.E.; Mantegazza, C.; Bosetti, A.; Zuccotti, G.V. Type 1 diabetes and celiac disease: The effects of gluten free diet on metabolic control. World J. Diabetes 2013, 4 (Suppl. 4), 130–134. [Google Scholar] [CrossRef] [PubMed]
- Salgin, B.; Meissner, T.; Beyer, P.; Haberland, H.; Borkenstein, M.; Fussenegger, J.; Holl, R.W. Lipoatrophy is associated with an increased risk of Hashimoto’s thyroiditis and coeliac disease in female patients with type 1 diabetes. Horm. Res. Paediatr. 2013, 79 (Suppl. 6), 368–372. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, F.H.; Murray, J.A.; Kudva, Y.C.; Zinsmeister, A.R.; Dierkhising, R.A.; Lahr, B.D.; Van Dyke, C.T. Celiac disease in type 1 diabetes mellitus in a North American community: Prevalence, serologic screening, and clinical features. Mayo Clin. Proc. 2005, 80 (Suppl. 11), 1429–1435. [Google Scholar] [CrossRef] [PubMed]
- Ludvigsson, J.F.; Ludvigsson, J.; Ekbom, A.; Montgomery, S.M. Celiac disease and risk of subsequent type 1 diabetes: A general population cohort study of children and adolescents. Diabetes Care 2006, 29 (Suppl. 11), 2483–2488. [Google Scholar] [CrossRef] [PubMed]
- Goh, C.; Banerjee, K. Prevalence of coeliac disease in children and adolescents with type 1 diabetes mellitus in a clinic based population. Postgrad. Med. J. 2007, 83 (Suppl. 976), 132–136. [Google Scholar] [CrossRef] [PubMed]
- Simmons, J.H.; Klingensmith, G.J.; McFann, K.; Rewers, M.; Taylor, J.; Emery, L.M.; Hoffenberg, E.J. Impact of celiac autoimmunity on children with type 1 diabetes. J. Pediatr. 2007, 150 (Suppl. 5), 461–466. [Google Scholar] [CrossRef] [PubMed]
- Pham-Short, A.; Donaghue, K.C.; Ambler, G.; Chan, A.K.; Craig, M.E. Coeliac disease in Type 1 diabetes from 1990 to 2009: Higher incidence in young children afterlonger diabetes duration. Diabet. Med. 2012, 29 (Suppl. 9), 286–289. [Google Scholar] [CrossRef] [PubMed]
- Levy-Shraga, Y.; Lerner-Geva, L.; Boyko, V.; Graph-Barel, C.; Mazor-Aronovitch, K.; Modan-Moses, D.; Pinhas-Hamiel, O. Type 1 diabetes in pre-school children—Long-term metabolic control, associated autoimmunity and complications. Diabet. Med. 2012, 29 (Suppl. 10), 1291–1296. [Google Scholar] [CrossRef] [PubMed]
- Van den Broeck, H.; de Jong, H.C.; Salentijn, E.M.; Dekking, L.; Bosch, D.; Hamer, R.J.; Smulders, M.J. Presence of celiac disease epitopes in modern and old hexaploid wheat varieties: Wheat breeding may have contributed to increased prevalence of celiac disease. Theor. Appl. Genet. 2010, 121 (Suppl. 8), 1527–1539. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, A.G.; Schmid, S.; Huber, D.; Hummel, M.; Bonifacio, E. Early infant feeding and risk of developing type 1 diabetes-associated autoantibodies. JAMA 2003, 290, 1721–1728. [Google Scholar] [CrossRef] [PubMed]
- Norris, J.M.; Barriga, K.; Klingensmith, G.; Hoffman, M.; Eisenbarth, G.S.; Erlich, H.A.; Rewers, M. Timing of initial cereal exposure in infancy and exposure in infancy and risk of islet autoimmunity. JAMA 2003, 290, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Funda, D.; Kaas, A.; Tlaskalova-Hogenova, H.; Buschard, K. Gluten-free but also gluten-enriched (gluten+) diet prevent diabetes in NOD mice; the gluten enigma in type 1 diabetes. Diabetes Metab. Res. Rev. 2008, 24, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Van den Broeck, H.; Hongbing, C.; Lacaze, X.; Dusautoir, J.C.; Gilissen, L.; Smulders, M.; Van Der Meer, I. In search of tetraploid wheat accessions reduced in celiac disease-related gluten epitopes. Mol. Biosyst. 2010, 6 (Suppl. 11), 2206–2213. [Google Scholar] [CrossRef] [PubMed]
- Wieser, H. Chemistry of gluten proteins. Food Microbiol. 2007, 24 (Suppl. 2), 115–119. [Google Scholar] [CrossRef] [PubMed]
- Poliarkova, H.; Blum, A. Land-races of wheat from the Northen Negev in Israel. Euphitica 1983, 32, 257–271. [Google Scholar] [CrossRef]
- Fedak, G. Alien Introgressions from wild Triticum species, T. monococcum, T. urartu, T. turgidum, T. dicoccum, T. dicoccoides, T. carthlicum, T. araraticum, T. timopheevii, and T. miguschovae. In Alien Introgression in Wheat; Springer: Cham, Switzerland, 2015; pp. 191–219. [Google Scholar]
- Peng, J.H.; Fahima, T.; Röder, M.S.; Li, Y.C.; Dahan, A.; Grama, A.; Ronin, Y.I.; Korol, A.B.; Nevo, E. Microsatellite tagging of the stripe-rust resistance gene YrH52 derived from wild emmer wheat, Triticum dicoccoides, and suggestive negative crossover interference on chromosome 1B. Theor. Appl. Genet. 1999, 98, 862–872. [Google Scholar] [CrossRef]
- King, A.J. The use of animal models in diabetes research. Br. J. Pharm. 2012, 166, 877–894. [Google Scholar] [CrossRef] [PubMed]
- Yi, Z.; Li, L.; Garland, A.; He, Q.; Wang, H.; Katz, J.D.; Tisch, R.; Wang, B. IFN-γ receptor deficiency prevents diabetes induction by diabetogenic CD4+, but not CD8+, T cells. Eur. J. Immunol. 2012, 42, 2010–2018. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.A.; Morgan, N.G. The impact of anti-inflammatory cytokines on the pancreatic β-cell. Islets 2014, 6, e950547. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Sanchez, J.P.; Cabrera-Chavez, F.; Calderon de la Barca, A.M.C. Maize Prolamins could induce a gluten-like cellular immune response in some celiac disease patients. Nutrients 2013, 5, 4174–4183. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, A.J.; Burghardt, K.M.; Kelly, J.; Simell, T.; Simell, O.; Altosaar, I.; Scott, F.W. A type 1 diabetes-related protein from wheat (Triticum aestivum). cDNA clone of a wheat storage globulin, Glb1, linked to islet damage. J. Biol. Chem. 2003, 278 (Suppl. 1), 54–63. [Google Scholar] [CrossRef] [PubMed]
- Loit, E.; Melnyk, C.W.; MacFarlane, A.J.; Scott, F.W.; Altosaar, I. Identification of three wheat globulin genes by screening a Triticum aestivum BAC genomic library with cDNA from a diabetes-associated globulin. BMC Plant Biol. 2009, 9, 93. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets Composition | Diet 1 | Diet 2 | Diet 3 | Diet 4 | Diet 5 |
|---|---|---|---|---|---|
| Maize | 35.00% | 21.06% | 21.06% | 27.85% | 21.06% |
| Wheat | 0 | 20.00% | 20.00% | 20.00% | 20.00% |
| Poultry Meat | 15.15% | 13.01% | 13.01% | 10.00% | 13.01% |
| Corn Gluten Meal | 5.27% | 5.17% | 5.17% | 4.19% | 5.17% |
| Sorghum | 15.18% | 14.92% | 14.92% | 11.63% | 14.92% |
| Soya Oil | 1.62% | 1.00% | 1.00% | 1.46% | 1.00% |
| Sunflower Meal | 15.00% | 15.00% | 15.00% | 15.00% | 15.00% |
| Tomato Fibers | 10.00% | 6.84% | 6.84% | 6.68% | 6.84% |
| ALIMET | 0.04% | 0.04% | 0.04% | 0.03% | 0.04% |
| L-Lys-Cl | 0.40% | 0.47% | 0.47% | 0.50% | 0.47% |
| NaCl | 0.25% | 0.27% | 0.27% | 0.29% | 0.27% |
| Vit. Conc. BR | 0.25% | 0.25% | 0.25% | 0.25% | 0.25% |
| Limestone | 1.20% | 1.24% | 1.24% | 1.26% | 1.24% |
| Na2SO4 | 0.10% | 0.10% | 0.10% | 0.10% | 0.10% |
| Dicalcium Phosphate | 0.54% | 0.62% | 0.62% | 0.75% | 0.62% |
| 100% | 100% | 100% | 100% | 100% |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorelick, J.; Yarmolinsky, L.; Budovsky, A.; Khalfin, B.; Klein, J.D.; Pinchasov, Y.; Bushuev, M.A.; Rudchenko, T.; Ben-Shabat, S. The Impact of Diet Wheat Source on the Onset of Type 1 Diabetes Mellitus—Lessons Learned from the Non-Obese Diabetic (NOD) Mouse Model. Nutrients 2017, 9, 482. https://doi.org/10.3390/nu9050482
Gorelick J, Yarmolinsky L, Budovsky A, Khalfin B, Klein JD, Pinchasov Y, Bushuev MA, Rudchenko T, Ben-Shabat S. The Impact of Diet Wheat Source on the Onset of Type 1 Diabetes Mellitus—Lessons Learned from the Non-Obese Diabetic (NOD) Mouse Model. Nutrients. 2017; 9(5):482. https://doi.org/10.3390/nu9050482
Chicago/Turabian StyleGorelick, Jonathan, Ludmila Yarmolinsky, Arie Budovsky, Boris Khalfin, Joshua D. Klein, Yosi Pinchasov, Maxim A. Bushuev, Tatiana Rudchenko, and Shimon Ben-Shabat. 2017. "The Impact of Diet Wheat Source on the Onset of Type 1 Diabetes Mellitus—Lessons Learned from the Non-Obese Diabetic (NOD) Mouse Model" Nutrients 9, no. 5: 482. https://doi.org/10.3390/nu9050482
APA StyleGorelick, J., Yarmolinsky, L., Budovsky, A., Khalfin, B., Klein, J. D., Pinchasov, Y., Bushuev, M. A., Rudchenko, T., & Ben-Shabat, S. (2017). The Impact of Diet Wheat Source on the Onset of Type 1 Diabetes Mellitus—Lessons Learned from the Non-Obese Diabetic (NOD) Mouse Model. Nutrients, 9(5), 482. https://doi.org/10.3390/nu9050482