Walnut Polyphenol Extract Attenuates Immunotoxicity Induced by 4-Pentylphenol and 3-methyl-4-nitrophenol in Murine Splenic Lymphocyte

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Animals
2.3. Preparation of WPE
2.4. Preparation of Splenocytes
2.5. Cell Viability Assay
2.6. Cell Staining/Flow Cytometry for Phenotypic Analysis
2.7. Measurement of Cytokine/Granzyme Production and Determination of ·OH, SOD, GSH-Px, and MDA Levels
2.8. Determination of the Total Phenolic Content
2.9. LC-MS Analyses (HPLC-ESI-IT-TOF-MS)
2.10. Statistical Analysis
3. Results
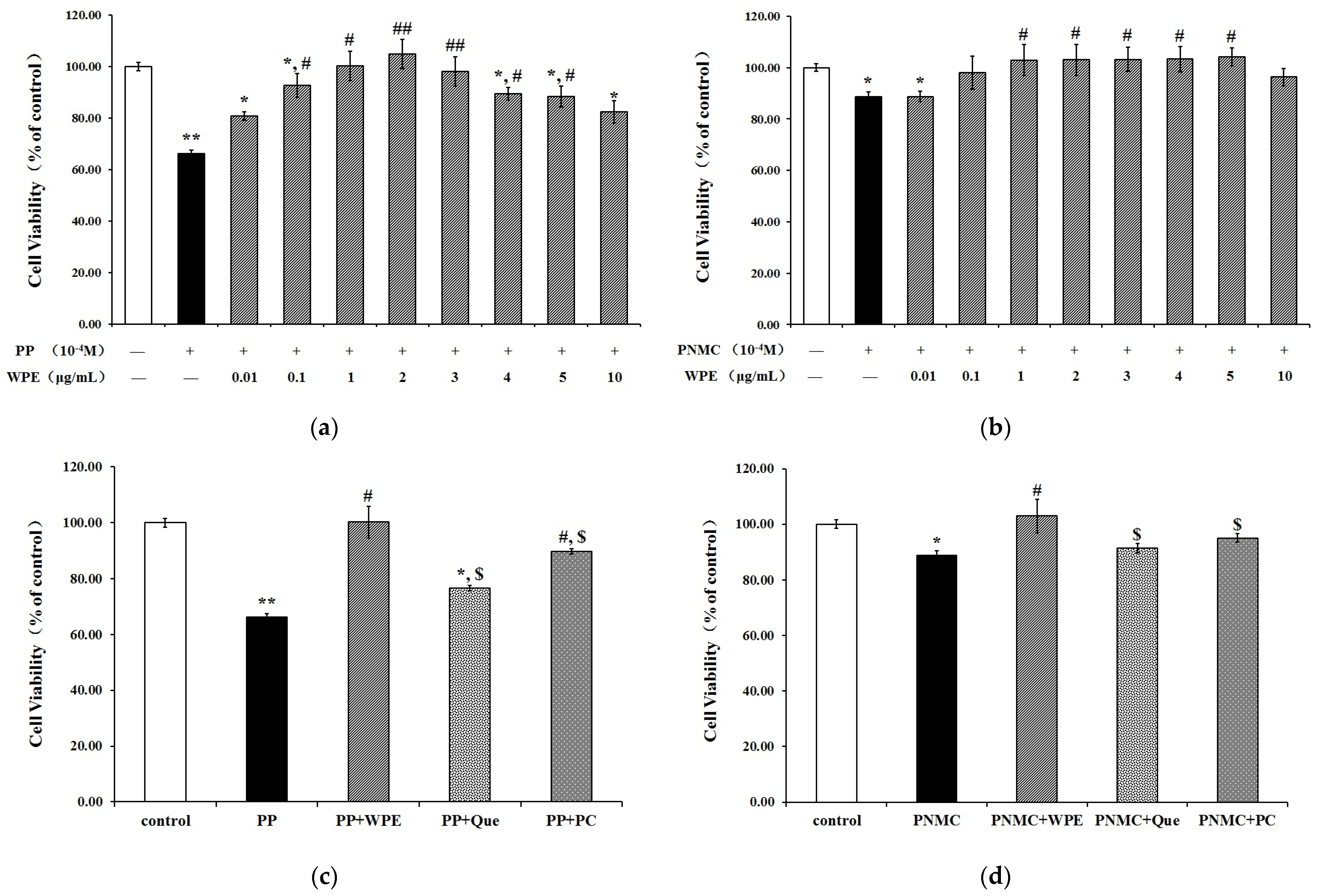
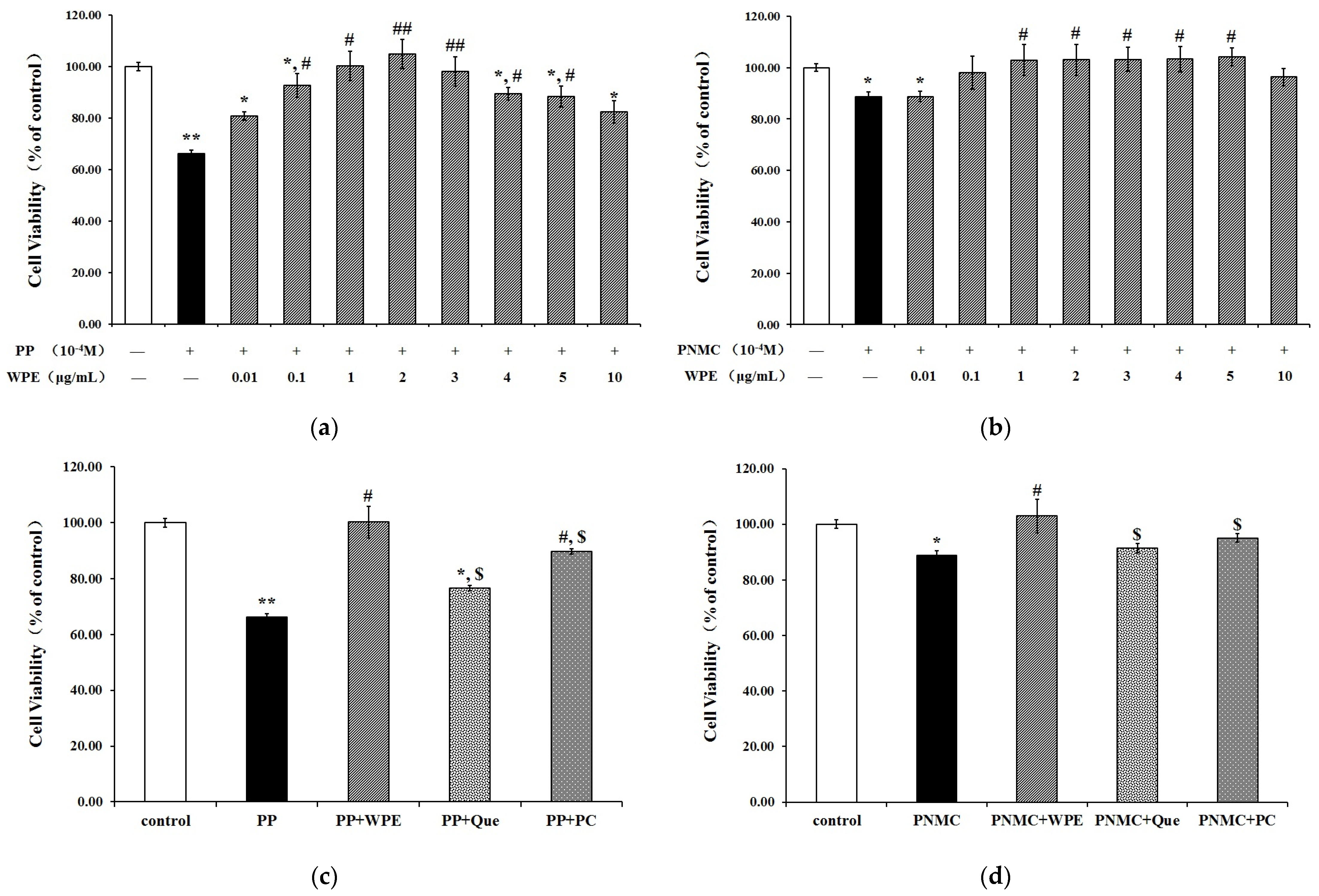
3.1. WPE Attenuates Cytotoxicity in Splenocytes
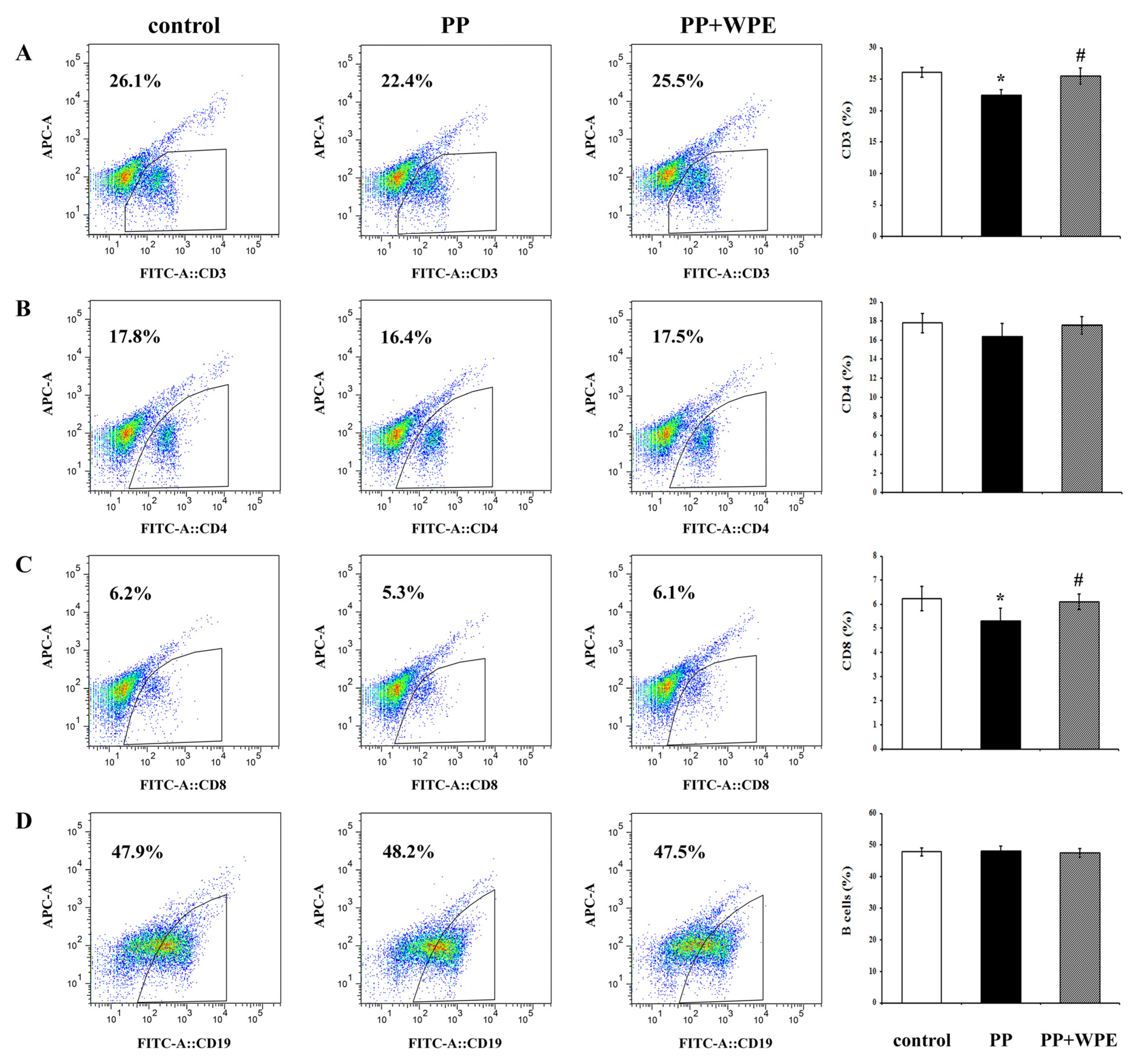
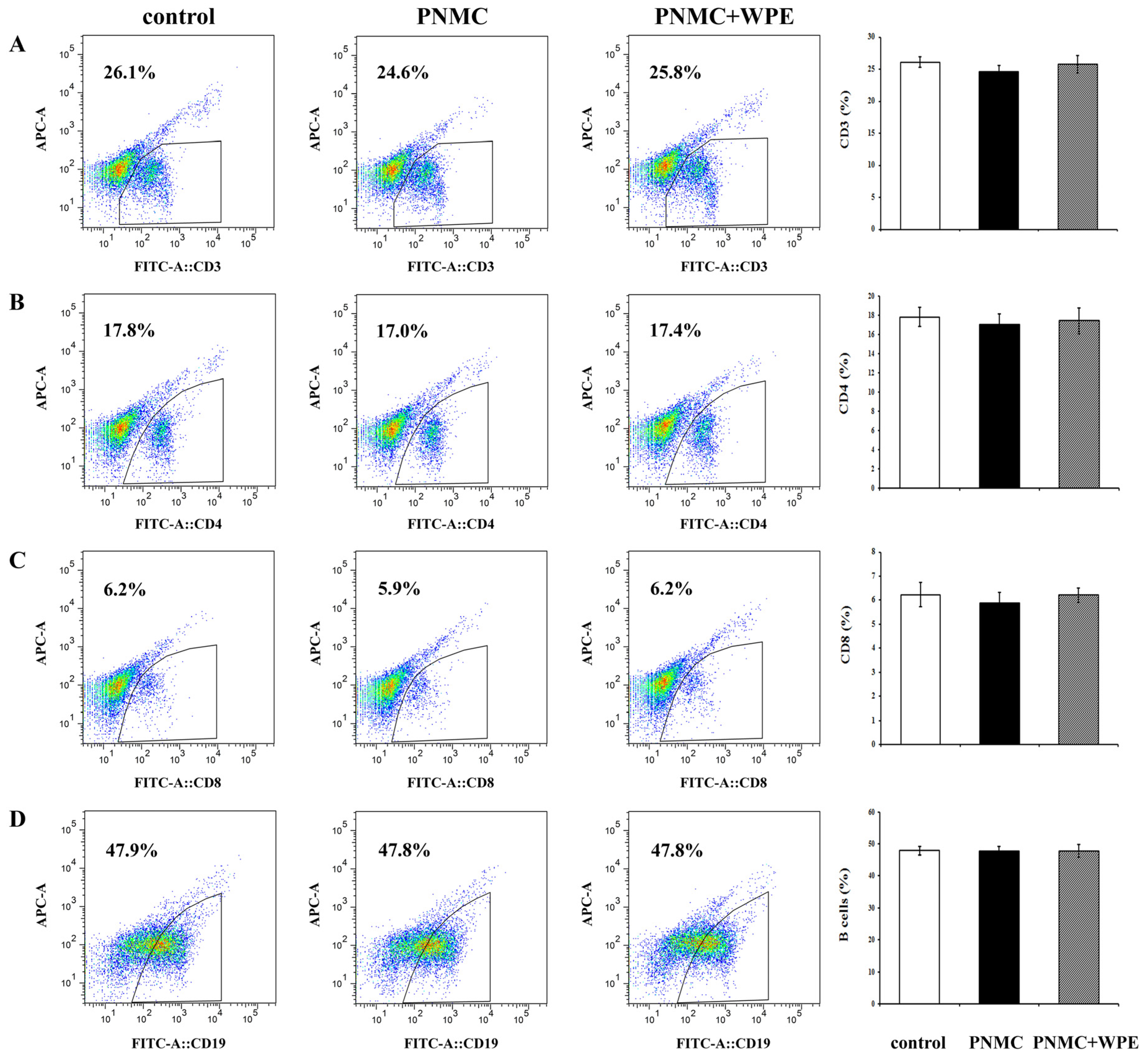
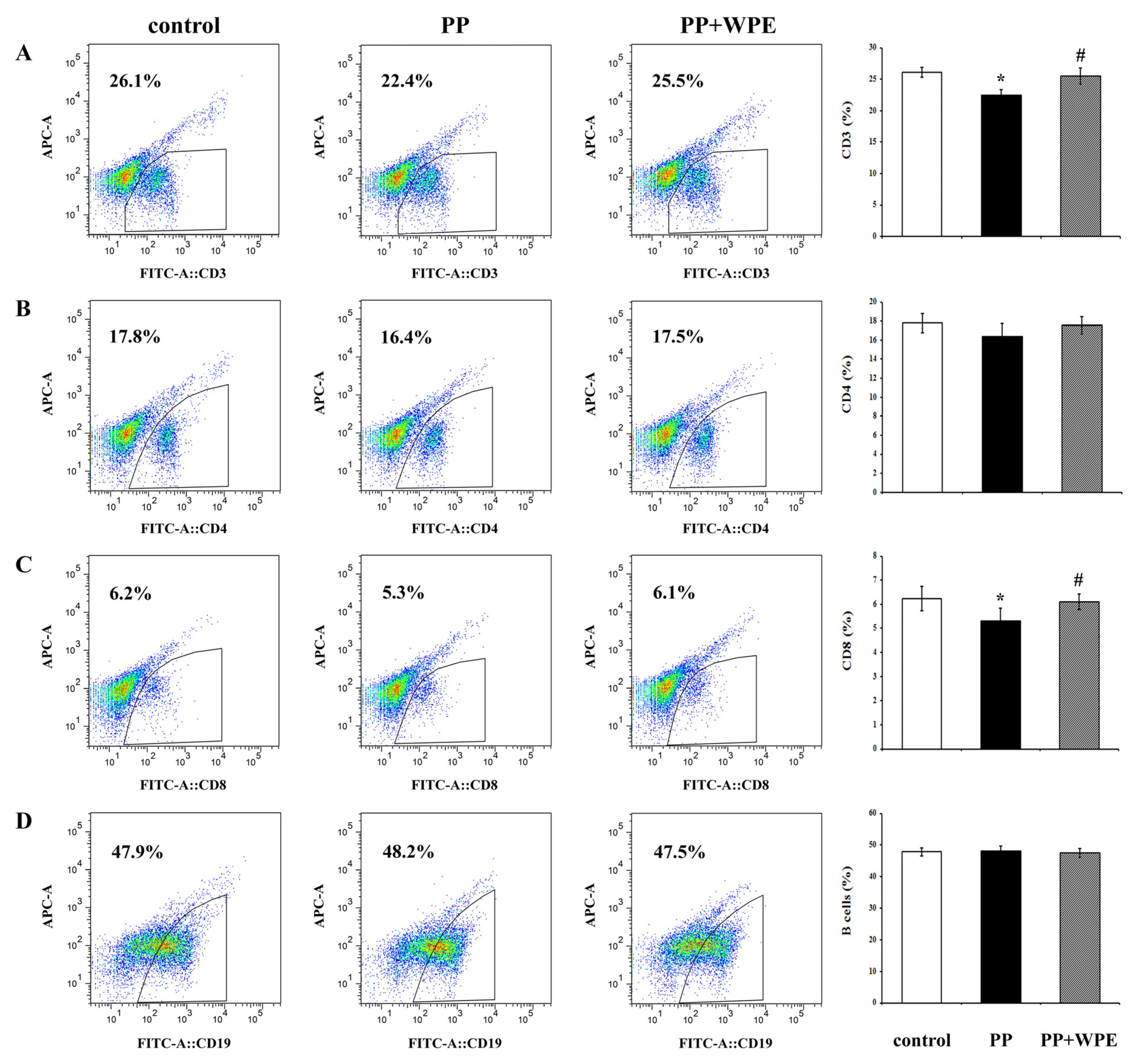
3.2. WPE Inhibited Decreases in Splenic T Cell Sub-Populations
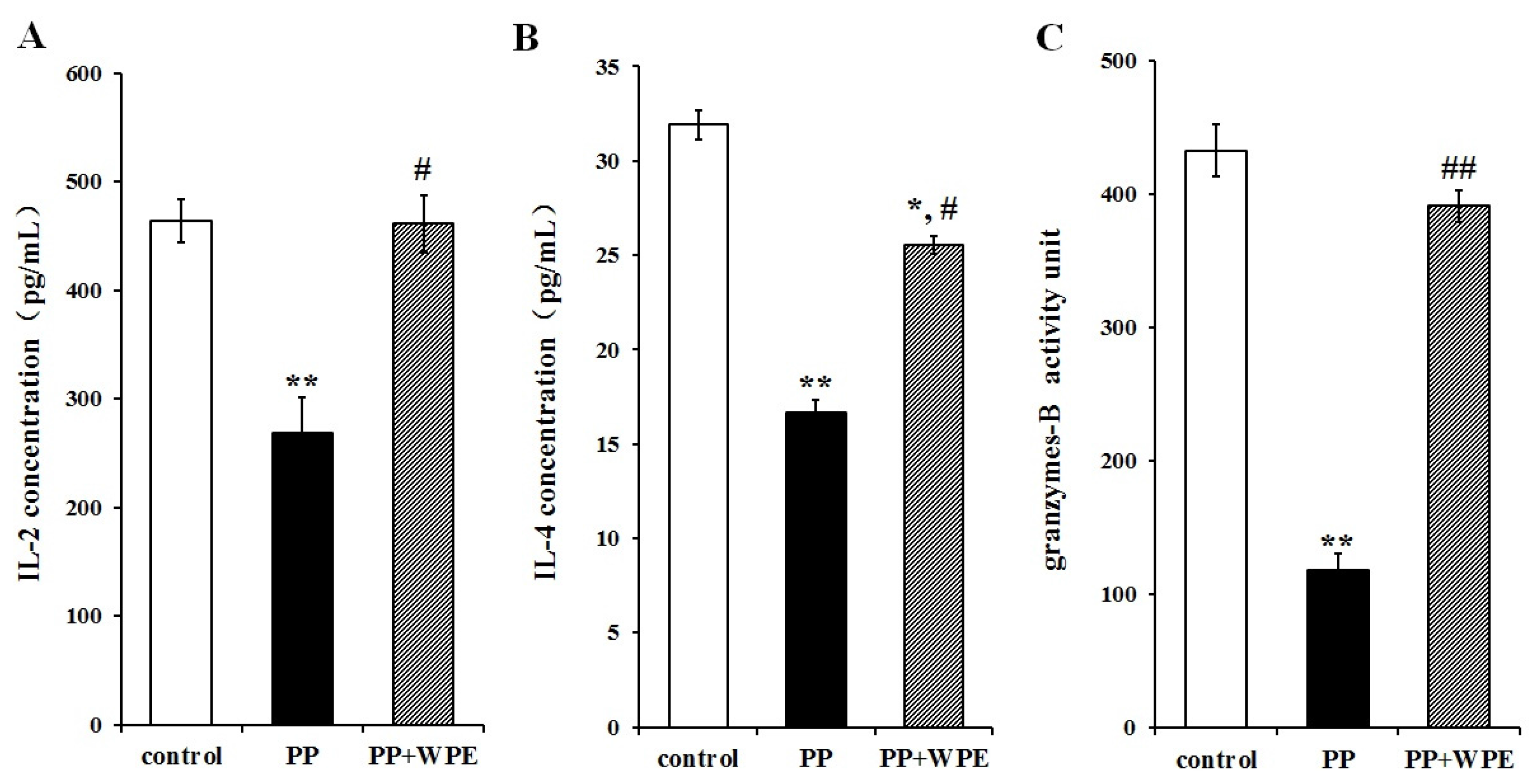
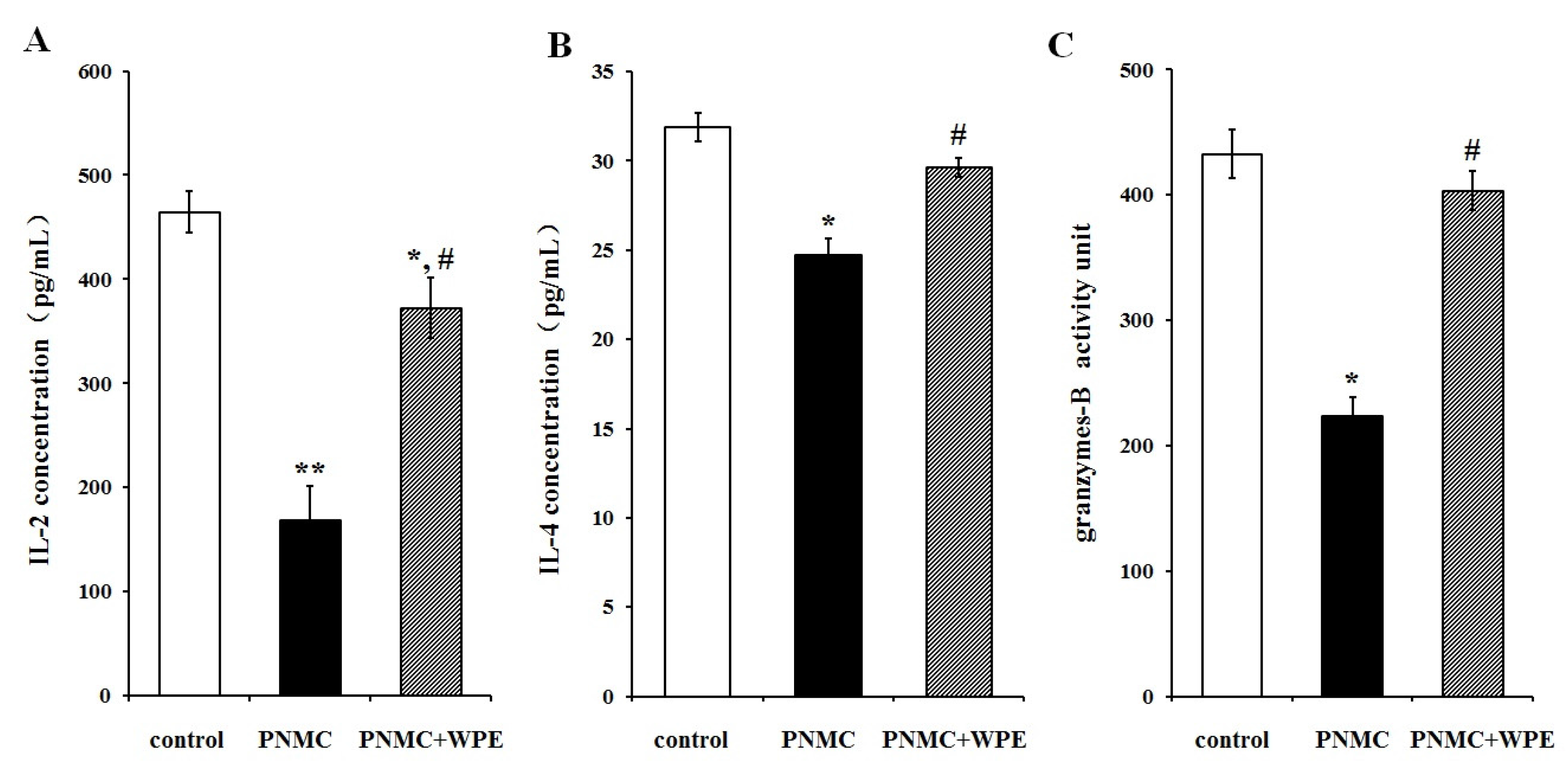
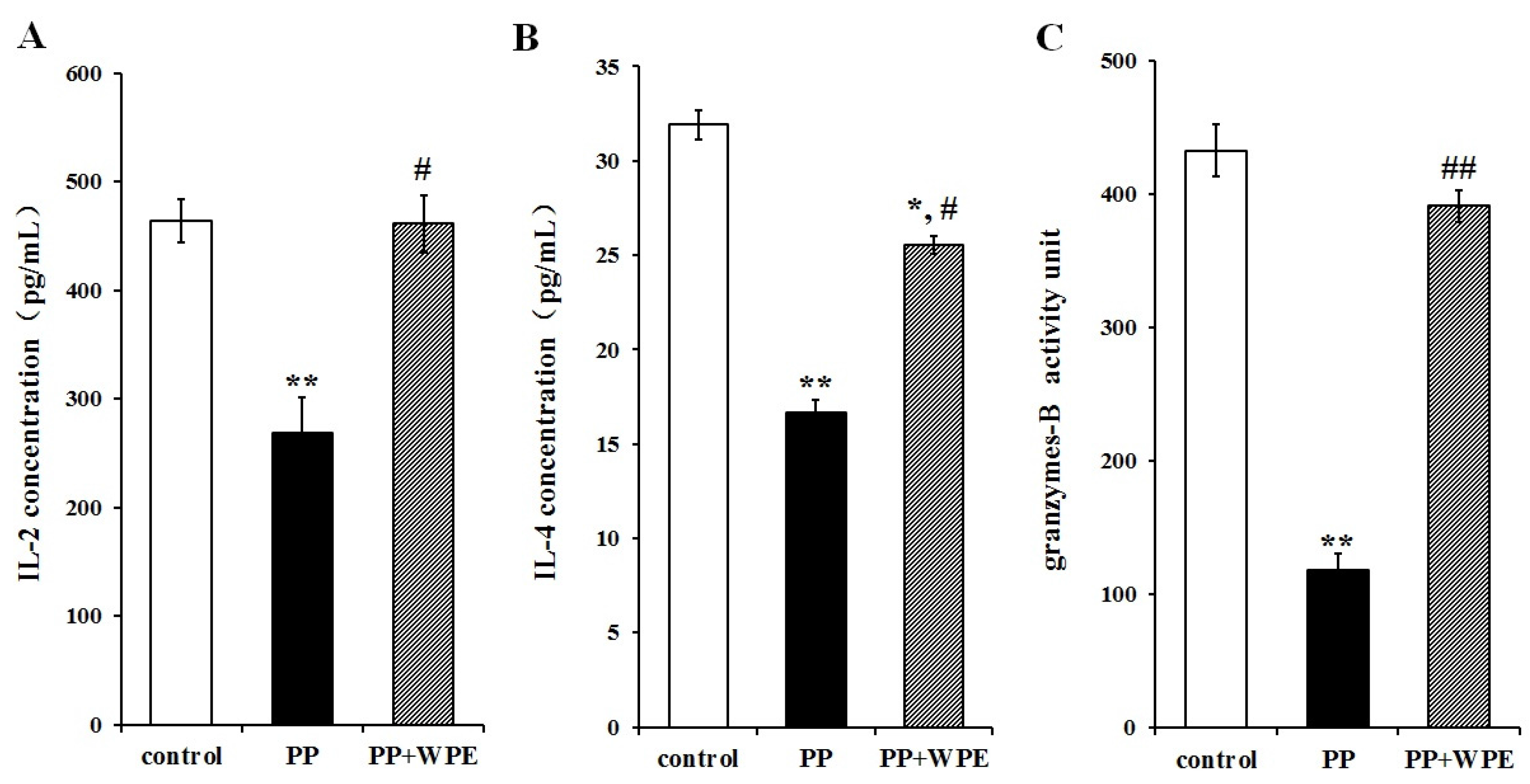
3.3. Treatment with WPE Prevents the Loss of Splenic T Cell Activity
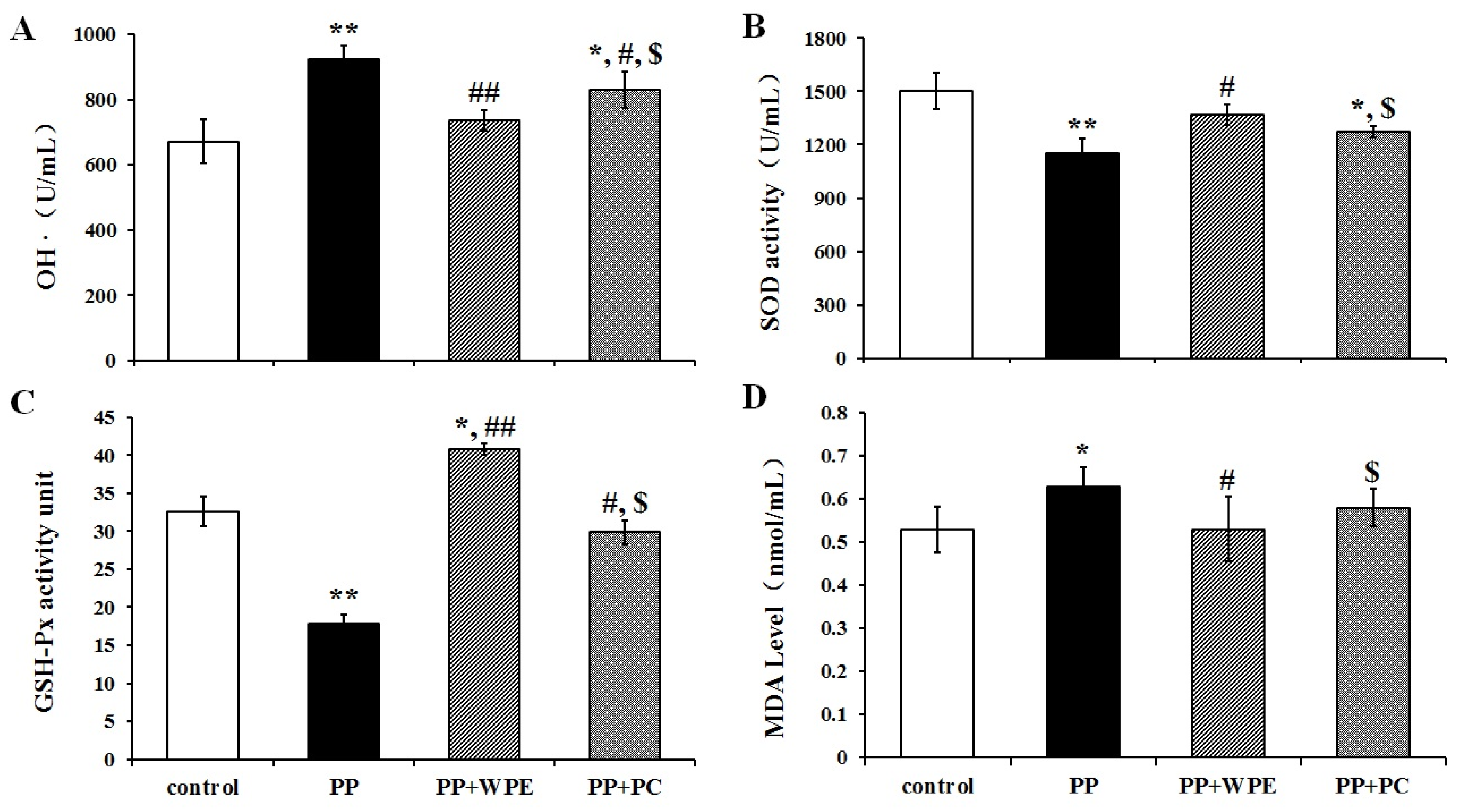
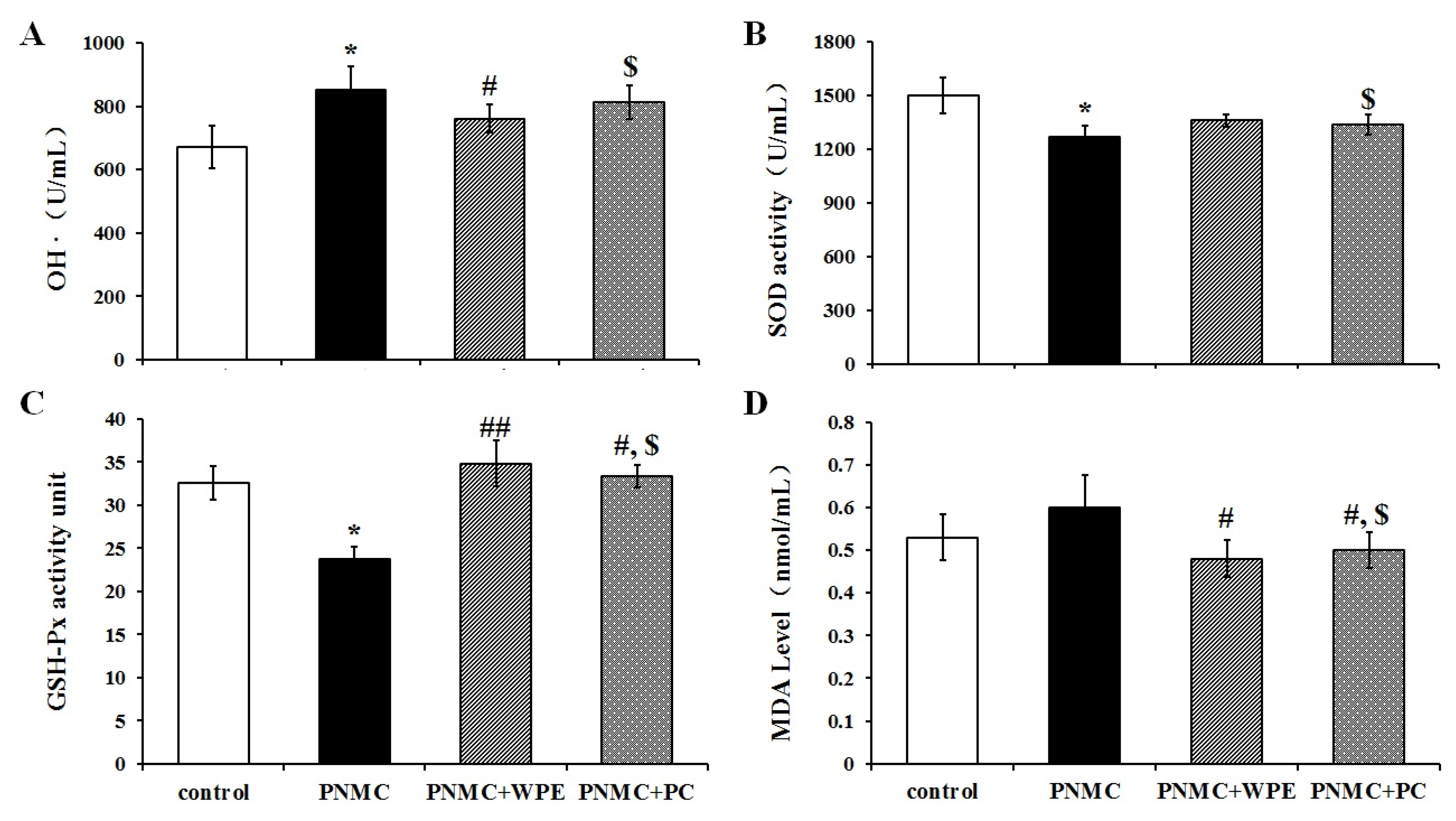
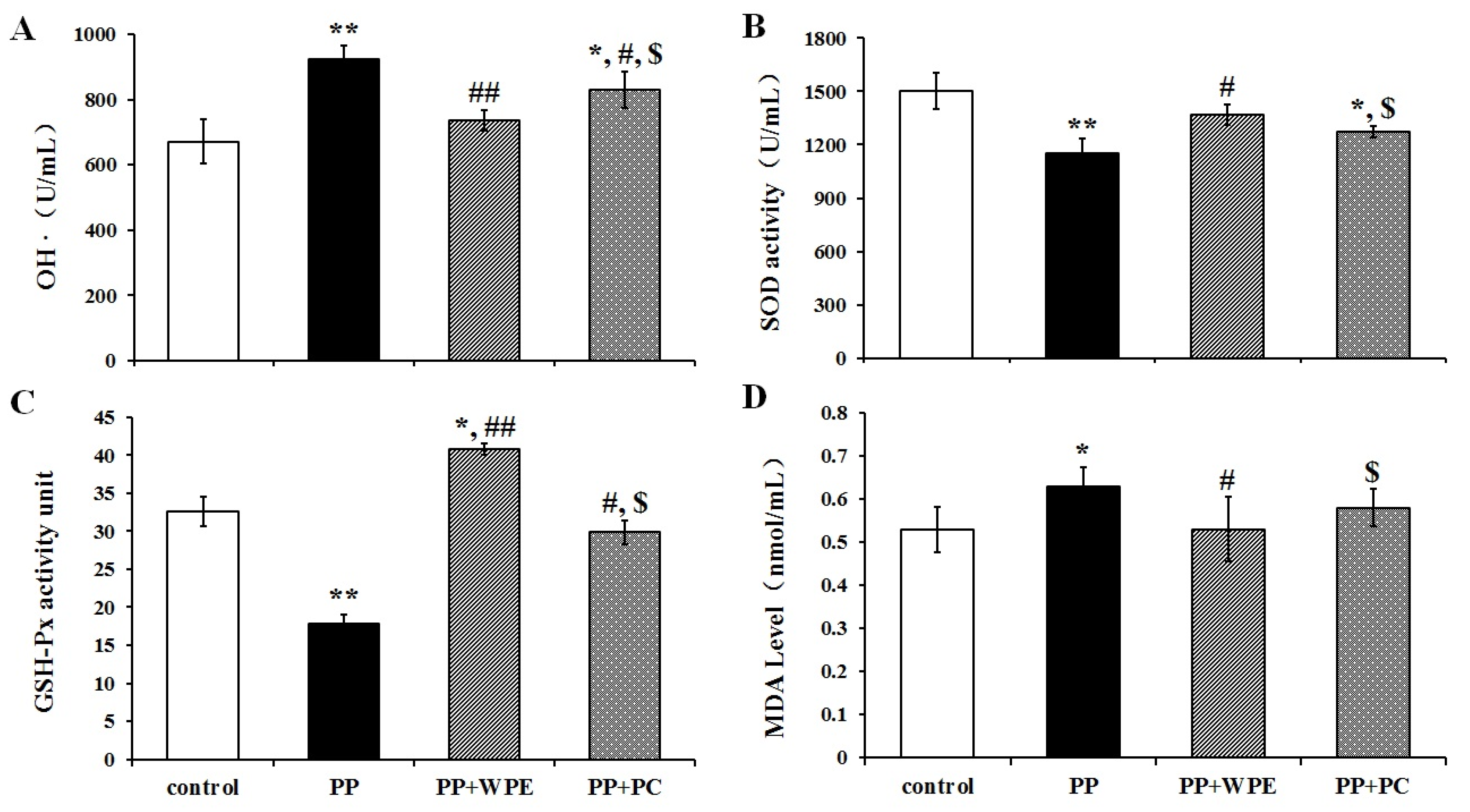
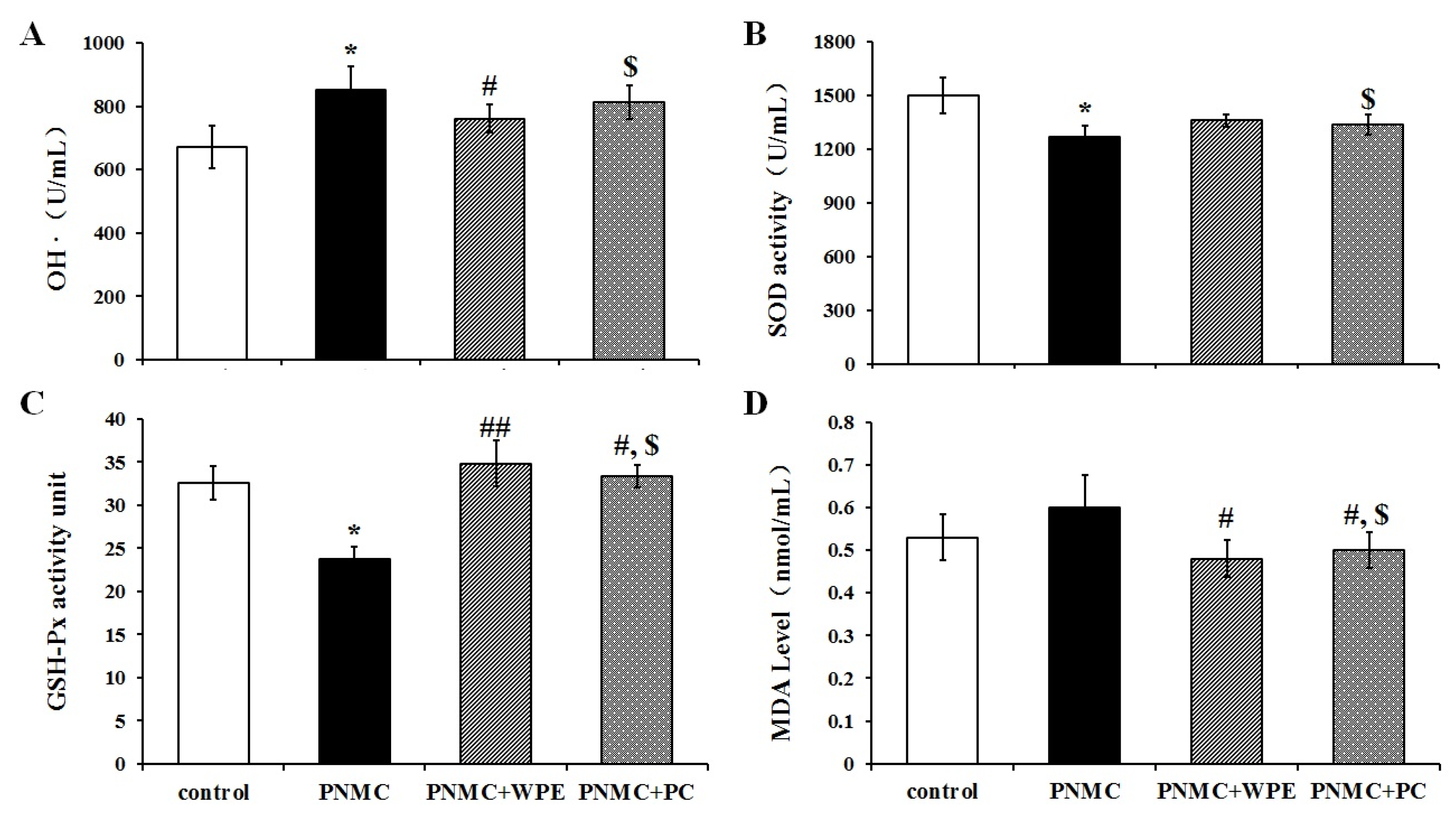
3.4. WPE Attenuates PP- and PNMC-Induced Oxidative Damage in Splenocytes
3.5. Quantification and Characterization of Phenolic Compounds in WPE
4. Discussion
5. Conclusions
Acknowledgments
Author Contribution
Conflicts of Interest
References
- Furuta, C.; Suzuki, A.K.; Taneda, S.; Kamata, K.; Hayashi, H.; Mori, Y.; Li, C.; Watanabe, G.; Taya, K. Estrogenic activities of nitrophenols in diesel exhaust particles. Biol. Reprod. 2004, 70, 1527–1533. [Google Scholar] [CrossRef] [PubMed]
- Furuta, C.; Li, C.; Taneda, S.; Suzuki, A.K.; Kamata, K.; Watanabe, G.; Taya, K. Immunohistological study for estrogenic activities of nitrophenols in diesel exhaust particles. Endocrine 2005, 27, 33–36. [Google Scholar] [CrossRef]
- Satoh, K.; Nonaka, R.; Ohyama, K.; Nagai, F. Androgenic and anti-androgenic effects of alkylphenols and parabens assessed using the reporter gene assay with stably transfected CHO-K1 Cells (AR-EcoScreen System). J. Health Sci. 2005, 51, 557–568. [Google Scholar] [CrossRef]
- Li, C.M.; Takahashi, S.; Taneda, S.; Furuta, C.; Watanabe, G.; Suzuki, A.K.; Taya, K. Impairment of testicular function in adult male Japanese quail (Coturnix japonica) after a single administration of 3-methyl-4-nitrophenol in diesel exhaust particles. J. Endocrinol. 2006, 189, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Li, C.M.; Takahashi, S.; Taneda, S.; Furuta, C.; Watanabe, G.; Suzuki, A.K.; Taya, K. Effects of 3-methyl-4-nitrophenol in diesel exhaust particles on the regulation of testicular function in immature male rats. J. Androl. 2007, 28, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Meier, S.; Morton, H.C.; Andersson, E.; Geffen, A.J.; Taranger, G.L.; Larsen, M.; Petersen, M.; Djurhuus, R.; Klungsøyr, J.; Svardal, A. Low-dose exposure to alkylphenols adversely affects sexual development of Atlantic cod (Gadus morhua): Acceleration of onset of puberty and delayed seasonal gonad development in mature female cod. Aquat. Toxicol. 2011, 105, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Yue, Z.; She, R.P.; Bao, H.H.; Li, W.G.; Wang, D.C.; Zhu, J.F.; Chang, L.L.; Yu, P. Exposure to 3-methyl-4-nitrophenol affects testicular morphology and induces spermatogenic cell apoptosis in immature male rats. Res. Vet. Sci. 2011, 91, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Che, W.J.; Liu, G.M.; Qiu, H.; Zhang, H.; Ran, Y.; Zeng, X.G.; Wen, W.H.; Shu, Y. Comparison of immunotoxic effects induced by the extracts from methanol and gasoline engine exhausts in vitro. Toxicol. Vitro 2010, 24, 1119–1125. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Kobayashi, M.; Inagaki, H.; Hirata, Y.; Sato, S.; Ishizaki, M.; Okamura, A.; Wang, D.; Nakajima, T.; Kamijima, M.; Kawada, T. Effect of oral exposure to fenitrothion and 3-methyl-4-nitrophenol on splenic cell populations and histopathological alterations in spleen in Wistar rats. Hum. Exp. Toxicol. 2010, 30, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Aam, B.B.; Fonnum, F. ROS scavenging effects of organic extract of diesel exhaust particles on human neutrophil granulocytes and rat alveolar macrophages. Toxicology 2007, 230, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Durga, M.; Nathiya, S.; Rajasekar, A.; Devasena, T. Effects of ultrafine petrol exhaust particles on cytotoxicity, oxidative stress generation, DNA damage and inflammation in human A549 lung cells and murine RAW 264.7 macrophages. Environ. Toxicol. Pharmacol. 2014, 38, 518–530. [Google Scholar] [CrossRef] [PubMed]
- Amakawa, K.; Terashima, T.; Matsuzaki, T.; Matsumaru, A.; Sagai, M.; Yamaguchi, K. Suppressive effects of diesel exhaust particles on cytokine release from human and murine alveolar macrophages. Exp. Lung Res. 2003, 29, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Chiang, D.; Sanjanwala, B.; Nadeau, K.C. Diesel exhaust particles impair regulatory T- cell function. In Proceedings of the International Conference on American Thoracic Society, San Diego, CA, USA, 15–20 May 2009. Abstract 4295.
- Sasaki, Y.; Ohtani, T.; Ito, Y.; Mizuashi, M.; Nakagawa, S.; Furukawa, T.; Horii, A.; Aiba, S. Molecular events in human T-cells treated with diesel exhaust particles or formaldehyde that underlie their diminished Interferon-γ and Interleukin-10 production. Int. Arch. Allergy Immunol. 2009, 148, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Karakaya, A.; Ates, I.; Yucesoy, B. Effects of occupational polycyclic aromatic hydro-carbon exposure on T-lymphocyte functions and natural killer cell activity in asphalt and coke oven workers. Hum. Exp. Toxicol. 2004, 23, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Mori, Y.; Kamata, K.; Toda, N.; Hayashi, H.; Seki, K.; Taneda, S.; Yoshino, S.; Sakushima, A.; Sakata, M.; Suzuki, A.K. Isolation of nitrophenols from diesel exhaust particles (DEP) as vasodilatation compounds. Biol. Pharm. Bull. 2003, 26, 394–395. [Google Scholar] [CrossRef] [PubMed]
- Murahashi, T.; Sasaki, S.; Nakajima, T. Determination of endocrine disruptors in automobile exhaust particulate matter. J. Health Sci. 2003, 49, 72–75. [Google Scholar] [CrossRef]
- Yang, L.B.; Ma, S.H.; Wan, Y.F.; Duan, S.Q.; Ye, S.Y.; Du, S.J.; Ruan, X.W.; Sheng, X.; Xu, M.Y.; Weng, Q.; Taya, K. In vitro effect of 4-pentylphenol and 3-methyl-4-nitrophenol on murine splenic lymphocyte populations and cytokine/granzyme production. J. Immunotoxicol. 2016, 31, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhang, R.; Li, W.M.; Niu, Y.J.; Guo, H.C.; Liu, X.H.; Hou, Y.C.; Zhao, L.J. The protective effect of grape seed procyanidin extract against cadmium-induced renal oxidative damage in mice. Environ. Toxicol Pharmcol. 2013, 36, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.M.; Ma, J.Q.; Liu, S.S.; Zheng, G.H.; Feng, Z.J.; Sun, J.M. Proanthocyanidins improves lead-induced cognitive impairments by blocking endoplasmic reticulum stress and nuclear factor-κB-mediated inflammatory pathways in rats. Food Chem. Toxicol. 2014, 72, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Fang, W.D.; Xiao, W.F.; Lu, L.P.; Jia, X.Y. Protective role of oligomeric proanthocyanidin complex against hazardous nodularin-induced oxidative toxicity in Carassius auratus lymphocytes. J. Hazard. Mater. 2014, 274, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Mi, Y.L.; Zhang, C.Q.; Li, C.M.; Taneda, S.; Watanabe, G.; Suzuki, A.K.; Taya, K. Quercetin protects embryonic chicken spermatogonial cells from oxidative damage intoxicated with 3-methyl-4-nitrophenol in primary culture. Toxicol. Lett. 2009, 190, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Bu, T.L.; Mi, Y.L.; Zeng, W.D.; Zhang, C.Q. Protective effect of quercetin on cadmium-induced oxidative toxicity on germ cells in male mice. Anat. Rec. 2011, 294, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Bu, T.L.; Jia, Y.D.; Lin, J.X.; Mi, Y.L.; Zhang, C.Q. Alleviative effect of quercetin on germ cells intoxicated by 3-methyl-4-nitrophenol from diesel exhaust particles. J. Zhejiang Univ. Sci. B 2012, 13, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Abarikwu, S.O. Protective effect of quercetin on atrazine-induced oxidative stress in the liver, kidney, brain, and heart of adult Wistar rats. Int. Toxicol. 2014, 21, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Kornsteiner, M.; Wagner, K.H.; Elmadfa, I. Tocopherols and total phenolics in 10 different nut types. Food Chem. 2006, 98, 381–387. [Google Scholar] [CrossRef]
- Halvorsen, B.L.; Carlsen, M.H.; Phillips, K.M.; Bøhn, S.K.; Holte, K.; Jacobs, D.R.; Blomhoff, R. Content of redox-active compounds (ie, antioxidants) in foods consumed in the United States. A. J. Clin. Nutr. 2006, 84, 95–135. [Google Scholar]
- Taha, N.A. Utility and importance of walnut, Juglans regia Linn: A review. Afr. J. Microbiol. Res. 2011, 5, 5796–5805. [Google Scholar]
- Eidi, A.; Moghadam, J.Z.; Mortazavi, P.; Rezazadeh, S.; Olamafar, S. Hepatoprotective effects of Juglans regia extract against CCl4-induced oxidative damage in rats. Pharm. Biol. 2013, 51, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Carey, A.N.; Fisher, D.R.; Joseph, J.A.; Shukitt, H.B. The ability of walnut extract and fatty acids to protect against the deleterious effects of oxidative stress and inflammation in hippocampal cells. Nutr. Neurosci. 2013, 16, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Muthaiyah, B.; Essa, M.; Chauhan, V.; Chauhan, A. Protective effects of walnut extract against amyloid beta peptide-induced cell death and oxidative stress in PC12 cells. Neurochem. Res. 2011, 36, 2096–2103. [Google Scholar] [CrossRef] [PubMed]
- Shabani, M.; Nazeri, M.; Parsania, S.; Razavinasab, M.; Zangiabadi, N.; Esmaeilpour, K.; Abareghi, F. Walnut consumption protects rats against cisplatin-induced neurotoxicity. NeuroToxicol 2012, 33, 1314–1321. [Google Scholar] [CrossRef] [PubMed]
- Benencia, F.; Courrèges, M.C.; Coulombié, F.C. In vivo and in vitro immunomodu-latory activities of Trichilia glabra aqueous leaf extracts. J. Ethnopharmacol. 2000, 69, 199–205. [Google Scholar] [CrossRef]
- Sun, H.X.; Qin, F.; Pan, Y.J. In vitro and in vivo immunosuppressive activity of Spica Prunellae ethanol extract on the immune responses in mice. J. Ethnopharmacol. 2005, 101, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Hirata, Y.; Piao, S.; Minami, M. Immunotoxicity of N,N-diethylaniline in mice: Effect on natural killer activity, cytotoxic T-lymphocyte activity, lymphocyte proliferation response, and cellular components of the spleen. Toxicology 2000, 150, 179–189. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, M.M.; Su, G.W.; Cai, M.S.; Zhou, C.M.; Huang, J.Y.; Lin, L.Z. The antioxidant activities and the xanthine oxidase inhibition effects of walnut (Juglans regia L.) fruit, stem and leaf. Int. J. Food Sci. Tech. 2015, 50, 233–239. [Google Scholar] [CrossRef]
- Grace, M.H.; Warlick, C.W.; Neff, S.A.; Lila, M.A. Efficient preparative isolation and identification of walnut bioactive components using high-speed counter-current chromatography and LC-ESI-IT-TOF-MS. Food Chem. 2014, 158, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Regueiro, J.; Sánchez-González, C.; Vallverdú-Queralt, A.; Simal-Gándara, J.; Lamuela-Raventós, R.; Izquierdo-Pulido, M. Comprehensive identification of walnut polyphenols by liquid chromatography coupled to linear ion trap–Orbitrap mass spectrometry. Food Chem. 2014, 152, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Slatnar, A.; Mikulic-Petkovsek, M.; Stampar, F.; Veberic, R.; Solar, A. Identification and quantification of phenolic compounds in kernels, oil and bagasse pellets of common walnut (Juglans regia L.). Food Res. Int. 2015, 67, 255–263. [Google Scholar] [CrossRef]
- Van der Hooft, J.J.; Akermi, M.; Unlu, F.Y.; Mihaleva, V.; Roldan, V.G.; Bino, R.J.; de Vos, R.C.; Vervoort, J. Structural annotation and elucidation of conjugated phenolic compounds in black, green, and white tea extracts. J. Agric. Food Chem. 2012, 60, 8841–8850. [Google Scholar] [CrossRef] [PubMed]
- Ou, K.; Gu, L. Absorption and metabolism of proanthocyanidins. J. Funct. Foods 2014, 7, 43–53. [Google Scholar] [CrossRef]
- Sanbongi, C.; Takano, H.; Osakabe, N.; Sasa, N.; Natsume, M.; Yanagisawa, R.; Inoue, K.; Kato, Y.; Osawa, T.; Yoshikawa, T. Rosmarinic acid inhibits lung injury induced by diesel exhaust particles. Free Radical. Biol. Med. 2003, 34, 1060–1069. [Google Scholar] [CrossRef]
- Bhullar, K.S.; Rupasinghe, H.P.V. Partridgeberry polyphenols protect primary cortical and hippocampal neurons against β-amyloid toxicity. Food Res. Int. 2015, 74, 237–249. [Google Scholar] [CrossRef]
- Sanbongi, C.; Suzuki, N.; Sakane, T. Polyphenols in chocolate, which have antioxidant activity, modulate immune functions in humansin vitro. Cell. Immunol. 1997, 177, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Nani, A.; Belarbi, M.; Ksouri-Megdiche, W.; Abdoul-Azize, S.; Benammar, C.; Ghiringhelli, F.; Hichami, A.; Khan, N.A. Effects of polyphenols and lipids from Pennisetum glaucum grains on T-cell activation: Modulation of Ca2+ and ERK1/ERK2 signaling. BMC Complement. Altern. Med. 2015, 15, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Krifa, M.; Bouhlel, I.; Ghedira-Chekir, L.; Ghedira, K. Immunomodulatory and cellular anti-oxidant activities of an aqueous extract of Limoniastrum guyonianum gall. J. Ethnopharmacol. 2013, 146, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, K.; Eto, N.; Sakakibara, Y.; Matsusita, Y.; Sugamoto, K.; Morishita, K.; Suiko, M. Oligomeric proanthocyanidins from rabbiteye blueberry leaves inhibits the proliferation of human T-cell lymphotropic virus type 1-associated cell lines via apoptosis and cell cycle arrest. J. Funct. Foods 2014, 6, 356–366. [Google Scholar] [CrossRef]
- Shen, R.; Deng, W.; Li, C.; Zeng, G. A natural flavonoid glucoside icariin inhibits Th1 and Th17 cell differentiation and ameliorates experimental autoimmune encephalomyelitis. Int. Immunopharmacol. 2015, 24, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Edfeldt, K.; Liu, P.T.; Chun, R.; Fabri, M.; Schenk, M.; Wheelwright, M.; Keegan, C.; Krutzik, S.R.; Adams, J.S.; Hewison, M.; et al. T-cell cytokines differentially control human monocyte antimicrobial responses by regulating vitamin D metabolism. Proc. Natl. Acad. Sci. USA 2010, 107, 22593–22598. [Google Scholar] [CrossRef] [PubMed]
- Magrone, T.; Fontana, S.; Laforgia, F.; Dragone, T.; Jirillo, E.; Passantino, L. Administration of a polyphenol-enriched feed to farmed sea bass (Dicentrarchus labrax L.) modulates intestinal and spleen immune responses. Oxid. Med. Cell. Longev. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.C.; Teuber, S.S. Ellagic acid and polyphenolics present in walnut kernels inhibit in vitro human peripheral blood mononuclear cell proliferation and alter cytokine production. Ann. N. Y. Acad. Sci. 2010, 1190, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.G.; Xu, P.; Liu, Q.; Yu, C.H.; Zhang, Y.; Chen, S.H.; Li, Y.M. Effect of tea polyphenol on cytokine gene expression in rats with alcoholic liver disease. HBPD Int. 2006, 5, 268–272. [Google Scholar] [PubMed]
- Varilek, G.W.; Yang, F.; Lee, E.Y.; deVilliers, W.J.; Zhong, J.; Oz, H.S.; Westberry, K.F.; McClain, C.J. Green tea polyphenol extract attenuates inflammation in interleukin-2-deficient mice, a model of autoimmunity. J. Nutr. 2001, 131, 2034–2039. [Google Scholar] [PubMed]
- Pereira, J.A.; Oliveira, I.; Sousa, A.; Ferreira, I.C.F.R.; Bento, A.; Estevinho, L. Bioactive properties and chemical composition of six walnut (Juglans regia L.) cultivars. Food Chem. Toxicol. 2008, 46, 2103–2111. [Google Scholar] [CrossRef] [PubMed]
- Vinson, J.A.; Cai, Y. Nuts, especially walnuts, have both antioxidant quantity and efficacy and exhibit significant potential health benefits. Food Funct. 2012, 3, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Scalbert, A. Ellagitannins—Nature, occurrence and dietary burden. J. Sci. Food Agric. 2000, 80, 1118–1125. [Google Scholar] [CrossRef]
- Chan, S.T.; Lin, Y.C.; Chuang, C.H.; Shiau, R.J.; Liao, J.W.; Yeh, S.L. Oral and intraperitoneal administration of quercetin decreased lymphocyte DNA damage and plasma lipid peroxidation induced by TSA in vivo. Biomed. Res. Int. 2009, 15, 197–243. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.C.; Lin, S.Y.; Yang, J.S.; Liu, K.C.; Tang, Y.J.; Yang, M.D.; Chiang, J.H.; Lu, C.C.; Wu, C.L.; Chiu, T.H. Gallic acid inhibits murine leukemia WEHI-3 cells in vivo and promotes macrophage phagocytosis. Vivo 2009, 23, 409–413. [Google Scholar]
- Gođevac, D.; Tešević, V.; Vajs, V.; Milosavljević, S.; Stanković, M. Antioxidant properties of raspberry seed extracts on micronucleus distribution in peripheral blood lymphocytes. Food Chem. Toxicology 2009, 47, 2853–2859. [Google Scholar]
- Mosbah, R.; Yousef, M.I.; Mantovani, A. Nicotine-induced reproductive toxicity, oxidative damage, histological changes and haematotoxicity in male rats: The protective effects of green tea extract. Exp. Toxicol. Pathol. 2015, 67, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Mi, Y.L.; Zhang, C.Q.; Li, C.M.; Taneda, S.; Watanabe, G.; Suzuki, A.K.; Taya, K. Quercetin attenuates oxidative damage induced by treatment of embryonic chicken spermatogonial cells with 4-nitro-3-phenylphenol in diesel exhaust particles. Biosci. Biotechnol. Biochem. 2010, 74, 934–938. [Google Scholar] [CrossRef] [PubMed]
- Mi, Y.L.; Zhang, C.Q.; Li, C.M.; Taneda, S.; Watanabe, G.; Suzuki, A.K.; Taya, K. Protective effect of quercetin on the reproductive toxicity of 4-nitrophenol in diesel exhaust particles on male embryonic chickens. J. Reprod. Dev. 2010, 56, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.D.; Chen, C.Y.; Zhao, S.L.; Ge, F.; Liu, D.Q.; Hao, S. Effects of walnut polyphenol on learning and memory functions in hypercholesterolemia mice. J. Food Nutr. Res. 2014, 2, 450–456. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | tR (min) | Measured [M − H]−(m/z) | Predicted [M − H]− (m/z) | λ (nm) | Molecular Formula | Identification | Expressed as | Reference |
|---|---|---|---|---|---|---|---|---|
| 1 | 12.663, 12.906 | 300.9993 | 300.9990 | 280 | C14H6O8 | Ellagic aci | Ellagitannins | [37,38,39] |
| 2 | 11.946 | 433.0432 | 433.0412 | 280 | C19H14O12 | Ellagic acid 4-O-xyloside | Ellagitannins | [37,38,39] |
| 3 | 12.507 | 433.1119 | 433.0772 | 280 | C20H18O11 | Quercetin pentoside isomer | Quercetin | [38] |
| 4 | 11.946 | 456.0383 | 457.0781 | 280 | C22H17O11 | Epigallocatechin-3-O-gallate | Ellagitannins | [40] |
| 5 | 13.74 | 447.0831 | 447.0938 | 280 | C21H19O11 | Kaempferol-3-O-glucoside | Ellagitannins | [40] |
| 6 | 12.507, 12.663, 12.906 | 469.0492 | 469.0049 | 280 | C21H10O13 | Valoneic acid dilactone | Valoneic acid dilactone | [38] |
| 7 | 11.946 | 481.5355 | 481.0620 | 280 | C20H18O14 | 2,3-O-HHDP-d-glucoside | Ellagitannins | [38] |
| 8 | 9.828, 10.020, 10.512, 10.784 | 483.5192 | 483.0777 | 280 | C20H20O14 | Digalloyl-glucose isomer | Gallic acid | [38,39] |
| 9 | 12.25 | 615.5905 | 615.0986 | 280 | C28H24O16 | Quercetin galloylhexoside isomer | Quercetin | [38] |
| 10 | 8.382, 8.751, 9.624 | 633.0720 | 633.0720 | 280 | C27H22O18 | Strictinin (galloyl-HHDP-glucose) | Ellagitannins | [37,38,39] |
| 11 | 10.784, 11.136 | 635.0876 | 635.0877 | 280 | C27H24O18 | Trigalloyl-glucose isomer | Ellagitannins | [38] |
| 12 | 8.382, 8.751, 9.003 | 783.0685 | 783.0681 | 280 | C34H24O22 | Pedunculagin (bis-HHDP-glucose) | Ellagitannins | [37,38,39] |
| 13 | 10.020, 10.236 | 785.0808 | 785.0840 | 280 | C34H26O22 | Tellimagrandin I isomer(digalloyl-HHDP-glucose) | Ellagitannins | [37,38,39] |
| 14 | 11.946, 12.250 | 787.1144 | 787.0996 | 280 | C34H28O22 | Tetragalloyl-glucose | Ellagitannins | [38] |
| 15 | 10.784, 11.488 | 933.0316 | 933.0630 | 280 | C41H26O26 | Praecoxin D | Ellagitannins | [37,38,39] |
| 16 | 11.136 | 935.0746 | 935.0786 | 280 | C41H28O26 | Casuarinin(Galloyl bis HHDP glucose) | Ellagitannins | [37,38,39] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Ma, S.; Han, Y.; Wang, Y.; Guo, Y.; Weng, Q.; Xu, M. Walnut Polyphenol Extract Attenuates Immunotoxicity Induced by 4-Pentylphenol and 3-methyl-4-nitrophenol in Murine Splenic Lymphocyte. Nutrients 2016, 8, 287. https://doi.org/10.3390/nu8050287
Yang L, Ma S, Han Y, Wang Y, Guo Y, Weng Q, Xu M. Walnut Polyphenol Extract Attenuates Immunotoxicity Induced by 4-Pentylphenol and 3-methyl-4-nitrophenol in Murine Splenic Lymphocyte. Nutrients. 2016; 8(5):287. https://doi.org/10.3390/nu8050287
Chicago/Turabian StyleYang, Lubing, Sihui Ma, Yu Han, Yuhan Wang, Yan Guo, Qiang Weng, and Meiyu Xu. 2016. "Walnut Polyphenol Extract Attenuates Immunotoxicity Induced by 4-Pentylphenol and 3-methyl-4-nitrophenol in Murine Splenic Lymphocyte" Nutrients 8, no. 5: 287. https://doi.org/10.3390/nu8050287