Dietary Natural Products for Prevention and Treatment of Liver Cancer



Abstract
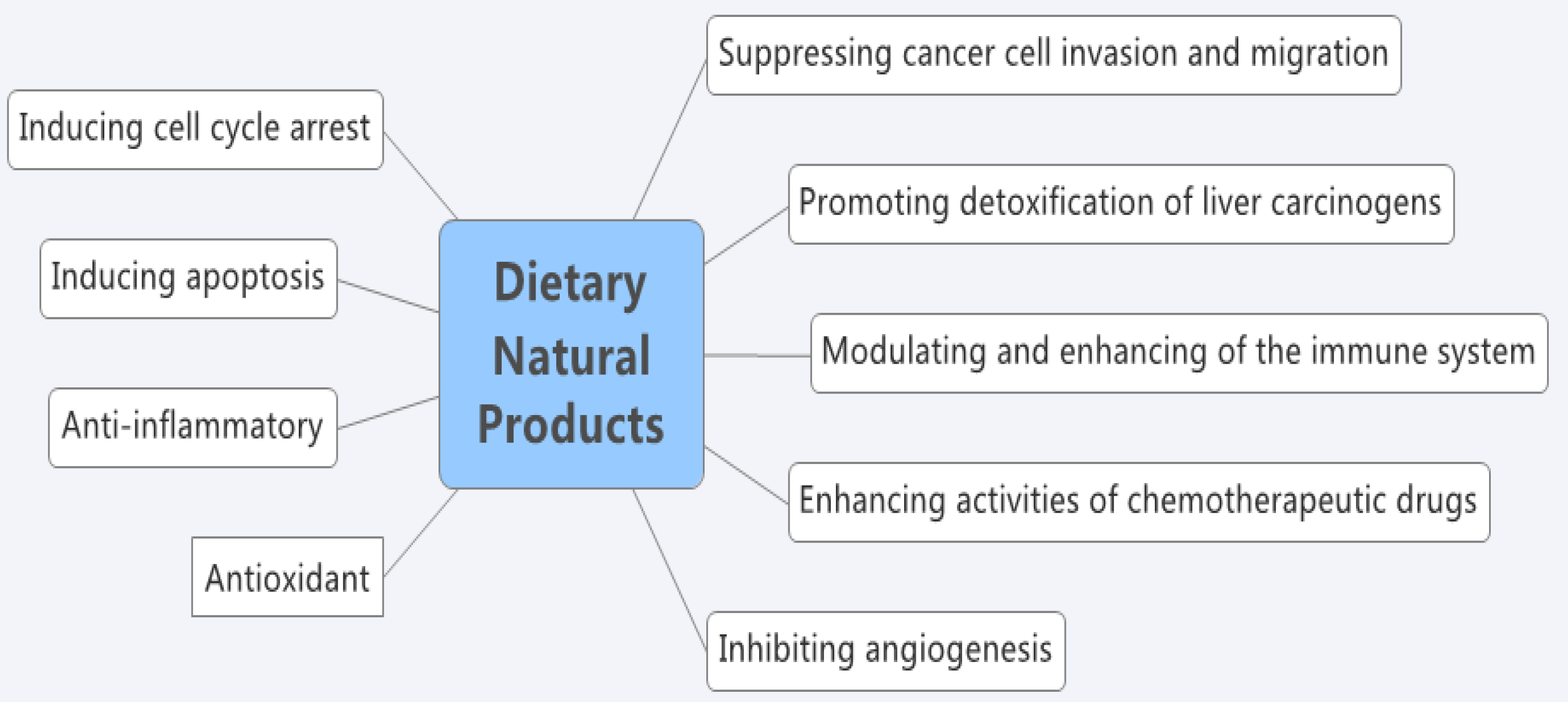
:1. Introduction
2. Fruits
2.1. Grape
2.2. Black Currant
2.3. Plum
2.4. Other Fruits
3. Vegetables
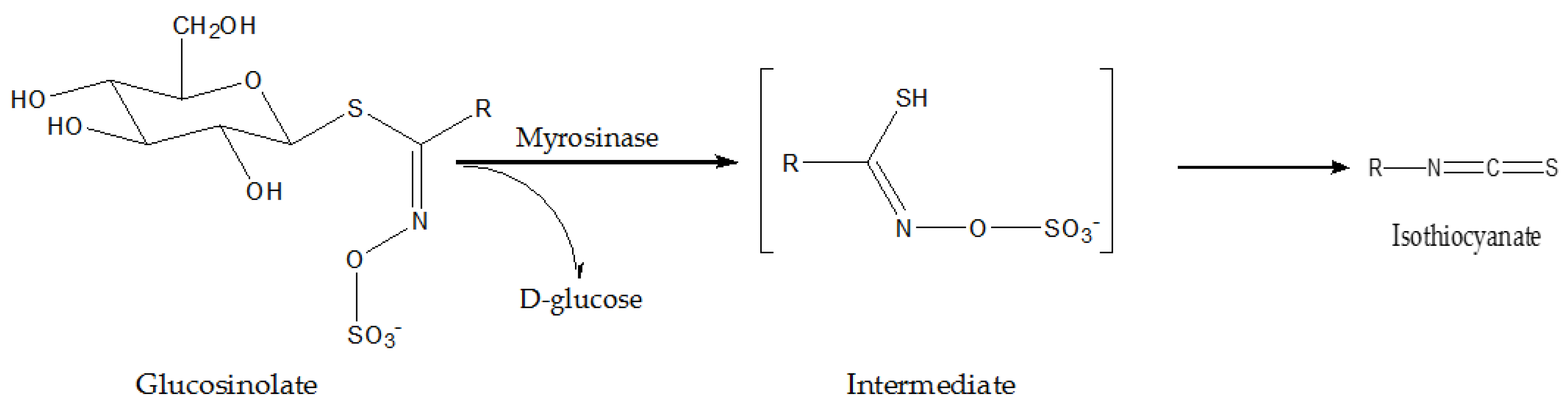
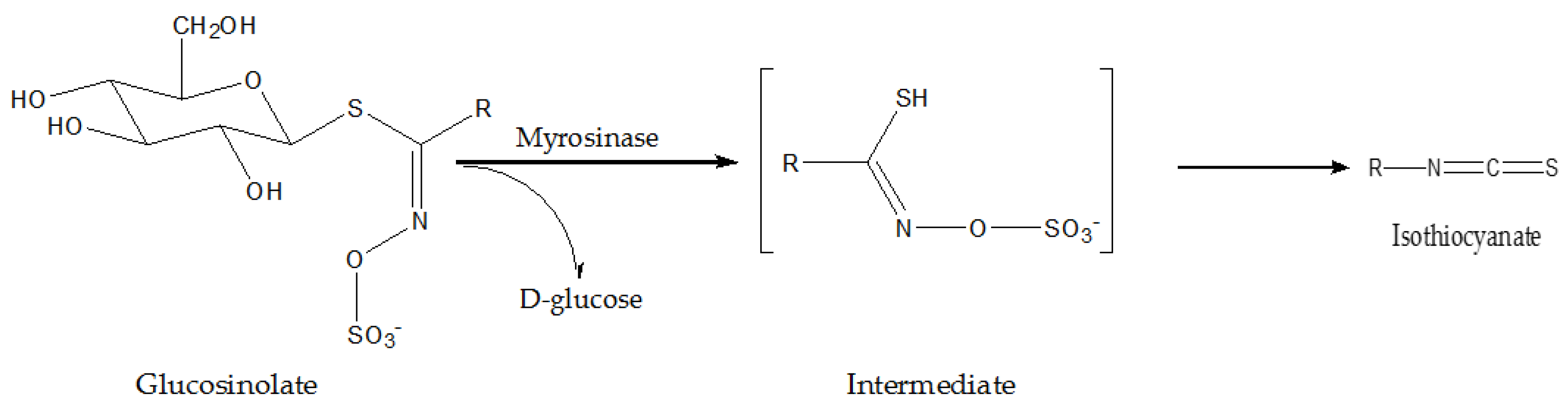
3.1. Cruciferous Vegetables
3.2. French Bean
3.3. Tomato
3.4. Asparagus
3.5. Other Vegetables
4. Spices
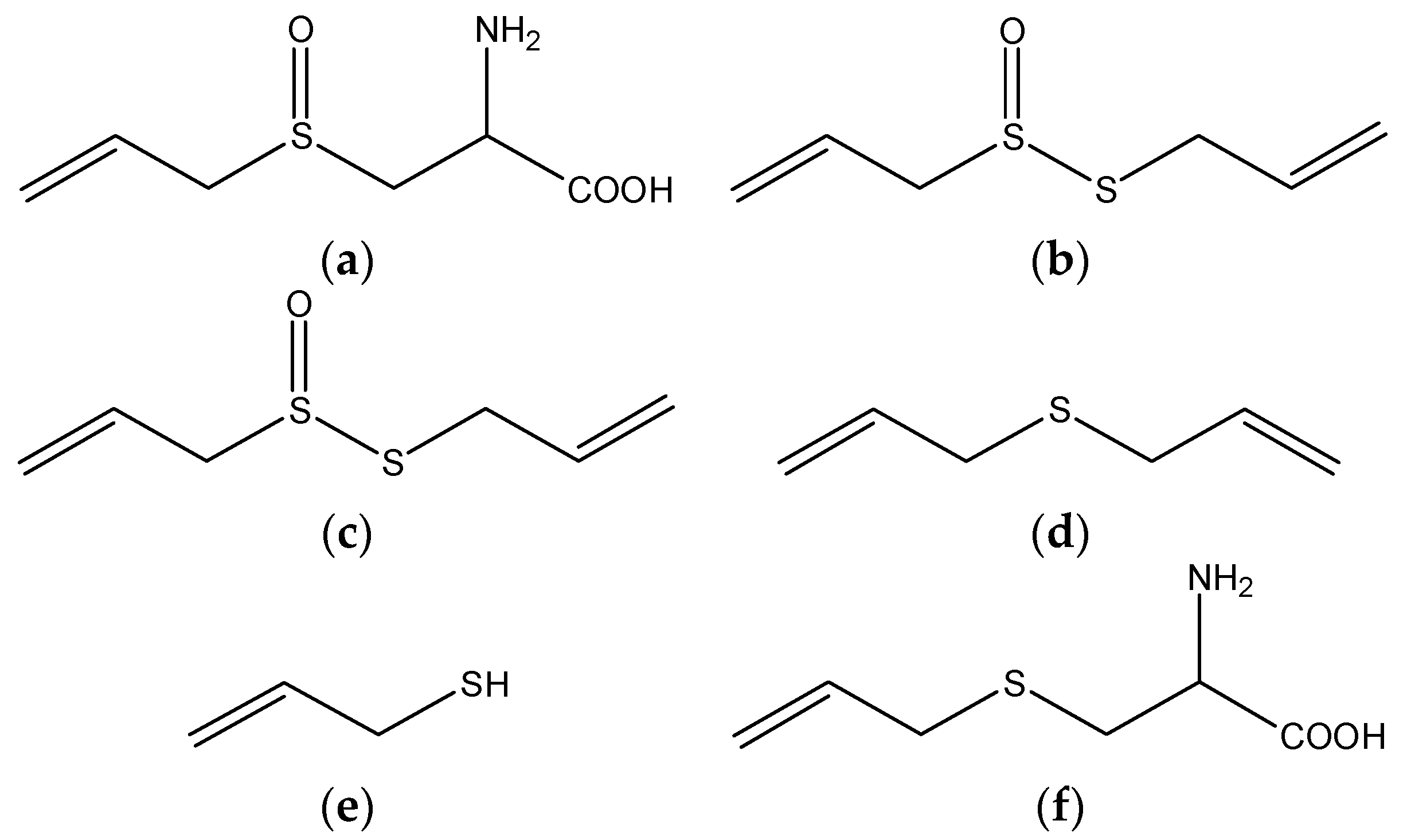
4.1. Garlic
4.2. Turmeric
4.3. Pepper
4.4. Ginger
4.5. Other Spices
5. Soy
6. Cereals
7. Edible Macro-Fungi
8. Effects of Combination of Dietary Natural Products with Anticancer Treatments
9. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Marquardt, J.U.; Andersen, J.B.; Thorgeirsson, S.S. Functional and genetic deconstruction of the cellular origin in liver cancer. Nat. Rev. Cancer 2015, 15, 653–667. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- De Jong, M.C.; Nathan, H.; Sotiropoulos, G.C.; Paul, A.; Alexandrescu, S.; Marques, H.; Pulitano, C.; Barroso, E.; Clary, B.M.; Aldrighetti, L.; et al. Intrahepatic cholangiocarcinoma: An international multi-institutional analysis of prognostic factors and lymph node assessment. J. Clin. Oncol. 2011, 29, 3140–3145. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, R.; Mitra, A. An overview of effective therapies and recent advances in biomarkers for chronic liver diseases and associated liver cancer. Int. Immunopharmacol. 2015, 24, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Bruix, J.; Raoul, J.L.; Sherman, M.; Mazzaferro, V.; Bolondi, L.; Craxi, A.; Galle, P.R.; Santoro, A.; Beaugrand, M.; Sangiovanni, A.; et al. Efficacy and safety of sorafenib in patients with advanced hepatocellular carcinoma: Subanalyses of a phase III trial. J. Hepatol. 2012, 57, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.l.; Zhao, Y.; Xu, F.; Jia, C.J.; Xu, Y.Q.; Dai, C.L. An updated meta-analysis of randomized controlled trials assessing the effect of sorafenib in advanced hepatocellular carcinoma. PLoS ONE 2014, 9, e112530. [Google Scholar] [CrossRef] [PubMed]
- Chen, J; Jin, R.A.; Zhao, J.; Liu, J.H.; Ying, H.N.; Yan, H.; Zhou, S.J.; Liang, Y.L.; Huang, D.Y.; Liang, X.; et al. Potential molecular, cellular and microenvironmental mechanism of Sorafenib resistance in hepatocellular carcinoma. Cancer Lett. 2015, 367, 1–11. [Google Scholar]
- Soerjomataram, I.; Oomen, D.; Lemmens, V.; Oenema, A.; Benetou, V.; Trichopoulou, A.; Coebergh, J.W.; Barendregt, J.; de Vries, E. Increased consumption of fruit and vegetable and future cancer incidence in selected European countries. Eur. J. Cancer 2010, 46, 2563–2580. [Google Scholar] [CrossRef] [PubMed]
- Turati, F.; Rossi, M.; Pelucchi, C.; Levi, F.; La Vecchia, C. Fruit and vegetables and cancer risk: A review of southern European studies. Br. J. Nutr. 2015, 113, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Thoppil, R.J.; Bhatia, D.; Barnes, K.F.; Haznagy-Radnai, E.; Hohmann, J.; Darvesh, A.S.; Bishayee, A. Black currant anthocyanins abrogate oxidative stress through Nrf2-mediated antioxidant mechanisms in a rat model of hepatocellular carcinoma. Curr. Cancer Drug Targets 2012, 12, 1244–1257. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.H.; Gwon, I.H.; Gyu, L.S.; Kim, D.I.; Jeong, S.H.; Lee, I.S. Inhibitory effect of immature plum on PMA-induced MMP-9 expression in human hepatocellular carcinoma. Nat. Prod. Res. 2009, 23, 704–718. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Ye, L.; Hao, X.; Ren, Z.; Ren, S.; Xu, K.; Li, J. Polysaccharides from Tricholoma matsutake and Lentinus edodes enhance 5-fluorouracil-mediated H22 cell growth inhibition. J. Tradit. Chin. Med. 2014, 34, 309–316. [Google Scholar] [CrossRef]
- Jo, J.Y.; de Mejia, E.G.; Lila, M.A. Cytotoxicity of bioactive polymeric fractions from grape cell culture on human hepatocellular carcinoma, murine leukemia and non-cancerous PK15 kidney cells. Food Chem. Toxicol. 2006, 44, 1758–1767. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Xu, B.T.; Xu, X.R.; Gan, R.Y.; Zhang, Y.; Xia, E.Q.; Li, H.B. Antioxidant capacities and total phenolic contents of 62 fruits. Food Chem. 2011, 129, 345–350. [Google Scholar] [CrossRef]
- Li, F.; Li, S.; Li, H.B.; Deng, G.F.; Ling, W.H.; Wu, S.; Xu, X.R.; Chen, F. Antiproliferative activity of peels, pulps and seeds of 61 fruits. J. Funct. Foods 2013, 5, 1298–1309. [Google Scholar] [CrossRef]
- Deng, G.F.; Shen, C.; Xu, X.R.; Kuang, R.D.; Guo, Y.J.; Zeng, L.S.; Gao, L.L.; Lin, X.; Xie, J.F.; Xia, E.Q.; et al. Potential of fruit wastes as natural resources of bioactive compounds. Int. J. Mol. Sci. 2012, 13, 8308–8323. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.Q.; Deng, G.F.; Guo, Y.J.; Li, H.B. Biological activities of polyphenols from grapes. Int. J. Mol. Sci. 2010, 11, 622–646. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.; Gonzalez De Mejia, E.; Lila, M.A. Effects of grape cell culture extracts on human topoisomerase II catalytic activity and characterization of active fractions. J. Agric. Food Chem. 2005, 53, 2489–2498. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.L.; Liu, B.X.; Zhong, J.Y.; Sun, L.B.; Yu, H.S. Effect of grape procyanidins on tumor angiogenesis in liver cancer xenograft models. Asian Pac. J. Cancer Prev. 2014, 15, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Scola, G.; Fernandes, C.L.C.; Menin, E.; Salvador, M. Suppression of oncoprotein Her-2 and DNA damage after treatment with Flavan-3-ol Vitis labrusca extract. Anticancer Agents Med. Chem. 2013, 13, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A.; Haznagy-Radnai, E.; Mbimba, T.; Sipos, P.; Morazzoni, P.; Darvesh, A.S.; Bhatia, D.; Hohmann, J. Anthocyanin-rich black currant extract suppresses the growth of human hepatocellular carcinoma cells. Nat. Prod. Commun. 2010, 5, 1613–1618. [Google Scholar] [PubMed]
- Bishayee, A.; Thoppil, R.J.; Mandal, A.; Darvesh, A.S.; Ohanyan, V.; Meszaros, J.G.; Haznagy-Radnai, E.; Hohmann, J.; Bhatia, D. Black currant phytoconstituents exert chemoprevention of diethylnitrosamine-initiated hepatocarcinogenesis by suppression of the inflammatory response. Mol. Carcinog. 2013, 52, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A.; Mbimba, T.; Thoppil, R.J.; Haznagy-Radnai, E.; Sipos, P.; Darvesh, A.S.; Folkesson, H.G.; Hohmann, J. Anthocyanin-rich black currant (Ribes nigrum L.) extract affords chemoprevention against diethylnitrosamine-induced hepatocellular carcinogenesis in rats. J. Nutr. Biochem. 2011, 22, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Ramos, S.; Alia, M.; Bravo, L.; Goya, L. Comparative effects of food-derived polyphenols on the viability and apoptosis of a human hepatoma cell line (HepG2). J. Agric. Food Chem. 2005, 53, 1271–1280. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.H.; Im, H.G.; Kim, H.I.; Lee, I.S. Induction of apoptosis by immature plum in human hepatocellular carcinoma. J. Med. Food 2009, 12, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Yu, M.H.; Lee, I.S. Inhibitory effects of methanol extract of plum (Prunus salicina L., cv. “Soldam”) fruits against benzo(α)pyrene-induced toxicity in mice. Food Chem. Toxicol. 2008, 46, 3407–3413. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A.; Thoppil, R.J.; Darvesh, A.S.; Ohanyan, V.; Meszaros, J.G.; Bhatia, D. Pomegranate phytoconstituents blunt the inflammatory cascade in a chemically induced rodent model of hepatocellular carcinogenesis. J. Nutr. Biochem. 2013, 24, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A.; Bhatia, D.; Thoppil, R.J.; Darvesh, A.S.; Nevo, E.; Lansky, E.P. Pomegranate-mediated chemoprevention of experimental hepatocarcinogenesis involves Nrf2-regulated antioxidant mechanisms. Carcinogenesis 2011, 32, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Kojima-Yuasa, A.; Xu, S.; Kennedy, D.O.; Hasuma, T.; Matsui-Yuasa, I. Combination of Zizyphus jujuba and green tea extracts exerts excellent cytotoxic activity in HepG2 cells via reducing the expression of APRIL. Am. J. Chin. Med. 2009, 37, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Kojima-Yuasa, A.; Xu, S.; Norikura, T.; Kennedy, D.O.; Hasuma, T.; Matsui-Yuasa, I. Green tea extract enhances the selective cytotoxic activity of Zizyphus jujuba extracts in HepG2 cells. Am. J. Chin. Med. 2008, 36, 729–744. [Google Scholar] [CrossRef] [PubMed]
- Sudan, S.; Rupasinghe, H.P. Flavonoid-enriched apple fraction AF4 induces cell cycle arrest, DNA topoisomerase II inhibition, and apoptosis in human liver cancer HepG2 cells. Nutr. Cancer 2014, 66, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xu, S.S.; Chen, J.W.; Wang, Y.; Xu, H.Q.; Fan, N.B.; Li, X. Anti-tumor activity of Annona squamosa seeds extract containing annonaceous acetogenin compounds. J. Ethnopharmacol. 2012, 142, 462–466. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Ohlander, M.; Jeppsson, N.; Björk, L.; Trajkovski, V. Changes in antioxidant effects and their relationship to phytonutrients in fruits of sea buckthorn (Hippophae rhamnoides L.) during maturation. J. Agr. Food Chem. 2000, 48, 1485–1490. [Google Scholar] [CrossRef]
- Teng, B.S.; Lu, Y.H.; Wang, Z.T.; Tao, X.Y.; Wei, D.Z. In vitro anti-tumor activity of isorhamnetin isolated from Hippophae rhamnoides L. against BEL-7402 cells. Pharmacol. Res. 2006, 54, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Tien, P.G.; Kayama, F.; Konishi, F.; Tamemoto, H.; Kasono, K.; Hung, N.T.; Kuroki, M.; Ishikawa, S.E.; Van, C.N.; Kawakami, M. Inhibition of tumor growth and angiogenesis by water extract of Gac fruit (Momordica cochinchinensis Spreng). Int. J. Oncol. 2005, 26, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.F.; Wu, C.H.; Yang, L.L. Antitumour and free radical scavenging effects of γ-mangostin isolated from Garcinia mangostana pericarps against hepatocellular carcinoma cell. J. Pharm. Pharmacol. 2013, 65, 1419–1428. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wei, Y.; Yuan, S.; Liu, G.; Zhang, Y.L.; Wang, W. Potential anticancer activity of litchi fruit pericarp extract against hepatocellular carcinoma in vitro and in vivo. Cancer Lett. 2006, 239, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Hara, A.; Sakata, K.; Yamada, Y.; Kuno, T.; Kitaori, N.; Oyama, T.; Hirose, Y.; Murakami, A.; Tanaka, T.; Mori, H. Suppression of β-catenin mutation by dietary exposure of auraptene, a citrus antioxidant, in N,N-diethylnitrosamine-induced hepatocellular carcinomas in rats. Oncol. Rep. 2005, 14, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Kalra, N.; Shukla, Y. Hepatoprotective effects of lupeol and mango pulp extract of carcinogen induced alteration in Swiss albino mice. Mol. Nutr. Food Res. 2007, 51, 352–359. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Liu, R.H. Cranberry phytochemicals: Isolation, structure elucidation, and their antiproliferative and antioxidant activities. J. Agric. Food Chem. 2006, 54, 7069–7074. [Google Scholar] [CrossRef] [PubMed]
- Luo, A.; Wang, F.; Luo, D.; Hu, D.; Mao, P.; Xie, W.; He, X.; Kan, W.; Wang, Y. Consumption of vegetables may reduce the risk of liver cancer: Results from a meta-analysis of case-control and cohort studies. Clin. Res. Hepatol. Gas. 2015, 39, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Hanlon, P.R.; Barnes, D.M. Phytochemical composition and biological activity of 8 varieties of radish (Raphanus sativus L.) sprouts and mature taproots. J. Food Sci. 2011, 76, C185–C192. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.T.; Yen, G.C. Effect of vegetables on human phenolsulfotransferases in relation to their antioxidant activity and total phenolics. Free Radic. Res. 2005, 39, 893–904. [Google Scholar] [CrossRef] [PubMed]
- Hanlon, P.R.; Webber, D.M.; Barnes, D.M. Aqueous extract from Spanish black radish (Raphanus sativus L. Var. niger) induces detoxification enzymes in the HepG2 human hepatoma cell line. J. Agric. Food Chem. 2007, 55, 6439–6446. [Google Scholar] [CrossRef] [PubMed]
- Abdull, R.A.; De Nicola, G.R.; Pagnotta, E.; Iori, R.; Ioannides, C. 4-Methylsulfanyl-3-butenyl isothiocyanate derived from glucoraphasatin is a potent inducer of rat hepatic phase II enzymes and a potential chemopreventive agent. Arch. Toxicol. 2012, 86, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Anwar-Mohamed, A.; El-Kadi, A.O. Sulforaphane induces CYP1A1 mRNA, protein, and catalytic activity levels via an AhR-dependent pathway in murine hepatoma Hepa 1c1c7 and human HepG2 cells. Cancer Lett. 2009, 275, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.S.; Jeffery, E.H. Induction of quinone reductase by sulforaphane and sulforaphane N-acetylcysteine conjugate in murine hepatoma cells. J. Med. Food 2005, 8, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.S.; Lee, H.J. Induction of quinone reductase by allylisothiocyanate (AITC) and the N-acetylcysteine conjugate of AITC in Hepa1c1c7 mouse hepatoma cells. Biofactors 2006, 26, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.S.; Lee, H.J. Allyl isothiocyanate and its N-acetylcysteine conjugate suppress metastasis via inhibition of invasion, migration, and matrix metalloproteinase-2/-9 activities in SK-Hep 1 human hepatoma cells. Exp. Biol. Med. (Maywood) 2006, 231, 421–430. [Google Scholar] [PubMed]
- Pasko, P.; Bukowska-Strakova, K.; Gdula-Argasinska, J.; Tyszka-Czochara, M. Rutabaga (Brassica napus L. var. napobrassica) seeds, roots, and sprouts: A novel kind of food with antioxidant properties and proapoptotic potential in HepG2 hepatoma cell line. J. Med. Food 2013, 16, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Spanou, C.; Stagos, D.; Aligiannis, N.; Kouretas, D. Influence of potent antioxidant leguminosae family plant extracts on growth and antioxidant defense system of Hep2 cancer cell line. J. Med. Food 2010, 13, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; He, X.; Liu, R.H. Phytochemicals of black bean seed coats: Isolation, structure elucidation, and their antiproliferative and antioxidative activities. J. Agric. Food Chem. 2007, 55, 6044–6051. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.K.; Ng, T.B. Isolation and characterization of a French bean hemagglutinin with antitumor, antifungal, and anti-HIV-1 reverse transcriptase activities and an exceptionally high yield. Phytomedicine 2010, 17, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Fang, E.F.; Pan, W.L.; Wong, J.H.; Chan, Y.S.; Ye, X.J.; Ng, T.B. A new Phaseolus vulgaris lectin induces selective toxicity on human liver carcinoma Hep G2 cells. Arch. Toxicol. 2011, 85, 1551–1563. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Wan, C.T.; Ng, T.B. Characterisation of a haemagglutinin from Hokkaido red bean (Phaseolus vulgaris cv. Hokkaido red bean). J. Sci. Food Agric. 2010, 90, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ausman, L.M.; Greenberg, A.S.; Russell, R.M.; Wang, X.D. Dietary lycopene and tomato extract supplementations inhibit nonalcoholic steatohepatitis-promoted hepatocarcinogenesis in rats. Int. J. Cancer 2010, 126, 1788–1796. [Google Scholar] [CrossRef] [PubMed]
- Koul, A.; Arora, N.; Tanwar, L. Lycopene mediated modulation of 7,12-Dimethlybenz(α) anthracene induced hepatic clastogenicity in male Balb/c mice. Nutr. Hosp. 2010, 25, 304–310. [Google Scholar] [PubMed]
- Gupta, P.; Bansal, M.P.; Koul, A. Lycopene modulates initiation of N-nitrosodiethylamine induced hepatocarcinogenesis: Studies on chromosomal abnormalities, membrane fluidity and antioxidant defense system. Chem. Biol. Interact. 2013, 206, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Bansal, M.P.; Koul, A. Evaluating the effect of lycopene from Lycopersicum esculentum on apoptosis during NDEA induced hepatocarcinogenesis. Biochem. Biophys. Res. Commun. 2013, 434, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Bansal, M.P.; Koul, A. Spectroscopic characterization of lycopene extract from Lycopersicum esculentum (Tomato) and its evaluation as a chemopreventive agent against experimental hepatocarcinogenesis in mice. Phytother. Res. 2013, 27, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.; Levin, C.E.; Lee, S.U.; Kim, H.J.; Lee, I.S.; Byun, J.O.; Kozukue, N. Tomatine-containing green tomato extracts inhibit growth of human breast, colon, liver, and stomach cancer cells. J. Agric. Food Chem. 2009, 57, 5727–5733. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Xiang, Y.; Lin, S.; Xin, D.; Liu, X.; Weng, L.; Chen, T.; Zhang, M. Anticancer effects of deproteinized asparagus polysaccharide on hepatocellular carcinoma in vitro and in vivo. Tumour Biol. 2014, 35, 3517–3524. [Google Scholar] [CrossRef] [PubMed]
- Weng, L.L.; Xiang, J.F.; Lin, J.B.; Yi, S.H.; Yang, L.T.; Li, Y.S.; Zeng, H.T.; Lin, S.M.; Xin, D.W.; Zhao, H.L.; et al. Asparagus polysaccharide and gum with hepatic artery embolization induces tumor growth and inhibits angiogenesis in an orthotopic hepatocellular carcinoma model. Asian Pac. J Cancer. Prev. 2014, 15, 10949–10955. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Lin, Y.; Kong, L. Steroids from the roots of Asparagus officinalis and their cytotoxic activity. J. Integr. Plant Biol. 2008, 50, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Huang, X.F.; Qi, Q.; Dai, Q.S.; Yang, L.; Nie, F.F.; Lu, N.; Gong, D.D.; Kong, L.Y.; Guo, Q.L. Asparanin A induces G2/M cell cycle arrest and apoptosis in human hepatocellular carcinoma HepG2 cells. Biochem. Biophys. Res. Commun. 2009, 381, 700–705. [Google Scholar] [CrossRef] [PubMed]
- Hafidh, R.R.; Abdulamir, A.S.; Bakar, F.A.; Jalilian, F.A.; Abas, F.; Sekawi, Z. Novel molecular, cytotoxical, and immunological study on promising and selective anticancer activity of mung bean sprouts. BMC Complement. Altern. Med. 2012, 12, 208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Z.; Fang, E.F.; Zhang, H.T.; Liu, L.L.; Yun, J.P. Momordica charantia lectin exhibits antitumor activity towards hepatocellular carcinoma. Investig. New Drugs 2015, 33, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fang, E.F.; Zhang, C.Z.; Wong, J.H.; Shen, J.Y.; Li, C.H.; Ng, T.B. The MAP30 protein from bitter gourd (Momordica charantia) seeds promotes apoptosis in liver cancer cells in vitro and in vivo. Cancer Lett. 2012, 324, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Fang, E.F.; Zhang, C.Z.; Zhang, L.; Fong, W.P.; Ng, T.B. In vitro and in vivo anticarcinogenic effects of RNase MC2, a ribonuclease isolated from dietary bitter gourd, toward human liver cancer cells. Int. J. Biochem. Cell Biol. 2012, 44, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, X.; Han, J.; Zheng, W.; Ma, W. Extract of Perilla frutescens inhibits tumor proliferation of HCC via PI3K/AKT signal pathway. Afr. J. Tradit. Complement. Alt. Med. 2013, 10, 251–257. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, Q.; He, M.; Mir, P.; Su, J.; Yang, Q. Inhibitory effect of antioxidant extracts from various potatoes on the proliferation of human colon and liver cancer cells. Nutr. Cancer 2011, 63, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.; Lee, K.R.; Kim, H.J.; Lee, I.S.; Kozukue, N. Anticarcinogenic effects of glycoalkaloids from potatoes against human cervical, liver, lymphoma, and stomach cancer cells. J. Agric. Food Chem. 2005, 53, 6162–6169. [Google Scholar] [CrossRef] [PubMed]
- Sultana, S.; Ahmed, S.; Jahangir, T.; Sharma, S. Inhibitory effect of celery seeds extract on chemically induced hepatocarcinogenesis: Modulation of cell proliferation, metabolism and altered hepatic foci development. Cancer Lett. 2005, 221, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Arnault, I.; Haffner, T.; Siess, M.H.; Vollmar, A.; Kahane, R.; Auger, J. Analytical method for appreciation of garlic therapeutic potential and for validation of a new formulation. J. Pharm. Biomed. Anal. 2005, 37, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Iciek, M.; Kwiecien, I.; Chwatko, G.; Sokolowska-Jezewicz, M.; Kowalczyk-Pachel, D.; Rokita, H. The effects of garlic-derived sulfur compounds on cell proliferation, caspase 3 activity, thiol levels and anaerobic sulfur metabolism in human hepatoblastoma HepG2 cells. Cell Biochem. Funct. 2012, 30, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Belloir, C.; Siess, M.H.; Le Bon, A.M. Inhibition of carcinogen-induced DNA damage in rat liver and colon by garlic powders with varying alliin content. Nutr. Cancer 2006, 55, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Belloir, C.; Singh, V.; Daurat, C.; Siess, M.H.; Le Bon, A.M. Protective effects of garlic sulfur compounds against DNA damage induced by direct- and indirect-acting genotoxic agents in HepG2 cells. Food Chem. Toxicol. 2006, 44, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.S.; Ko, M.; Ishizuka, M.; Fujita, S.; Yabuki, A.; Hossain, M.A.; Yamato, O. Sodium 2-propenyl thiosulfate derived from garlic induces phase II detoxification enzymes in rat hepatoma H4IIE cells. Nutr. Res. 2010, 30, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.T.; Guo, D.Y.; Cheng, Q.; Geng, W.; Ling, C.C.; Li, C.X.; Liu, X.B.; Ma, Y.Y.; Lo, C.M.; Poon, R.T.; et al. A garlic derivative, S-allylcysteine (SAC), suppresses proliferation and metastasis of hepatocellular carcinoma. PLoS ONE 2012, 7, e31655. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.L.; Ho, C.T.; Chung, J.G.; Raghu, R.; Lo, Y.C.; Sheen, L.Y. Allicin induces anti-human liver cancer cells through the p53 gene modulating apoptosis and autophagy. J. Agric. Food Chem. 2013, 61, 9839–9848. [Google Scholar] [CrossRef] [PubMed]
- De Martino, A.; Filomeni, G.; Aquilano, K.; Ciriolo, M.R.; Rotilio, G. Effects of water garlic extracts on cell cycle and viability of HepG2 hepatoma cells. J. Nutr. Biochem. 2006, 17, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Uda, N.; Kashimoto, N.; Sumioka, I.; Kyo, E.; Sumi, S.; Fukushima, S. Aged garlic extract inhibits development of putative preneoplastic lesions in rat hepatocarcinogenesis. J. Nutr. 2006, 136, 855S–860S. [Google Scholar] [PubMed]
- Ishikawa, H.; Saeki, T.; Otani, T.; Suzuki, T.; Shimozuma, K.; Nishino, H.; Fukuda, S.; Morimoto, K. Aged garlic extract prevents a decline of NK cell number and activity in patients with advanced cancer. J. Nutr. 2006, 136, 816S–820S. [Google Scholar] [PubMed]
- Kim, J.; Ha, H.L.; Moon, H.B.; Lee, Y.W.; Cho, C.K.; Yoo, H.S.; Yu, D.Y. Chemopreventive effect of Curcuma longa Linn on liver pathology in HBx transgenic mice. Integr. Cancer Ther. 2011, 10, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Sreepriya, M.; Bali, G. Chemopreventive effects of embelin and curcumin against N-nitrosodiethylamine/phenobarbital-induced hepatocarcinogenesis in Wistar rats. Fitoterapia 2005, 76, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Strofer, M.; Jelkmann, W.; Depping, R. Curcumin decreases survival of Hep3B liver and MCF-7 breast cancer cells: The role of HIF. Strahlenther. Onkol. 2011, 187, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Shoji, M.; Nakagawa, K.; Watanabe, A.; Tsuduki, T.; Yamada, T.; Kuwahara, S.; Kimura, F.; Miyazawa, T. Comparison of the effects of curcumin and curcumin glucuronide in human hepatocellular carcinoma HepG2 cells. Food Chem. 2014, 151, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shi, X.; Zhang, J.; Zhang, X.; Martin, R.C. Hepatic protection and anticancer activity of curcuma: A potential chemopreventive strategy against hepatocellular carcinoma. Int. J. Oncol. 2014, 44, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.B.; Wu, L.C.; Hsieh, Y.C.; Wu, C.H.; Chan, Y.J.; Chang, L.H.; Chang, C.M.; Hsu, S.L.; Teng, C.L.; Wu, C.C. Supercritical carbon dioxide extraction of aromatic turmerone from Curcuma longa L. inn. induces apoptosis through reactive oxygen species-triggered intrinsic and extrinsic pathways in human hepatocellular carcinoma HepG2 cells. J. Agric. Food Chem. 2012, 60, 9620–9630. [Google Scholar] [CrossRef] [PubMed]
- Lizcano, L.J.; Siles, M.; Trepiana, J.; Hernandez, M.L.; Navarro, R.; Ruiz-Larrea, M.B.; Ruiz-Sanz, J.I. Piper and vismia species from Colombian Amazonia differentially affect cell proliferation of hepatocarcinoma cells. Nutrients 2015, 7, 179–195. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, S.J.; Lim, K.T. ZPDC glycoprotein (24 kDa) induces apoptosis and enhances activity of NK cells in N-nitrosodiethylamine-injected Balb/c. Cell Immunol. 2014, 289, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Dung, T.D.; Feng, C.C.; Kuo, W.W.; Pai, P.; Chung, L.C.; Chang, S.H.; Hsu, H.H.; Tsai, F.J.; Lin, Y.M.; Huang, C.Y. Suppression of plasminogen activators and the MMP-2/-9 pathway by a Zanthoxylum avicennae extract to inhibit the HA22T human hepatocellular carcinoma cell migration and invasion effects in vitro and in vivo via phosphatase 2A activation. Biosci. Biotechnol. Biochem. 2013, 77, 1814–1821. [Google Scholar] [CrossRef] [PubMed]
- Dung, T.D.; Chang, H.C.; Binh, T.V.; Lee, M.R.; Tsai, C.H.; Tsai, F.J.; Kuo, W.W.; Chen, L.M.; Huang, C.Y. Zanthoxylum avicennae extracts inhibit cell proliferation through protein phosphatase 2A activation in HA22T human hepatocellular carcinoma cells in vitro and in vivo. Int. J. Mol. Med. 2012, 29, 1045–1052. [Google Scholar] [PubMed]
- Dung, T.D.; Chang, H.C.; Chen, C.Y.; Peng, W.H.; Tsai, C.H.; Tsai, F.J.; Kuo, W.W.; Chen, L.M.; Huang, C.Y. Zanthoxylum avicennae extracts induce cell apoptosis through protein phosphatase 2A activation in HA22T human hepatocellular carcinoma cells and block tumor growth in xenografted nude mice. Int. J. Mol. Med. 2011, 28, 927–936. [Google Scholar] [PubMed]
- Paik, S.Y.; Koh, K.H.; Beak, S.M.; Paek, S.H.; Kim, J.A. The essential oils from Zanthoxylum schinifolium pericarp induce apoptosis of HepG2 human hepatoma cells through increased production of reactive oxygen species. Biol. Pharm. Bull. 2005, 28, 802–807. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, D.; Das, A.; Bhattacharya, A.; Chakrabarti, G. Aqueous extract of ginger shows antiproliferative activity through disruption of microtubule network of cancer cells. Food Chem. Toxicol. 2010, 48, 2872–2880. [Google Scholar] [CrossRef] [PubMed]
- Manju, V.; Nalini, N. Chemopreventive efficacy of ginger, a naturally occurring anticarcinogen during the initiation, post-initiation stages of 1,2 dimethylhydrazine-induced colon cancer. Clin. Chim. Acta 2005, 358, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Park, K.K.; Chun, K.S.; Lee, J.M.; Lee, S.S.; Surh, Y.J. Inhibitory effects of 6-gingerol, a major pungent principle of ginger, on phorbol ester-induced inflammation, epidermal ornithine decarboxylase activity and skin tumor promotion in ICR mice. Cancer Lett. 1998, 129, 139–144. [Google Scholar] [CrossRef]
- Mansour, M.A.; Bekheet, S.A.; Al-Rejaie, S.S.; Al-Shabanah, O.A.; Al-Howiriny, T.A.; Al-Rikabi, A.C.; Abdo, A.A. Ginger ingredients inhibit the development of diethylnitrosoamine induced premalignant phenotype in rat chemical hepatocarcinogenesis model. Biofactors 2010, 36, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Habib, S.H.; Makpol, S.; Abdul, H.N.; Das, S.; Ngah, W.Z.; Yusof, Y.A. Ginger extract (Zingiber officinale) has anti-cancer and anti-inflammatory effects on ethionine-induced hepatoma rats. Clinics 2008, 63, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Vijaya, P.V.; Arul, D.C.S.; Ramkuma, K.M. Induction of apoptosis by ginger in HEp-2 cell line is mediated by reactive oxygen species. Basic Clin. Pharmacol. Toxicol. 2007, 100, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Weng, C.J.; Wu, C.F.; Huang, H.W.; Ho, C.T.; Yen, G.C. Anti-invasion effects of 6-shogaol and 6-gingerol, two active components in ginger, on human hepatocarcinoma cells. Mol. Nutr. Food Res. 2010, 54, 1618–1627. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Liu, T.Z.; Liu, Y.W.; Tseng, W.C.; Liu, R.H.; Lu, F.J.; Lin, Y.S.; Kuo, S.H.; Chen, C.H. 6-shogaol (alkanone from ginger) induces apoptotic cell death of human hepatoma p53 mutant Mahlavu subline via an oxidative stress-mediated caspase-dependent mechanism. J. Agric. Food Chem. 2007, 55, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.S.; Bhatnagar, D. Chemo-preventive effect of Star anise in N-nitrosodiethylamine initiated and phenobarbital promoted hepato-carcinogenesis. Chem. Biol. Interact. 2007, 169, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Tavakkol-Afshari, J.; Brook, A.; Mousavi, S.H. Study of cytotoxic and apoptogenic properties of saffron extract in human cancer cell lines. Food Chem. Toxicol. 2008, 46, 3443–3447. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.; Hamza, A.A.; Bajbouj, K.; Ashraf, S.S.; Daoud, S. Saffron: A potential candidate for a novel anticancer drug against hepatocellular carcinoma. Hepatology 2011, 54, 857–867. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.T.; Wu, J.; Wen, M.; Su, L.J.; Luo, H. Galangin induces apoptosis in hepatocellular carcinoma cells through the caspase 8/t-Bid mitochondrial pathway. J. Asian Nat. Prod. Res. 2012, 14, 626–633. [Google Scholar] [CrossRef] [PubMed]
- An, N.; Zou, Z.M.; Tian, Z.; Luo, X.Z.; Yang, S.L.; Xu, L.Z. Diarylheptanoids from the rhizomes of Alpinia officinarum and their anticancer activity. Fitoterapia 2008, 79, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Liu, T.Z.; Chen, C.H.; Wu, C.C.; Cheng, J.T.; Yiin, S.J.; Shih, M.K.; Wu, M.J.; Chern, C.L. Isoobtusilactone A-induced apoptosis in human hepatoma HepG2 cells is mediated via increased NADPH oxidase-derived reactive oxygen species (ROS) production and the mitochondria-associated apoptotic mechanisms. Food Chem. Toxicol. 2007, 45, 1268–1276. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Yiin, S.J.; Hsu, J.L.; Wang, W.C.; Lin, S.C.; Chern, C.L. Isoobtusilactone A sensitizes human hepatoma HepG2 cells to TRAIL-induced apoptosis via ROS and CHOP-mediated up-regulation of DR5. J. Agric. Food Chem. 2012, 60, 3533–3539. [Google Scholar] [CrossRef] [PubMed]
- Jeurissen, S.M.; Punt, A.; Delatour, T.; Rietjens, I.M. Basil extract inhibits the sulfotransferase mediated formation of DNA adducts of the procarcinogen 1'-hydroxyestragole by rat and human liver S9 homogenates and in HepG2 human hepatoma cells. Food Chem. Toxicol. 2008, 46, 2296–2302. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.; Utan, A.; Speroni, E.; Cervellati, R.; Piva, G.; Prandini, A.; Guerra, M.C. Carnosic acid from rosemary extracts: A potential chemoprotective agent against aflatoxin B1. An in vitro study. J. Appl. Toxicol. 2007, 27, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Limer, J.L.; Speirs, V. Phyto-oestrogens and breast cancer chemoprevention. Breast Cancer Res. 2004, 6, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Kampkotter, A.; Wiegand, C.; Timpel, C.; Rohrdanz, E.; Chovolou, Y.; Kahl, R.; Watjen, W. Increased expression of catalase in human hepatoma cells by the soy isoflavone, daidzein. Basic Clin. Pharmacol. Toxicol. 2008, 102, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Ho, V.S.; Ng, T.B. A Bowman-Birk trypsin inhibitor with antiproliferative activity from Hokkaido large black soybeans. J. Pept. Sci. 2008, 14, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Ng, T.B. A trypsin-chymotrypsin inhibitor with antiproliferative activity from small glossy black soybeans. Planta Med. 2009, 75, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Helferich, W.G.; Andrade, J.E.; Hoagland, M.S. Phytoestrogens and breast cancer: A complex story. Inflammopharmacology 2008, 16, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Kannan, A.; Hettiarachchy, N.; Johnson, M.G.; Nannapaneni, R. Human colon and liver cancer cell proliferation inhibition by peptide hydrolysates derived from heat-stabilized defatted rice bran. J. Agric. Food Chem. 2008, 56, 11643–11647. [Google Scholar] [CrossRef] [PubMed]
- LIU, Z. Grain phytic acid content in japonica rice as affected by cultivar and environment and its relation to protein content. Food Chem. 2005, 89, 49–52. [Google Scholar] [CrossRef]
- Al-Fatlawi, A.A.; Al-Fatlawi, A.A.; Irshad, M.; Zafaryab, M.; Rizvi, M.M.; Ahmad, A. Rice bran phytic acid induced apoptosis through regulation of Bcl-2/Bax and p53 genes in HepG2 human hepatocellular carcinoma cells. Asian Pac. J. Cancer Prev. 2014, 15, 3731–3736. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.F.; Xu, X.R.; Zhang, Y.; Li, D.; Gan, R.Y.; Li, H.B. Phenolic compounds and bioactivities of pigmented rice. Crit. Rev. Food Sci. 2013, 53, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Banjerdpongchai, R.; Wudtiwai, B.; Sringarm, K. Cytotoxic and apoptotic-inducing effects of purple rice extracts and chemotherapeutic drugs on human cancer cell lines. Asian Pac. J. Cancer Prev. 2014, 14, 6541–6548. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, X.; Xue, Y.; Wang, N.; Liu, W. Anti-hepatoma activity and mechanism of corn silk polysaccharides in H22 tumor-bearing mice. Int. J. Biol. Macromol. 2014, 64, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, B.Y.; Jia, Z.X.; Wu, W.J.; Lu, Z.Q. Hepatocellular carcinoma HepG2 cell apoptosis and caspase-8 and Bcl-2 expression induced by injectable seed extract of Coix lacryma-jobi. Hepatob. Pancreatic. Dis. Int. 2011, 10, 303–307. [Google Scholar] [CrossRef]
- Kim, S.H.; Cui, C.B.; Kang, I.J.; Kim, S.Y.; Ham, S.S. Cytotoxic effect of buckwheat (Fagopyrum esculentum Moench) hull against cancer cells. J. Med. Food. 2007, 10, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.J.; Deng, G.F.; Xu, X.R.; Wu, S.; Li, S.; Xia, E.Q.; Li, F.; Chen, F.; Ling, W.H.; Li, H.B. Antioxidant capacities, phenolic compounds and polysaccharide contents of 49 edible macro-fungi. Food Funct. 2012, 3, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Ahn, W.S.; Kim, D.J.; Chae, G.T.; Lee, J.M.; Bae, S.M.; Sin, J.I.; Kim, Y.W.; Namkoong, S.E.; Lee, I.P. Natural killer cell activity and quality of life were improved by consumption of a mushroom extract, Agaricus blazei Murill Kyowa, in gynecological cancer patients undergoing chemotherapy. Int. J. Gynecol. Cancer 2004, 14, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Sorimachi, K.; Akimoto, K.; Koge, T. Inhibitory effect of Agaricu blazei Murill components on abnormal collagen fiber formation in human hepatocarcinoma cells. Biosci. Biotechnol. Biochem. 2008, 72, 621–623. [Google Scholar] [CrossRef]
- Wu, M.F.; Lu, H.F.; Hsu, Y.M.; Tang, M.C.; Chen, H.C.; Lee, C.S.; Yang, Y.Y.; Yeh, M.Y.; Chung, H.K.; Huang, Y.P.; et al. Possible reduction of hepatoma formation by Smmu 7721 cells in SCID mice and metastasis formation by B16F10 melanoma cells in C57BL/6 mice by Agaricus blazei Murill extract. Vivo 2011, 25, 399–404. [Google Scholar]
- Angeli, J.P.; Ribeiro, L.R.; Bellini, M.F.; Mantovani, M.S. β-glucan extracted from the medicinal mushroom Agaricus blazei prevents the genotoxic effects of benzo(α)pyrene in the human hepatoma cell line HepG2. Arch. Toxicol. 2009, 83, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.Y.; Hwang, L.S.; Kuo, Y.H.; Shu, C.H.; Sheen, L.Y. Black soybean promotes the formation of active components with antihepatoma activity in the fermentation product of Agaricus blazei. J. Agric. Food Chem. 2008, 56, 9447–9454. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.Y.; Tung, Y.C.; Hwang, L.S.; Sheen, L.Y. Blazeispirol A from Agaricus blazei fermentation product induces cell death in human hepatoma Hep3B cells through caspase-dependent and caspase-independent pathways. J. Agric. Food Chem. 2011, 59, 5109–5116. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.W.; Li, B.; Lai, E.T.; Chen, L.; Huang, J.J.; Cheung, A.L.; Cheung, P.C. Water extract from Pleurotus pulmonarius with antioxidant activity exerts in vivo chemoprophylaxis and chemosensitization for liver cancer. Nutr. Cancer 2014, 66, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Huang, J.J.; Cheung, P.C. Extract of Pleurotus pulmonarius suppresses liver cancer development and progression through inhibition of VEGF-induced PI3K/AKT signaling pathway. PLoS ONE 2012, 7, e34406. [Google Scholar] [CrossRef] [PubMed]
- Mariga, A.M.; Yang, W.J.; Mugambi, D.K.; Pei, F.; Zhao, L.Y.; Shao, Y.N.; Hu, Q. Antiproliferative and immunostimulatory activity of a protein from Pleurotus eryngii. J. Sci. Food Agric. 2014, 94, 3152–3162. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.R.; Ng, T.B.; Li, L.; Fang, J.C.; Jiang, Y.; Wen, T.Y.; Qiao, W.T.; Li, N.; Liu, F. Isolation of a polysaccharide with antiproliferative, hypoglycemic, antioxidant and HIV-1 reverse transcriptase inhibitory activities from the fruiting bodies of the abalone mushroom Pleurotus abalonus. J. Pharm. Pharmacol. 2011, 63, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Yukawa, H.; Ishikawa, S.; Kawanishi, T.; Tamesada, M.; Tomi, H. Direct cytotoxicity of Lentinula edodes mycelia extract on human hepatocellular carcinoma cell line. Biol. Pharm. Bull. 2012, 35, 1014–1021. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Chen, Y.; Wang, M.; Yin, Y.; Pan, Y.; Gu, B.; Yu, G.; Li, Y.; Wong, B.H.; Liang, Y.; et al. A novel lectin from Agrocybe aegerita shows high binding selectivity for terminal N-acetylglucosamine. Biochem. J. 2012, 443, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.W.; Lan, J.L.; Meng, Q.; Cheng, Y.W.; Huang, H.M.; Tsai, J.J. Eosinophil apoptosis induced by fungal immunomodulatory peptide-fve via reducing IL-5α receptor. J. Formos. Med. Assoc. 2007, 106, 36–43. [Google Scholar] [CrossRef]
- Chang, H.H.; Hsieh, K.Y.; Yeh, C.H.; Tu, Y.P.; Sheu, F. Oral administration of an Enoki mushroom protein FVE activates innate and adaptive immunity and induces anti-tumor activity against murine hepatocellular carcinoma. Int. Immunopharmacol. 2010, 10, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.Q.; Chen, Y.; Yan, Y.X.; Zhao, J.X. Iso-suillin isolated from Suillus luteus, induces G1 phase arrest and apoptosis in human hepatoma SMMC-7721 cells. Asian Pac. J. Cancer Prev. 2014, 15, 1423–1428. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.T.; Liu, W.H. o-Orsellinaldehyde from the submerged culture of the edible mushroom Grifola frondosa exhibits selective cytotoxic effect against Hep 3B cells through apoptosis. J. Agric. Food Chem. 2006, 54, 7564–7569. [Google Scholar] [CrossRef] [PubMed]
- Norikura, T.; Fujiwara, K.; Yanai, T.; Sano, Y.; Sato, T.; Tsunoda, T.; Kushibe, K.; Todate, A.; Morinaga, Y.; Iwai, K.; Matsue, H. p-terphenyl derivatives from the mushroom Thelephora aurantiotincta suppress the proliferation of human hepatocellular carcinoma cells via iron chelation. J. Agric. Food Chem. 2013, 61, 1258–1264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Bai, D.C.; Wu, Y.J.; Li, W.G.; Liu, N.F. Proanthocyanidin from grape seeds enhances anti-tumor effect of doxorubicin both in vitro and in vivo. Pharmazie 2005, 60, 533–538. [Google Scholar] [PubMed]
- Liang, G.; Tang, A.; Lin, X.; Li, L.; Zhang, S.; Huang, Z.; Tang, H.; Li, Q.Q. Green tea catechins augment the antitumor activity of doxorubicin in an in vivo mouse model for chemoresistant liver cancer. Int. J. Oncol. 2010, 37, 111–123. [Google Scholar] [PubMed]
- Lee, J.S.; Hong, E.K. Agaricus blazei Murill enhances doxorubicin-induced apoptosis in human hepatocellular carcinoma cells by NFκB-mediated increase of intracellular doxorubicin accumulation. Int. J. Oncol. 2011, 38, 401–408. [Google Scholar] [PubMed]
- Lee, J.S.; Hong, E.K. Hericium erinaceus enhances doxorubicin-induced apoptosis in human hepatocellular carcinoma cells. Cancer Lett. 2010, 297, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Karabulut, A.B.; Karadag, N.; Gurocak, S.; Kiran, T.; Tuzcu, M.; Sahin, K. Apricot attenuates oxidative stress and modulates of Bax, Bcl-2, caspases, NFκ-B, AP-1, CREB expression of rats bearing DMBA-induced liver damage and treated with a combination of radiotherapy. Food Chem. Toxicol. 2014, 70, 128–133. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Natural Products | Bioactive Components | Study Type | Bioactivities and Potential Mechanisms | References |
|---|---|---|---|---|
| Fruits | ||||
| Grape | procyanidins | in vitro | selective cytotoxicity to cancer cells | [13] |
| in vivo | inhibiting tumor angiogenesis; promoting doxorubicin induced apoptosis | [19,144] | ||
| Flavan-3-ol | in vitro | inducing apoptosis, DNA damage and Suppressing expression of oncoprotein Her-2 | [20] | |
| Black currant | anthocyanins | in vivo | protecting against diethylnitrosamine induced hepatocarcinogenesis by inducting apoptosis and suppressing oxidative stress and inflammation | [10] |
| Plum | polyphenols | in vitro | inducing extrinsic apoptosis and inhibiting migration | [11,25] |
| in vivo | protecting against B(α)P liver damage through regulating enzymes involved detoxification | [26] | ||
| Pomegranate | polyphenols | in vivo | protecting against diethylnitrosamine induced hepatocarcinogenesis by suppressing oxidative stress and inflammatory responses | [27,28] |
| Apple | polyphenols | in vitro | inducing apoptosis, G2/M cell cycle arrest and inhibiting DNA topoisomerase II in cancer cells | [31] |
| Sweetsop | annonaceous acetogenins | in vitro and in vivo | exerting cytotoxicity against HepG2 cells and inhibiting tumor growth in hepatoma bearing mice | [32] |
| Sea buckthorn | isorhamnetin | in vitro | promoting apoptosis of human hepatoma cells | [34] |
| Gac fruit | a water soluble protein | in vitro | inducing S phase arrest in cancer cells | [35] |
| Mangosteen | γ-mangostin | in vitro | inducing apoptosis in cancer cells | [36] |
| Citrus fruit | auraptene | in vivo | suppressing tumor progression in N,N-diethylnitrosamine challenged rats by negative selection for cancer cells with β-catenin mutation | [38] |
| Mango | lupeol | in vivo | ameliorating DMBA insult induced alterations in liver | [39] |
| Vegetables | ||||
| Radish | Glucoraphasa-tin, isothiocyanate | in vitro | upregulating hepatic phase II detoxification enzymes involved in the metabolism of chemical carcinogens | [45] |
| Broccoli | sulforaphane | in vitro | upregulating CYP1A1 and quinone reductase | [46,47] |
| Rutabaga | NA | in vitro | exerting selective antiproliferative and pro-apoptotic effects in cancer cells | [50] |
| French bean | triterpenoids, flavonoids | in vitro | exhibiting antiproliferative activities against cancer cells | [52] |
| lectins | in vitro | exerting selectively cytotoxicity to cancer cells via promoting apoptosis, necrosis, NO production and release of proinflammatory cytokines | [54] | |
| Tomato | lycopene | in vivo | protecting against chemical induced liver carcinogenesis through inducing apoptosis | [59,60] |
| tomatine | in vivo | inducing antigen-specific cellular immunity and direct destructing cancer cell membranes | [61] | |
| Asparagus | polysaccharides | in vitro and in vivo | selectively inhibiting cancer cell proliferation and enhancing the tumoricidal activities of mitomycin | [62] |
| in vivo | serving as a good embolic candidate in transcatheter arterial chemoembolization | [63] | ||
| asparanin A | in vitro | inducing G2/M cell cycle arrest and apoptosis | [65] | |
| Mung bean sprouts | NA | in vitro | increasing apoptosis, anti-tumor cytokines (TNF-α and IFN-β), IFN-γ production and upregulating cell-mediated immunity | [66] |
| Bitter gourd | lectin | in vitro | inducing G2/M phase arrest, apoptosis, autophagy and enhancing the anti-tumor effects of Sorafenib | [67] |
| MAP30 | in vitro and in vivo | inducing apoptosis and S phase cell cycle arrest | [68] | |
| Purple perilla | Isoegomake-tone | in vitro and in vivo | inhibiting cell growth and xenograft tumor formation probably through blocking the PI3K/Akt signaling pathway | [70] |
| Potato | glycoalkaloids | in vitro | selectively inhibiting cancer cell growth | [72] |
| Celery | pigenin, linamarose, Vitamins A/C | in vivo | dose dependently suppressing chemically induced hepatocarcinogenesis | [73] |
| Spices | ||||
| Garlic | Organo-sulphur compounds | in vitro | inhibiting chemical induced DNA damage | [77] |
| sodium 2-propenyl thiosulfate | in vitro | upregulating quinone reductase | [78] | |
| allicin | in vitro | inducing apoptosis through overproduction of ROS | [80] | |
| S-allylcysteine | in vitro and in vivo | inducing apoptosis and S phase arrest, inhibiting cancer cell migration and invasion | [78] | |
| aged garlic extract | in vivo | inhibiting diethylnitrosamine induced preneoplastic lesions in liver | [81] | |
| clinical trial | enhancing natural-killer cell activities | [83] | ||
| Turmeric | NA | in vivo | protecting against HBV-related liver cancer | [84] |
| curcumin | in vitro and in vivo | demonstrating anti-tumor activity against chemical induced hepatocarcinogenesis | [85] | |
| Sesquiterpe-noids | in vitro and in vivo | alleviating concanavalin A induced oxidative stress and inflammation, inhibiting cancer cell growth | [88] | |
| aromatic tumerone | in vitro | inducing apoptotic cell death via ROS-mediated ERK and JNK kinases activation | [89] | |
| Pepper | NA | in vitro | selective cytotoxicity against rat hepatoma cells through intracellular accumulation of ROS | [90] |
| glycoprotein | in vivo | preventing chemical induced liver carcinogenesis by immunomodulation and promotion of apoptosis | [91] | |
| NA | in vitro | inducing G2/M cell cycle arrest and apoptosis, inhibiting cell metastasis, invasion via down-regulating MMP-2/-9 and up-regulating TIMP-1/-2 | [92,93,94] | |
| geranyl acetate, citronella, sabinene | in vitro and in vivo | increasing apoptotic cell death through ROS production | [95] | |
| Ginger | NA | in vivo | inhibiting inflammation and promoting apoptosis | [100] |
| geraniol, pinostrobin, clavatol | in vitro | inhibiting cancer cell proliferation though ROS-mediated apoptotic death | [101] | |
| 6-shogaol, 6-gingerol | in vitro | suppressing metastasis via down-regulation of MMP-9, urokinase-type plasminogen and up-regulation of TIMP-1 | [102] | |
| Star anise | NA | in vivo | ameliorating tumor burden, oxidative stress and upregulating phase II detoxifying enzymes | [104] |
| Saffron | NA | in vitro and in vivo | inducing apoptosis and decreasing tumor burden, oxidative damage and inflammatory responses | [105,106] |
| Galangal | galagin | in vitro | promoting mitochondrial apoptotic death | [107] |
| Cinnamon | Isoobtusilac-tone A | in vitro | inducing apoptotic cancer cell death through overproduction of ROS | [109,110] |
| Basil | NA | in vitro and in vivo | inhibiting sulfotransferase induced procarcinogenesis by suppressing DNA adducts formation | [111] |
| Rosemary | carnosic acid | in vitro | protecting against aflatoxin B1 through reduced oxidative stress | [112] |
| Soy | trypsin inhibitor | in vitro | inhibiting cancer cell growth | [115,116] |
| Cereals | ||||
| Rice bran | peptide hydrolysates, phytic acid | in vitro | inhibiting cancer cell growth | [118,120] |
| Pigmented rice | anthocyanin | in vitro | synergistically promoting the cytotoxicity of vinblastine through a mitochondrial apoptosis pathway | [122] |
| Corn silk | polysaccharides | in vivo | enhancing immune function and extending survival time | [123] |
| Semen coicis | NA | in vitro | dose-dependently promoting apoptotic death of cancer cells through upregulation of capase-8 | [124] |
| Edible macro-fungi | ||||
| Agaricus blazei | NA | in vitro | sensitizing doxorubicin induced apoptotic death of cancer cells; decreasing formation of abnormal collagen fiber | [128,146] |
| β-glucan | in vitro | protecting against B(α)P induced DNA damage by binding to the carcinogen, scavenging free radicals and probably modulating cell metabolism | [130] | |
| blazeispirols A and C | in vitro | displaying potent antiproliferative activities against Hep3B cells and HepG2 cells | [131] | |
| Pleurotus pulmonarius | NA | in vitro and in vivo | reducing the incidence and size of tumor; inhibiting invasion and drug-resistance of hepatoma cells; enhancing cytotoxicity of cisplatin | [133,134] |
| Lentinula edodes | polysaccharide | in vitro | selectively killing HepG2 cells through the capase-3/-8 mediated extrinsic apoptosis pathway | [137] |
| Agrocybe aegerita | lectin | in vitro | binding to the surface of liver cancer cells, resulting apoptotic cell death | [138] |
| Flammulina velutipes | FVE (glycoprotein) | in vivo | prolonging survival time and reduced tumor size through inducing cytotoxic immune response | [140] |
| Suillus luteus | iso-suillin | in vitro | selectively inducing G1 cell cycle arrest and apoptotic death in cancer cells | [141] |
| Grifola frondosa | O-orsellinaldehyde | in vitro | exhibiting selective potent cytotoxicity against Hep3B cells | [142] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Li, Y.; Zhou, T.; Zheng, J.; Li, S.; Li, H.-B. Dietary Natural Products for Prevention and Treatment of Liver Cancer. Nutrients 2016, 8, 156. https://doi.org/10.3390/nu8030156
Zhou Y, Li Y, Zhou T, Zheng J, Li S, Li H-B. Dietary Natural Products for Prevention and Treatment of Liver Cancer. Nutrients. 2016; 8(3):156. https://doi.org/10.3390/nu8030156
Chicago/Turabian StyleZhou, Yue, Ya Li, Tong Zhou, Jie Zheng, Sha Li, and Hua-Bin Li. 2016. "Dietary Natural Products for Prevention and Treatment of Liver Cancer" Nutrients 8, no. 3: 156. https://doi.org/10.3390/nu8030156
APA StyleZhou, Y., Li, Y., Zhou, T., Zheng, J., Li, S., & Li, H.-B. (2016). Dietary Natural Products for Prevention and Treatment of Liver Cancer. Nutrients, 8(3), 156. https://doi.org/10.3390/nu8030156