Readily Available Sources of Long-Chain Omega-3 Oils: Is Farmed Australian Seafood a Better Source of the Good Oil than Wild-Caught Seafood?

Abstract
:1. Introduction
2. Experimental Section
2.1. Sample Collection
2.2. Lipid Extraction
2.3. Fatty Acid Analysis
3. Results
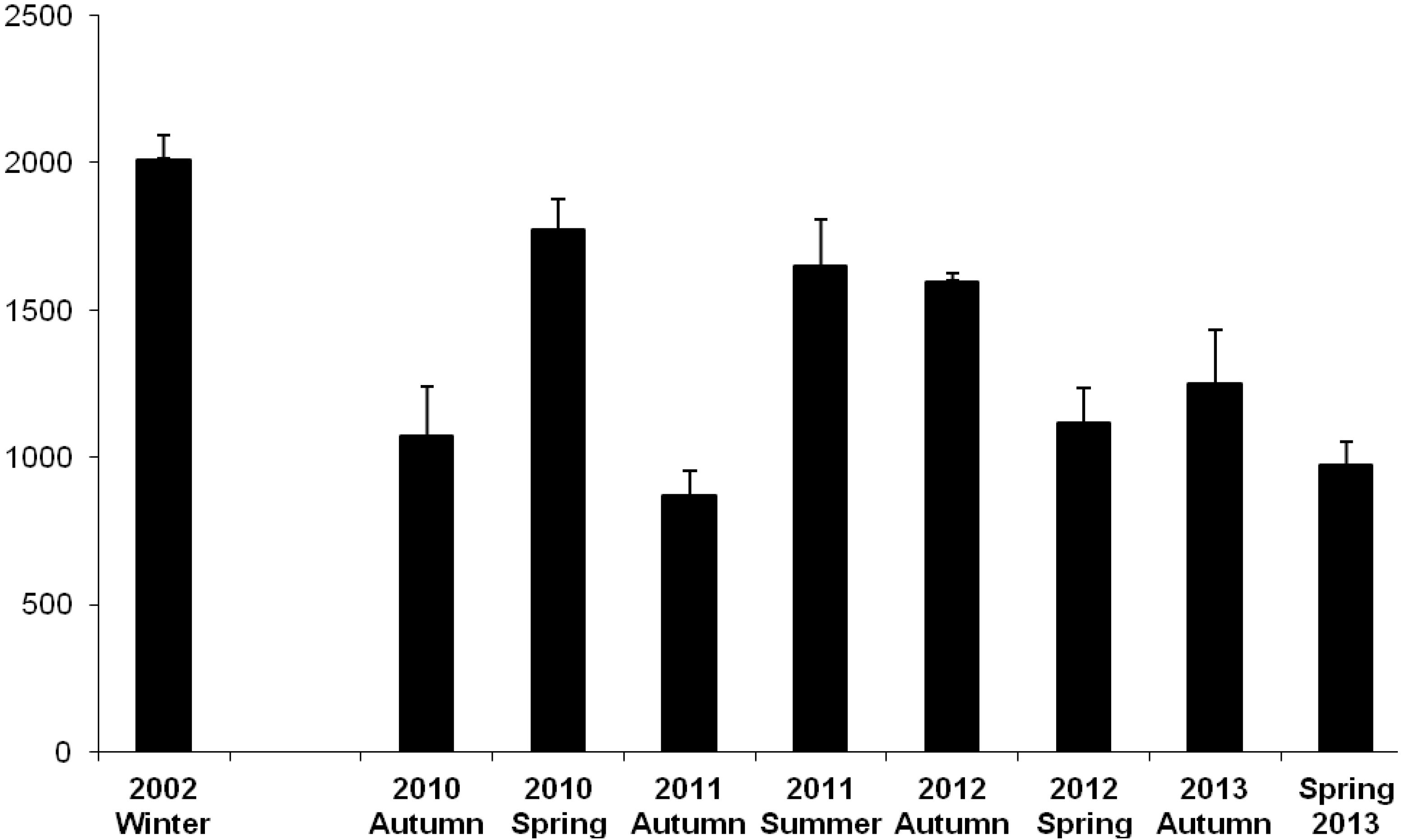
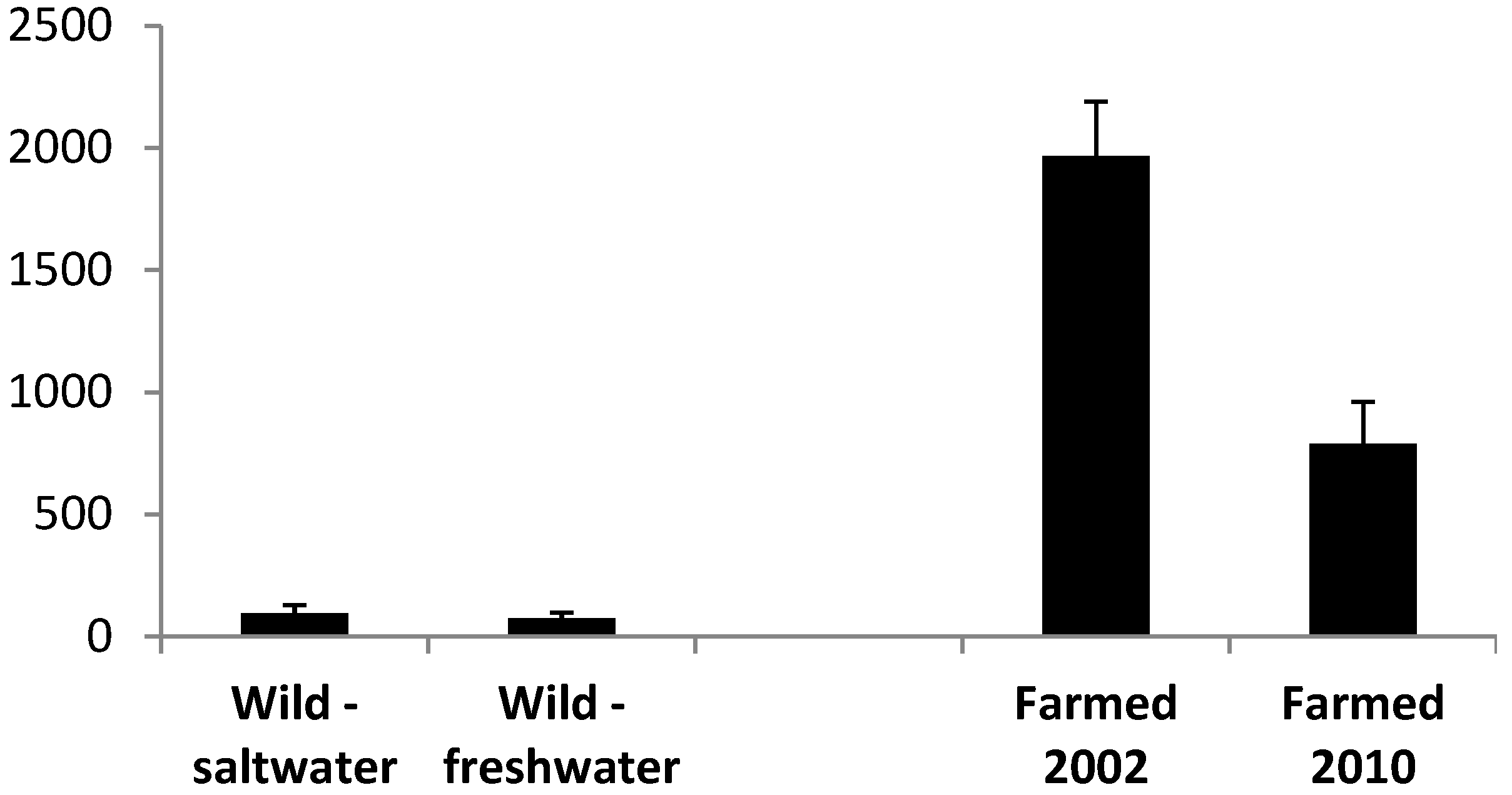
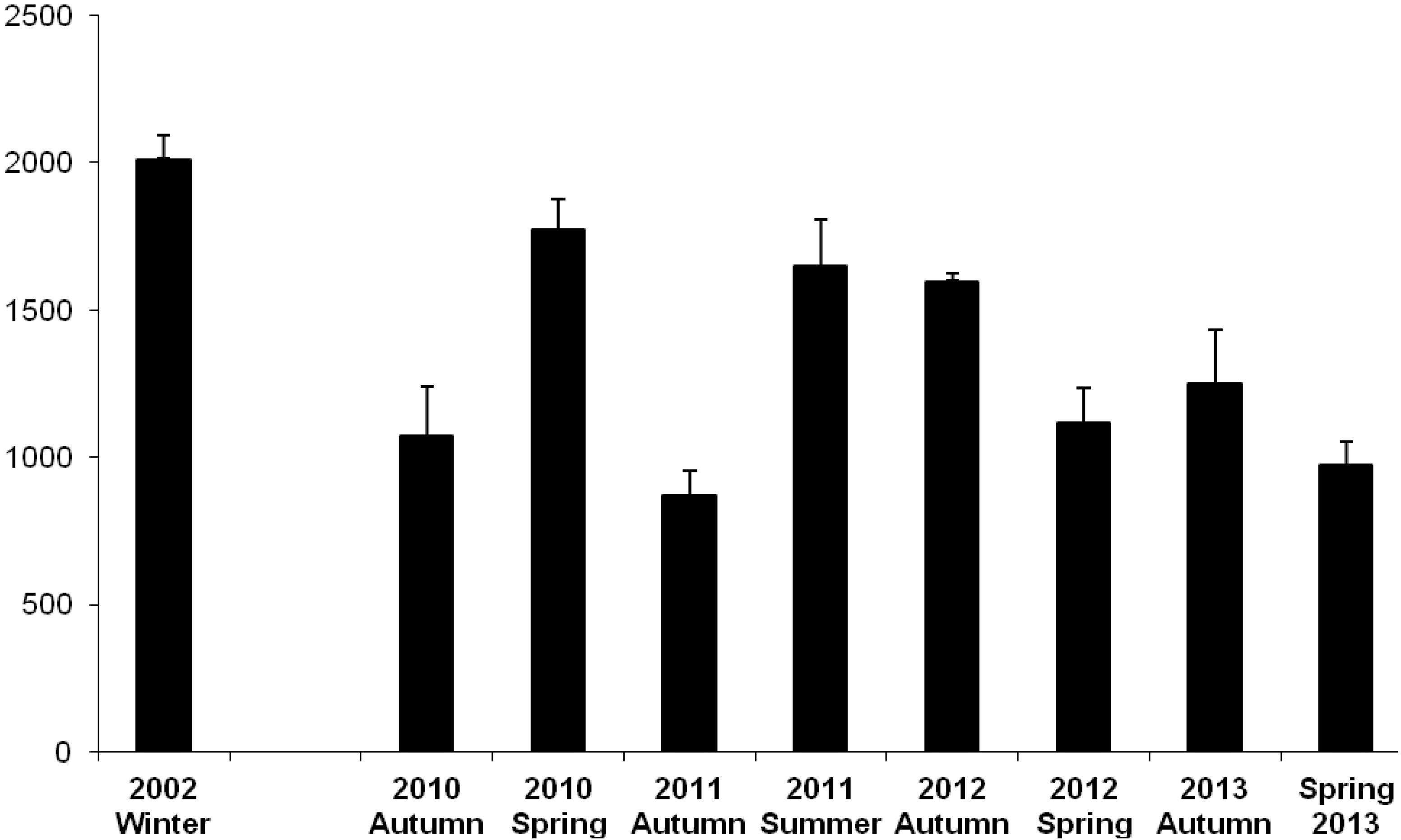
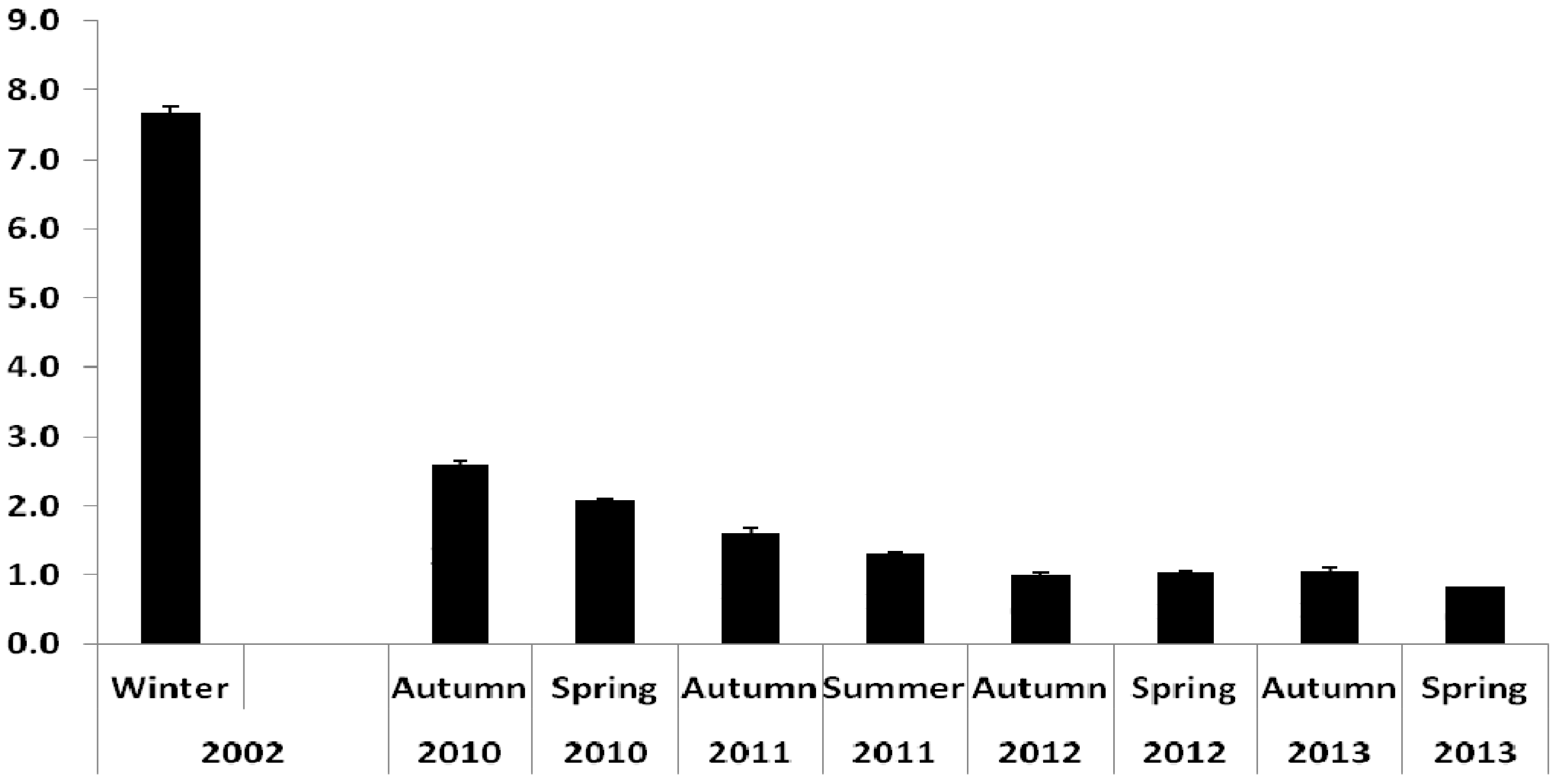
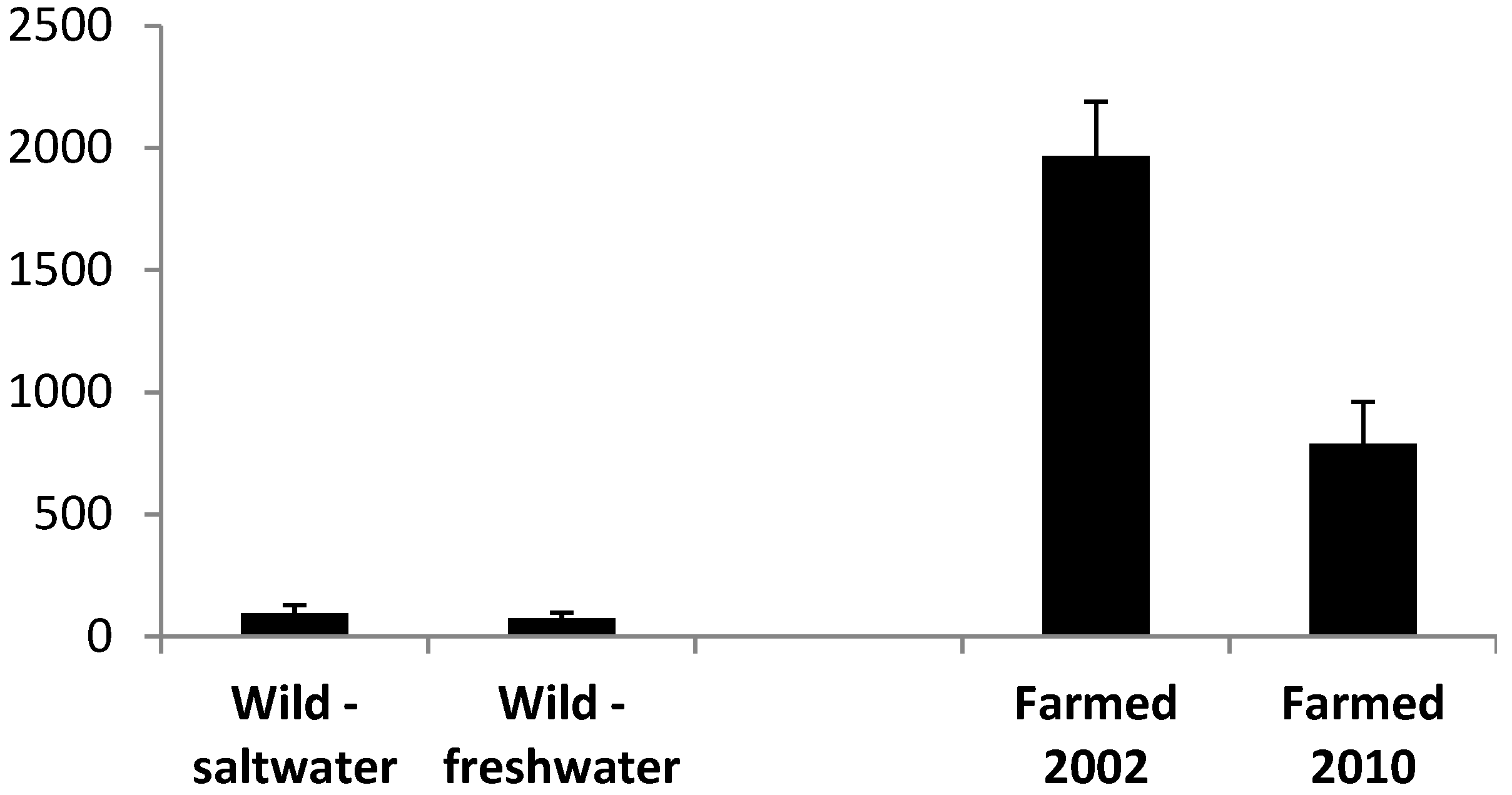
3.1. Oil Content
3.2. Fatty Acid Composition and Content
3.2.1. Farmed Fish

| Fatty acid | Sample Date | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 2002 | 2010 | 2010 | 2011 | 2011 | 2012 | 2012 | 2013 | 2013 | |
| Winter | Autumn | Spring | Autumn | Summer | Autumn | Spring | Autumn | Spring | |
| 14:0 | 5.5 | 2.1 | 2.7 | 2.8 | 2.5 | 2.4 | 1.8 | 1.9 | 1.7 |
| 15:0 | 0.5 | 0.2 | 0.2 | 0.3 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| 16:4 | 0.3 | 0.7 | 0.6 | 0.5 | 0.5 | 0.3 | 0.2 | 0.3 | 0.2 |
| 16:3 | 0.4 | 0.8 | 0.6 | 0.6 | 0.5 | 0.4 | 0.2 | 0.4 | 0.3 |
| 16:1ω7c | 6.2 | 8.3 | 7.0 | 7.4 | 7.2 | 6.4 | 6.8 | 6.9 | 4.9 |
| 16:0 | 18.0 | 14.4 | 15.1 | 13.9 | 15.7 | 15.4 | 15.5 | 14.7 | 13.0 |
| 17:1ω8c + a17:0 | 0.4 | 0.3 | 0.3 | 0.4 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 |
| 17:0 | 0.5 | 0.2 | 0.3 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| 18:3ω6 | 0.1 | 0.1 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| 18:4ω3 | 3.0 | 1.4 | 1.3 | 1.0 | 1.0 | 0.9 | 0.7 | 0.8 | 0.7 |
| 18:2ω6 | 2.8 | 7.2 | 8.2 | 8.4 | 10.0 | 12.8 | 9.6 | 9.8 | 12.4 |
| 18:3ω3 | 0.0 | 0.9 | 1.1 | 1.1 | 1.0 | 0.7 | 1.1 | 0.9 | 2.1 |
| 18:1ω9c | 14.5 | 27.0 | 29.2 | 33.8 | 33.8 | 33.0 | 39.0 | 38.8 | 42.7 |
| 18:1ω7c | 3.3 | 4.0 | 3.9 | 4.0 | 3.5 | 3.1 | 3.5 | 3.4 | 3.4 |
| 18:1ω5c | 0.1 | 0.1 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.1 |
| 18:0 | 4.2 | 3.9 | 4.4 | 4.2 | 4.1 | 4.5 | 4.5 | 4.5 | 4.0 |
| 20:4ω6 | 0.8 | 0.8 | 0.7 | 0.6 | 0.5 | 0.3 | 0.4 | 0.5 | 0.4 |
| 20:5ω3 | 9.6 | 8.5 | 7.4 | 5.9 | 4.9 | 4.4 | 3.1 | 3.4 | 2.7 |
| 20:3ω6 | 0.2 | 0.4 | 0.3 | 0.3 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 |
| 20:4ω3 | 1.5 | 0.9 | 0.8 | 0.7 | 0.6 | 0.5 | 0.4 | 0.5 | 0.5 |
| C20PUFA | 0.0 | 0.0 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.0 |
| 20:2ω6 | 0.3 | 0.4 | 0.5 | 0.5 | 0.5 | 0.7 | 0.6 | 0.5 | 0.0 |
| 20:1ω9c | 1.7 | 1.4 | 1.4 | 1.7 | 1.5 | 1.8 | 1.8 | 1.7 | 1.7 |
| 20:1ω7c | 0.2 | 0.3 | 0.3 | 0.3 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| 20:0 | 0.2 | 0.2 | 0.2 | 0.2 | 0.1 | 0.2 | 0.2 | 0.1 | 0.2 |
| 21:5ω3 | 0.6 | 0.4 | 0.4 | 0.3 | 0.3 | 0.3 | 0.2 | 0.2 | 0.1 |
| 22:5ω6 | 0.3 | 0.1 | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| 22:6ω3 | 17.0 | 7.8 | 6.7 | 5.0 | 5.1 | 5.4 | 4.8 | 4.9 | 4.0 |
| 22:4ω6 | 0.1 | 0.2 | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| 22:5ω3 | 3.8 | 3.8 | 2.8 | 2.5 | 2.2 | 1.9 | 1.4 | 1.6 | 1.2 |
| 22:1ω11c | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.6 | 0.2 | 0.1 | 0.1 |
| 22:1ω9c | 0.2 | 0.1 | 0.1 | 0.2 | 0.1 | 0.2 | 0.2 | 0.2 | 0.2 |
| 24:5ω3 | 0.0 | 0.2 | 0.2 | 0.0 | 0.2 | 0.1 | 0.1 | 0.1 | 0.0 |
| 24:1ω11c | 0.0 | 0.1 | 0.0 | 0.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| 24:1ω9c | 0.5 | 0.2 | 0.2 | 0.0 | 0.2 | 0.2 | 0.2 | 0.1 | 0.2 |
| 24:0 | 0.0 | 0.0 | 0.0 | 0.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Other | 2.7 | 2.0 | 1.8 | 1.5 | 1.4 | 1.4 | 1.4 | 1.5 | 1.4 |
| Sum | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| Sum SFA | 29.4 | 21.9 | 23.4 | 22.4 | 23.2 | 23.3 | 22.7 | 21.9 | 19.7 |
| Sum MUFA | 29.2 | 43.1 | 44.1 | 49.6 | 48.2 | 46.8 | 53.3 | 52.8 | 54.7 |
| Sum Omega-3 PUFA | 35.5 | 24.0 | 20.9 | 16.5 | 15.4 | 14.3 | 11.9 | 12.4 | 11.3 |
| Sum Omega-6 PUFA | 4.6 | 9.3 | 10.0 | 10.4 | 11.8 | 14.6 | 11.6 | 11.8 | 13.6 |

| Fatty Acid | 1998 | 1998 | 2002 | 2010 |
|---|---|---|---|---|
| Freshwater | Saltwater | Farmed | Farmed | |
| 14:0 | 2.8 | 0.9 | 5.9 | 3.6 |
| 15:0 | 0.9 | 0.6 | 0.6 | 0.3 |
| 16:1ω7c | 7.4 | 2.1 | 6.5 | 7.5 |
| 16:0 | 27.8 | 22.4 | 19 | 17.7 |
| 17:1ω8c + a17:0 | 0.9 | 0.7 | 0.5 | 0.3 |
| 18:3ω6 | 0.4 | 0.3 | 0.3 | 0.3 |
| 18:4ω3 | 0.4 | 0.6 | 1.9 | 1.3 |
| 18:2ω6 | 4.8 | 1 | 5.5 | 9.8 |
| 18:3ω3 | 1 | 0.2 | 0 | 1.1 |
| 18:1ω9c | 17.9 | 11.7 | 19.6 | 26.2 |
| 18:1ω7c | 4 | 2.1 | 2.9 | 3.2 |
| 18:0 | 8.8 | 12.5 | 4.4 | 4.8 |
| 20:4ω6 | 7.1 | 12.2 | 0.6 | 0.7 |
| 20:5ω3 | 1.3 | 3.1 | 6.2 | 6.9 |
| 20:3ω6 | 0.5 | 0.2 | 0.1 | 0.2 |
| 20:4ω3 | 0.3 | 0 | 0.8 | 0.5 |
| 20:2ω6 | 0.3 | 0.1 | 0.2 | 0.2 |
| 20:1ω9c | 0.6 | 0.3 | 3.5 | 1.1 |
| 20:1ω7c | 0.1 | 0 | 0.3 | 0.2 |
| 20:0 | 0.3 | 0.2 | 0.2 | 0.3 |
| 22:5ω6 | 0.4 | 0 | 0.3 | 0.2 |
| 22:6ω3 | 6.1 | 21.6 | 10.2 | 5.4 |
| 22:4ω6 | 1.2 | 1.2 | 0.1 | 0.1 |
| 22:5ω3 | 1.6 | 2.2 | 2.5 | 2.1 |
| 22:1ω11c | 0 | 0 | 1.6 | 0.5 |
| 22:1ω9c | 0 | 0 | 0.5 | 0.1 |
| 24:1ω9c | 0.2 | 0.8 | 0.5 | 0.2 |
| Other | 2.9 | 3 | 5.3 | 2.3 |
| Sum SFA | 42.7 | 39.3 | 31.1 | 27.9 |
| Sum MUFA | 31.7 | 17.7 | 37 | 40.7 |
| Sum Omega-3 PUFA | 11.1 | 28.1 | 21.6 | 17.9 |
| Sum Omega-6 PUFA | 14.5 | 14.9 | 7.2 | 10 |
| Omega-3/Omega-6 | 0.8 | 1.9 | 3 | 1.55 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3.2.2. Fish Oil Capsules
| Brand | Description | Lab Code | Capsule Size (mg) | Number of Capsules | Cost ($) | Cost ($) per Capsule | EPA mg per Capsule | DHA mg per Capsule | EPA + DHA mg per Capsule | Number of Caps for 500 mg EPA + DHA |
|---|---|---|---|---|---|---|---|---|---|---|
| Group 1 | ||||||||||
| Blackmores | Omega Joint | FO1 | 1000 | 60 | 22.39 | 0.37 | 550 | 120 | 670 | 1 |
| Blackmores | Omega Brain | FO2 | 1000 | 60 | 22.39 | 0.37 | 100 | 500 | 600 | 1 |
| Healthy Care | Fish oil One a Day | FO3 | 1000 | 50 | 14.99 | 0.30 | 360 | 240 | 600 | 1 |
| Cenovis | Fish oil Plus | FO4 | 1500 | 62 | 16.69 | 0.27 | 335 | 185 | 520 | 2 |
| Natures Own | Omega-3 Ultra | FO5 | 1000 | 60 | 22.69 | 0.38 | 302 | 201 | 503 | 1 |
| Group 2 | ||||||||||
| Cenovis | Fish oil with Omega-3 | FO6 | 1000 | 180 | 13.69 | 0.08 | 180 | 120 | 300 | 2 |
| Blackmores | Odorless Fish Oil + Vitamin D3 | FO7 | 1000 | 100 | 12.99 | 0.13 | 180 | 120 | 300 | 2 |
| Healthy Care | Fish oil | FO8 | 1000 | 400 | 12.99 | 0.03 | 180 | 120 | 300 | 2 |
| Swisse | Odorless Fish Oil | FO9 | 1000 | 200 | 22.99 | 0.11 | 180 | 120 | 300 | 2 |
| Group 3 | ||||||||||
| Swisse | Wild salmon oil | FO10 | 1000 | 200 | 22.99 | 0.11 | 80 | 80 | 160 | 4 |
| Blackmores | Pregnancy | FO11 | 1000 | 120 | 33.99 | 0.28 | 25 | 125 | 150 | 4 |
| Natures Own | Fish oil + Glucosamine | FO12 | 1100 | 90 | 20.99 | 0.23 | 90 | 60 | 150 | 4 |
| Swisse | Wild krill oil (NKO) | FO13 | 333 | 50 | 43.99 | 0.88 | 50 | 30 | 80 | 7 |
| Cenovis | Cod liver oil * | FO14 | 275 | 90 | 3.99 | 0.04 | 23 | 29 | 52 | 10 |
| Sample | Group 1 | Group 2 | Group 3 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FO1 | FO2 | FO3 | FO4 | FO5 | FO6 | FO7 | FO8 | FO9 | FO10 | FO11 | FO12 | FO13 | FO14 | |
| 14:0 | 0.1 | 0.2 | 0.9 | 4.2 | 0.5 | 5.7 | 5.8 | 5.7 | 6.2 | 4.8 | 6.9 | 6.6 | 9.1 | 3.3 |
| 15:0 | 0.0 | 0.1 | 0.0 | 0.3 | 0.0 | 0.5 | 0.4 | 0.5 | 0.5 | 0.4 | 0.7 | 0.4 | 0.4 | 0.3 |
| 16:4 | 2.7 | 0.1 | 0.4 | 2.2 | 0.4 | 2.2 | 2.4 | 2.3 | 2.2 | 0.5 | 0.1 | 1.8 | 1.3 | 0.4 |
| 16:3 | 1.8 | 0.1 | 0.4 | 1.6 | 0.2 | 1.5 | 1.8 | 1.5 | 1.6 | 0.5 | 0.1 | 1.4 | 0.3 | 0.3 |
| 16:1ω7c | 0.5 | 0.8 | 1.7 | 6.9 | 1.4 | 9.6 | 9.4 | 9.6 | 9.7 | 6.8 | 3.8 | 8.0 | 6.8 | 6.6 |
| 16:1ω5c | 0.0 | 0.0 | 0.0 | 0.1 | 0.0 | 0.2 | 0.2 | 0.2 | 0.2 | 0.4 | 0.1 | 0.2 | 0.5 | 0.3 |
| 16:0 | 0.3 | 3.2 | 2.6 | 11.3 | 3.5 | 15.1 | 15.9 | 15.0 | 15.6 | 13.4 | 18.5 | 15.5 | 19.7 | 10.9 |
| i17:0 | 0.0 | 0.1 | 0.0 | 0.1 | 0.0 | 0.2 | 0.5 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.3 | 0.2 |
| 17:1ω8c + a17:0 | 0.0 | 0.2 | 0.1 | 0.2 | 0.1 | 0.4 | 0.4 | 0.4 | 0.4 | 0.5 | 0.6 | 0.3 | 0.4 | 0.7 |
| 17:0 | 0.0 | 0.3 | 0.1 | 0.2 | 0.2 | 0.5 | 0.4 | 0.4 | 0.4 | 0.3 | 0.8 | 0.4 | 0.1 | 0.2 |
| 18:3ω6 | 0.6 | 0.1 | 0.1 | 0.2 | 0.1 | 0.3 | 0.2 | 0.3 | 0.2 | 0.0 | 0.1 | 0.2 | 0.2 | 0.1 |
| 18:4ω3 | 8.1 | 0.7 | 1.5 | 2.2 | 2.0 | 3.3 | 3.3 | 3.3 | 3.3 | 2.4 | 0.8 | 2.7 | 3.9 | 2.3 |
| 18:2ω6 | 0.2 | 1.1 | 0.9 | 2.5 | 1.6 | 1.4 | 1.1 | 1.6 | 1.4 | 2.5 | 8.3 | 7.8 | 2.1 | 2.2 |
| 18:3ω3 | 0.2 | 0.4 | 0.6 | 0.6 | 0.6 | 0.8 | 0.6 | 0.8 | 0.8 | 0.9 | 1.2 | 1.4 | 1.0 | 0.7 |
| 18:1ω9c | 0.2 | 7.7 | 6.6 | 7.8 | 10.5 | 8.5 | 10.0 | 8.4 | 8.4 | 16.8 | 12.7 | 10.0 | 12.6 | 20.0 |
| 18:1ω7c | 0.0 | 1.7 | 2.8 | 3.2 | 3.0 | 3.9 | 3.5 | 3.9 | 4.0 | 4.0 | 2.3 | 3.7 | 8.4 | 4.7 |
| 18:1ω5c | 0.0 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.2 | 0.1 | 0.6 | 0.1 | 0.2 | 0.4 | 0.4 |
| 18:0 | 0.1 | 3.9 | 2.8 | 2.7 | 3.5 | 3.4 | 3.2 | 3.4 | 3.5 | 2.4 | 7.8 | 5.3 | 1.4 | 2.6 |
| 20:4ω6 | 1.2 | 2.6 | 0.8 | 1.0 | 0.8 | 0.8 | 0.7 | 0.6 | 0.6 | 0.4 | 1.5 | 0.7 | 0.2 | 0.3 |
| 20:5ω3 EPA | 60.4 | 14.5 | 35.6 | 27.4 | 31.8 | 18.9 | 20.0 | 19.0 | 18.7 | 9.1 | 5.4 | 15.1 | 16.9 | 8.8 |
| 20:3ω6 | 0.2 | 0.3 | 0.4 | 0.3 | 0.4 | 0.3 | 0.3 | 0.3 | 0.2 | 0.1 | 0.1 | 0.2 | 0.0 | 0.1 |
| 20:4ω3 | 0.9 | 0.9 | 1.8 | 1.2 | 1.5 | 1.0 | 0.9 | 1.0 | 1.0 | 1.2 | 0.4 | 0.8 | 0.4 | 0.8 |
| C20PUFA | 0.1 | 0.1 | 0.8 | 0.4 | 0.4 | 0.3 | 0.3 | 0.3 | 0.3 | 0.1 | 0.0 | 0.3 | 0.0 | 0.1 |
| 20:2ω6 | 0.1 | 0.6 | 0.5 | 0.3 | 0.4 | 0.3 | 0.2 | 0.4 | 0.3 | 0.3 | 0.2 | 0.3 | 0.0 | 0.3 |
| 20:1w11c | 0.0 | 0.6 | 0.4 | 0.2 | 0.3 | 0.2 | 0.1 | 0.2 | 0.2 | 7.2 | 0.4 | 0.0 | 0.0 | 1.2 |
| 20:1ω9c | 0.5 | 2.3 | 2.2 | 1.3 | 2.2 | 1.3 | 0.5 | 1.3 | 1.1 | 3.0 | 1.0 | 1.3 | 0.9 | 8.8 |
| 20:1ω7c | 0.0 | 0.4 | 1.1 | 0.7 | 0.9 | 0.4 | 0.3 | 0.5 | 0.5 | 0.5 | 0.1 | 0.4 | 0.4 | 0.5 |
| 20:0 | 0.0 | 0.7 | 0.6 | 0.3 | 0.5 | 0.3 | 0.2 | 0.3 | 0.3 | 0.3 | 0.4 | 0.4 | 0.1 | 0.1 |
| 21:5ω3 | 1.5 | 0.6 | 1.4 | 1.1 | 1.6 | 0.7 | 0.8 | 0.7 | 0.8 | 0.4 | 0.2 | 0.6 | 0.5 | 0.4 |
| 22:5ω6 | 0.0 | 1.1 | 0.5 | 0.4 | 0.4 | 0.3 | 0.3 | 0.3 | 0.3 | 0.1 | 1.3 | 0.3 | 0.0 | 0.1 |
| 22:6ω3 DHA | 17.7 | 46.8 | 23.2 | 13.2 | 23.2 | 11.8 | 10.9 | 11.6 | 11.7 | 6.9 | 18.8 | 9.4 | 8.2 | 10.3 |
| 22:4ω6 | 0.1 | 0.5 | 0.3 | 0.2 | 0.2 | 0.2 | 0.1 | 0.2 | 0.2 | 0.0 | 0.3 | 0.1 | 0.0 | 0.1 |
| 22:5ω3 | 1.5 | 2.9 | 4.5 | 2.9 | 4.3 | 2.3 | 2.1 | 2.3 | 2.2 | 1.8 | 1.2 | 1.8 | 0.4 | 1.2 |
| 22:1ω11c | 0.0 | 0.6 | 1.6 | 0.9 | 1.0 | 0.7 | 0.1 | 0.7 | 0.6 | 7.6 | 0.5 | 0.5 | 0.0 | 7.5 |
| 22:1ω9c | 0.0 | 0.4 | 0.4 | 0.2 | 0.3 | 0.2 | 0.1 | 0.1 | 0.2 | 0.7 | 0.1 | 0.1 | 0.5 | 0.4 |
| 22:0 | 0.0 | 0.4 | 0.2 | 0.2 | 0.2 | 0.1 | 0.3 | 0.1 | 0.1 | 0.1 | 0.2 | 0.2 | 0.0 | 0.1 |
| 24:6ω3 | 0.0 | 0.2 | 0.1 | 0.1 | 0.1 | 0.1 | 0.0 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| 24:5ω3 | 0.0 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.0 | 0.1 | 0.1 | 0.1 | 0.0 | 0.1 | 0.1 | 0.2 |
| 24:1ω9c | 0.0 | 1.0 | 0.7 | 0.4 | 0.6 | 0.4 | 0.3 | 0.4 | 0.4 | 0.6 | 0.7 | 0.4 | 0.4 | 0.3 |
| 24:0 | 0.0 | 0.3 | 0.1 | 0.0 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.0 | 0.4 | 0.1 | 0.0 | 0.0 |
| Sum other | 1.0 | 1.2 | 0.9 | 1.0 | 1.0 | 1.8 | 2.3 | 1.7 | 1.7 | 2.0 | 1.5 | 0.9 | 1.6 | 2.2 |
| Sum | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Sum EPA + DPA + DHA | 79.7 | 64.3 | 63.2 | 43.5 | 59.4 | 33.0 | 33.0 | 32.9 | 32.6 | 17.8 | 25.4 | 26.3 | 25.5 | 20.2 |
| Omega-3/Omega-6 | 16.6 | 9.3 | 11.2 | 4.9 | 9.9 | 6.8 | 7.4 | 6.7 | 7.4 | 4.5 | 2.0 | 1.9 | 6.2 | 5.8 |
4. Discussion
4.1. Farmed Fish—Oil Content and Fatty Acid Profiles
4.2. Fish Oil Capsules as Sources of EPA + DHA

5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Bang, H.O.; Dyerberg, J.E. Lipid metabolism and ischaemic heart disease in Greenland Eskimos. Adv. Nutr. Res. 1980, 3, 1–22. [Google Scholar] [CrossRef]
- Nichols, P.D.; Petrie, J.; Singh, S. Long-chain omega-3 oils—An update on sustainable sources. Nutrients 2010, 2, 572–585. [Google Scholar] [CrossRef]
- Tacon, A.G.J.; Metian, M. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects. Aquaculture 2008, 285, 146–158. [Google Scholar] [CrossRef]
- Glencross, B.D. Exploring the nutritional demand for essential fatty acids by aquaculture species. Rev. Aquac. 2009, 1, 71–124. [Google Scholar]
- Naylor, R.L.; Hardy, R.W.; Bureau, D.P.; Chiu, A.; Elliott, M.; Farrell, A.P.; Forster, I.; Gatlin, D.M.; Goldburg, R.J.; Hua, K.; et al. Feeding aquaculture in an era of finite resources. Proc. Natl. Acad. Sci. USA 2009, 106, 15103–15110. [Google Scholar] [CrossRef]
- Glencross, B.; Turchini, G.M. Fish Oil Replacement in Starter, Grow-Out and Finishing Feeds for Farmed Aquatic Animals. In Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds; Turchini, G.M., Ng, W.K., Tocher, D.R., Eds.; CRC Publishing Ltd.: Boca Raton, FL, USA, 2010; pp. 373–404. [Google Scholar]
- Turchini, G.M.; Torstensen, B.E.; Ng, W.K. Fish oil replacement in finfish nutrition. Rev. Aquac. 2009, 1, 10–57. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Christie, W.W. Lipid Analysis; Pergamon Press: Oxford, UK, 1982. [Google Scholar]
- Nichols, P.D.; Mooney, B.D.; Elliott, N.G. Nutritional Value of Australian Seafood II. Factors Affecting Oil Composition of Edible Species; Prepared for the Fisheries Research and Development Corporation: Hobart, Australia, September 2002. [Google Scholar]
- Nichols, P.D.; Mooney, B.; Virtue, P.; Elliott, N. Nutritional Value of Australian Fish: Oil, Fatty Acid and Cholesterol of Edible Species; Report Prepared for the Fisheries Research and Development Corporation: Hobart, Australia, August 1998. [Google Scholar]
- Glencross, B.D. Nutritional management of barramundi, Lates calcarifer—A review. Aquac. Nutr. 2006, 12, 291–309. [Google Scholar] [CrossRef]
- Percival, S.; Drabsch, P.; Glencross, B.D. Determining factors affecting muddy-flavour taint in farmed barramundi, Lates calcarifer. Aquaculture 2008, 284, 136–143. [Google Scholar] [CrossRef]
- Glencross, B.D.; Michael, R.; Austen, K.; Hauler, R. Productivity, carcass composition, waste output and sensory characteristics of large barramundi Lates calcarifer fed high-nutrient density diets. Aquaculture 2008, 284, 167–173. [Google Scholar] [CrossRef]
- Ramsden, C.E.; Hibbeln, J.R.; Majchrzak, S.F.; Davis, J.M. n-6 Fatty acid-specific and mixed polyunsaturate dietary interventions have different effects on CHD risk: A meta-analysis of randomized controlled trials. Br. J. Nutr. 2010, 104, 1586–1600. [Google Scholar] [CrossRef]
- Harris, W.S.; Mozaffarian, D.; Rimm, E.; Kris-Etherton, P.; Rudel, L.L.; Appel, L.J.; Engler, M.M.; Engler, M.B.; Sacks, F. n-6 Fatty acids and risk for cardiovascular disease: A science advisory from the American Heart Association Nutrition Subcommittee of the Council on Nutrition. Physical Activity, and Metabolism; Council on Cardiovascular Nursing; and Council on Epidemiology and Prevention. Circulation 2009, 119, 902–907. [Google Scholar] [CrossRef]
- Rosenlund, G.; Corraze, G.; Izquierdo, M.; Torstensen, B. The Effects of Fish oil Replacement on Nutritional and Organoleptic Qualities of Farmed Fish. In Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds; Turchini, G.M., Ng, W.K., Tocher, D.R., Eds.; CRC Publishing Ltd.: Boca Raton, FL, USA, 2010; pp. 487–522. [Google Scholar]
- Rosewarne, G.; New Zealand King Salmon Co., Ltd., Nelson, New Zealand. Personal communication. 2013. [Google Scholar]
- Seierstad, S.L.; Seljeflot, I.; Johansen, O.; Hansen, R.; Haugen, M.; Rosenlund, G.; Frøyland, L.; Arnesen, H. Dietary intake of differently fed salmon; the influence on markers of human atherosclerosis. Eur. J. Clin. Investig. 2005, 35, 52–59. [Google Scholar]
- Codabaccus, B.M.; Carter, C.G.; Bridle, A.R.; Nichols, P.D. The “n-3 LC-PUFA sparing effect” of modified dietary n-3 LC-PUFA content and DHA to EPA ratio in Atlantic salmon smolt. Aquaculture 2012, 356–357, 135–140. [Google Scholar]
- Petrie, J.R.; Shrestha, P.; Mansour, M.P.; Nichols, P.D.; Liu, Q.; Singh, S.P. Metabolic engineering of omega-3 long-chain polyunsaturated fatty acids in plants using an acyl-CoA Δ6-desaturase with ω3-preference from the marine microalga Micromonas pusilla. Metab. Eng. 2010, 12, 233–240. [Google Scholar] [CrossRef]
- Petrie, J.R.; Nichols, P.D.; Devine, M.; Singh, S.P. Engineered oil seed crops with fish oil DHA levels. INFORM 2013, 24, 648–652. [Google Scholar]
- Petrie, J.R.; Shrestha, P.; Belide, S.; Kennedy, Y.; Lester, G.; Liu, Q.; Divi, U.K.; Mulder, R.J.; Mansour, M.P.; Nichols, P.D.; Singh, S.P. Metabolic engineering Camelina sativa with fish oil like levels of DHA. PLoS One 2014, 9, e85061. [Google Scholar] [CrossRef]
- Petrie, J.R.; Singh, S.P. Expanding the docosahexaenoic acid food web for sustainable production: Engineering lower plant pathways into higher plants. AoB Plants 2011. [CrossRef]
- Qi, B.X.; Fraser, T.; Mugford, S.; Dobson, G.; Sayanova, O.; Butler, J.; Napier, J.A.; Stobart, A.K.; Lazarus, C.M. Production of very long chain polyunsaturated omega-3 and omega-6 fatty acids in plants. Nat. Biotechnol. 2004, 22, 739–745. [Google Scholar] [CrossRef]
- Venegas-Caleron, M.; Sayanova, O.; Napier, J.A. An alternative to fish oils: Metabolic engineering of oil-seed crops to produce omega-3 long chain polyunsaturated fatty acids. Prog. Lipid Res. 2010, 49, 108–119. [Google Scholar] [CrossRef]
- Ruiz-Lopez, N.; Haslam, R.P.; Napier, J.A.; Sayanova, O. Successful high-level accumulation of fish oil omega-3 long-chain polyunsaturated fatty acids in a transgenic oilseed crop. Plant J. 2014, 77, 198–208. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nichols, P.D.; Glencross, B.; Petrie, J.R.; Singh, S.P. Readily Available Sources of Long-Chain Omega-3 Oils: Is Farmed Australian Seafood a Better Source of the Good Oil than Wild-Caught Seafood? Nutrients 2014, 6, 1063-1079. https://doi.org/10.3390/nu6031063
Nichols PD, Glencross B, Petrie JR, Singh SP. Readily Available Sources of Long-Chain Omega-3 Oils: Is Farmed Australian Seafood a Better Source of the Good Oil than Wild-Caught Seafood? Nutrients. 2014; 6(3):1063-1079. https://doi.org/10.3390/nu6031063
Chicago/Turabian StyleNichols, Peter D., Brett Glencross, James R. Petrie, and Surinder P. Singh. 2014. "Readily Available Sources of Long-Chain Omega-3 Oils: Is Farmed Australian Seafood a Better Source of the Good Oil than Wild-Caught Seafood?" Nutrients 6, no. 3: 1063-1079. https://doi.org/10.3390/nu6031063
APA StyleNichols, P. D., Glencross, B., Petrie, J. R., & Singh, S. P. (2014). Readily Available Sources of Long-Chain Omega-3 Oils: Is Farmed Australian Seafood a Better Source of the Good Oil than Wild-Caught Seafood? Nutrients, 6(3), 1063-1079. https://doi.org/10.3390/nu6031063