Effect of Ethyl Pyruvate on Skeletal Muscle Metabolism in Rats Fed on a High Fat Diet


Abstract
:1. Introduction
2. Experimental Section
2.1. Animals and Diets
2.2. Enzymes Activities and Sulfhydryl Groups Oxidation
2.3. Plasma Biochemical Analyses
2.4. Chemicals
2.5. Statistical Analyses
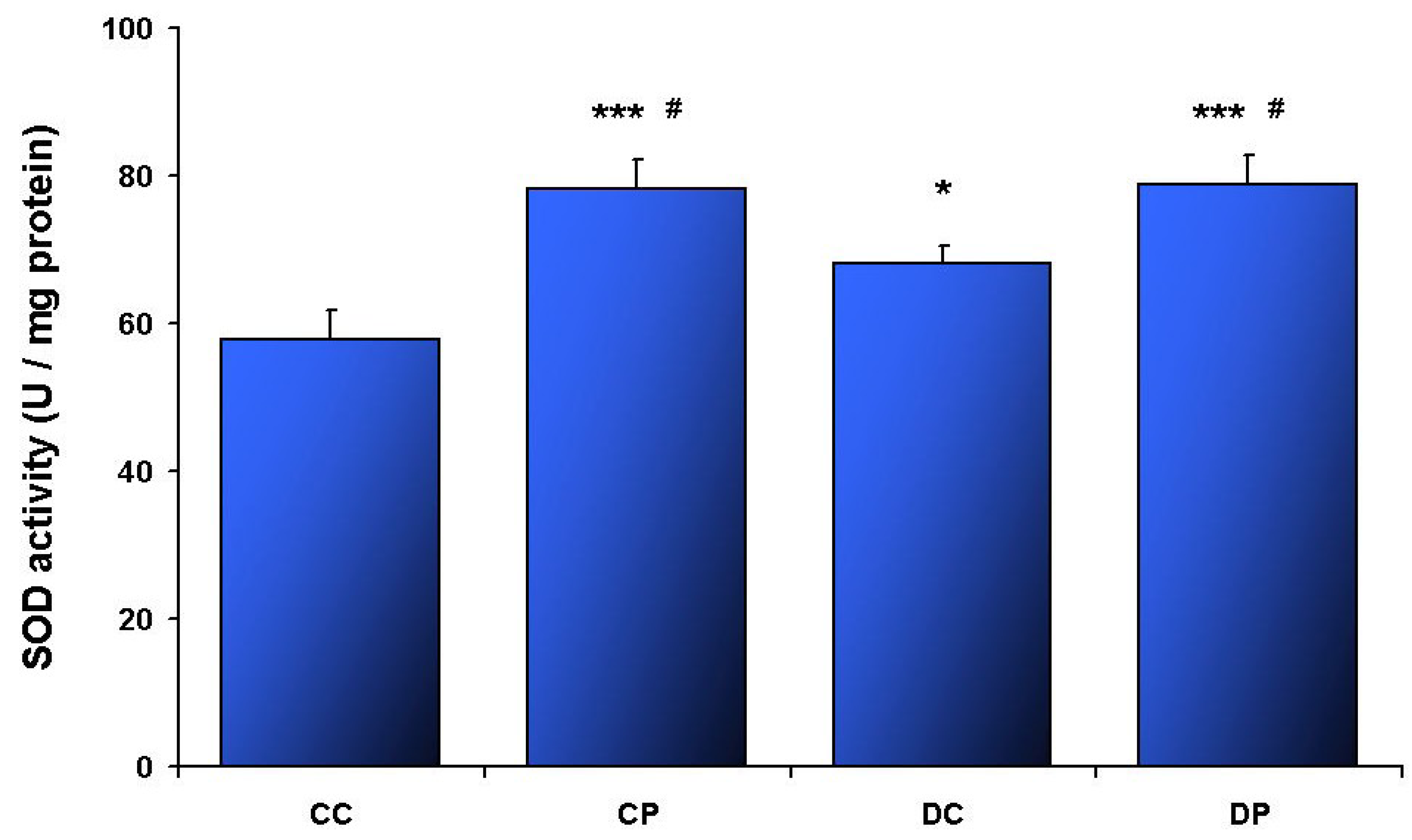
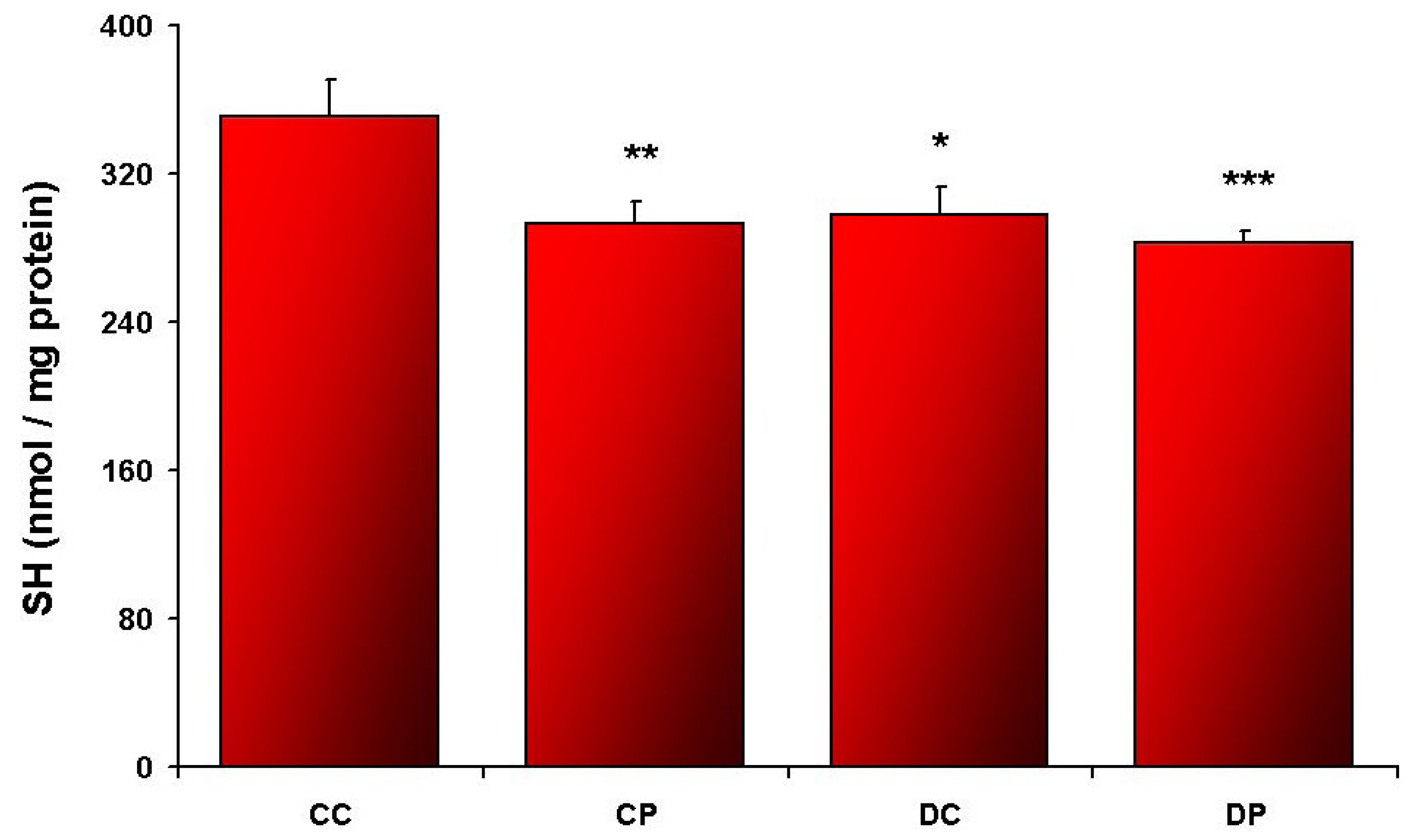
3. Results

{kind=link}
{kind=link}
{kind=link}
| Groups | ||||
|---|---|---|---|---|
| CC | CP | DC | DP | |
| SOL | ||||
| HADH (mU/mg protein) * | 133 ± 9 | 134 ± 6 | 180 ± 12 | 184 ± 5 |
| CS (mU/mg protein) ** | 151 ± 16 | 150 ± 4 | 179 ± 14 | 168 ± 8 |
| Fum (mU/mg protein) ** | 150 ± 12 | 136 ± 4 | 169 ± 11 | 155 ± 10 |
| EDL | ||||
| HADH (mU/mg protein) | 61 ± 5 | 62 ± 5 | 69 ± 9 | 63 ± 6 |
| CS (mU/mg protein) | 163 ± 14 | 151 ± 10 | 158 ± 18 | 140 ± 8 |
| Fum (mU/mg protein) | 124 ± 7 | 118 ± 8 | 119 ± 9 | 108 ± 8 |

| Groups | ||||
|---|---|---|---|---|
| CC | CP | DC | DP | |
| EDL | ||||
| CAT (U/mg protein) | 1.86 ± 0.07 | 1.54 ± 0.13 | 1.90 ± 0.20 | 1.87 ± 0.15 |
| SOD (U/mg protein) | 50.6 ± 2.0 | 51.0 ± 3.3 | 51.1 ± 7.6 | 45.1 ± 2.4 |
| SH (nmol/mg protein) | 272 ± 5 | 262 ± 7 | 275 ± 11 | 257 ± 2 |


| Groups | ||||
|---|---|---|---|---|
| CC | CP | DC | DP | |
| Insulin (ng/mL) | 0.49 ± 0.14 | 1.48 ± 0.28 * | 1.19 ± 0.27 ** | 1.52 ± 0.19 *** |
| Glucose (mg/dL) | 92.6 ± 1.8 | 94.5 ± 2.2 | 94.1 ± 2.8 | 96.3 ± 2.4 |
| HbA1c (%) | 4.83 ± 0.13 | 4.73 ± 0.10 | 4.70 ± 0.05 | 4.71 ± 0.05 |
4. Discussion
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Johannsen, D.L.; Ravussin, E. The role of mitochondria in health and disease. Curr. Opin. Pharmacol. 2009, 9, 780–786. [Google Scholar] [CrossRef]
- Parise, G.; de Lisio, M. Mitochondrial theory of aging in human age-related sarcopenia. Interdiscip. Top. Gerontol. 2010, 37, 142–156. [Google Scholar]
- Iossa, S.; Lionetti, L.; Mollica, M.P.; Crescenzo, R.; Botta, M.; Barletta, A.; Liverini, G. Effect of high-fat feeding on metabolic efficiency and mitochondrial oxidative capacity in adult rats. Br. J. Nutr. 2003, 90, 953–960. [Google Scholar] [CrossRef] [Green Version]
- Chanseaume, E.; Malpuech-Brugere, C.; Patrac, V.; Bielicki, G.; Rousset, P.; Couturier, K.; Salles, J.; Renou, J.P.; Boirie, Y.; Morio, B. Diets high in sugar, fat, and energy induce muscle type-specific adaptations in mitochondrial functions in rats. J. Nutr. 2006, 136, 2194–2200. [Google Scholar]
- Lionetti, L.; Mollica, M.P.; Crescenzo, R.; D’Andrea, E.; Ferraro, M.; Bianco, F.; Liverini, G.; Iossa, S. Skeletal muscle subsarcolemmal mitochondrial dysfunction in high-fat fed rats exhibiting impaired glucose homeostasis. Int. J. Obes. (Lond.) 2007, 31, 1596–1604. [Google Scholar] [CrossRef]
- Chanseaume, E.; Tardy, A.L.; Salles, J.; Giraudet, C.; Rousset, P.; Tissandier, A.; Boirie, Y.; Morio, B. Chronological approach of diet-induced alterations in muscle mitochondrial functions in rats. Obesity (Silver Spring) 2007, 15, 50–59. [Google Scholar]
- Takada, S.; Kinugawa, S.; Hirabayashi, K.; Suga, T.; Yokota, T.; Takahashi, M.; Fukushima, A.; Homma, T.; Ono, T.; Sobirin, M.A.; et al. Angiotensin II receptor blocker improves the lowered exercise capacity and impaired mitochondrial function of the skeletal muscle in type 2 diabetic mice. J. Appl. Physiol. 2013, 114, 844–857. [Google Scholar] [CrossRef]
- Yokota, T.; Kinugawa, S.; Hirabayashi, K.; Matsushima, S.; Inoue, N.; Ohta, Y.; Hamaguchi, S.; Sobirin, M.A.; Ono, T.; Suga, T.; et al. Oxidative stress in skeletal muscle impairs mitochondrial respiration and limits exercise capacity in type 2 diabetic mice. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1069–H1077. [Google Scholar] [CrossRef]
- Yuzefovych, L.V.; Musiyenko, S.I.; Wilson, G.L.; Rachek, L.I. Mitochondrial DNA damage and dysfunction, and oxidative stress are associated with endoplasmic reticulum stress, protein degradation and apoptosis in high fat diet-induced insulin resistance mice. PLoS One 2013, 8, e54059. [Google Scholar] [CrossRef]
- St Pierre, J.; Buckingham, J.A.; Roebuck, S.J.; Brand, M.D. Topology of superoxide production fromdifferent sites in the mitochondrial electron transport chain. J. Biol. Chem. 2002, 277, 44784–44790. [Google Scholar]
- Barazzoni, R.; Zanetti, M.; Cappellari, G.G.; Semolic, A.; Boschelle, M.; Codarin, E.; Pirulli, A.; Cattin, L.; Guarnieri, G. Fatty acids acutely enhance insulin-induced oxidative stress and cause insulin resistance by increasing mitochondrial reactive oxygen species (ROS) generation and nuclear factor-κB inhibitor (IκB)-nuclear factor-κB (NFκB) activation in rat muscle, in the absence of mitochondrial dysfunction. Diabetologia 2012, 55, 773–782. [Google Scholar] [CrossRef]
- Wells, G.D.; Noseworthy, M.D.; Hamilton, J.; Tarnopolski, M.; Tein, I. Skeletal muscle metabolic dysfunction in obesity and metabolic syndrome. Can. J. Neurol. Sci. 2008, 35, 31–40. [Google Scholar]
- Rogge, M.M. The role of impaired mitochondrial lipid oxidation in obesity. Biol. Res. Nurs. 2009, 10, 356–373. [Google Scholar] [CrossRef]
- Chanseaume, E.; Morio, B. Potential mechanisms of muscle mitochondrial dysfunction in aging and obesity and cellular consequences. Int. J. Mol. Sci. 2009, 10, 306–324. [Google Scholar] [CrossRef]
- Zorzano, A.; Hernandez-Alvarez, M.I.; Palacin, M.; Mingrone, G. Alterations in the mitochondrial regulatory pathways constituted by the nuclear co-factors PGC-1α or PGC-1β and mitofusin 2 in skeletal muscle in type 2 diabetes. Biochim. Biophys. Acta 2010, 1797, 1028–1033. [Google Scholar] [CrossRef]
- Wilson, L.; Yang, Q.; Szustakowski, J.D.; Gullicksen, P.S.; Halse, R. Pyruvate induces mitochondrial biogenesis by a PGC-1 α-independent mechanism. Am. J. Physiol. Cell Physiol. 2007, 292, C1599–C1605. [Google Scholar] [CrossRef]
- Owen, L.; Sunram-Lea, S.I. Metabolic agents that enhance ATP can improve cognitive functioning: A review of the evidence for glucose, oxygen, pyruvate, creatine, and l-carnitne. Nutrients 2011, 3, 735–755. [Google Scholar]
- Ivy, J.L.; Cortez, M.Y.; Chandler, R.M.; Byrne, H.K.; Miller, R.H. Effects of pyruvate on the metabolism and insulin resistance of obese Zucker rats. Am. J. Clin. Nutr. 1994, 59, 331–337. [Google Scholar]
- Das, U.N. Pyruvate is an endogenous anti-inflammatory and anti-oxidant molecule. Med. Sci. Monit. 2006, 12, RA79–RA84. [Google Scholar]
- Bunton, C.A. Oxidation of a-diketones and a-keto-acids by hydrogen peroxide. Nature 1949, 163, 444. [Google Scholar]
- Dobsak, P.; Courderot-Masuyer, C.; Zeller, M.; Vergely, C.; Laubriet, A.; Assem, M.; Eicher, J.C.; Teyssier, J.R.; Wolf, J.E.; Rochette, L. Antioxidative properties of pyruvate and protection of the ischemic rat heart during cardioplegia. J. Cardiovasc. Pharmacol. 1999, 34, 651–659. [Google Scholar] [CrossRef]
- Cruz, R.J., Jr.; Harada, T.; Sasatomi, E.; Fink, M.P. Effects of ethyl pyruvate and other alpha-keto carboxylic acid derivatives in a rat model of multivisceral ischemia and reperfusion. J. Surg. Res. 2011, 165, 151–157. [Google Scholar] [CrossRef]
- Olek, R.A.; Ziolkowski, W.; Kaczor, J.J.; Wierzba, T.H.; Antosiewicz, J. Higher hypochlorous acid scavenging activity of ethyl pyruvate compared to its sodium salt. Biosci. Biotechnol. Biochem. 2011, 75, 500–504. [Google Scholar] [CrossRef]
- Johansson, A.S.; Johansson-Haque, K.; Okret, S.; Palmblad, J. Ethyl pyruvate modulates acute inflammatory reactions in human endothelial cells in relation to the NF-kappaB pathway. Br. J. Pharmacol. 2008, 154, 1318–1326. [Google Scholar] [CrossRef]
- Kim, J.B.; Yu, Y.M.; Kim, S.W.; Lee, J.K. Anti-inflammatory mechanism is involved in ethyl pyruvate-mediated efficacious neuroprotection in the postischemic brain. Brain Res. 2005, 1060, 188–192. [Google Scholar]
- Zeng, J.; Liu, J.; Yang, G.Y.; Kelly, M.J.; James, T.L.; Litt, L. Exogenous ethyl pyruvate versus pyruvate during metabolic recovery after oxidative stress in neonatal rat cerebrocortical slices. Anesthesiology 2007, 107, 630–640. [Google Scholar]
- Fedeli, D.; Falcioni, G.; Olek, R.A.; Massi, M.; Cifani, C.; Polidori, C.; Gabbianelli, R. Protective effect of ethyl pyruvate on msP rat leukocytes damaged by alcohol intake. J. Appl. Toxicol. 2007, 27, 561–570. [Google Scholar] [CrossRef]
- Olek, R.A.; Ziolkowski, W.; Flis, D.J.; Fedeli, D.; Fiorini, D.; Wierzba, T.H.; Gabbianelli, R. The effect of ethyl pyruvate supplementation on rat fatty liver induced by high fat diet. J. Nutr. Sci. Vitaminol. (Tokyo) 2013, in press. [Google Scholar]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Hancock, C.R.; Han, D.H.; Chen, M.; Terada, S.; Yasuda, T.; Wright, D.C.; Holloszy, J.O. High-fat diets cause insulin resistance despite an increase in muscle mitochondria. Proc. Natl. Acad. Sci. USA 2008, 105, 7815–7820. [Google Scholar]
- Van den Broek, N.M.; Ciapaite, J.; de Feyter, H.M.; Houten, S.M.; Wanders, R.J.; Jeneson, J.A.; Nicolay, K.; Prompers, J.J. Increased mitochondrial content rescues in vivo muscle oxidative capacity in long-term high-fat-diet-fed rats. FASEB J. 2010, 24, 1354–1364. [Google Scholar] [CrossRef]
- Turner, N.; Bruce, C.R.; Beale, S.M.; Hoehn, K.L.; So, T.; Rolph, M.S.; Cooney, G.J. Excess lipid availability increases mitochondrial fatty acid oxidative capacity in muscle: Evidence against a role for reduced fatty acid oxidation in lipid-induced insulin resistance in rodents. Diabetes 2007, 56, 2085–2092. [Google Scholar] [CrossRef]
- Wright, L.E.; Brandon, A.E.; Hoy, A.J.; Forsberg, G.B.; Lelliott, C.J.; Reznick, J.; Lofgren, L.; Oscarsson, J.; Stromstedt, M.; Cooney, G.J.; et al. Amelioration of lipid-induced insulin resistance in rat skeletal muscle by overexpression of Pgc-1β involves reductions in long-chain acyl-CoA levels and oxidative stress. Diabetologia 2011, 54, 1417–1426. [Google Scholar] [CrossRef]
- Iossa, S.; Mollica, M.P.; Lionetti, L.; Crescenzo, R.; Botta, M.; Liverini, G. Skeletal muscle oxidative capacity in rats fed high-fat diet. Int. J. Obes. Relat. Metab. Disord. 2002, 26, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Zou, B.; Suwa, M.; Nakano, H.; Higaki, Y.; Ito, T.; Katsuta, S.; Kumagai, S. Adaptation of skeletal muscle characteristics to a high-fat diet in rats with different intra-abdominal-obesity susceptibilities. J. Nutr. Sci. Vitaminol. (Tokyo) 2003, 49, 241–246. [Google Scholar] [CrossRef]
- Ritchie, I.R.; Dyck, D.J. Rapid loss of adiponectin-stimulated fatty acid oxidation in skeletal muscle of rats fed a high fat diet is not due to altered muscle redox state. PLoS One 2012, 7, e52193. [Google Scholar] [CrossRef]
- Campbell, S.E.; Tandon, N.N.; Woldegiorgis, G.; Luiken, J.J.; Glatz, J.F.; Bonen, A. A novel function for fatty acid translocase (FAT)/CD36: Involvement in long chain fatty acid transfer into the mitochondria. J. Biol. Chem. 2004, 279, 36235–36241. [Google Scholar]
- Bonen, A.; Holloway, G.P.; Tandon, N.N.; Han, X.X.; McFarlan, J.; Glatz, J.F.; Luiken, J.J. Cardiac and skeletal muscle fatty acid transport and transporters and triacylglycerol and fatty acid oxidation in lean and Zucker diabetic fatty rats. Am. J. Physiol. 2009, 297, R1202–R1212. [Google Scholar]
- Seifert, E.L.; Estey, C.; Xuan, J.Y.; Harper, M.E. Electron transport chain-dependent and -independent mechanisms of mitochondrial H2O2 emission during long-chain fatty acid oxidation. J. Biol. Chem. 2010, 285, 5748–5758. [Google Scholar]
- Montgomery, C.M.; Webb, J.L. Metabolic studies on heart mitochondria. II. The inhibitory action of parapyruvate on the tricarboxylic acid cycle. J. Biol. Chem. 1956, 221, 359–368. [Google Scholar]
- Kao, K.K.; Fink, M.P. The biochemical basis for the anti-inflammatory and cytoprotective actions of ethyl pyruvate and related compounds. Biochem. Pharmacol. 2010, 80, 151–159. [Google Scholar] [CrossRef]
- Kim, S.Y.; Choi, J.S.; Park, C.; Jeong, J.W. Ethyl pyruvate stabilizes hypoxia-inducible factor 1 alpha via stimulation of the TCA cycle. Cancer Lett. 2010, 295, 236–241. [Google Scholar] [CrossRef]
- Chen, J.; Ma, H.T.; Wang, M.; Kong, Y.L.; Zou, S.X. Creatine pyruvate enhances lipolysis and protein synthesis in broiler chicken. J. Integr. Agric. 2011, 10, 1977–1985. [Google Scholar]
- Tsung, A.; Kaizu, T.; Nakao, A.; Shao, L.; Bucher, B.; Fink, M.P.; Murase, N.; Geller, D.A. Ethyl pyruvate ameliorates liver ischemia-reperfusion injury by decreasing hepatic necrosis and apoptosis. Transplantation 2005, 79, 196–204. [Google Scholar] [CrossRef]
- Mollen, K.P.; McCloskey, C.A.; Tanaka, H.; Prince, J.M.; Levy, R.M.; Zuckerbraun, B.S.; Billiar, T.R. Hypoxia activates c-Jun N-terminal kinase via Rac1-dependent reactive oxygen species production in hepatocytes. Shock 2007, 28, 270–277. [Google Scholar] [CrossRef]
- Hommelberg, P.P.; Langen, R.C.; Schols, A.M.; Mensink, R.P.; Plat, J. Inflammatory signaling in skeletal muscle insulin resistance: Green signal for nutritional intervention? Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 647–655. [Google Scholar] [CrossRef]
- Reade, M.C.; Fink, M.P. Bench-to-bedside review: Amelioration of acute renal impairment using ethyl pyruvate. Crit. Care 2005, 9, 556–560. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Olek, R.A.; Ziolkowski, W.; Wierzba, T.H.; Kaczor, J.J. Effect of Ethyl Pyruvate on Skeletal Muscle Metabolism in Rats Fed on a High Fat Diet. Nutrients 2013, 5, 2372-2383. https://doi.org/10.3390/nu5072372
Olek RA, Ziolkowski W, Wierzba TH, Kaczor JJ. Effect of Ethyl Pyruvate on Skeletal Muscle Metabolism in Rats Fed on a High Fat Diet. Nutrients. 2013; 5(7):2372-2383. https://doi.org/10.3390/nu5072372
Chicago/Turabian StyleOlek, Robert A., Wieslaw Ziolkowski, Tomasz H. Wierzba, and Jan J. Kaczor. 2013. "Effect of Ethyl Pyruvate on Skeletal Muscle Metabolism in Rats Fed on a High Fat Diet" Nutrients 5, no. 7: 2372-2383. https://doi.org/10.3390/nu5072372